Genomic stability of self-inactivating rabies
- MRC Laboratory of Molecular Biology, United Kingdom
Figures
Figure 1
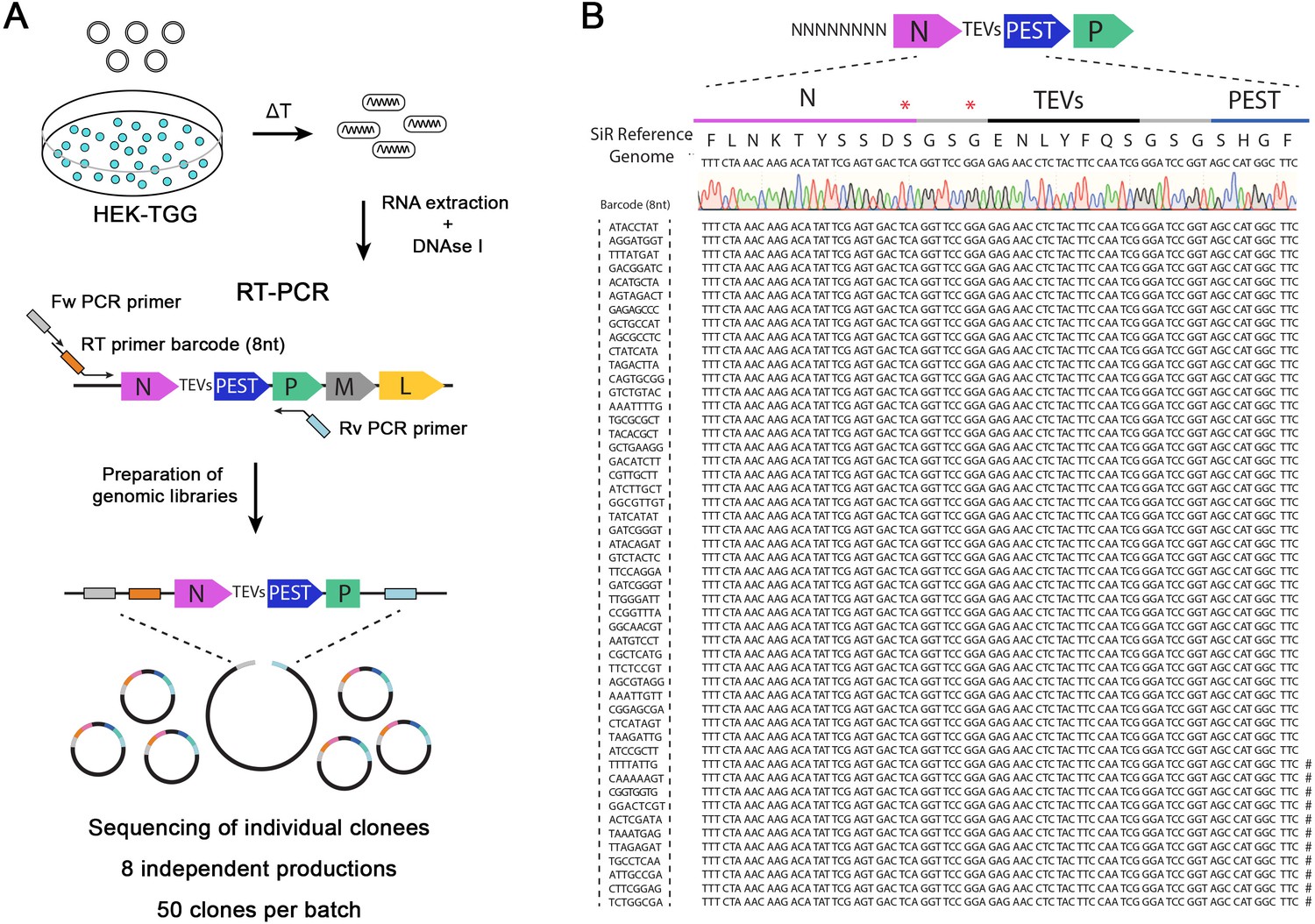
SiR production from cDNA leads to revertant-free viral preparations.
(A) Scheme of experimental strategy to identify the emergence of “revertant” mutations during SiR production. 8 independent SiR preparations were rescued from cDNA and genomic RNA were extracted, treated with DNAse I, subjected to RT-PCR to amplify N-TEVs-PEST coding sequence and used to generate libraries for Sanger sequencing (50 clones per preparation were sequenced). (B) Example of sequencing results from one SiR preparation showing no mutations at the end of N. Symbols (#) show the position of previously identified mutations, marks on the sequences indicates the presence of mutations in different positions.
Figure 2 with 2 supplements
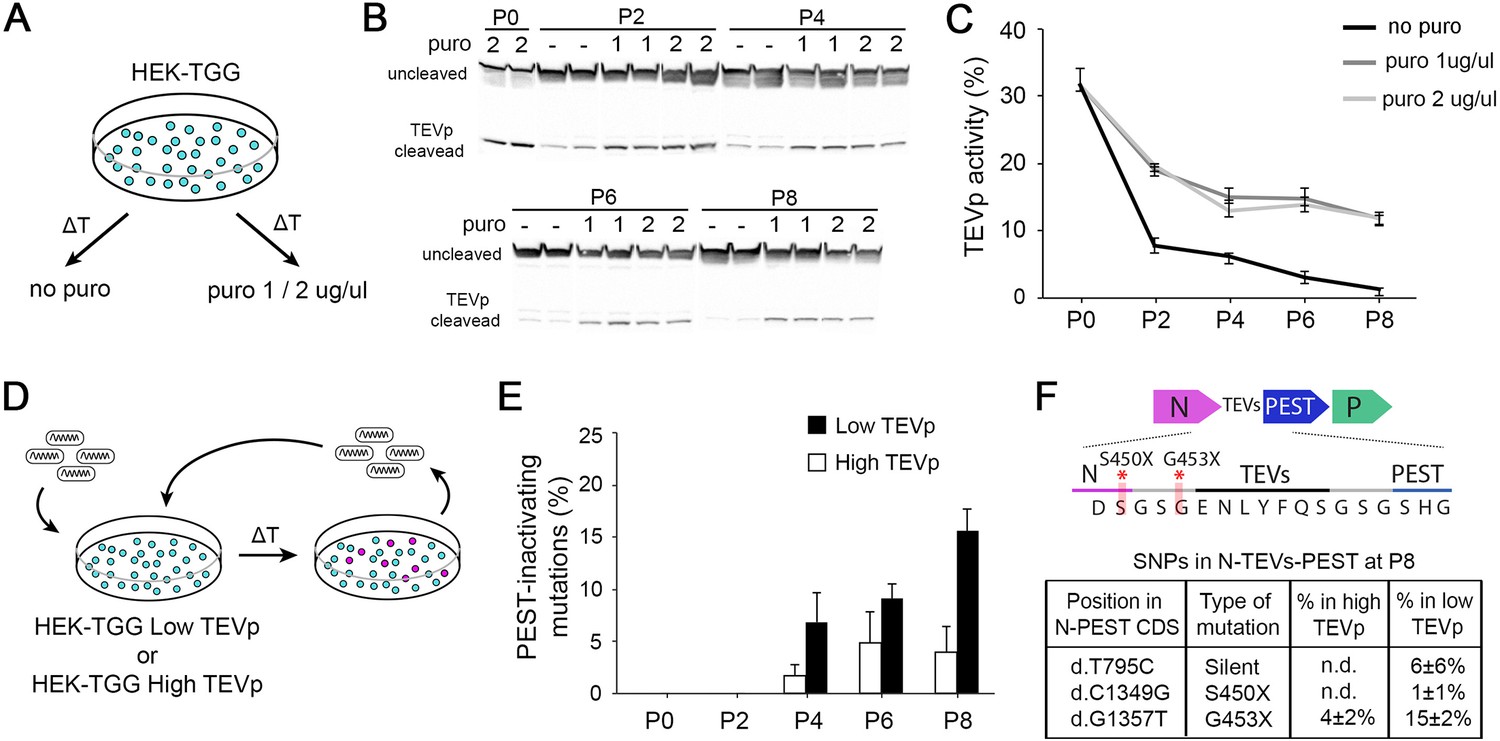
High TEVp activity in packaging cells prevents accumulation of PEST-mutations.
(A) HEK-TGG packaging cells were amplified for several passages in absence or presence (1 or 2 μg/ml) of puromycin selection. (B) TEVp-dependent cleavage of TEVp-activity reporter was analysed by western blot in HEK-TGG at different amplification passages. (C) Quantification of TEVp-activity in packaging cells over time in presence or absence of antibiotic pressure. (mean ± SEM, n=3) (D) Experimental design to assess emergence of mutations in SiR preparations after multiple passages of amplification in high TEVp (HEK-TGG P0) or low TEVp HEK-TGG (HEK-TGG P8, without puromycin selection). (E) Quantification of frequency of the accumulation of PEST-targeting mutations over time that prevent translation of PEST domain (mean ± SEM, n=4 independent viral preparation). (F) Summary of the single nucleotide polymorphisms (SNPs) in the coding sequence (CDS) of N-TEVsPEST that reached threshold at P8 (mean ± SEM, n=4; n.d. indicates that the mutations were not detected above threshold). Top scheme shows the position of PEST-inactivating mutations.
-
Figure 2—source data 1
Individual Western Blots used in Figure 2B.
- https://cdn.elifesciences.org/articles/83459/elife-83459-fig2-data1-v2.zip
-
Figure 2—source data 2
TEVp-activity in HEK-TGG packaging cells over time.
- https://cdn.elifesciences.org/articles/83459/elife-83459-fig2-data2-v2.xlsx
Figure 2—figure supplement 1
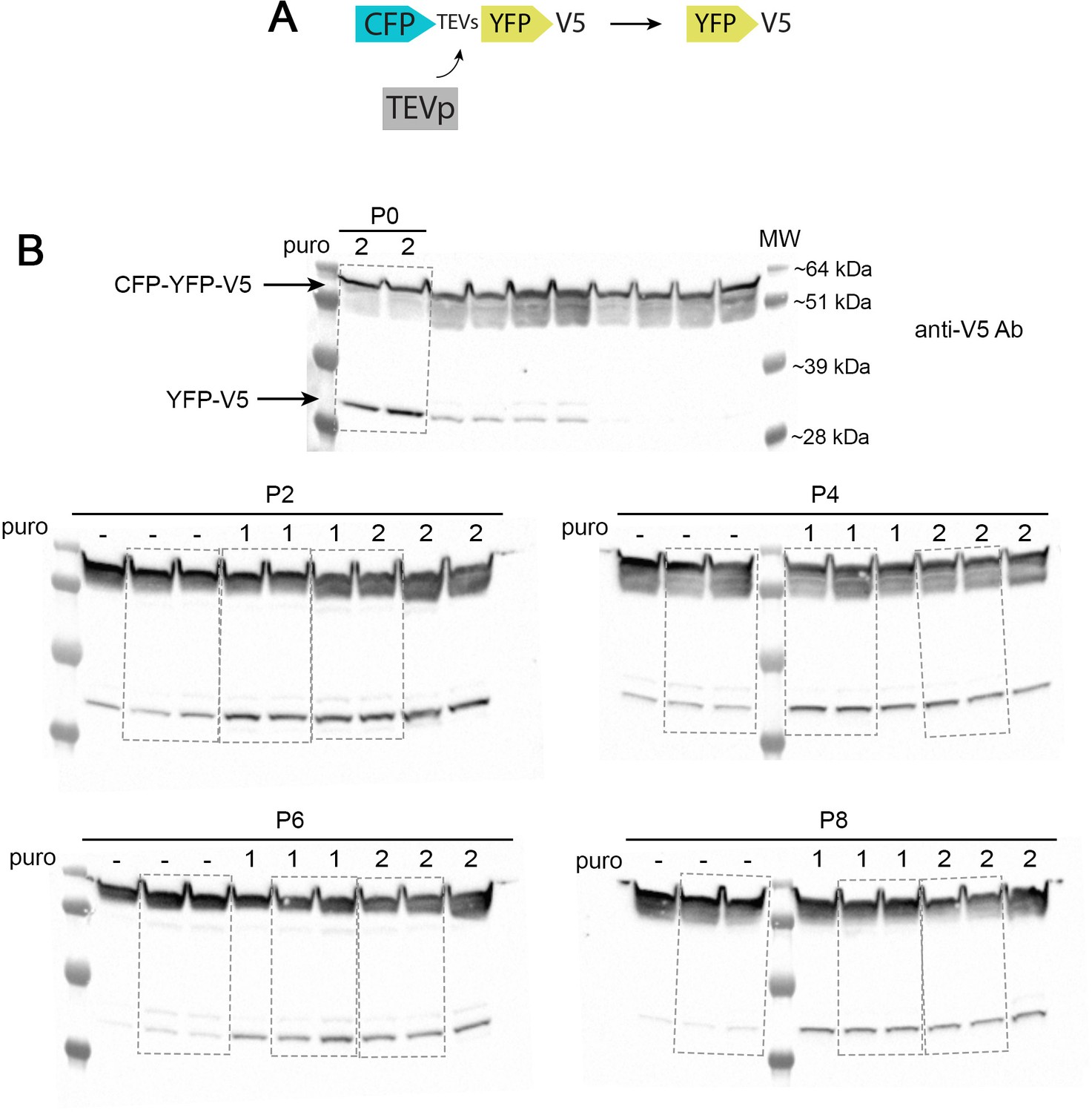
Western blots to test TEVp in packaging cells over time.
(A) Scheme of the TEVp activity reporter. (B) Original western blots stained with an anti-V5 antibody with the representative lanes used to generate Figure 2B.
Figure 2—figure supplement 2

SMRT sequencing of SiR genomic libraries.
Scheme of the strategy to sequence SiR preparations using SMRT NGS technology from Pacbio. Amplicons of the entire coding sequence of N-TEVs-PEST gene are generated by RT-PCR. Unique Molecular Identifier (UMI) of 10 nucleotides is added during retrotranscription to each genomic molecule and sample specific barcodes of 16 nucleotides are added at the two ends during subsequent PCR. SMRT bell libraries are generated by ligating the provided adapters to generate circular DNA molecules that are sequenced continuously for multiple passages. Subreads are used to generate high-fidelity consensus sequences that are demultiplexed using the 16 nt barcodes, deduplicated using the UMIs and aligned to the reference for variant calling.
Figure 3 with 2 supplements
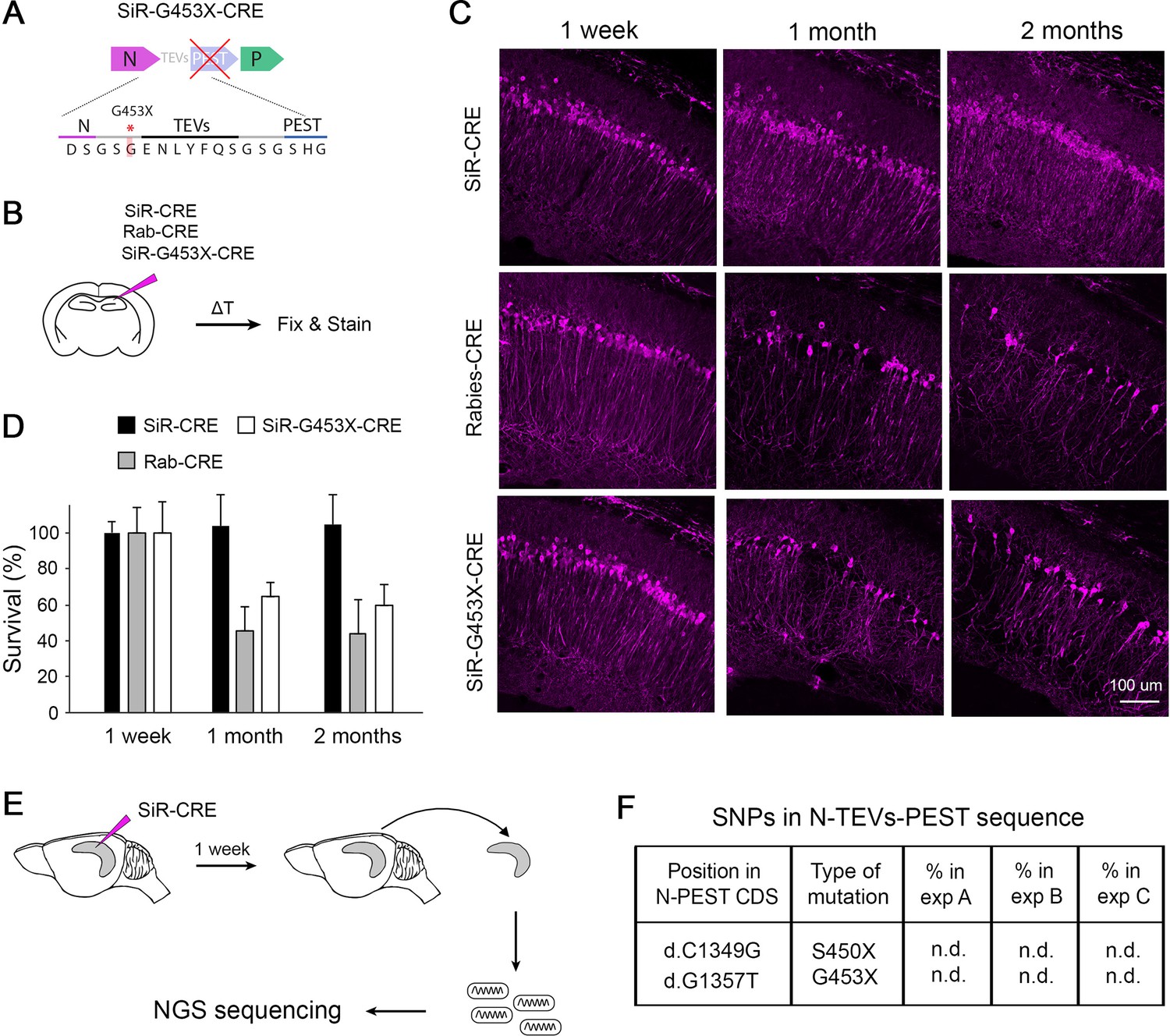
Revertant-free SiR, but not PEST-mutant, is non-toxic and does not accumulate PEST-targeting mutations in vivo.
(A) Scheme of the engineered PEST-mutant SiR (SiR-G453X). (B) Experimental procedure. (C) Confocal images of hippocampal sections of Rosa26LSL-tdTomato mice infected with SiR-CRE, Rab-CRE, SiR-G453X and imaged at 1 week, 1 month and 2 months p.i. Scale bar, 50 μm. (D) Number of tdTomato positive neurons at 1 week, 1 months, and 2 months p.i. normalized to 1 week time point (mean ± SEM, n=4 animals per virus per time point). (E) Experimental procedure for the sequencing of SiR particles from injected hippocampi at 1 week p.i. (F) List of PEST-inactivating mutations above 2% thresholds with relative frequency in each animal (n.d. indicates that the mutation was not detected above threshold; n=3 animals).
-
Figure 3—source data 1
tdTomato+ positive neurons in injected Hippocampi with Rab, SiR or Pest-mutant SiR.
- https://cdn.elifesciences.org/articles/83459/elife-83459-fig3-data1-v2.xlsx
Figure 3—figure supplement 1
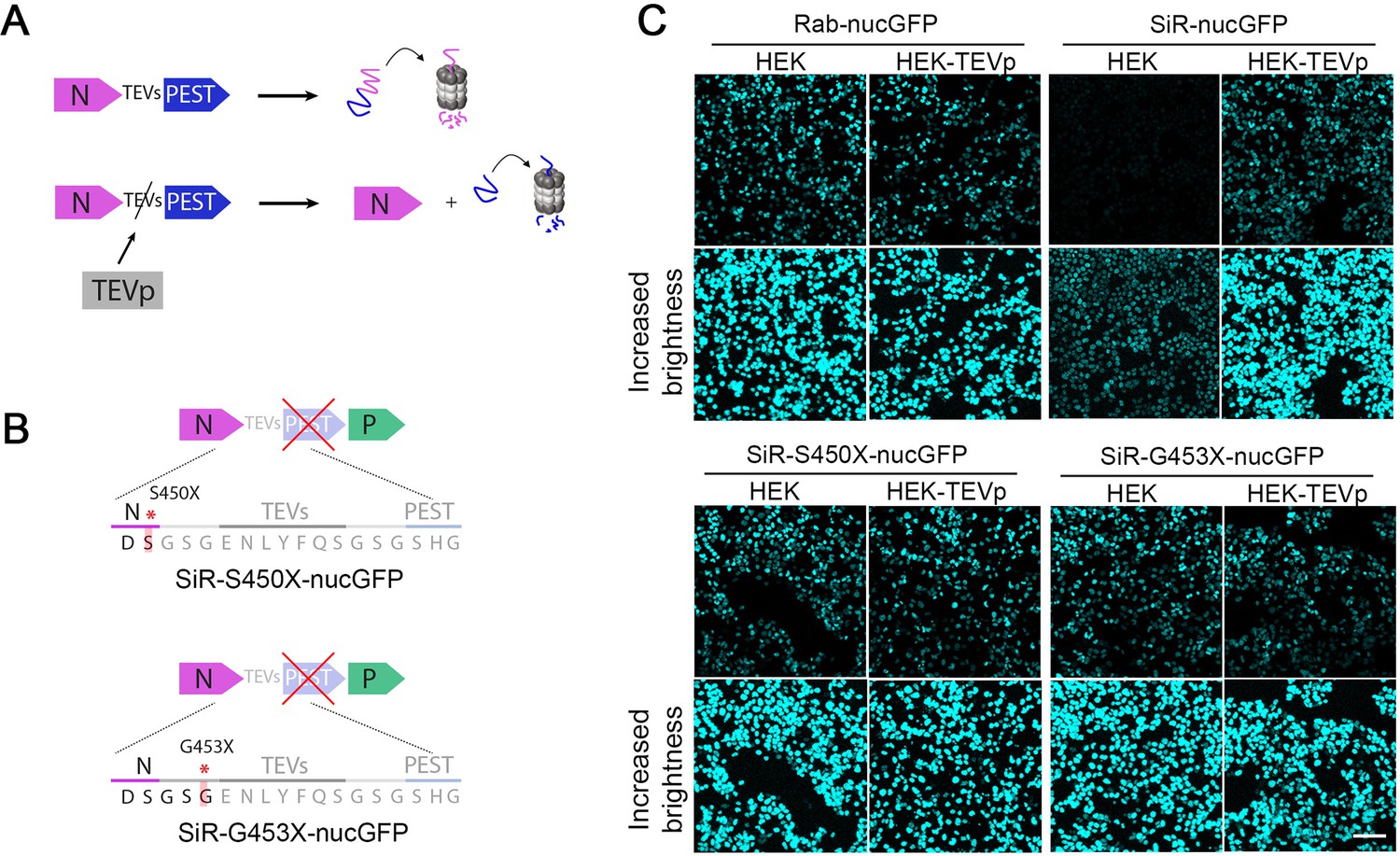
SiR revertants lose functional TEVs and PEST domain.
(A) The conditional destabilization of N can be prevented by TEVp expression in the infected cells leading to cleavage of the TEVs-containing linker. (B) Engineered revertant SiR viruses containing the reporter PEST-inactivating substitutions in their cDNA. (C) Confocal images of HEK and HEK-TEVp at 48 hrs p.i. All images were acquired with same settings. Bottom panels have been equally adjusted in brightness in all conditions. Scale bar 100 μm.
Figure 3—figure supplement 2
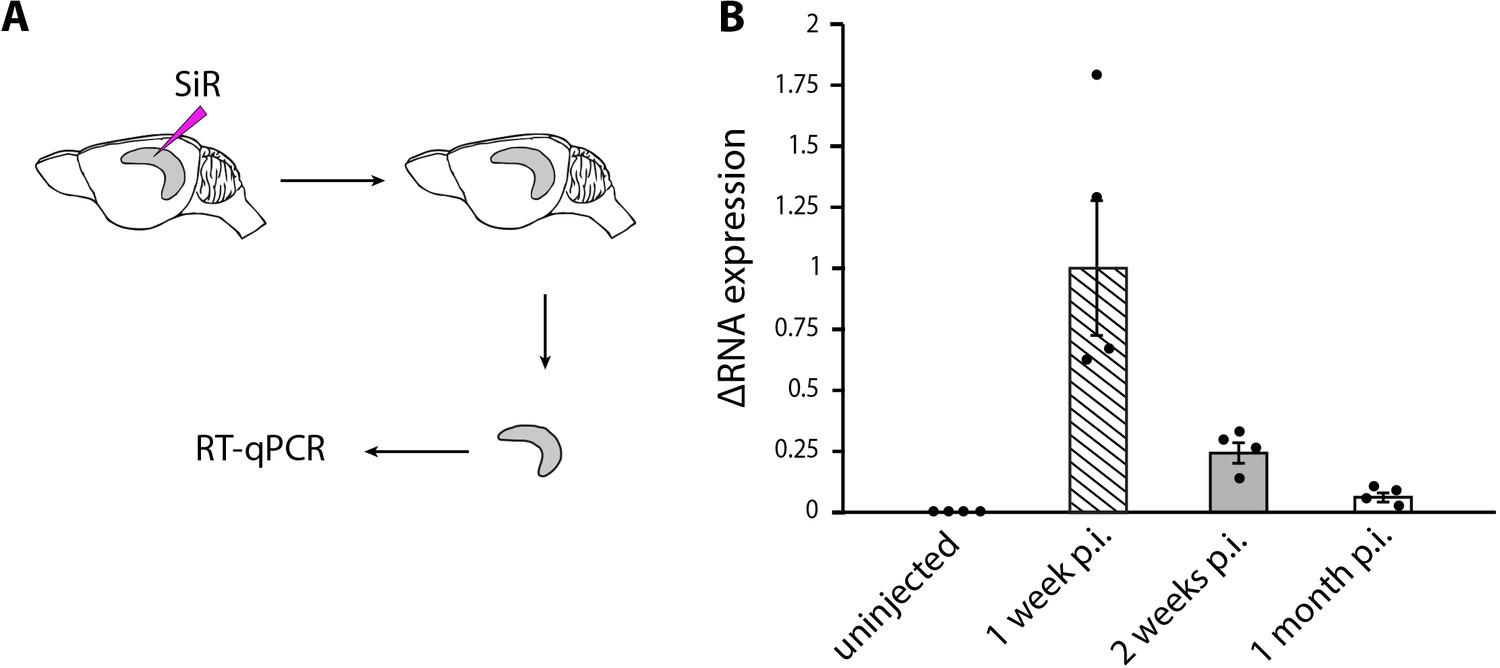
SiR RNA in injected hippocampi.
(A) Schematic of SiR-CRE injection in the hippocampus mice followed by total RNA extraction and RT-qPCR. (B) Levels of viral RNA normalized to 1 week RNA level (mean ± SEM, n=4 animals per time point).
Figure 4 with 1 supplement
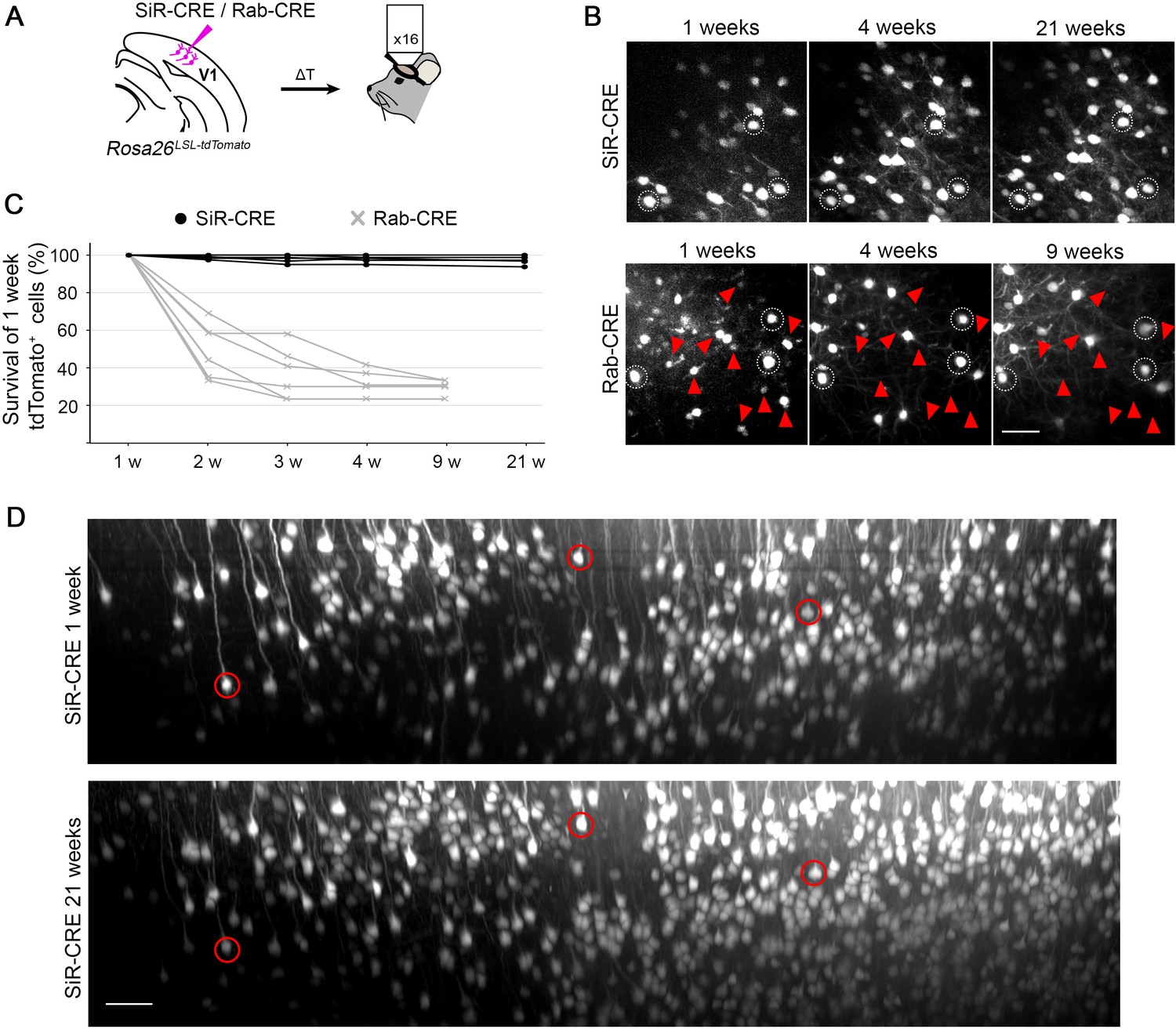
2-photon in vivo longitudinal imaging of revertant-free SiR-infected cortical neurons reveals no toxicity and unaltered neuronal morphology after 5 months.
(A) Schematic of SiR-CRE or Rab-CRE injection in Rosa26LSL-tdTomato mice in V1 followed by in vivo imaging. (B) Two-photon maximal projection of the same field in SiR-CRE and RabCRE injected cortices at 1, 4, and 21 weeks p.i. or 1, 4, and 9 weeks, respectively. Red arrowheads mark tdTomato positive neurons detected at 1 week that disappear in later recordings. Scale bar 50 μm. (C) Survival of the tdTomato-positive cells recorded at 1 week over time. (ROIs = 6 per virus. n=2 animals per virus). (D) Two-photon maximal projection of the same large field in SiR-CRE injected cortices at 1 week and 21 weeks p.i. Scale bar 50 μm.
-
Figure 4—source data 1
tdTomato+ positive neurons in injected cortices with Rab or SiR.
- https://cdn.elifesciences.org/articles/83459/elife-83459-fig4-data1-v2.xlsx
Figure 4—figure supplement 1

Two-photon in vivo longitudinal imaging of revertant-free SiR-infected cortical neurons.
Two-photon maximal projection of the same fields in SiR-CRE injected cortices at 1-2-3-4-21 weeks p.i.
Figure 5
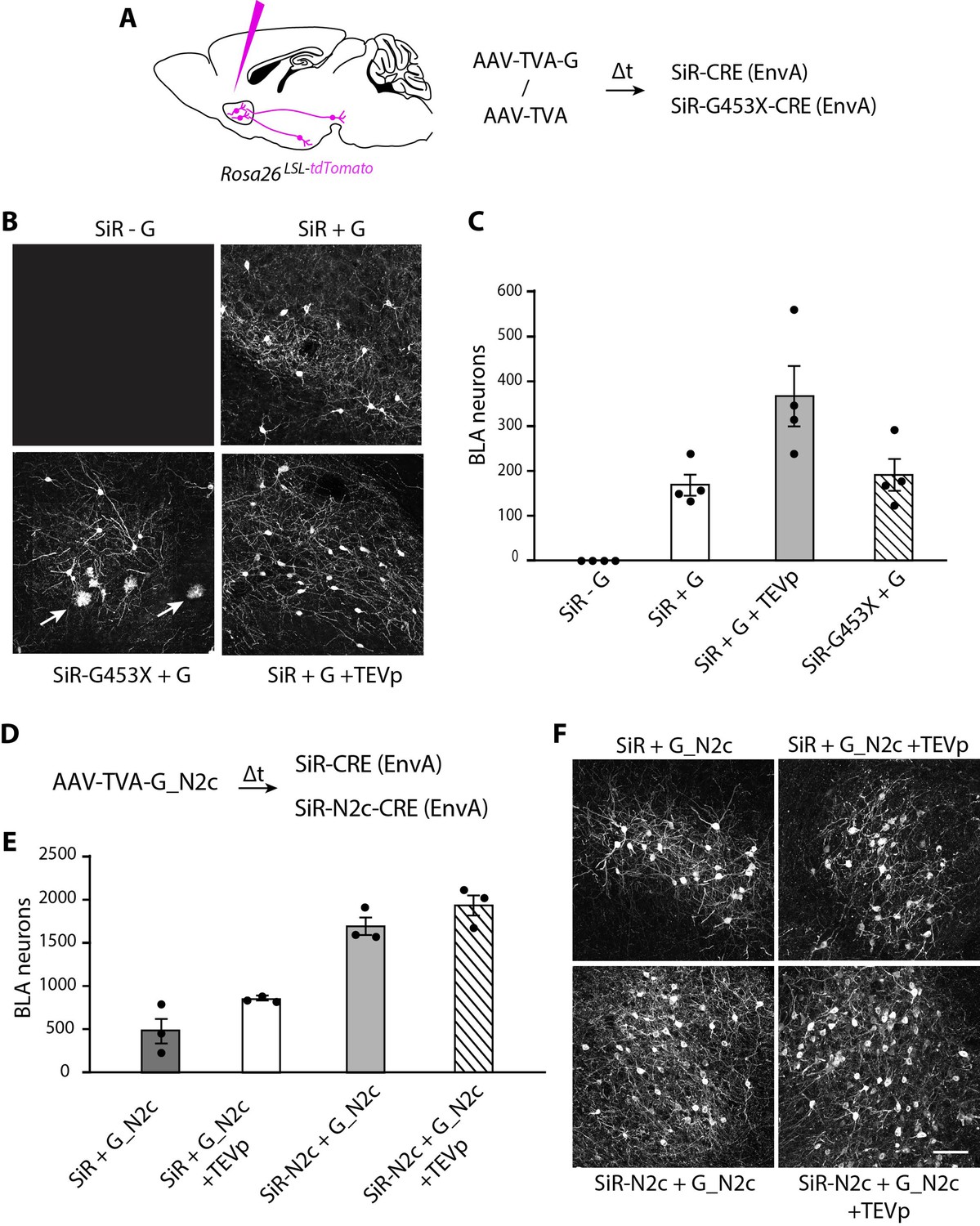
SiR vectors transsynaptic tracing of neural circuits in the central nervous system.
(A) Experimental design for the transsynaptic tracing of NAc inputs using EnvA-pseudotyped SiR-CRE or SiR-G453X-CRE in Rosa26LSL-tdTomato mice. (B) Confocal images of BLA area of Rosa26LSL-tdTomato mice infected with SiR-CRE or SiR-G453X-CRE. Arrows point to tdTomato+ microglia. (C) Number of tdTomato-positive neurons in the BLA at 1 month post SiR injection (mean ± SEM, n=4 animals per condition). (E) Number of tdTomato+ neurons in the BLA at 1 month post SiR injection (mean ± SEM, n=3 animals per condition). (F) Confocal images of BLA area of Rosa26LSL-tdTomato mice infected with SiR-CRE or SiR-N2c-CRE. Scale bar, 100 μm.
-
Figure 5—source data 1
tdTomato+ positive BLA neurons upon transsynaptically tracing with SiR, Pest-mutant SiR or SiR-N2c.
- https://cdn.elifesciences.org/articles/83459/elife-83459-fig5-data1-v2.xlsx
Tables
Table 1
List of detected mutations in SiR viruses rescued from cDNA divided by batch (50 individual clones per batch).
The position of the mutations is calculated referring to +1 as the first base of the nucleoprotein N coding sequence.
| Sanger sequencing results of SiRs rescued from cDNA | |||||
|---|---|---|---|---|---|
| Batch A | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 1/50 | GAT >GAC | –54 | Substitution | - |
| 1/50 | AAA >AAG | –18 | Substitution | - | |
| N gene | 1/50 | GCC >GCT | +186 | Substitution | Synonymous A62 |
| 1/50 | TTT >TTTT | +243 | Insertion | Frameshift | |
| 1/50 | AAG >A-G | +485 | Deletion | Frameshift | |
| 1/50 | ATG >CTG | +562 | Substitution | Missense M188L | |
| 1/50 | GTG >G-- | +677/8 | Deletion | Frameshift | |
| 1/50 | ACG >ACCG | +983 | Insertion | Frameshift | |
| 1/50 | GAA >AAA | +1,093 | Substitution | E365K | |
| 1/50 | TCA >CCA | +1,276 | Substitution | S426P | |
| TEVs-PEST | - | - | - | - | - |
| Intergenic N/P | 4/50 | AAA >AAAA | +1,571 | Insertion | - |
| 1/50 | CCC >CCA | +1,581 | Substitution | - | |
| P | - | - | - | - | - |
| Batch B | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 1/50 | AAC >A-C | –63 | Deletion | - |
| 1/50 | CAA >CA- | –60 | Deletion | ||
| 1/50 | CTA >CTG | -3 | Substitution | - | |
| N gene | 1/50 | TTT >TTTT | +243 | Insertion | Frameshift |
| 1/50 | GAC >GAA | +501 | Substitution | D167E | |
| 1/50 | AAT >AAC | +588 | Substitution | Synonymous N196 | |
| 1/50 | GCT >GCC | +1,002 | Substitution | Synonymous A334 | |
| 1/50 | AAA >AAAA | +1,056 | Insertion | Frameshift | |
| TEVs-PEST | 1/50 | TCC >TGC | +1,385 | Substitution | Missense S462C in GSG linker after TEVs |
| Intergenic N/P | 1/50 | TAT >TAA | +1,554 | Substitution | - |
| 2/50 | AAA >AAAA | +1,571 | Insertion | - | |
| P | 1/50 | GAA >GAG | +1,671 | Substitution | Synonymous E23 |
| 1/50 | CTG >CCG | +1,775 | Substitution | Missense L58P | |
| 1/50 | GGA >TGA | +2014 | Deletion | Nonsense G138>STOP | |
| Batch C | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 2/50 | AAA >AAAA | –43 | Insertion | - |
| N gene | 1/50 | TGT >TTT | +212 | Substitution | Missense C71F |
| 1/50 | AGA >AGG | +1,074 | Substitution | Synonymous R358 | |
| 1/50 | GGT >GAT | +1,190 | Substitution | Missense G397D | |
| TEVs-PEST | - | - | - | - | - |
| Intergenic N/P | 1/50 | AAA >AAG | +1,569 | Substitution | - |
| 3/50 | AAA >AAAA | +1,571 | Insertion | - | |
| 1/50 | AAA >AA- | +1,571 | Deletion | ||
| P | 1/50 | CAA >AAA | +1,720 | Substitution | Missense Q40K |
| Batch D | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | - | - | - | - | - |
| N gene | 1/50 | AAG >AGG | +113 | Substitution | Missense K38R |
| 1/50 | AAA >CAA | +295 | Substitution | Missense K99Q | |
| 1/50 | CAT >AAT | +655 | Substitution | Missense H219N | |
| 1/50 | TCA >TCC | +873 | Substitution | Synonymous S291 | |
| 1/50 | ACC >AAC | +1,196 | Substitution | Missense T399N | |
| TEVs-PEST | - | - | - | - | - |
| Intergenic N/P | 3/50 | AAA >AAAA | +1,571 | Insertion | - |
| 1/50 | ATC >ATT | +1,596 | Substitution | - | |
| P | 1/50 | AAA >AAAA | +1,671 | Insertion | Frameshift |
| 1/50 | CGT >CTA | +1,878 | Substitution | Synonymous L92 | |
| 1/50 | AGA >AGT | +1941 | Substitution | Missense R113S | |
| 1/50 | GGA >GGG | +2016 | Substitution | Synonymous G138 | |
| 1/50 | ACT >ACA | +2046 | Substitution | Synonymous T148 | |
| Batch E | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 1/50 | CCA >CC- | –57 | Deletion | - |
| N gene | 1/50 | CCT >CAT | +200 | Substitution | Missense P67H |
| 1/50 | TTT >TTTT | +243 | Insertion | Frameshift | |
| 1/50 | GGA >GAA | +371 | Substitution | Missense G124E | |
| 1/50 | ACA >ACG | +387 | Substitution | Synonymous T129 | |
| 2/50 | GAC >GAT | +393 | Substitution | Synonymous D131 | |
| 1/50 | CAC >C-- | +551/2 | Deletion | Frameshift | |
| 1/50 | ACT >AAT | +557 | Substitution | T186N | |
| 1/50 | TTT >TTTT | +779 | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - |
| Intergenic N/P | 1/50 | CAT >CAC | +1,560 | Substitution | - |
| 1/50 | AAA >AAC | +1,570 | Substitution | ||
| 4/50 | AAA >AAAA | +1,571 | Insertion | ||
| 1/50 | ATC >ATT | +1,596 | Substitution | - | |
| P | 1/50 | GAA >GGA | +1,667 | Substitution | Missense E22G |
| Batch F | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 1/50 | ACC >AC- | –58 | Deletion | - |
| 1/50 | CAG >CA- | –56 | Deletion | - | |
| 1/50 | TCA >TCG | –52 | Substitution | - | |
| 1/50 | AAA >AAAA | –43 | Insertion | - | |
| 1/50 | AAG >AA- | –22 | Deletion | - | |
| N gene | 1/50 | TTT >TTTTT | +243/4 | Insertion | Frameshift |
| 1/50 | TTG >TCG | +434 | Substitution | Missense L145S | |
| 1/50 | TTT >TT- | +534 | Deletion | Frameshift | |
| 1/50 | GCA >GTA | +767 | Substitution | Missense A256V | |
| 1/50 | ACA >ATA | +836 | Substitution | Missense T279I | |
| 1/50 | AAA >AAAA | +908 | Insertion | Frameshift | |
| 1/50 | 321 bp | +1041–1,362 | Deletion | Deletion of C-terminal of N in frame with PEST domain | |
| 1/50 | GGA >GGG | +1,038 | Substitution | Synonymous G346 | |
| TEVs-PEST | - | - | - | - | - |
| Intergenic N/P | 4/50 | AAA >AAAA | +1,571 | Insertion | - |
| P | 1/50 | CCT >CCC | +1,626 | Substitution | Synonymous P8 |
| 1/50 | GAA >GGA | +1,727 | Substitution | Missense E42G | |
| 1/50 | TTT >TTC | +1,845 | Substitution | Synonymous F81 | |
| Batch G | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 1/50 | CCA >CC- | –57 | Deletion | - |
| 1/50 | AAA >AA- | –16 | Deletion | - | |
| N gene | 1/50 | GCA >GTA | +290 | Substitution | Missense A97V |
| 1/50 | CAT >GAT | +409 | Substitution | Missense H137D | |
| 1/50 | TTT >TT- | +534 | Deletion | Frameshift | |
| 1/50 | TAT >TGT | +1,271 | Substitution | Missense Y424C | |
| 1/50 | GCC >GTC | +1,316 | Substitution | Missense A439V | |
| TEVs-PEST | - | - | - | - | - |
| Intergenic N/P | 4/50 | AAA >AAAA | +1,571 | Insertion | - |
| P | 1/50 | AAA >CAA | +1,786 | Substitution | Missense K62Q |
| 1/50 | GAA >GGA | +1,823 | Substitution | Missense E74G | |
| 1/50 | CGA >CAA | +1,834 | Substitution | Missense R78Q | |
| Batch H | |||||
| Clones | Sequence | Position | Mutation | Effect on CDS | |
| Upstream N | 1/50 | AAA >AAAA | –43 | Insertion | - |
| 1/50 | AAC >AA- | –42 | Deletion | ||
| N gene | 1/50 | TTA >CTA | +145 | Substitution | Synonymous L49 |
| 1/50 | ATG >ATA | +234 | Substitution | Missense M78I | |
| 1/50 | TTT >TTTT | +243 | Insertion | Frameshift | |
| 1/50 | AAA >CAA | +295 | Substitution | Missense K99Q | |
| 1/50 | GAT >AAT | +301 | Substitution | Missense D101N | |
| 1/50 | GGA >AGA | +622 | Substitution | Missense G208R | |
| 1/50 | GCT >TCT | +838 | Substitution | Missense A280S | |
| 1/50 | GGC >G-C | +1,028 | Deletion | Frameshift | |
| 1/50 | GAC >AAC | +1,132 | Substitution | Missense D378N | |
| TEVs-PEST | 1/50 | CTG >CTA | +1,437 | Substitution | Synonymous L16 in PEST domain |
| Intergenic N/P | 3/50 | AAA >AAAA | +1,571 | Insertion | - |
| 1/50 | AAC >AAA | +1,592 | Substitution | - | |
| P | 1/50 | AAA >AAAA | +1,788 | Insertion | Frameshift |
Table 2
List of detected mutations above 2% thresholds in SiR viruses amplified in high- and low-TEVp packaging cells sequenced by SMRT NGS sequencing.
The position of the mutations is defined considering +1 the first base of the nucleoprotein N coding sequence.
| NGS sequencing results of SiRs amplified for multiple passages in vitro | ||||||
|---|---|---|---|---|---|---|
| SIR-A-P0 bc1—bc2 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 302/6608 | 4.5% | Insertion | - |
| N gene | +237 | +T | 266/6598 | 4.0% | Insertion | Frameshift |
| +636 | +T | 190/6595 | 2.9% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 732/6556 | 11.1% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-B-P0 bc1—bc3 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 276/6045 | 4.6% | Insertion | - |
| N gene | +237 | +T | 274/6037 | 4.5% | Insertion | Frameshift |
| +636 | +T | 180/6036 | 3.0% | Insertion | Frameshift | |
| TEVs-PEST | +1,359 | A>T | 246/5879 | 4.2% | Substitution | Silent G453 |
| Intergenic | +1,564 | +A | 729/6556 | 12.1% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-C-P0 bc1—bc4 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 256/5137 | 5.0% | Insertion | - |
| N gene | +237 | +T | 227/5137 | 4.4% | Insertion | Frameshift |
| +636 | +T | 167/5138 | 3.3% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 598/5140 | 11.6% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-D-P0 bc1—bc5 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 249/5419 | 4.6% | Insertion | - |
| N gene | +237 | +T | 229/5419 | 4.2% | Insertion | Frameshift |
| +636 | +T | 125/5422 | 2.3% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 612/5420 | 11.3% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-A-HighTEVp-P2 bc2—bc4 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 245/5934 | 4.1% | Insertion | - |
| N gene | +237 | +T | 297/5933 | 5.0% | Insertion | Frameshift |
| +636 | +T | 157/5938 | 2.6% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 634/5935 | 10.7% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-B-HighTEVp-P2 bc2—bc5 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 281/5750 | 4.9% | Insertion | - |
| N gene | +237 | +T | 272/5752 | 4.7% | Insertion | Frameshift |
| +636 | +T | 170/5752 | 3.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 625/5749 | 10.9% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-C-HighTEVp-P2 bc2—bc6 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 236/4773 | 4.9% | Insertion | - |
| N gene | +237 | +T | 241/4772 | 5.1% | Insertion | Frameshift |
| +636 | +T | 137/4774 | 2.9% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 489/4776 | 10.2% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-D-HighTEVp-P2 bc2—bc6 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 260/5591 | 4.7% | Insertion | - |
| N gene | +237 | +T | 238/5595 | 4.3% | Insertion | Frameshift |
| +636 | +T | 150/5597 | 2.7% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 550/5594 | 9.8% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-A-LowTEVp-P2 bc1—bc6 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 197/3891 | 5.1% | Insertion | - |
| N gene | +237 | +T | 194/3891 | 5.0% | Insertion | Frameshift |
| +636 | +T | 116/3892 | 3.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 447/3891 | 11.5% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-B-LowTEVp-P2 bc1—bc7 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 244/5050 | 4.8% | Insertion | - |
| N gene | +237 | +T | 227/5055 | 4.5% | Insertion | Frameshift |
| +636 | +T | 162/5055 | 3.2% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 503/5055 | 10.0% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-C-LowTEVp-P2 bc1—bc8 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 266/5050 | 5.3% | Insertion | - |
| N gene | +237 | +T | 248/5050 | 4.9% | Insertion | Frameshift |
| +636 | +T | 146/5056 | 2.9% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 547/5054 | 10.8% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-D-LowTEVp-P2 bc1—bc9 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 200/5295 | 3.8% | Insertion | - |
| N gene | +237 | +T | 204/5295 | 3.9% | Insertion | Frameshift |
| +636 | +T | 141/5297 | 2.7% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 456/5297 | 8.6% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-A-HighTEVp-P4 bc2—bc8 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 225/5803 | 3.9% | Insertion | - |
| N gene | +108 | +A | 154/5805 | 2.7% | Insertion | Frameshift |
| +237 | +T | 276/5806 | 4.8% | Insertion | Frameshift | |
| +636 | +T | 158/5807 | 2.7% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 134/5745 | 2.3% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 536/5803 | 9.2% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-B-HighTEVp-P4 bc2—bc10 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 270/5572 | 4.8% | Insertion | - |
| N gene | +237 | +T | 223/5572 | 4.0% | Insertion | Frameshift |
| +636 | +T | 155/5571 | 2.8% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 590/5576 | 10.6% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-C-HighTEVp-P4 bc2—bc11 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 233/5581 | 4.2% | Insertion | - |
| –21 | -N | 114/5581 | 2.0% | Deletion | - | |
| –19 | A>G | 272/5499 | 4.9% | Substitution | - | |
| N gene | +237 | +T | 252/5582 | 4.5% | Insertion | Frameshift |
| +636 | +T | 149/5581 | 2.7% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 248/5528 | 4.5% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 573/5579 | 10.3% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-D-HighTEVp-P4 bc2—bc12 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 200/6116 | 3.3% | Insertion | - |
| N gene | +237 | +T | 219/6117 | 3.6% | Insertion | Frameshift |
| +636 | +T | 160/6119 | 2.6% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 456/6120 | 7.5% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-A-LowTEVp-P4 bc1—bc10 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 239/4681 | 5.1% | Insertion | - |
| N gene | +108 | +A | 114/4682 | 2.4% | Insertion | Frameshift |
| +237 | +T | 242/4683 | 5.2% | Insertion | Frameshift | |
| +636 | +T | 131/4684 | 2.8% | Insertion | Frameshift | |
| +1,053 | +A | 97/4683 | 2.1% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 170/4650 | 3.7% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 570/4683 | 12.2% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-B-LowTEVp-P4 bc1—bc11 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 255/4757 | 5.4% | Insertion | - |
| N gene | +237 | +T | 245/4758 | 5.1% | Insertion | Frameshift |
| +636 | +T | 141/4758 | 3.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 551/4757 | 11.6% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-C-LowTEVp-P4 bc1—bc12 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 268/5461 | 4.9% | Insertion | - |
| –19 | A>G | 160/5403 | 3.0% | Substitution | - | |
| N gene | +237 | +T | 231/5463 | 4.2% | Insertion | Frameshift |
| +636 | +T | 156/5466 | 2.9% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 705/5286 | 13.3% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 538/5464 | 9.8% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-D-LowTEVp-P4 bc2—bc3 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 266/5841 | 4.6% | Insertion | - |
| N gene | +237 | +T | 246/5838 | 4.2% | Insertion | Frameshift |
| +574 | -N | 140/5834 | 2.4% | Deletion | Frameshift | |
| +636 | +T | 156/5833 | 2.7% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 200/5737 | 3.5% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 529/5818 | 9.1% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-A-HighTEVp-P6 bc5—bc6 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 604/6567 | 9.2% | Insertion | - |
| –19 | A>G | 555/6349 | 8.7% | Substitution | - | |
| N gene | +108 | +A | 227/6565 | 3.5% | Insertion | Frameshift |
| +166 | +T | 157/6565 | 2.4% | Insertion | Frameshift | |
| +237 | +T | 543/6565 | 8.3% | Insertion | Frameshift | |
| +245 | +G | 132/6565 | 2.0% | Insertion | Frameshift | |
| +466 | +A | 175/6566 | 2.7% | Insertion | Frameshift | |
| +636 | +T | 337/6569 | 5.1% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 767/6317 | 12.1% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 1032/6583 | 15.7% | Insertion | - |
| P | +1,669 | +A | 155/6584 | 2.4% | Insertion | Frameshift |
| SIR-B-HighTEVp-P6 bc5—bc7 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 624/6752 | 9.2% | Insertion | - |
| –21 | -N | 202/6754 | 3.0% | Deletion | - | |
| –20 | +G | 243/6754 | 3.6% | Insertion | - | |
| –19 | A>G | 1180/6296 | 18.7% | Substitution | - | |
| N gene | +108 | +A | 216/6752 | 3.2% | Insertion | Frameshift |
| +166 | +T | 185/6751 | 2.7% | Insertion | Frameshift | |
| +237 | +T | 559/6751 | 8.3% | Insertion | Frameshift | |
| +245 | +G | 138/6751 | 2.0% | Insertion | Frameshift | |
| +466 | +A | 197/6753 | 2.9% | Insertion | Frameshift | |
| +612 | +T | 147/6753 | 2.2% | Insertion | Frameshift | |
| +636 | +T | 330/6753 | 4.9% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 965/6766 | 14.3% | Insertion | - |
| P | +1,669 | +A | 187/6769 | 2.8% | Insertion | Frameshift |
| SIR-C-HighTEVp-P6 bc5—bc8 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 578/6166 | 9.4% | Insertion | - |
| –21 | -N | 205/6166 | 3.3% | Deletion | - | |
| –20 | +G | 298/6166 | 4.8% | Insertion | - | |
| –19 | A>G | 3305/5625 | 58.8% | Substitution | - | |
| N gene | +108 | +A | 179/6166 | 2.9% | Insertion | Frameshift |
| +166 | +T | 171/6165 | 2.8% | Insertion | Frameshift | |
| +237 | +T | 514/6164 | 8.3% | Insertion | Frameshift | |
| +466 | +A | 158/6166 | 2.6% | Insertion | Frameshift | |
| +636 | +T | 318/6170 | 5.2% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 436/5995 | 7.3% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 1019/6184 | 16.5% | Insertion | - |
| P | +1,669 | +A | 165/6185 | 2.7% | Insertion | Frameshift |
| SIR-D-HighTEVp-P6 bc5—bc9 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 562/6355 | 8.8% | Insertion | - |
| –21 | -N | 228/6356 | 3.6% | Deletion | - | |
| –20 | +G | 314/6356 | 4.9% | Insertion | - | |
| –19 | A>G | 2816/5789 | 48.6% | Substitution | - | |
| -9 | A>T | 139/6104 | 2.3% | Substitution | - | |
| -6 | C>T | 176/6275 | 2.8% | Substitution | - | |
| -5 | C>A | 121/5995 | 2.0% | Substitution | - | |
| N gene | +108 | +A | 175/6357 | 2.8% | Insertion | Frameshift |
| +237 | +T | 474/6358 | 7.5% | Insertion | Frameshift | |
| +245 | +G | 131/6358 | 2.1% | Insertion | Frameshift | |
| +466 | +A | 167/6359 | 2.6% | Insertion | Frameshift | |
| +636 | +T | 316/6360 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 947/6365 | 14.9% | Insertion | - |
| P | +1,669 | +A | 139/6365 | 2.2% | Insertion | Frameshift |
| SIR-A-LowTEVp-P6 bc4—bc5 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 588/6703 | 8.8% | Insertion | - |
| –19 | A>G | 369/6525 | 5.7% | Substitution | - | |
| N gene | +108 | +A | 259/6704 | 3.9% | Insertion | Frameshift |
| +166 | +T | 173/6704 | 2.6% | Insertion | Frameshift | |
| +237 | +T | 584/6703 | 8.7% | Insertion | Frameshift | |
| +246 | +G | 145/6703 | 2.2% | Insertion | Frameshift | |
| +466 | +A | 196/6704 | 2.9% | Insertion | Frameshift | |
| +636 | +T | 366/6705 | 5.5% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 681/6468 | 10.5% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 1035/6711 | 15.4% | Insertion | - |
| P | +1,669 | +A | 161/6711 | 2.4% | Insertion | Frameshift |
| SIR-B-LowTEVp-P6 bc4—bc6 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 550/6112 | 9.0% | Insertion | - |
| –19 | A>G | 317/5985 | 5.3% | Substitution | - | |
| N gene | +108 | +A | 186/6117 | 3.0% | Insertion | Frameshift |
| +166 | +T | 131/6117 | 2.1% | Insertion | Frameshift | |
| +237 | +T | 486/6116 | 7.9% | Insertion | Frameshift | |
| +466 | +A | 148/6118 | 2.4% | Insertion | Frameshift | |
| +612 | +T | 125/6120 | 2.0% | Insertion | Frameshift | |
| +636 | +T | 303/6119 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 360/5983 | 6.0% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 946/6133 | 15.4% | Insertion | - |
| P | +1,669 | +A | 138/6133 | 2.3% | Insertion | Frameshift |
| SIR-C-LowTEVp-P6 bc4—bc7 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 494/5209 | 9.5% | Insertion | - |
| –20 | +G | 123/5209 | 2.4% | Insertion | - | |
| –19 | A>G | 2864/4984 | 5.7% | Substitution | - | |
| N gene | +108 | +A | 167/5210 | 3.2% | Insertion | Frameshift |
| +166 | +T | 136/5210 | 2.6% | Insertion | Frameshift | |
| +237 | +T | 400/5210 | 7.7% | Insertion | Frameshift | |
| +245 | +G | 123/5210 | 2.4% | Insertion | Frameshift | |
| +466 | +A | 146/5213 | 2.8% | Insertion | Frameshift | |
| +636 | +T | 261/5214 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 546/5066 | 10.8% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 816/5212 | 15.7% | Insertion | - |
| P | +1,669 | +A | 120/5212 | 2.3% | Insertion | Frameshift |
| SIR-D-LowTEVp-P6 bc4—bc7 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 492/5279 | 9.3% | Insertion | - |
| –21 | -N | 114/5279 | 2.2% | Deletion | - | |
| –20 | +G | 119/5279 | 2.3% | Insertion | - | |
| –19 | A>G | 1553/5049 | 30.8% | Substitution | - | |
| -9 | A>T | 104/5189 | 2.0% | Substitution | - | |
| N gene | +108 | +A | 163/5279 | 3.1% | Insertion | Frameshift |
| +166 | +T | 129/5279 | 2.4% | Insertion | Frameshift | |
| +237 | +T | 434/5279 | 8.2% | Insertion | Frameshift | |
| +245 | +G | 106/5279 | 2.0% | Insertion | Frameshift | |
| +466 | +A | 148/5281 | 2.8% | Insertion | Frameshift | |
| +612 | +T | 120/5281 | 2.3% | Insertion | Frameshift | |
| +636 | +T | 279/5281 | 5.3% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | - | - | - | - | - |
| Intergenic | +1,564 | +A | 831/5281 | 15.7% | Insertion | - |
| P | +1,669 | +A | 123/5281 | 2.3% | Insertion | Frameshift |
| SIR-A-HighTEVp-P8 bc6—bc7 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 541/6868 | 7.9% | Insertion | - |
| –21 | -N | 299/6868 | 4.4% | Deletion | - | |
| –20 | +G | 431/6868 | 6.3% | Insertion | - | |
| –19 | A>G | 3684/6150 | 60.0% | Substitution | - | |
| N gene | +108 | +A | 198/6867 | 2.9% | Insertion | Frameshift |
| +166 | +T | 157/6867 | 2.3% | Insertion | Frameshift | |
| +237 | +T | 583/6867 | 8.5% | Insertion | Frameshift | |
| +245 | +G | 138/6867 | 2.0% | Insertion | Frameshift | |
| +466 | +A | 181/6868 | 2.6% | Insertion | Frameshift | |
| +636 | +T | 342/6870 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 651/6620 | 9.8% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 952/6896 | 13.8% | Insertion | - |
| P | +1,669 | +A | 144/6898 | 2.1% | Insertion | Frameshift |
| SIR-B-HighTEVp-P8 bc6—bc8 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 571/6246 | 9.1% | Insertion | - |
| –21 | -N | 182/6246 | 2.9% | Deletion | - | |
| –20 | +G | 319/6246 | 5.1% | Insertion | - | |
| –19 | A>G | 3836/5763 | 66.6% | Substitution | - | |
| –18 | A>C | 171/5940 | 2.9% | Substitution | - | |
| N gene | +108 | +A | 197/6247 | 3.2% | Insertion | Frameshift |
| +166 | +T | 167/6247 | 2.7% | Insertion | Frameshift | |
| +237 | +T | 486/6247 | 7.8% | Insertion | Frameshift | |
| +245 | +G | 145/6248 | 2.3% | Insertion | Frameshift | |
| +466 | +A | 149/6249 | 2.4% | Insertion | Frameshift | |
| +636 | +T | 323/6251 | 5.2% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 365/6068 | 6.0% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 927/6259 | 14.8% | Insertion | - |
| P | +1,669 | +A | 152/6259 | 2.4% | Insertion | Frameshift |
| SIR-C-HighTEVp-P8 bc6—bc9 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 598/6403 | 9.3% | Insertion | - |
| –19 | A>G | 6024/6304 | 95.6% | Substitution | - | |
| N gene | +108 | +A | 200/6404 | 3.1% | Insertion | Frameshift |
| +166 | +T | 146/6404 | 2.3% | Insertion | Frameshift | |
| +237 | +T | 518/6405 | 8.1% | Insertion | Frameshift | |
| +245 | +G | 158/6405 | 2.5% | Insertion | Frameshift | |
| +466 | +A | 172/6406 | 2.7% | Insertion | Frameshift | |
| +636 | +T | 311/6407 | 4.9% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 986/6410 | 15.4% | Insertion | - |
| P | +1,669 | +A | 139/6408 | 2.2% | Insertion | Frameshift |
| SIR-D-HighTEVp-P8 bc6—bc10 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 482/5760 | 8.4% | Insertion | - |
| –19 | A>G | 5092/5625 | 9.1% | Substitution | - | |
| –18 | A>G | 155/5609 | 2.8% | Substitution | - | |
| -9 | A>T | 247/5402 | 4.6% | Substitution | - | |
| -9 | A>G | 449/5402 | 8.3% | Substitution | - | |
| -9 | +G | 120/5761 | 2.1% | Insertion | - | |
| -6 | C>T | 680/5586 | 12.2% | Substitution | - | |
| -6 | +T | 167/5761 | 2.9% | Insertion | - | |
| -5 | C>A | 153/5412 | 2.8% | Substitution | - | |
| N gene | +108 | +A | 163/5763 | 2.8% | Insertion | Frameshift |
| +166 | +T | 119/5763 | 2.1% | Insertion | Frameshift | |
| +237 | +T | 414/5763 | 7.2% | Insertion | Frameshift | |
| +466 | +A | 119/5764 | 2.1% | Insertion | Frameshift | |
| +612 | +T | 127/5764 | 2.2% | Insertion | Frameshift | |
| +636 | +T | 291/5764 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 861/5766 | 14.9% | Insertion | - |
| P | +1,669 | +A | 137/5766 | 2.4% | Insertion | Frameshift |
| SIR-A-LowTEVp-P8 bc4—bc9 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 646/7058 | 9.2% | Insertion | - |
| –21 | -N | 252/7059 | 3.6% | Deletion | - | |
| –20 | +G | 417/7059 | 5.9% | Insertion | - | |
| –19 | A>G | 2752/6358 | 43.3% | Substitution | - | |
| -6 | C>T | 171/6942 | 2.5% | Substitution | - | |
| -5 | C>A | 542/6530 | 8.3% | Substitution | - | |
| N gene | +108 | +A | 346/7058 | 4.9% | Insertion | Frameshift |
| +166 | +T | 178/7058 | 2.5% | Insertion | Frameshift | |
| +237 | +T | 622/7058 | 8.8% | Insertion | Frameshift | |
| +245 | +G | 161/7058 | 2.3% | Insertion | Frameshift | |
| +466 | +A | 194/7059 | 2.7% | Insertion | Frameshift | |
| +612 | +T | 150/7060 | 2.1% | Insertion | Frameshift | |
| +636 | +T | 345/7060 | 4.9% | Insertion | Frameshift | |
| +795 | T>C | 1604/6265 | 25.6% | Substitution | Silent F265 | |
| +795 | +C | 318/7061 | 4.5% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 1122/6684 | 16.8% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 1079/7085 | 15.2% | Insertion | - |
| P | +1,669 | +A | 161/7090 | 2.3% | Insertion | Frameshift |
| SIR-B-LowTEVp-P8 bc4—bc10 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 647/6759 | 9.6% | Insertion | - |
| –21 | -N | 242/6761 | 3.6% | Deletion | - | |
| –20 | +G | 371/6761 | 5.5% | Insertion | - | |
| –19 | A>G | 2200/6168 | 35.7% | Substitution | - | |
| –18 | A>C | 400/6309 | 6.3% | Substitution | - | |
| N gene | +108 | +A | 224/6761 | 3.3% | Insertion | Frameshift |
| +166 | +T | 157/6761 | 2.3% | Insertion | Frameshift | |
| +237 | +T | 575/6760 | 8.5% | Insertion | Frameshift | |
| +466 | +A | 189/6764 | 2.8% | Insertion | Frameshift | |
| +636 | +T | 353/6763 | 5.2% | Insertion | Frameshift | |
| +1,349 | C>A | 144/6671 | 2.2% | Substitution | Missense S450X | |
| TEVs-PEST | +1,357 | G>T | 1192/6372 | 18.7% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 1026/6769 | 15.2% | Insertion | - |
| P | +1,669 | +A | 173/6772 | 2.6% | Insertion | Frameshift |
| SIR-C-LowTEVp-P8 bc4—bc11 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 614/6893 | 8.9% | Insertion | - |
| –20 | +G | 261/6893 | 3.8% | Insertion | - | |
| –19 | A>G | 5317/6466 | 82.2% | Substitution | - | |
| N gene | +108 | +A | 215/6894 | 3.1% | Insertion | Frameshift |
| +237 | +T | 564/6894 | 8.2% | Insertion | Frameshift | |
| +466 | +A | 207/6895 | 3.0% | Insertion | Frameshift | |
| +636 | +T | 364/6895 | 5.3% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 1013/6551 | 15.5% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 1053/6920 | 15.2% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-D-LowTEVp-P8 bc4—bc12 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 541/5872 | 9.2% | Insertion | - |
| –20 | +G | 190/5872 | 3.2% | Insertion | - | |
| –19 | A>G | 4259/5565 | 76.5% | Substitution | - | |
| -9 | A>T | 141/5738 | 2.5% | Substitution | - | |
| N gene | +108 | +A | 168/5876 | 2.9% | Insertion | Frameshift |
| +166 | +T | 154/5876 | 2.6% | Insertion | Frameshift | |
| +237 | +T | 491/5876 | 8.4% | Insertion | Frameshift | |
| +245 | +G | 133/5876 | 2.3% | Insertion | Frameshift | |
| +332 | +A | 123/5876 | 2.1% | Insertion | Frameshift | |
| +466 | +A | 152/5876 | 2.6% | Insertion | Frameshift | |
| +612 | +T | 134/5876 | 2.3% | Insertion | Frameshift | |
| +636 | +T | 324/5876 | 5.5% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 521/5707 | 9.1% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 996/5881 | 17.0% | Insertion | - |
| P | +1,669 | +A | 150/5882 | 2.6% | Insertion | Frameshift |
Table 3
List of detected mutations above 2% threshold in purified SiR viruses recovered from injected hippocampi sequenced by SMRT NGS sequencing.
The position of the mutations is defined considering +1 the first base of the nucleoprotein N coding sequence.
| NGS sequencing results of purified viruses used in vivo | ||||||
|---|---|---|---|---|---|---|
| SIR-CRE purified bc3—bc5 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 238/5196 | 4.6% | Insertion | - |
| N gene | +237 | +T | 199/5196 | 3.8% | Insertion | Frameshift |
| +636 | +T | 150/5200 | 2.9% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 544/5205 | 10.5% | Insertion | - |
| P | - | - | - | - | - | - |
| SIR-CRE purified, 1 week p.i. in vivo (A) bc5—bc10 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 474/5211 | 9.1% | Insertion | - |
| –21 | +A | 110/5211 | 2.1% | Insertion | - | |
| N gene | +108 | +A | 176/5211 | 3.4% | Insertion | Frameshift |
| +166 | +T | 132/5211 | 2.5% | Insertion | Frameshift | |
| +237 | +T | 389/5211 | 7.5% | Insertion | Frameshift | |
| +245 | +G | 108/5211 | 2.1% | Insertion | Frameshift | |
| +466 | +A | 135/5211 | 2.6% | Insertion | Frameshift | |
| +612 | +T | 108/5210 | 2.1% | Insertion | Frameshift | |
| +636 | +T | 288/5210 | 5.5% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 773/5213 | 14.8% | Insertion | - |
| P | +1,669 | +A | 128/5213 | 2.5% | Insertion | Frameshift |
| SIR-CRE purified, 1 week p.i. in vivo (B) bc5—bc11 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 482/5542 | 8.7% | Insertion | - |
| N gene | +108 | +A | 157/5543 | 2.8% | Insertion | Frameshift |
| +166 | +T | 125/5543 | 2.3% | Insertion | Frameshift | |
| +237 | +T | 402/5543 | 7.3% | Insertion | Frameshift | |
| +245 | +G | 123/5543 | 2.2% | Insertion | Frameshift | |
| +466 | +A | 157/5543 | 2.8% | Insertion | Frameshift | |
| +612 | +T | 112/5543 | 2.0% | Insertion | Frameshift | |
| +636 | +T | 276/5543 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 744/5542 | 13.4% | Insertion | - |
| P | +1,669 | +A | 144/5542 | 2.6% | Insertion | Frameshift |
| SIR-CRE purified, 1 week p.i. in vivo (C) bc5—bc12 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 481/5150 | 9.3% | Insertion | - |
| N gene | +108 | +A | 137/5150 | 2.7% | Insertion | Frameshift |
| +166 | +T | 118/5150 | 2.3% | Insertion | Frameshift | |
| +237 | +T | 390/5150 | 7.6% | Insertion | Frameshift | |
| +245 | +G | 104/5150 | 2.0% | Insertion | Frameshift | |
| +466 | +A | 140/5150 | 2.7% | Insertion | Frameshift | |
| +612 | +T | 116/5150 | 2.3% | Insertion | Frameshift | |
| +636 | +T | 255/5150 | 5.0% | Insertion | Frameshift | |
| TEVs-PEST | - | - | - | - | - | - |
| Intergenic | +1,564 | +A | 739/5148 | 14.4% | Insertion | - |
| P | +1,669 | +A | 130/5148 | 2.5% | Insertion | Frameshift |
| SIR-G453X-CRE purified bc3—bc11 | ||||||
| Position | Variant | N (q>20) | Freq % | Mutation | Effect on CDS | |
| Upstream N | –49 | +A | 211/4886 | 4.3% | Insertion | - |
| N gene | +237 | +T | 244/4890 | 5.0% | Insertion | Frameshift |
| +636 | +T | 138/4911 | 2.8% | Insertion | Frameshift | |
| TEVs-PEST | +1,357 | G>T | 4780/4912 | 97.3% | Substitution | Missense G453X |
| Intergenic | +1,564 | +A | 502/4924 | 10.2% | Insertion | - |
| P | - | - | - | - | - | - |
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (mouse Rosa26LSL-tdTomato) | B6.CgGt(ROSA)26Sortm14(CAG-tdTomato)Hze/J | Jackson Labs (H.Zeng) | 007914 | |
| Cell line (Homo-sapiens) | HEK293T | ATTC | CRL-3216 | |
| Cell line (Homo-sapiens) | HEK-GG | This paper | See Methods. | |
| Cell line (Homo-sapiens) | HEK-TGG | This paper | See Methods. | |
| Cell line (Homo-sapiens) | HEK-TEVp | This paper | See Methods. | |
| Recombinant DNA reagent (plasmid) | pLenti-puro-2A-TEV | Ciabatti et al., 2017 | Addgene: 99610 | |
| Recombinant DNA reagent (plasmid) | pLenti-H2BGFP-2A-GlySAD | This paper | See Methods. | |
| Recombinant DNA reagent (plasmid) | pSiR-CRE | This Paper | Derived from Addgene: 99608. See Methods. | |
| Recombinant DNA reagent (plasmid) | pSiR-S450X-nucGFP | This Paper | Derived from Addgene: 99608. See Methods. | |
| Recombinant DNA reagent (plasmid) | pSiR-G453X-nucGFP | This Paper | Derived from Addgene: 99608. See Methods. | |
| Recombinant DNA reagent (plasmid) | pSiR-G453X-CRE | This Paper | Derived from Addgene: 99608. See Methods. | |
| Recombinant DNA reagent (plasmid) | pSiR-N2c-CRE | Lee et al., 2023 | Addgene: 194456 | |
| Recombinant DNA reagent (plasmid) | pΔG-Rabies-CRE | This paper | See Methods. | |
| Recombinant DNA reagent (plasmid) | pAAV-CMV-nucGFP-2A-TVA (AAV-TVA) | This paper | See Methods. | |
| Recombinant DNA reagent (plasmid) | pAAV-hSyn1-TVAmCherry-2A-G(N2c) (AAV-TVA-G_N2c) | Lee et al., 2023 | Addgene: 194354 | |
| Recombinant DNA reagent (plasmid) | pAAV-hSyn1-TVAmCherry-2A-oG(AAV-TVA-G) | This paper | Derived from Addgene: 194354. See Methods. | |
| Antibody | Anti-V5 tag antibody (mouse monoclonal) | Sigma Aldrich | V8012 | 1:5000 dilution |
| Antibody | anti-Mouse IgG (H+L) HRP-conjugated (goat polyclonal) | Invitrogen | 32430 | 1:2000 dilution |
| Sequence-based reagent | qPCR assay against Actb gene (HEX-conjugated) | IDT | Mm.PT.39a.22214843.g | |
| Sequence-based reagent | qPCR assay against Rabies N gene (6-FAM-conjugated) | IDT | FW: CAGGTTCTCTGGTGGAGATAAA Probe: TGACAGGAGGCATGGAACTGACAA RV: CTCAAGAGAAGACCGACTAAGG |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Genomic stability of self-inactivating rabies
eLife 12:e83459.
https://doi.org/10.7554/eLife.83459




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}