Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds
- School of Molecular Biosciences, Washington State University, United States
- Center for Reproductive Biology, Washington State University, United States
Abstract
Scars are a serious health concern for burn victims and individuals with skin conditions associated with wound healing. Here, we identify regenerative factors in neonatal murine skin that transforms adult skin to regenerate instead of only repairing wounds with a scar, without perturbing development and homeostasis. Using scRNA-seq to probe unsorted cells from regenerating, scarring, homeostatic, and developing skin, we identified neonatal papillary fibroblasts that form a transient regenerative cell type that promotes healthy skin regeneration in young skin. These fibroblasts are defined by the expression of a canonical Wnt transcription factor Lef1 and using gain- and loss of function genetic mouse models, we demonstrate that Lef1 expression in fibroblasts primes the adult skin macroenvironment to enhance skin repair, including regeneration of hair follicles with arrector pili muscles in healed wounds. Finally, we share our genomic data in an interactive, searchable companion website (https://skinregeneration.org/). Together, these data and resources provide a platform to leverage the regenerative abilities of neonatal skin to develop clinically tractable solutions that promote the regeneration of adult tissue.
Introduction
Understanding how to induce skin regeneration instead of scarring will have broad implications clinically and cosmetically (Walmsley et al., 2015b). One of the main characteristics of scars is the absence of hair follicles, indicating that their regeneration in a wound may be a critical step in achieving scar-less skin repair (Yang and Cotsarelis, 2010). Interestingly, human embryonic skin has the capacity to regenerate without scars (Lo et al., 2012). Similarly, neonatal and adult mouse skin has the capacity to regenerate small non-functional hair follicles under specific conditions (Figure 1c–d; Ito et al., 2007; Rognoni et al., 2016; Telerman et al., 2017). These insights have prompted efforts in the field to define the molecular triggers that promote hair development in skin, with the ultimate goal of devising a way to regenerate fully functional hairs in adult skin wounds as a therapeutic modality (Yang and Cotsarelis, 2010).
Figure 1 with 2 supplements see all
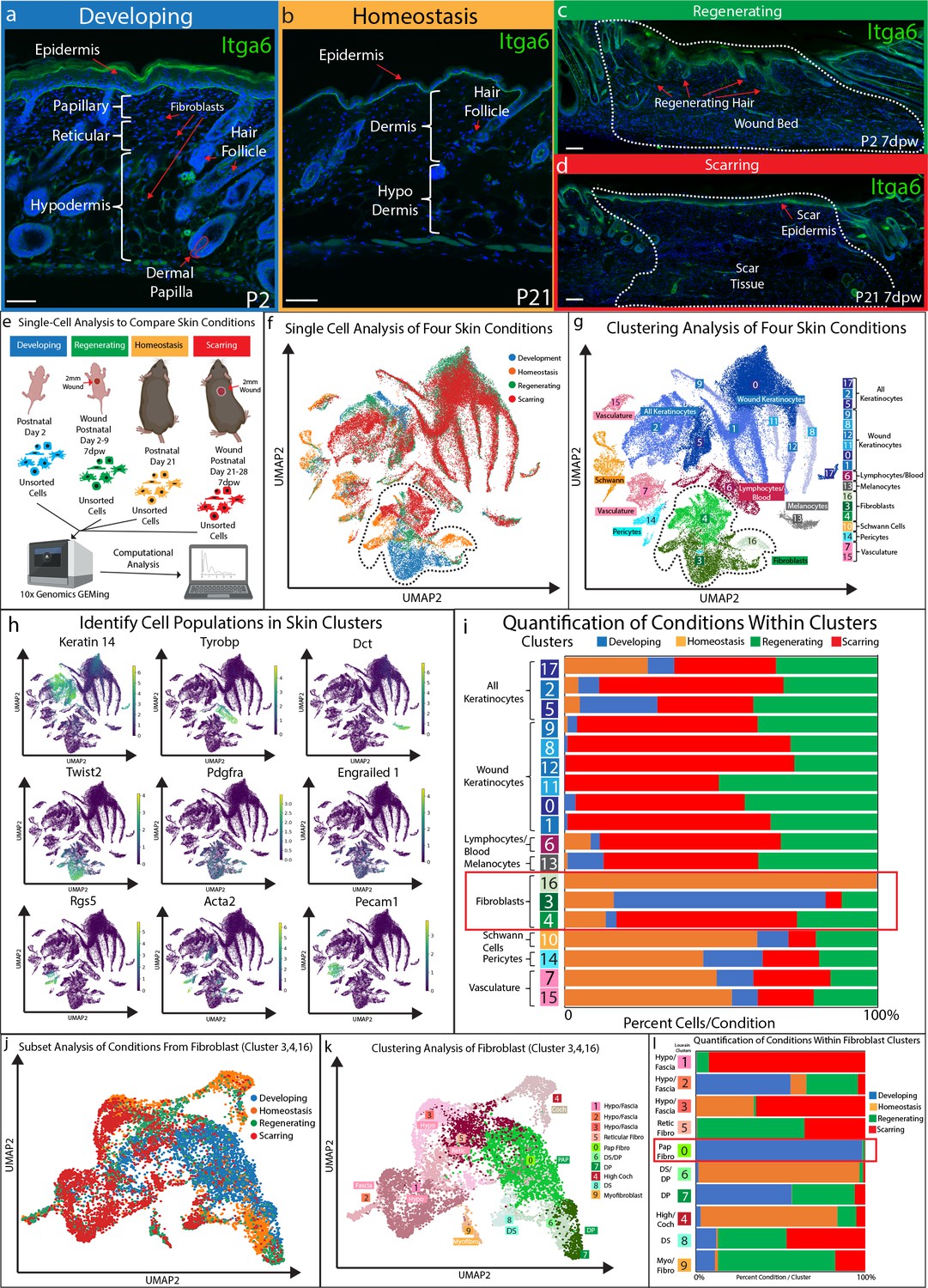
Fibroblasts in developing skin are distinct from regenerating, scarring, and homeostatic conditions.
(a–d) Immunostaining analysis and histological map of the dermis in developing Post-natal day 2 (P2) skin (a), homeostatic skin at P21 (b), skin undergoing regeneration P2 7 days post wound (7dwp) (c), and skin repairing a wound via scarring P21 7dpw (d). (e) Schematic representation of single-cell isolation, library preparation, and sequencing analysis for unsorted cells from developing (P2), regenerating (P2 7dpw), homeostatic (P21), and scarring (P21 7dpw) conditions. (f) UMAP visualization of all cell populations for all conditions. Each cell is color coded based according to condition as labeled. (g) Clustering analysis of the UMAP plot, color coded based on cell types. (h) Overlaying gene expression on UMAP clusters to identify cell types. (i) Quantification of the percentage of all cells represented within a cluster. (j) Subset and re-clustering of Clusters 3, 4, and 16 by computational integration. (k) Cluster analysis of integrated fibroblast clusters 3, 4, and 16. (l) Quantitation of fibroblasts within each cluster represented by condition. n = 3 biological replicates for each condition. Each biological replicate was used to generate a single library. Scale bars are 100 μm.
Human and mouse skin are similar in their overall structural complexity, indicating that mouse skin can be a useful model to study skin development and wound repair (Chen et al., 2013). Murine hair follicle and skin development primarily occurs between embryonic day 12.5 (E12.5) to post-natal day 21 (P21) (Müller-Röver et al., 2001). During this time, different fibroblast lineages are established that respond to the changes in the environment to support hair follicle and skin development (Driskell et al., 2013; Jiang et al., 2018; Rinkevich et al., 2015; Rognoni et al., 2016). Fibroblasts that support hair follicle development differentiate from the papillary fibroblast lineage, into dermal papilla (DP), dermal sheath (DS), and arrector pili (AP) cells (Driskell et al., 2013; Plikus et al., 2017). Reticular fibroblasts, which cannot differentiate into papillary fibroblast lineages, secrete extra-cellular-matrix (ECM) and form adipocytes (Driskell et al., 2013; Schmidt and Horsley, 2013). By post-natal day 2 (P2) fibroblast heterogeneity is fully established with the presence of the defined layers of the dermis (Figure 1a; Driskell et al., 2013). Skin maturation occurs after P2 with the formation of the AP and the completion of the first hair follicle cycle, which results in the loss of a defined papillary fibroblast layer (Figure 1b; Driskell et al., 2013; Rognoni et al., 2016; Salzer et al., 2018). We have previously shown that papillary fibroblasts are the primary source of de-novo dermal papilla during skin development, which are required for hair formation (Driskell et al., 2013). Furthermore, it has been suggested that adult murine skin form scars due to the lack of a defined papillary layer (Driskell et al., 2013; Driskell and Watt, 2015). Consequently, expanding this fibroblast layer in adult skin might support skin regeneration in adult mice.
The use of scRNA-seq in the murine skin has established useful cell atlases of the skin during development and homeostasis (Guerrero-Juarez et al., 2019; Haensel et al., 2020; Joost et al., 2020; Joost et al., 2018; Joost et al., 2016; Mok et al., 2019). In addition, scRNA-seq studies investigating wound healing have so far focused on comparing scarring, non-scarring, or regenerating conditions (Guerrero-Juarez et al., 2019; Haensel et al., 2020; Joost et al., 2018). These studies have helped to identified key markers of the newly discovered skin fascia, including Gpx3, Plac8, and Mest, which recently has been shown to contribute to scar formation (Correa-Gallegos et al., 2019; Grachtchouk et al., 2000; Joost et al., 2020). These, scRNA-seq studies have revealed that transgenic activation of the Shh pathway in the alpha-smooth-actin cells in scarring wounds, which includes pericytes, blood vessels, and myofibroblasts, can support small non-functional hair regeneration (Lim et al., 2018). However, activation of Shh pathway in dermal fibroblasts is associated with malignant phenotypes and may perturb development and homeostasis such that it may not be a safe target to support human skin regeneration clinically (Fan et al., 1997; Grachtchouk et al., 2000; Oro et al., 1997; Sun et al., 2020). Altogether, these findings suggest that an overall comparison of developing, homeostatic, scarring, and regenerating skin conditions will yield important discoveries for molecular factors that can safely support skin regeneration without harmful side effects.
The Wnt signaling pathway is involved in regulating development, wound healing, disease and cancer (Nusse and Clevers, 2017). Studies that activated Wnt and beta-catenin in skin have led to important discoveries for wound healing but have produced contrasting results in the context of fibroblast biology and hair follicle formation (Chen et al., 2012; Enshell-Seijffers et al., 2010; Hamburg-Shields et al., 2015; Lim et al., 2018; Rognoni et al., 2016). Wnts are a group of secreted protein that activates a cascade of events that stabilizes nuclear beta-catenin, which operates as a powerful co-transcription factor of gene expression. Importantly, beta-catenin alone cannot activate the expression of Wnt target genes without co-transcription factors. There are four Wnt co-transcription factors Tcf7 (Tcf1), Lef1, Tcf3 (Tcf7l1), and Tcf4 (Tcf7l2). These co-transcription factors modulate the functional outcome of Wnt signaling by binding to different target genes (Adam et al., 2018; Nguyen et al., 2009; Yu et al., 2012). In the context of wound healing and regeneration, it is not known how differential expression of Tcf/Lef can modulate fibroblast activity via Wnt signaling.
Since it has been shown that embryonic and neonatal skin have the potential to regenerate hair follicles upon wounding (Hu et al., 2018; Rognoni et al., 2016), we set out to identify the cell types and molecular factors that define this ability in order to transfer this regenerative potential to adult tissue. Our work has identified Lef1, as the factor in fibroblast of developing skin, that can transform adult tissue to regenerate.
Results
Developing fibroblasts are distinct cells that are associated with the ability to support hair follicle regeneration
We and others have previously shown that there are differences between neonatal and adult skin in terms of their cellular and biological properties (Ge et al., 2020; Rognoni et al., 2016; Salzer et al., 2018). These differences influence how skin can repair or if it can regenerate hair follicles in a wound. We showed that neonatal developing skin at P2 (Figure 1a) can regenerate small hair follicles 7 days after wounding (7dpw) (Figure 1c), while adult homeostatic skin at P21 (Figure 1b) heals by scarring 7dpw (Figure 1d). In order to identify the cell types and molecular pathways that define the ability of developing skin to regenerate, we performed a comparison of unsorted cells from four conditions using scRNA-seq (Figure 1e). A total of 66,407 cells from 12 libraries (n = 3 for each condition) met the preprocessing threshold, showed little batch variation (Figure 1—figure supplement 2), and were used for downstream analysis. Each condition was represented as different colors (Figure 1e–f). UMAP plotting of the 66,407 sequenced cells revealed distinct clusters (Figure 1f–g). To identify cell clusters, we used Louvain modulating analysis (Joost et al., 2016; Wolf et al., 2018), which resulted in 18 distinct clusters (Figure 1g). We assigned seven main classes of cells to the clusters. Keratinocytes represented the largest portion of cell types in the analysis (blue clusters - 0, 1, 2, 5, 8, 9, 11, 12, 17) (Figure 1h). We identified three distinct fibroblast clusters (green clusters - 3, 4, 16). These clusters expressed Twist2, Pdgfra, and En1 (Figure 1h). We also identified other clusters based on their distinct gene expression profile (Figure 1h; Supplementary file 1), such as immune cells (cluster 6), vasculature (clusters - 7, 15), Schwann cells (cluster 10), melanocytes (cluster 13), and a pericyte population (cluster 14).
To identify a unique cell type that supports the ability to regenerate hair follicles, we quantified the number of cells for each condition within a cluster and graphed them as a percentage of cell condition within a cluster (Figure 1i). Unexpectedly, there was no cluster exclusively defined by over-representation of the regenerating condition. Regenerating (green) and scarring (red) conditions were found in similar proportions amongst Keratinocytes, lymphocytes/blood, melanocytes, Schwann cells, blood vessels and fibroblasts (Figure 1i). However, there were two clusters that were distinct based on the percentage of cell types from a specific condition. Cluster 16 consisted of 99.9% of fibroblasts from homeostatic skin, while Cluster 3 consisted of 67.8% of developing fibroblasts (Figure 1i). In addition, cluster 4 consisted of 25.7% of regenerating condition, 57.6% of the scarring conditions. Since fibroblasts are known to play a critical role in regulating hair follicle reformation in wounds by differentiating into de-novo DP (Driskell et al., 2013; Plikus et al., 2017; Rognoni et al., 2016), we chose to further analyze clusters 3, 4, and 16.
We subset the fibroblast clusters 3, 4, 16 and re-clustered them by performing an integration analysis (Polański et al., 2020). This allowed us to test if different fibroblast subtypes, such as the papillary and reticular/hypodermal/fascia lineages, would be represented uniquely within a condition (Figure 1j–l). UMAP plotting of the integrated analysis revealed 10 distinct clusters (Figure 1k). The largest cluster (cluster 0) expressed markers of the papillary fibroblast lineage (Figure 1e–g). Four clusters (clusters 1, 2, 3, 4) distinctly expressed markers of the reticular/hypodermal/fascia layers of the dermis (Figure 1k; Figure 1—figure supplement 1a–b; Supplementary file 1; Correa-Gallegos et al., 2019; Joost et al., 2018). Three clusters (clusters 6, 7, 8) expressed markers of the dermal papilla and dermal sheath (Figure 1e–g). One cluster was distinctly represented by Acta2 indicating a myofibroblast subtype (cluster 9).
To identify conditions that are represented within specific fibroblast subtypes, we quantified the percentage of cells from each condition within the sub-clusters (Figure 1l). Our results revealed that four clusters were uniquely represented by four conditions. The myofibroblast sub-cluster (cluster 9) contained 69% of cells from the regenerating condition, while a hypodermal fascia cluster (cluster 1) contained 92.2% of cells from the scarring condition. Cluster 4, identified by the high expression of Cochlin (Coch), an inner ear ECM protein, contained 80.8% of cells from the homeostasis condition and a DP/DS sub-clusters (cluster 6) contained 95.0% of cells from the homeostasis condition. The papillary fibroblast cluster (cluster 0) contained 98.1% of cells from the developing skin condition. The DP sub-cluster (cluster 7) consisted of 56.5% cells from the developing condition and 37.1% cells from the regenerating condition. These results indicate that there is a distinct separation based on condition between fibroblast sub-types neonatal papillary fibroblasts versus reticular/hypodermal/fascia. Moreover, regenerating, and scarring conditions were present in similar proportions across multiple fibroblasts subtypes, besides clusters of hypodermal and fascia cells that were mainly composed of the scarring condition. This suggested that both wounding conditions shared similar cellular and molecular signatures that may be driven by the wound healing process.
Since our goal was to identify a cell type that uniquely represented the ability to support hair follicle formation that does not include a wound induced signature, we focused on cluster 0 which are developing fibroblasts. Developing fibroblasts migrate into wounds and support the reformation of hair follicles (Driskell et al., 2013; Rognoni et al., 2016). In homeostatic skin DP and DS are critical for hair follicle growth and cycling. However, they do not migrate into wounds to support hair follicle reformation (Johnston et al., 2013; Kaushal et al., 2015; Abbasi et al., 2020). Myofibroblasts are a major contributor to scar formation but may not normally differentiate into dermal papilla during hair follicle regeneration (Lim et al., 2018; Plikus et al., 2017). Furthermore, reticular and fascia fibroblasts contribute to scar formation but do not differentiate into dermal papilla (Correa-Gallegos et al., 2019; Driskell et al., 2013). We conclude that the developing fibroblasts are unique and might harbor the potential to support hair follicle regeneration in wounds.
Lef1 drives neonatal papillary lineage trajectory in developing fibroblasts
Neonatal developing skin has been extensively studied in the context of functional fibroblasts heterogeneity (Driskell et al., 2013). To identify fibroblasts lineages in scRNA-seq data, we isolated cell clusters from developing skin using pan fibroblast marker Pdgfra and Twist2 (Figure 2d, Figure 2—figure supplement 1; Collins et al., 2011) and then overlaid previously identified molecular markers of different lineages including papillary (Dpp4+/Dlk1-/Ly6a-), reticular (Dpp4-/Dlk1+/Ly6a+/-) and hypodermal fibroblasts (Dpp4-/Dlk1-/Ly6a+) on the developing scRNA-seq UMAP (Figure 2a–e). We also identified small populations of fascia utilizing the new marker Gpx3 (Figure 2d; Joost et al., 2020). Furthermore, the dermal sheath cluster was identified by expression of Acan and Acta2 (Heitman et al., 2020; Figure 2d, Figure 2—figure supplement 1), which is plotted between the dermal papilla (DP) and papillary fibroblasts clusters (Figure 2d–e). The DP clusters were defined by expression of Corin, a well-established marker for DP (Enshell-Seijffers et al., 2008). Interestingly, we also found novel heterogeneity within papillary fibroblast clusters. Besides the Dpp4+ clusters, Lef1 and Dkk1 expressions marked distinct papillary fibroblast clusters (Figure 2i, Figure 2—figure supplement 1). Overall, we concluded that the previously identified fibroblasts heterogeneity and the lineages could be identified in the scRNA-seq analysis of developing skin, with the additional subclusters of papillary fibroblasts (Figure 2e; Driskell et al., 2013).
Figure 2 with 2 supplements see all
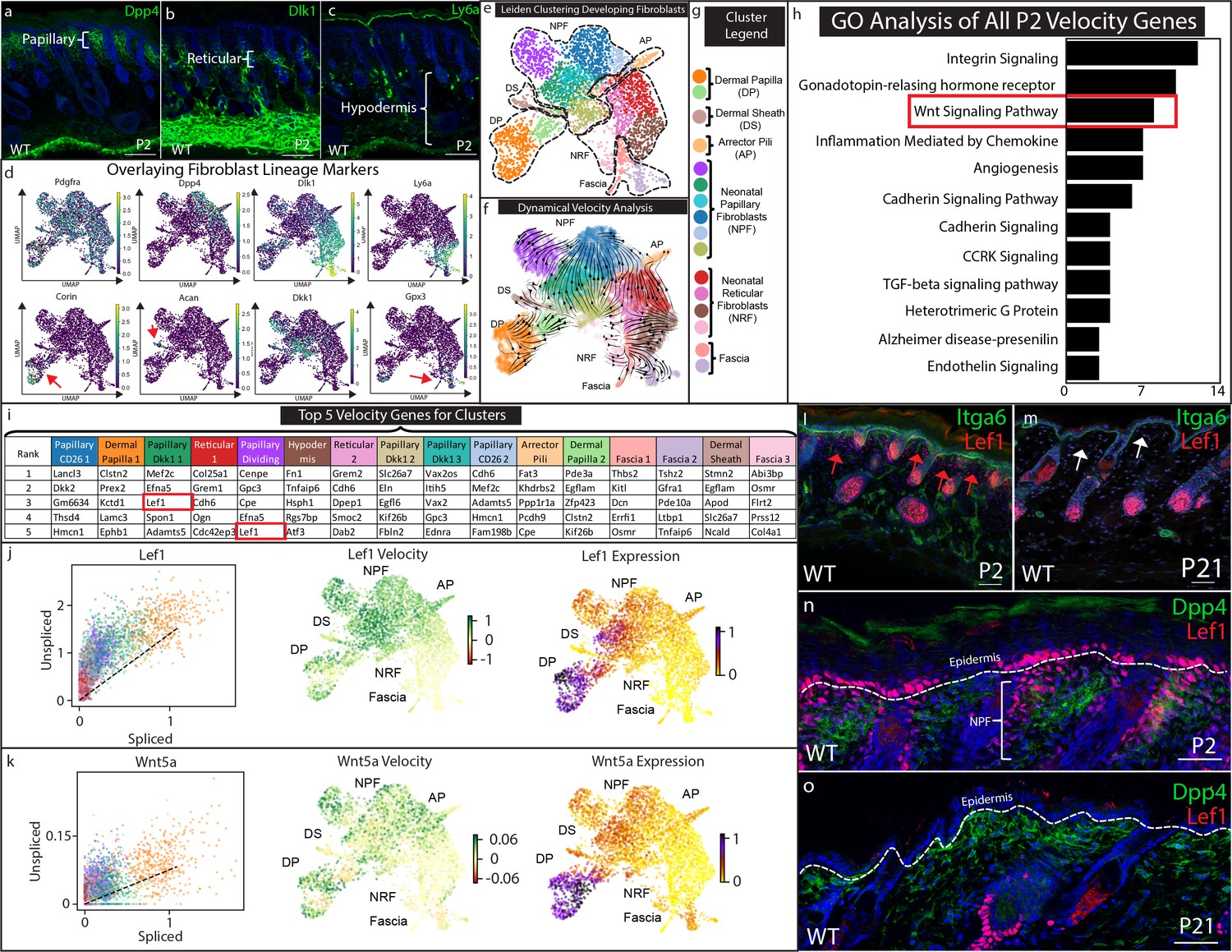
Papillary fibroblast lineage trajectory is defined by Lef1 in RNA Velocity analysis.
(a–c) Immunostaining development (P2) skin for Papillary (a) (Dpp4), Reticular/Hypodermal (b) (Dlk1/Pref1), and Hypodermal makers (c) (Ly6a). (d) Overlaying fibroblast lineage markers on UMAP projections of developing fibroblasts. (e) UMAP analysis of developing fibroblasts (f–g) RNA Velocity analysis overlaid on UMAP projection of developing fibroblasts. Clusters are colored coded and labeled. (h) GO Analysis of 195 driver genes of RNA Velocity for developing fibroblasts. (i) Top 5 Velocity driver genes of developing fibroblast clusters. (j) Phase contrast analysis of Lef1 and Wnt5a. (i–m) Immunostaining developing (P2) and homeostatic (P21) time points for Lef1 expression and counterstained with Itga6. Red arrows indicate areas of papillary dermis expressing Lef1. White arrows indicate papillary dermal region. (n–o) Immunostaining developing (P2) and homeostatic (P21) skin for Lef1 and Dpp4. Scale bars are 100 μm.
To estimate and predict the future state of developing fibroblasts and to identify the driver genes for fibroblast heterogeneity, we performed RNA velocity analysis (Figure 2f; Bergen et al., 2020). RNA velocity computationally compares the ratio of spliced and un-spliced mRNA from scRNA-seq data to estimate differentiation trajectories. The overall velocity analysis of developing fibroblasts is projected as arrows on the UMAP plot and it is defined by a list of putative driver genes (Supplementary file 2). Velocity analysis predicts that the dermal papilla is a differentiation endpoint, while papillary fibroblasts converge into the Dpp4 subcluster. Interestingly, the predicted differentiation trajectory of reticular fibroblasts suggests that they differentiate into arrector pili and also converge into the Dpp4 papillary subcluster. This contradicts previous in vivo lineage-tracing studies of the Dlk1+ fibroblast populations but agrees with ex vivo studies that tested the differentiation potential of Dpp4-/Dlk1+/Ly6a- fibroblast populations in chamber grafting and hydrogel assays (Driskell et al., 2013). Hypodermal and fascia fibroblast clusters were predicted to maintain the lower lineage (Figure 2f).
RNA velocity analysis predicts cellular trajectories using a list of the same driver genes that are ranked differently in each cluster (Figure 2i, Supplementary file 2). These genes are ranked based on their roles in driving the velocity trajectories. We performed PANTHER pathway classification on this list genes and found that the most highly represented pathways included Integrin signaling, Wnt signaling, Cadherin signaling and TGFb signaling pathways (Figure 2h; Supplementary file 5). We next ranked the top five genes that drive the velocity of each cluster (Figure 2i). We found that Dkk2, Mef2c, Lef1, and Egfl6 were identified to drive the velocity for different papillary clusters (Figure 2i). Col25a1, Smoc2, Fn1, Dcn, and Col4a1 were found to drive velocity to reticular and fascia clusters. Interestingly, Lef1 was found in the Papillary Dkk1 fibroblasts and the Dividing Papillary subclusters (Figure 2i). This suggests that the expression of Lef1 may drive RNA velocity toward papillary fibroblast and dermal papilla. We further analyzed Wnt signaling genes found in the papillary and dermal papilla clusters to determine the direction of the trajectory by phase contrast analysis (Figure 2j) Phase contrast analysis predicts that Lef1 and Wnt5a velocity increases towards to the papillary fibroblast and dermal papilla clusters (Figure 2j–k).
To validate the expression and localization of Lef1 in developing skin fibroblasts, we visualized the in vivo expression pattern via immunostaining (Figure 2l–o). Our results confirmed the population of neonatal papillary fibroblasts that is found only in developing skin (Figure 2j, red arrows) and not in juvenile homeostatic skin (Figure 2k, white arrows). Co-staining with Dpp4 confirmed that Lef1 marked a subset of neonatal papillary fibroblasts (Figure 2l) that was also not found in juvenile skin (Figure 2m).
The transient expression of Lef1 in neonatal papillary fibroblast correlates with the ability of neonatal developing skin to regenerate hair follicles in wounds (Rognoni et al., 2016). In addition, velocity analysis predicts that Lef1 drives developing fibroblast velocity to papillary fibroblast and DP. Importantly, subset analysis of regenerating and scarring fibroblasts did not identify fibroblasts defined by Lef1 expression, indicating that this transient population is unique to developing skin (Figure 2—figure supplement 2a–f; Supplementary file 4). We hypothesize that Lef1 expression in fibroblasts marks the ability of developing skin to regenerate hair follicles during the wound healing process.
Neonatal skin regeneration requires Lef1 expression in fibroblasts
Based on our scRNA-seq findings, we postulated that Lef1 defines a neonatal papillary fibroblast that supports skin regeneration during neonatal wound healing. To test if Lef1 in fibroblasts is required to support neonatal regeneration, we produced a tissue-specific knockout model. We utilized the dermal specific promoter Twist2 to drive Cre expression, bred with a mouse line with flox sites flanking Exon 7 and 8 of the Lef1 locus (Yu et al., 2012). We called this mouse line Twist-Lef1cKO (Figure 3a). Twist-Lef1cKO mice were viable and fertile with small shifts in hair follicle phenotypes that resulted in less dense fur (Fine et al., 2020). We confirmed dermal Lef1 ablation from the papillary fibroblast at P2 and from adult DP by immunostaining (Figure 3b–e). We also performed 2 mm wounds on P2 Twist-Lef1cKO and wild-type littermates harvesting at 7dpw (Figure 3f). Our analyses revealed that Lef1 was expressed in the de-novo DP and regenerating hair follicle buds of wild-type mice (Figure 3g,i), but that wounded Twist-Lef1cKO mice lacked regeneration (Figure 3h,j,k). We conclude that Lef1 expression in neonatal fibroblasts is required for neonatal hair follicle regeneration in wounds.
Figure 3
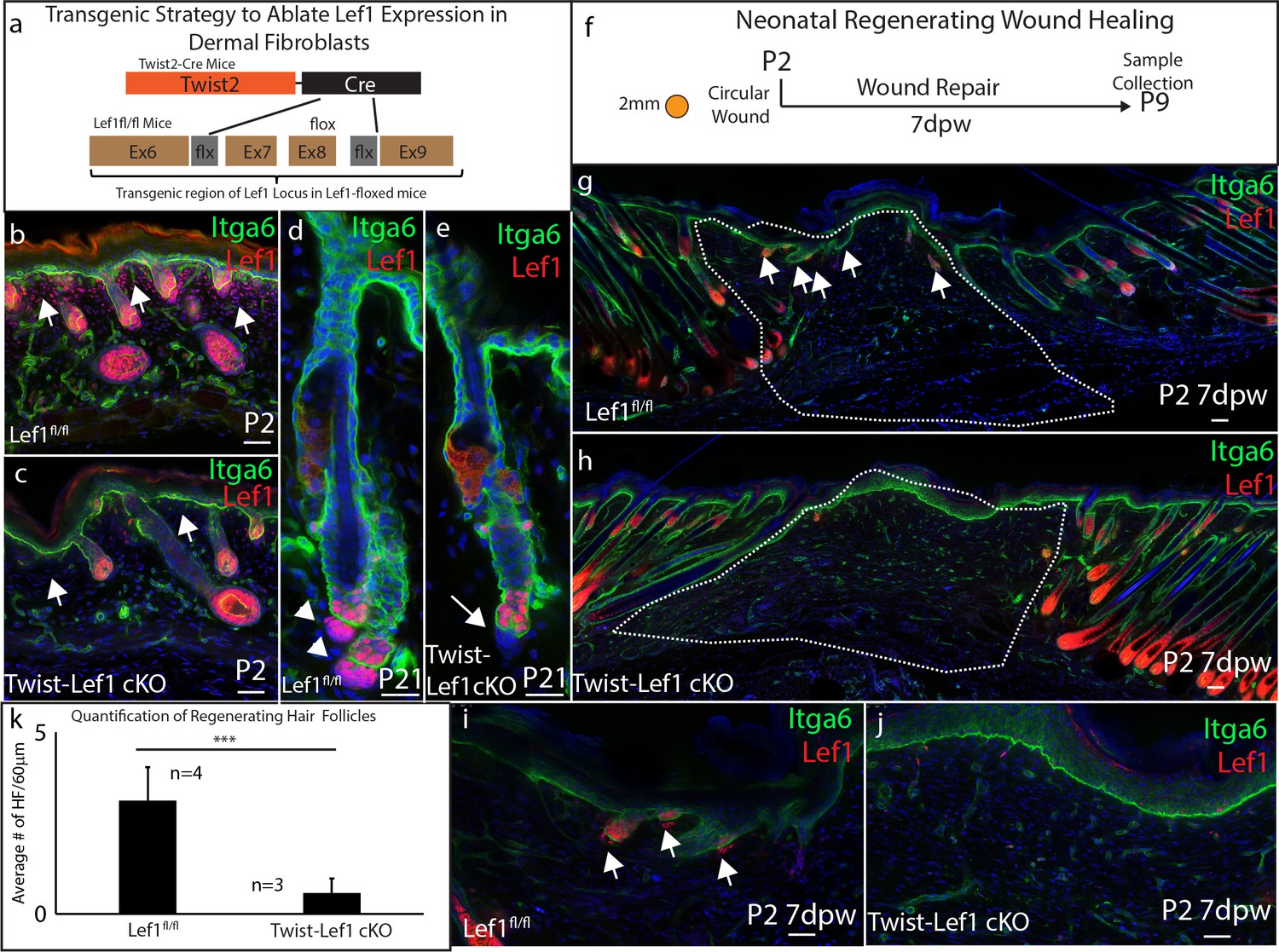
Dermal Lef1 is required to support hair follicle regeneration in neonatal wounds.
(a) Schematic representation of transgenic strategy to ablate Lef1 expression in dermal fibroblasts. Scale bars are 50 μm (b–e) Verifying dermal Lef1 ablation by immunostaining. White arrows indicate papillary regions (b–c) or dermal papilla (d–e). Scale bars are 50 μm (f) Schematic describing 2 mm circular wounds and harvest times to test if dermal Lef1 is required for regeneration. (g–i) Immunostaining of P2 7 days post wound (7dpw) skin for Itga6 and Lef1. (g-h Scale bars are 100 μm) Wound beds are highlighted by white outline. White arrows indicate regenerating hair follicles. (i-j Scale bars are 50 μm) (k) Quantification of regenerating hair follicles in wounds beds of n = 4 Lef1fl/fl (WT) and n = 3 Twist-Lef1cKO mice. p<0.003.
Dermal Lef1 expression primes adult skin to enhance regeneration in large wounds
Our scRNA-seq data, together with tissue-specific knockout, implies that Lef1 expression in the dermis could convey the regenerative ability of neonatal papillary fibroblasts in adult skin. We hypothesize that constitutively active Lef1 expression in dermal fibroblasts will prime skin to support regeneration. To induce Lef1 expression in the dermis of the skin, we utilized a previously published transgenic mouse model Lef1KI (Lynch et al., 2016) bred with Twist2-Cre mouse line called the Twist-Lef1KI line (Figure 4a). We verified that Lef1 was expressed in all dermal fibroblast compartments in Twist-Lef1KI mice during development and in adult homeostatic skin conditions (Figure 4b–e). Importantly, the only phenotype detected was the early entry of the hair follicle cycling into anagen at P21 in Twist-Lef1KI mice (Figure 2d–e). In addition, Twist-Lef1KI mice have similar life spans as wild type mice without any adverse effects (Figure 4—figure supplement 1). Our results reveal that induced expression of Lef1 in fibroblasts does not negatively affect the development and homeostasis of murine skin.
Figure 4 with 1 supplement see all
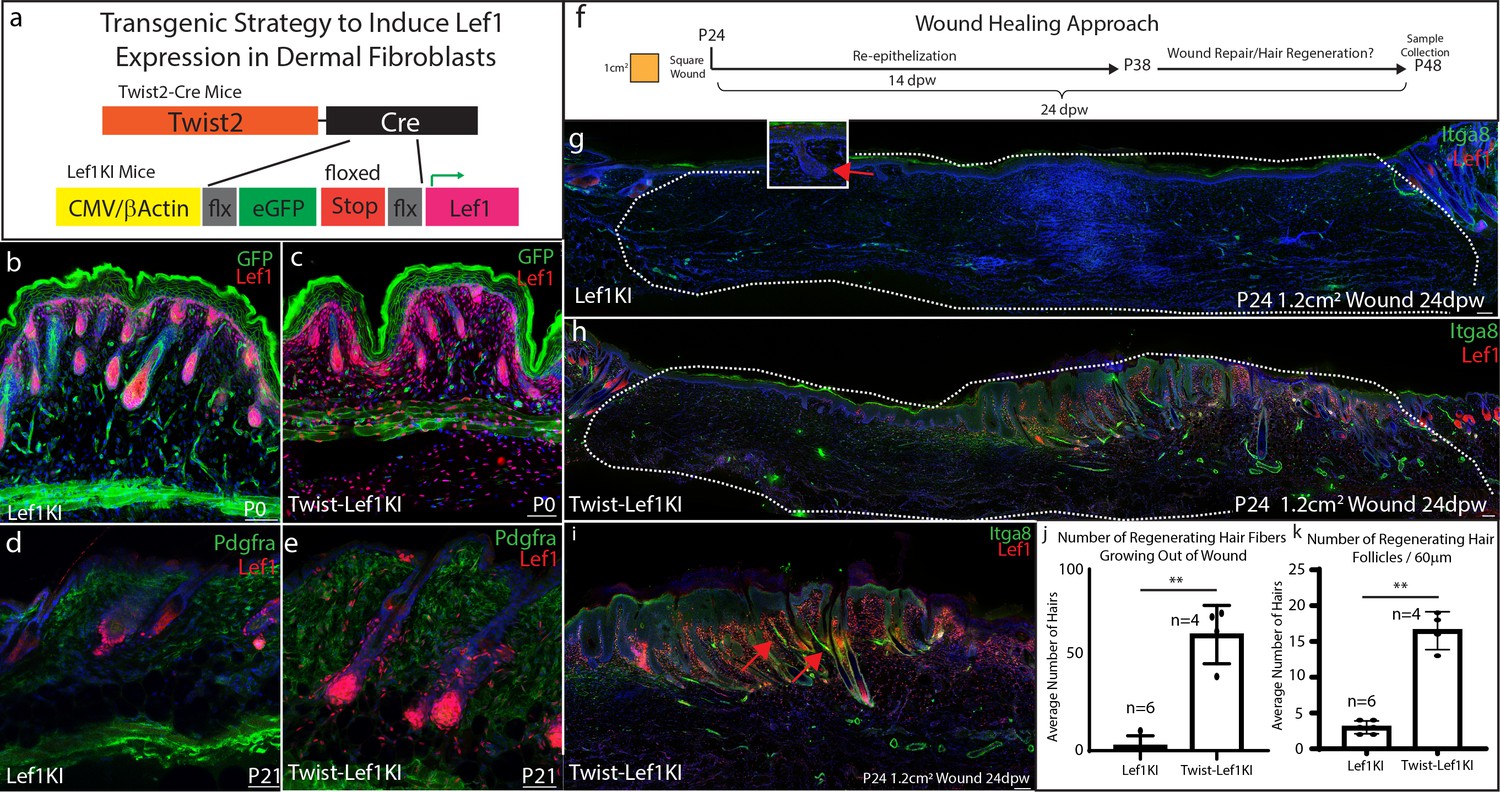
Lef1 expression in the dermis enhances skin regeneration in large wounds.
(a) Schematic representation of transgenic strategy to ectopically express Lef1 in fibroblasts during development, homeostasis, and wound healing. (b–c) Immunostaining for Lef1 and GFP in P0 Lef1KI and Twist-Lef1KI (Twist-Cre+/Lef1KI+) mice. (d–e) Immunostaining for Lef1 and PDGFRa in P21 Lef1KI and Twist-Lef1KI mice. (f) Schematic representation of wound healing approach. (g–k) Immunostaining tissue for Lef1 and Itga8 (arrector pili) from the wounds of Lef1KI and Twist-Lef1KI mice. Regenerating hair follicles in Lef1KI mice can be found by red arrows. Wound beds are marked by white dotted outlines. High resolution area of regenerating hair follicles in Twist-Lef1KI mice (i). (j) Quantification of regenerating hair follicles growing out of the wound beds of Lef1KI n = 6 and Twist-Lef1KI n = 4 mice p<0.004. (k) Quantification of regenerating hair follicles detected in 60 μm sections of wound beds. p<0.001. Scale bars at b-d are 100 μm. Scale bars at g-i are 200 μm.
Adult skin normally does not regenerate but has the capacity to reform non-functional hair follicles (small and lack arrector pili) in wounds 1 cm2 or larger (Ito et al., 2007). To test if overexpression of Lef1 in adult dermal fibroblasts enhances hair regeneration in adult skin, we wounded wild type (Lef1KI) and Twist-Lef1KI mice at P24 with 1.2 cm2 wounds and harvested at 24dpw (Figure 4f). Our analysis comparing Lef1KI and Twist-Lef1KI wound beds revealed enhanced hair regeneration (Figure 4f–k). There were five times more regenerating hair follicles on average in Twist-Lef1KI wound beds compared to wild type wounds (Figure 4j–k). The non-functional hair follicles that typically regenerate in large adult wounds are small and lack the arrector pili, a smooth muscle that makes hair stand up when contracted. Strikingly, the hair follicles regenerating in the Twist-Lef1KI wound beds contained arrector pili (Figure 4i) and were larger than hair follicles found in wild-type wounds. We conclude that Lef1 expression in the dermis of the skin leads to enhances regeneration.
Dermal Lef1 transforms scarring wounds to be regenerative
Wound healing studies utilize a standardized approach to account for the hair follicle cycle, which influences the wound healing process (Ansell et al., 2011). Consequently, the bulk of wound healing and regeneration studies are performed at 3 weeks of age (~P21) or at 7–8 weeks of age (~P57) (Gay et al., 2013; Guerrero-Juarez et al., 2019; Ito et al., 2007; Lim et al., 2018; Plikus et al., 2017; Plikus et al., 2008). In addition, a standard size wound of roughly 8 mm in diameter or smaller is used, which heal by scarring (Driskell et al., 2013; Lim et al., 2018; Rognoni et al., 2016). Since Lef1KI mice showed enhanced regenerative capacity in large wounds, we explored the regeneration potential of Twist-Lef1KI skin in wounds that normally scar. To determine if the hair follicle cycle influences skin regeneration we performed wounds at different time points: P24 which undergoes a hair follicle cycle during wound healing; P40 which heals during the resting phase of the hair cycle; and P90 which is the beginning of the un-synchronized hair follicle cycling that occurs in adult mice (Figure 5a). Scars formed in wounds performed at P24 in both WT and Twist-Lef1KI mice (Figure 5b–c,l). However, hair follicle regeneration with arrector pili occurred in wounds performed at P40 and P90 in Twist-Lef1KI mice (Figure 5d–k,l; Figure 5—figure supplement 1; Figure 5—figure supplement 2). Occasionally, we observed cyst-like structures in regenerated wounds, similar to that observed in WIHN (Ito et al., 2007). We conclude that Lef1 expression in the dermis induces skin to support hair follicle regeneration in wounds with variation depends on the timepoints and hair follicle cycle.
Figure 5 with 2 supplements see all
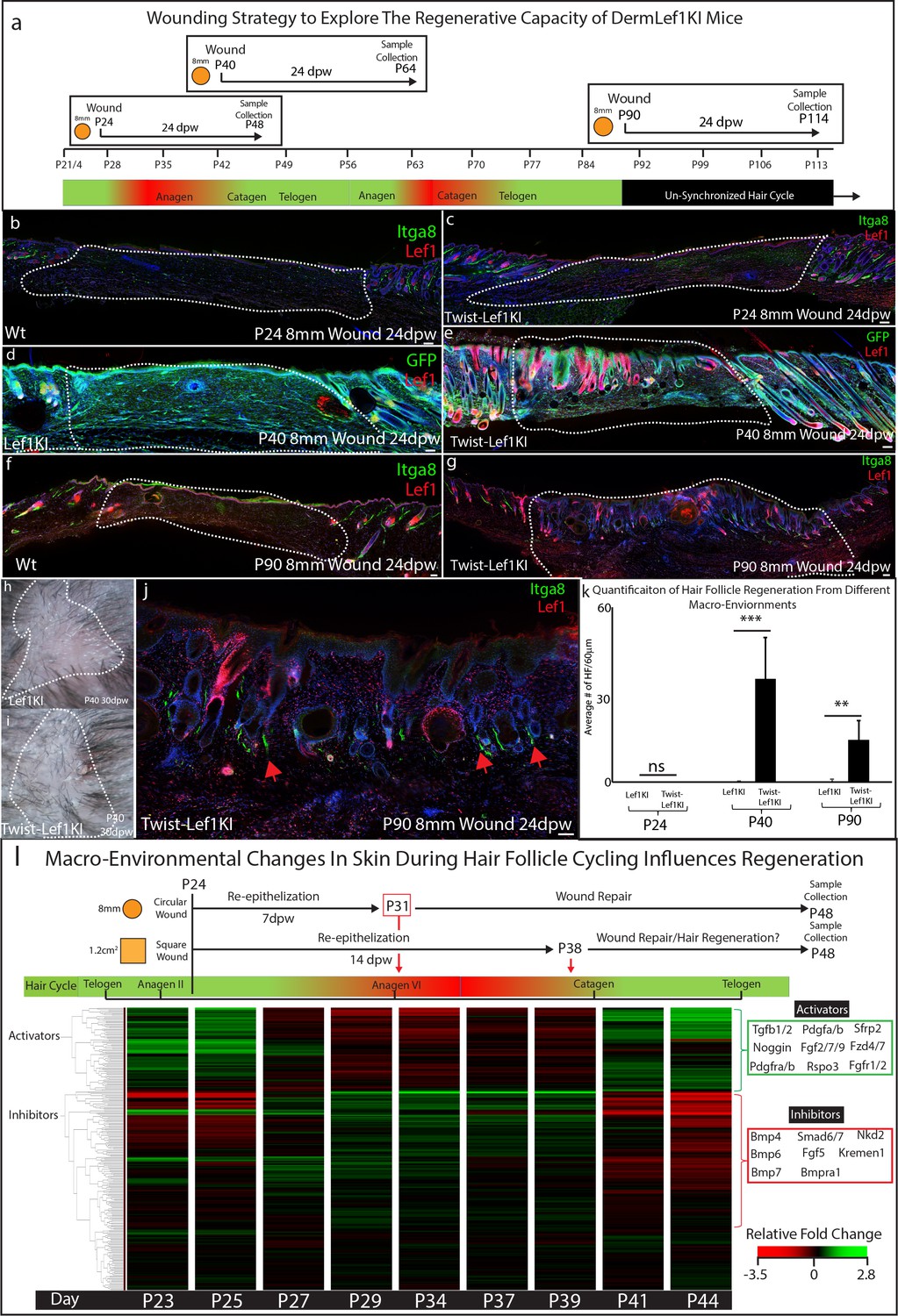
Lef1 expression in the dermis enhances skin regeneration in permissive macro environments.
(a) Schematic of the differential wound healing analysis from different time points of the hair follicle cycle in murine skin. (b–g,j) Immunostaining analysis of wounds 24dpw from P24 (b–d), P40 (d–e), and P90 (f–g,j). Sections were stained for either Itga8 and Lef1 (b–c,f–g,j) or GFP and Lef1 (d–e). Itga8 labels arrector pili. (h–i) Macroscopic analysis of wound beds from WT (Lef1KI) and Twist-Lef1KI mice. Hair follicles could be seen growing out of the wound bed of Twist-Lef1KI mice. (k) Quantification of the average number of regenerating hair follicles detected in wounds beds of 60 μm sections from P24 24dpw (ns, WT-n = 4, Twist-Lef1KI -n = 4), P40 24dpw (p<0.0009 WT-n = 4, Twist-Lef1KI -n = 4), and P90 24dpw (p<0.007 WT-n = 3, Twist-Lef1KI -n = 4). (l) Schematic representation of different wound healing approaches above a heatmap of microarray data (Lin et al., 2009) from key time points during hair follicle cycling. Heatmap is generated from a list of Activator and Inhibitor genes reported to regulate the progression of the hair follicle cycle (Wang et al., 2017). Scale bars are 100 μm.
Our wound healing studies mirror previous work suggesting that the size of a wound can dictate the potential of skin to regenerate instead of scar (Ito et al., 2007). To explain ‘why size matters’ we investigated and mapped the timing of wound closure and regeneration in the context of molecular signals within the dermal macro-environment that are dynamically changing throughout the hair follicle cycle (Plikus et al., 2008; Wang et al., 2017). Previous studies have defined the activators and inhibitors that regulate hair follicle cycling during this time (Wang et al., 2017; Supplementary file 3). Activators include the Wnt signaling pathway, Noggin, Tgfb, and fibroblast markers such as Pdgfra. Inhibitors include the BMP signaling pathway. These activators and inhibitors have been previously shown to modulate Lef1 activity (Jamora et al., 2003). To explain why Twist-Lef1KI mice scar at P24, we generated a diagram of wound re-epithelization and healing time points comparing 8 mm circular wounds to 1.2 cm2 wound in the context of gene expression for inhibitors and activators at all time points during the hair follicle cycle (Figure 5l). 8 mm circular wounds re-epithelialize 7 dpw (Driskell et al., 2013) in a macro-environment with high levels of Lef1 inhibitors, which are not inductive to hair follicle formation (Figure 5l). In contrast, large 1.2 cm2 wounds re-epithelialize during the time point of the hair cycle that has high activators and low inhibitors (Figure 5l). We conclude that the size of a wound dictates the time point when wound closure occurs which is associated with inhibitor or inductive regenerative signals.
Discussion
It is well established that embryonic and neonatal skin has the potential to heal in a scar-less manner (Walmsley et al., 2015a). However, the cell types and signals that can transfer this ability to adult skin have not been identified. In this study, we identify developing papillary fibroblasts as a transient cell population that is defined by Lef1 expression. Inducing Lef1 expression in the adult dermis will prime skin to support enhanced hair follicle regeneration. Interestingly, ectopic Lef1 expression in the dermis did not result in any adverse phenotypes during development (Supplementary file 4).
scRNA-seq has provided an unprecedented insight in the cell types and molecular pathways that are represented within regenerating and scarring wounds (Guerrero-Juarez et al., 2019; Haensel et al., 2020; Joost et al., 2018; Abbasi et al., 2020). For example, comparisons of these different conditions have identified the activation of the Shh pathway as a key pathway to support hair follicle regeneration in wounds of different sizes (Lim et al., 2018). However, the activation of ectopic Shh in either the epidermis or dermis is associated with unwanted phenotypes and cancer (Fan et al., 1997; Grachtchouk et al., 2000; Oro et al., 1997; Sun et al., 2020). Unexpectedly, we discovered a neonatal papillary fibroblast population that is defined by Lef1 expression in developing skin instead of in regenerating conditions. Importantly, our comparative analysis of regenerating and scarring wounds reveals their similarities (Figure 1i, Figure 2—figure supplement 2) more than their differences suggesting that the wound environment is a powerful driving force for gene expression even with a heterogeneous population of cells. Consequently, the basic mechanisms occurring during development and maintenance of neonatal tissue may hold the keys to transforming adult tissue to regenerate instead of scarring.
Studies involving the activation of the Wnt and beta-catenin pathways in skin have led to important discoveries in wound healing research, but have produced contrasting results in the context of fibroblast biology and hair follicle formation (Chen et al., 2012; Hamburg-Shields et al., 2015; Ito et al., 2007; Lim et al., 2018; Mastrogiannaki et al., 2016; Rognoni et al., 2016). The activation of beta-catenin in fibroblasts during wound healing has been shown to inhibit hair follicle reformation in scars (Lim et al., 2018; Rognoni et al., 2016) while deactivating Wnt signaling inhibited hair reformation (Myung et al., 2013). Moreover, persistent Wnt activity in the dermis has been recently shown to drive WIHN toward fibrotic wound healing (Gay et al., 2020). In contrast, the activation of beta-catenin during development increases hair formation and size (Chen et al., 2012; Enshell-Seijffers et al., 2010). One explanation for these conflicting results is the presence of different fibroblast lineages present in either development or wounds, which express different Lef/Tcf factors to direct a Wnt/beta-catenin-specific response (Driskell et al., 2013; Philippeos et al., 2018; Rognoni et al., 2016). We and others have shown that Lef1 is expressed in embryonic and neonatal papillary fibroblasts, which is lost in adult fibroblasts (Driskell et al., 2013; Mastrogiannaki et al., 2016; Philippeos et al., 2018; Rognoni et al., 2016). Here, we demonstrate that Lef1 is a key transcription factor in developing fibroblasts that supports hair follicle formation in adult wounds. Our data indicates that Lef1 expression in the dermis of the skin primes a response to support hair follicle neogenesis and arrector pili reformation in the wound. Our results align with studies that induced Lef1 expression in cultured human dermal papilla cells to enhance hair follicle formation in vitro (Abaci et al., 2018). In humans, at the age of 50+, the quality of the papillary dermis gradually deteriorates, which correlates with the aging skin decreased wound healing abilities (Haydont et al., 2019; Marcos-Garcés et al., 2014). Our data suggests that activating the papillary region to retain its identity by expression of Lef1 has the potential to enhance wound healing in humans.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (M. musculus) | Dermo1-Cre or Twist2-Cre, C57Bl6/BalbC | Jackson Laboratory | RRID:MGI:3840442 | |
| Strain, strain background (M. musculus) | Lef1KI, C57Bl6/BalbC | Engelhardt lab - University of Iowa | Other | A gift from the Engelhardt lab |
| Strain, strain background (M. musculus) | Lef1KO, C57Bl6/BalbC | Xue lab - University of Iowa | RRID:MGI:5447957 | A gift from the Xue lab |
| Sequencing kit | Single Cell 3’ Library and Gel Bead Kit v2 | 10X Genomics | Cat#: PN-120267 | |
| Antibody | Anti-Integrin alpha 8 (Itga8) (goat polyclonal) | R and D Systems | Cat#: AF4076 | IF(1:200) |
| Antibody | Anti-Pref1/Dlk1 (rabbit monoclonal) | R and D Systems | Cat#: AF8277 | IF(1:200) |
| Antibody | Anti-Dpp4/CD26 (goat polyclonal) | R and D Systems | Cat#: AF954 | IF(1:200) |
| Antibody | Anti-Pdgfra (goat polyclonal) | R and D Systems | Cat#: AF1062 | IF(1:200) |
| Antibody | Anti-Lef1 (rabbit monoclonal) | Cell Signaling | Cat#: 2230S | IF(1:200) |
| Antibody | Anti-Alpha smooth muscle actin (Acta2) (rabbit polyclonal) | abcam | Cat#: ab5694 | IF(1:200) |
| Antibody | Anti GFP (chicken polyclonal) | abcam | Cat#: ab13970 | IF(1:200) |
| Antibody | Anti-Ly6a/Sca1 (rat monoclonal) | Biolegend | Cat#: 108108 | IF(1:200) |
| Antibody | Anti-CD49f/Integrin alpha 6 (Itga6) (rat monoclonal) | BD Biosciences | Cat#: 555735 | IF(1:200) |
| Antibody | Anti-goat Alexa Fluor 488 (Donkey polyclonal) | Invitrogen | Cat#: A-11055 | IF(1:500) |
| Antibody | Anti-goat Alexa Fluor 555 (Donkey polyclonal) | Invitrogen | Cat#: A-21432 | IF(1:500) |
| Antibody | Anti-goat Alexa fluor 647 (Donkey polyclonal) | Invitrogen | Cat#: A-21447 | IF(1:500) |
| Antibody | Anti-rabbit Alexa fluor Plus 555 (Donkey polyclonal) | Invitrogen | Cat#: A32794 | IF(1:500) |
| Antibody | Anti-rabbit Alexa fluor 647 (Donkey polyclonal) | Invitrogen | Cat#: A-31573 | IF(1:500) |
| Antibody | Anti-chicken Alexa fluor 488 (Goat polyclonal) | Invitrogen | Cat#: A-11039 | IF(1:500) |
| Peptide, recombinant protein | Dispase | Corning | Cat#: 3254235 | |
| Peptide, recombinant protein | Collagenase Type I | ThermoFisher | Cat#: 17100017 | |
| Software, algorithm | Cellranger v3.0.2 | 10X Genomics | RRID:SCR_017344 | |
| Software, algorithm | scanpy | scanpy | RRID:SCR_018139 | |
| Software, algorithm | scvelo | scVelo | RRID:SCR_018168 |
Mouse models
Request a detailed protocolAll mice were outbred on a C57BL6/CBA background, with male and female mice used in all experiments. The following transgenic mouse lines were used in this study, Twist2-Cre (RRID:MGI:3840442) (Cat# 008712). The ROSA26-CAG-flox-GFP-STOP-flox-Lef1 and Lef1-flox-Exon7,8-flox mice (RRID:MGI:5447957) were a kind gift and have been previously described (Lynch et al., 2016; Yu et al., 2012). Wild-type mice utilized in scRNA-seq experiments were an outbred background of C57BL6/CBA mice.
Wounding experiments
Request a detailed protocolAll wounding experiments were done in accordance with the guidelines from approved protocols Washington State University IACUC. Mice were wounded at post-natal day (P) 2, P24, P40 and P90 as stated in the results section. All wounding experiments were carried out with mice under anesthetization. The dorsal hair was shaved, and wounding areas were disinfected. For full-thickness circular wound, 2 mm and 8 mm punch biopsy and surgical scissors were used to create circular wounds on P2 and P24/P40 mice, respectively. For large wounds, 1.44 cm2 (1.2 × 1.2 cm) squares were excised using surgical scissors. Wounds were harvested at 7 dpw, 14 dpw, 24 dpw, and 30 dpw.
Horizontal whole mount
Request a detailed protocolThis procedure was described in Salz and Driskell, 2017. Briefly, full thickness skin samples were collected and fixed in 4% Paraformaldehyde (PFA), before being frozen in OCT compound in cryo-block. Samples were sectioned using cryostat at 60 micro m thickness. Skin sections were then stained with primary antibodies at 4°C overnight in PB buffer and with secondary antibodies in PB buffer for 1 hr. Samples were mounted on glass cover slip with glycerol as mounting medium.
Generation of single-cell RNA-sequencing data
Request a detailed protocolTissues were collected from dorsal regions of unwounded P2 and P21 mice, and right on the edge of 2 mm wounds 7 days post wound from P2 wound and P21 wound. Cells isolation procedure was previously described in Jensen and Driskell 2009 Nature Protocols (Jensen et al., 2010). Single-cell cDNA libraries were prepared using the 10X Genomics Chromium Single Cell 3’ kit. For each condition, three mice were used to generate three separate libraries, making it a total of 12 libraries across all four conditions. All samples were processed individually. The prepared libraries were sequenced on Illumina HiSeq 4000 (100 bp Paired-End). Demultiplexed Paired-End fastq files were aligned to mm10 reference genome using 10X Genomics CellRanger function (RRID:SCR_017344)(Cellranger version 3.0.2). FASTQ files for this manuscript can be access at NCBI Geo-Datasets: GSE153596. The outputs of CellRanger alignment were used to create loom files by Velocyto ‘run10x’ function (La Manno et al., 2018). Loom files are the preferred input data for scVelo package (RRID:SCR_018168) to perform RNA Velocity analysis (Bergen et al., 2020).
Single-cell RNA-seq data preprocessing
Request a detailed protocolFor basic filtering of our data, we filtered out cells have expressed less than 200 genes and genes that are expressed in less than three cells. To ensure that the data is comparable among cells, we normalized the number of counts per cell to 10,000 reads per cell as suggested by the Scanpy pipeline (RRID:SCR_018139) (Wolf et al., 2018). Data were then log-transformed for downstream analysis and visualization. We also regressed out effects of total counts per cell and the percentage of mitochondrial genes expressed, then scaled the data to unit variance with the maximum standard deviation 10.
Single-cell RNA-seq data analysis
Request a detailed protocolNeighborhood graph of cells was computed using PCA presentation (n PCs = 40, n neighbors = 10). The graph was embedded in two dimensions using Uniform Manifold Approximation and Projection (UMAP) as suggested by Scanpy developers. Clusters of cell types were defined by Louvain and Leiden method for community detection on the generated UMAP graph at resolution of 0.2. The analysis pipeline is published on the Driskell lab Github page as Jupyter Notebook (https://github.com/DriskellLab/Priming-Skin-to-Regenerate-by-Inducing-Lef1-Expression-in-Fibroblasts-; copy archived at https://github.com/elifesciences-publications/Priming-Skin-to-Regenerate-by-Inducing-Lef1-Expression-in-Fibroblasts-; DriskellLab, 2020).
Microarray analysis
Request a detailed protocolThe microarray data were previously published (GEO GSE11186) (Lin et al., 2009). RMA normalization was used, and the bottom twentieth percentile of genes were excluded from the analysis.
Data availability
We have generated a web-resource to easily query our large datasets: https://skinregeneration.org/ We have uploaded our data to GEO-Datasets: GSE153596 Our source code can be found on our Github webpage: https://github.com/DriskellLab/Priming-Skin-to-Regenerate-by-Inducing-Lef1-Expression-in-Fibroblasts- (copy archived at https://github.com/elifesciences-publications/Priming-Skin-to-Regenerate-by-Inducing-Lef1-Expression-in-Fibroblasts-).
-
NCBI Gene Expression OmnibusID GSE153596. Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds.
-
NCBI Gene Expression OmnibusID GSE11186. Expression profiling of mouse dorsal skin during hair follicle cycling.
References
-
Exploring the "hair growth-wound healing connection": anagen phase promotes wound re-epithelializationJournal of Investigative Dermatology 131:518–528.https://doi.org/10.1038/jid.2010.291
-
Generalizing RNA velocity to transient cell states through dynamical modelingNature Biotechnology 14:0591-3.https://doi.org/10.1038/s41587-020-0591-3
-
Murine models of human wound healingMethods in Molecular Biology 1037:265–274.https://doi.org/10.1007/978-1-62703-505-7_15
-
Understanding fibroblast heterogeneity in the skinTrends in Cell Biology 25:92–99.https://doi.org/10.1016/j.tcb.2014.10.001
-
Lef1 and Tcf1 expression in dermal fibroblasts regulates the group size of hair follicles in developing skinArticle in Preparation.
-
Basal cell carcinomas in mice overexpressing Gli2 in skinNature Genetics 24:216–217.https://doi.org/10.1038/73417
-
Age-related evolutions of the dermis: clinical signs, fibroblast and extracellular matrix dynamicsMechanisms of Ageing and Development 177:150–156.https://doi.org/10.1016/j.mad.2018.03.006
-
Embryonic skin development and repairOrganogenesis 14:46–63.https://doi.org/10.1080/15476278.2017.1421882
-
Fate of Prominin-1 expressing dermal papilla cells during homeostasis, wound healing and wnt activationJournal of Investigative Dermatology 135:2926–2934.https://doi.org/10.1038/jid.2015.319
-
Scarless fetal skin wound healing updateBirth Defects Research. Part C, Embryo Today : Reviews 96:237–247.https://doi.org/10.1002/bdrc.21018
-
β-Catenin stabilization in skin fibroblasts causes fibrotic lesions by preventing adipocyte differentiation of the reticular DermisJournal of Investigative Dermatology 136:1130–1142.https://doi.org/10.1016/j.jid.2016.01.036
-
A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stagesJournal of Investigative Dermatology 117:3–15.https://doi.org/10.1046/j.0022-202x.2001.01377.x
-
Epithelial wnt ligand secretion is required for adult hair follicle growth and regenerationJournal of Investigative Dermatology 133:31–41.https://doi.org/10.1038/jid.2012.230
-
Tcf3 and Tcf4 are essential for long-term homeostasis of skin epitheliaNature Genetics 41:1068–1075.https://doi.org/10.1038/ng.431
-
Spatial and Single-Cell transcriptional profiling identifies functionally distinct human dermal fibroblast subpopulationsJournal of Investigative Dermatology 138:811–825.https://doi.org/10.1016/j.jid.2018.01.016
-
Horizontal whole mount: a novel processing and imaging protocol for thick, Three-dimensional tissue Cross-sections of skinJournal of Visualized Experiments 2:56106.https://doi.org/10.3791/56106
-
Dermal Blimp1 acts downstream of epidermal tgfβ and wnt/β-Catenin to Regulate Hair Follicle Formation and GrowthJournal of Investigative Dermatology 137:2270–2281.https://doi.org/10.1016/j.jid.2017.06.015
-
A mouse fetal skin model of scarless wound repairJournal of Visualized Experiments 16:52297.https://doi.org/10.3791/52297
-
Scarless wound healing: chasing the holy grailPlastic and Reconstructive Surgery 135:907–917.https://doi.org/10.1097/PRS.0000000000000972
-
Review of hair follicle dermal cellsJournal of Dermatological Science 57:2–11.https://doi.org/10.1016/j.jdermsci.2009.11.005
Article and author information
Author details
Funding
National Institute of Arthritis and Musculoskeletal and Skin Diseases (1 R56 AR073778-01A1)
- Ryan R Driskell
WSU New Faculty Seed (133408)
- Ryan R Driskell
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
This work was supported by NIAMS R56 AR073778-01A1 and a WSU New Faculty Seed Grant 2428–9926. The authors acknowledge Sean Thompson, Jonathan Jones, Kai Kretzschmar, and Klaas Mulder for their discussion and help.The authors acknowledge the Washington State University Franceschi Microscopy Imaging Center for the continued support in microscopic imaging, and Jared Brannan for generation of our web resource.
Ethics
Animal experimentation: This study was performed in strict accordance with recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All of the animals were handled according to approved institutional animal care and use committee (IACUC) protocols (#5033/6724 and #5002/6723) of Washington State University.
Version history
- Received: June 16, 2020
- Accepted: September 9, 2020
- Version of Record published: September 29, 2020 (version 1)
Copyright
© 2020, Phan et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 14,684
- views
-
- 1,300
- downloads
-
- 92
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds
eLife 9:e60066.
https://doi.org/10.7554/eLife.60066
Further reading
-
- Neuroscience
- Stem Cells and Regenerative Medicine
While accumulated publications support the existence of neurogenesis in the adult human hippocampus, the homeostasis and developmental potentials of neural stem cells (NSCs) under different contexts remain unclear. Based on our generated single-nucleus atlas of the human hippocampus across neonatal, adult, aging, and injury, we dissected the molecular heterogeneity and transcriptional dynamics of human hippocampal NSCs under different contexts. We further identified new specific neurogenic lineage markers that overcome the lack of specificity found in some well-known markers. Based on developmental trajectory and molecular signatures, we found that a subset of NSCs exhibit quiescent properties after birth, and most NSCs become deep quiescence during aging. Furthermore, certain deep quiescent NSCs are reactivated following stroke injury. Together, our findings provide valuable insights into the development, aging, and reactivation of the human hippocampal NSCs, and help to explain why adult hippocampal neurogenesis is infrequently observed in humans.
-
- Stem Cells and Regenerative Medicine
We developed a 96-well plate assay which allows fast, reproducible, and high-throughput generation of 3D cardiac rings around a deformable optically transparent hydrogel (polyethylene glycol [PEG]) pillar of known stiffness. Human induced pluripotent stem cell-derived cardiomyocytes, mixed with normal human adult dermal fibroblasts in an optimized 3:1 ratio, self-organized to form ring-shaped cardiac constructs. Immunostaining showed that the fibroblasts form a basal layer in contact with the glass, stabilizing the muscular fiber above. Tissues started contracting around the pillar at D1 and their fractional shortening increased until D7, reaching a plateau at 25±1%, that was maintained up to 14 days. The average stress, calculated from the compaction of the central pillar during contractions, was 1.4±0.4 mN/mm2. The cardiac constructs recapitulated expected inotropic responses to calcium and various drugs (isoproterenol, verapamil) as well as the arrhythmogenic effects of dofetilide. This versatile high-throughput assay allows multiple in situ mechanical and structural readouts.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}