Role of D-aminoacyl-tRNA deacylase beyond chiral proofreading as a cellular defense against glycine mischarging by AlaRS
- CSIR–Centre for Cellular and Molecular Biology, India
Abstract
Strict L-chiral rejection through Gly-cisPro motif during chiral proofreading underlies the inability of D-aminoacyl-tRNA deacylase (DTD) to discriminate between D-amino acids and achiral glycine. The consequent Gly-tRNAGly ‘misediting paradox’ is resolved by EF-Tu in the cell. Here, we show that DTD’s active site architecture can efficiently edit mischarged Gly-tRNAAla species four orders of magnitude more efficiently than even AlaRS, the only ubiquitous cellular checkpoint known for clearing the error. Also, DTD knockout in AlaRS editing-defective background causes pronounced toxicity in Escherichia coli even at low-glycine levels which is alleviated by alanine supplementation. We further demonstrate that DTD positively selects the universally invariant tRNAAla-specific G3•U70. Moreover, DTD’s activity on non-cognate Gly-tRNAAla is conserved across all bacteria and eukaryotes, suggesting DTD’s key cellular role as a glycine deacylator. Our study thus reveals a hitherto unknown function of DTD in cracking the universal mechanistic dilemma encountered by AlaRS, and its physiological importance.
https://doi.org/10.7554/eLife.24001.001eLife digest
Proteins are made up of many different building blocks called amino acids, which are linked together in chains. The exact order of amino acids in a protein chain is important for the protein to work properly. When a cell makes proteins, molecules known as transfer ribonucleic acids (or tRNAs for short) bind to specific amino acids to guide them to the growing protein chains in the correct order.
Most amino acids – except one called glycine – have two forms that are mirror images of one another, known as left-handed (L-amino acids) and right-handed (D-amino acids). However, only L-amino acids and glycine are used to make proteins. This is because of the presence of multiple quality control checkpoints in the cell that prevent D-amino acids from being involved. One such checkpoint is an enzyme called D-amino acid deacylase (DTD), which removes D-amino acids that are attached to tRNAs.
Other enzymes are responsible for linking a particular amino acid to its correct tRNA. Along with mistaking D-amino acids for L-amino acids, these enzymes can also make errors when they have to distinguish between amino acids that are similar in shape and size. For example, the enzyme that attaches L-alanine to its tRNA can also mistakenly attach larger L-serine or smaller glycine to it instead. Previous research has shown that attaching L-serine to this tRNA can lead to neurodegeneration in mice, whereas attaching glycine does not seem to cause any harm. It is not clear why this is the case.
Pawar et al. investigated how incorrectly attaching glycine or L-serine to the tRNA that usually binds to L-alanine affects a bacterium called Escherichia coli. The experiments show that, if the mistake is not corrected, glycine can be just as harmful to the cells as L-serine. The reason that glycine appears to be less of a problem is that the DTD enzyme is able to remove glycine, but not L-serine, from the tRNA. Further experiments show that DTD can play a similar role in a variety of organisms from bacteria to mammals.
The findings of Pawar et al. extend the role of DTD beyond preventing D-amino acids from being incorporated into proteins. The next step is to understand the role of this enzyme in humans and other multicellular organisms, especially in the context of nerve cells, where it is present at high levels.
https://doi.org/10.7554/eLife.24001.002Introduction
D-aminoacyl-tRNA deacylase (DTD) is a key factor that keeps chiral errors away from the translational machinery by allowing only L-amino acids to form proteins and has therefore been implicated in perpetuation of homochirality in the protein world (Calendar and Berg, 1967; Soutourina et al., 1999, 2000). The design principle by which this remarkable configurational specificity is achieved by DTD involves only strict L-chiral rejection, rather than D-chiral selection. An invariant cross-subunit Gly-cisPro motif forms the structural and mechanistic basis for DTD’s enantioselection (Ahmad et al., 2013). Thus, the architecture of DTD’s chiral proofreading site is such that it cannot prevent misediting of achiral glycine charged on tRNAGly and seems to have an inherent flaw. The glycine ‘misediting paradox’ is, however, effectively resolved through protection of the cognate achiral substrate by elongation factor thermo unstable (EF-Tu) (Routh et al., 2016).
While occasional chiral errors that occur during aminoacylation are cleared by DTD to ensure the accuracy of aminoacyl-tRNAs present in the cellular pool, a major role is played by editing functions associated with about half of the 20 aminoacyl-tRNA synthetases (aaRSs) to rectify the incorrect pairing of a similar non-cognate L-amino acid with a tRNA (Guo and Schimmel, 2012; Ibba and Soll, 2000). These aaRSs can edit the non-cognate amino acid at the aminoacylation site itself after the amino acid has been activated (i.e. formation of aminoacyl-AMP using ATP as a substrate) but prior to its transfer to the tRNA (pre-transfer editing). Alternatively, proofreading can happen at a distinct editing site after the activated non-cognate amino acid has been esterified with the tRNA (post-transfer editing). These proofreading processes are so crucial that even mild defects can lead to adverse cellular outcomes like cell growth retardation, neurodegeneration, cardiomyopathy and cell death (Bacher et al., 2005; Bullwinkle et al., 2014; Karkhanis et al., 2007; Korencic et al., 2004; Lee et al., 2006; Liu et al., 2014; Lu et al., 2014; Moghal et al., 2016; Nangle et al., 2002; Roy et al., 2004), although a compromise in editing can also be beneficial as it helps the organism to tide over stress conditions (Moghal et al., 2014).
Being comparatively small and similar to the cognate alanine, glycine and serine are misactivated by alanyl-tRNA synthetase (AlaRS) at significantly high frequencies of 1/240 and 1/500, respectively, relative to alanine (Tsui and Fersht, 1981) (Figure 1). However, these misactivation rates are much higher than the overall error rates of ~10−4–10−3 observed during protein biosynthesis (Ogle and Ramakrishnan, 2005). Once (mis)activated, these non-cognate amino acids are mischarged on tRNAAla by AlaRS. This creates a unique mechanistic challenge for the editing domain of AlaRS, which has to specifically remove two non-cognate amino acids—the larger serine and the smaller glycine—attached to tRNAAla without acting on the cognate alanine, which is intermediate in size between serine and glycine. This 3.5-billion-year-old double-discrimination problem is shown to be unavoidable for AlaRS in all forms of life (Guo et al., 2009). It has also been shown that serine mischarging on tRNAAla is detrimental to the cell and even a mild deficiency in the proofreading activity of AlaRS leads to cell death and severe neuropathologies in mouse (Lee et al., 2006; Liu et al., 2014). The problem is so severe that several standalone trans-editing modules (collectively called AlaXs), which are homologous to AlaRS cis-editing domain, have come into being. However, these trans-editing factors are not ubiquitously present; their distribution is more in archaea than in eukaryotes and bacteria (Guo and Schimmel, 2012). Surprisingly, only archaeal AlaXs are known to clear both Ser-tRNAAla and Gly-tRNAAla (Ahel et al., 2003); eukaryotic AlaXs have been shown to act as cellular redundancies to edit only Ser-tRNAAla (Guo et al., 2009), whereas biochemical activity of bacterial AlaXs is yet to be probed (Figure 1). These findings corroborated the notion that only serine mischarging by AlaRS poses the major threat to the cell (Guo et al., 2009; Lee et al., 2006; Liu et al., 2014).
Figure 1
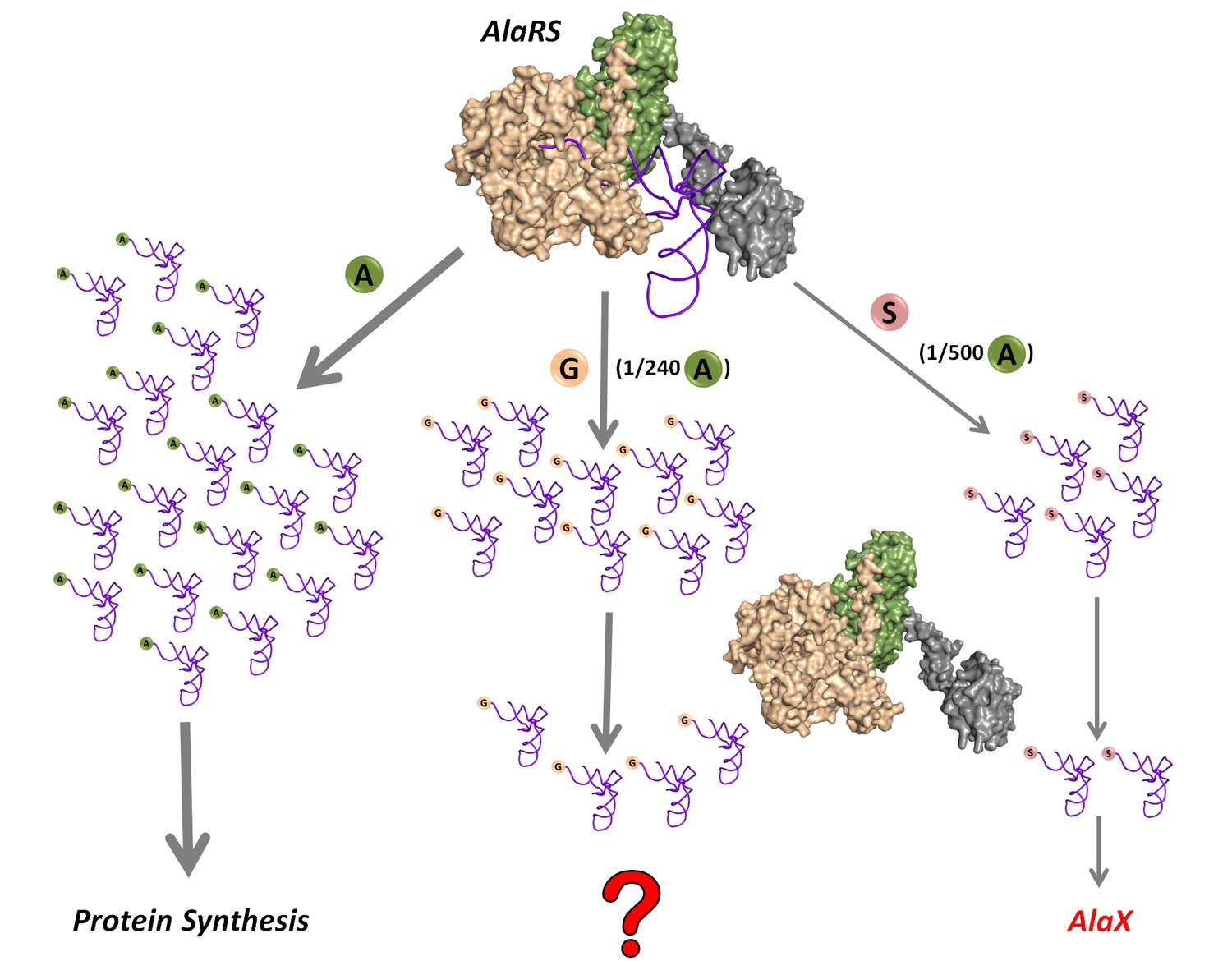
Mischarging by AlaRS.
AlaRS activates and charges alanine (A) to form cognate Ala-tRNAAla which is routed for protein synthesis. In this process, AlaRS also misactivates glycine (G) and serine (S) at frequencies of 1 per 240 alanine and 1 per 500 alanine, respectively (Tsui and Fersht, 1981). The two non-cognate amino acids are then charged on tRNAAla to produce Gly-tRNAAla and Ser-tRNAAla species, with glycine mischarging being nearly twice that of serine. Since AlaRS does not distinguish much between Gly-tRNAAla and Ser-tRNAAla while clearing the two, higher levels of Gly-tRNAAla might accumulate in the cell. However, there are additional free-standing trans-editing factors called AlaX (found in all domains of life but not in all organisms), which are known to edit mainly Ser-tRNAAla. This leads to a fundamental question as to how the problem of Gly-tRNAAla editing is solved in the cellular context.
In the current study, we show that there is significant glycine mischarging by AlaRS in the presence of EF-Tu which can be equally pernicious as serine mischarging. We demonstrate that DTD plays an active and crucial role in preventing the accumulation of mischarged Gly-tRNAAla species. A cell lacking DTD in AlaRS editing-defective background displays pronounced toxicity toward even low levels of glycine which is, nevertheless, alleviated by alanine supplementation. Our data also indicate that DTD has selectivity for the G3•U70 wobble base pair that is unique to tRNAAla, suggesting that in the primordial scenario, DTD could have been recruited primarily as a glycine-removing factor. Our study thus brings to the fore three important aspects of translational fidelity, which were underappreciated or unknown so far. Firstly, glycine, like serine, can be toxic and deleterious to the cell under conditions wherein the cell is deficient in disposing of the mischarged Gly-tRNAAla species. Secondly, how the design of the active site of DTD, notwithstanding its unwarranted activity on Gly-tRNAGly, is used to efficiently decouple glycine mischarged on tRNAAla despite the presence of EF-Tu, thereby fortifying translational fidelity. Thirdly, there is a positive selection of the element(s) of tRNAAla by DTD, indicating for the first time the role of tRNA elements in modulating DTD’s activity.
Results
Mischarging by AlaRS leads to significant accumulation of Gly-tRNAAla
To test whether Gly-tRNAAla is accumulated due to mischarging of glycine on tRNAAla by AlaRS, we performed aminoacylation assays in the presence of EF-Tu. In comparison to alanine charging, significant glycine mischarging was observed. Furthermore, the level of glycine mischarging was about twice that of serine mischarging (Figure 2a). This clearly indicated that even with full AlaRS editing potential, there can be significant accumulation of Gly-tRNAAla species in the cell. Moreover, this is in accordance with the twofold higher misactivation rate of glycine by AlaRS when compared to that of serine (Tsui and Fersht, 1981). We then checked the accumulation of Gly-tRNAAla when AlaRS editing was compromised, and it was found to be significantly high, almost equal to the level of Ala-tRNAAla formation (Figure 2—figure supplement 1). To accomplish a compromise in the proofreading activity of AlaRS, a known editing site mutation (viz., C666A) in AlaRS from Escherichia coli was used (Beebe et al., 2003). The above data lead to a couple of fundamental questions: (a) why does a defect in the same editing domain that edits both serine and glycine from tRNAAla cause toxicity only due to serine? and (b) how does the cell tackle the problem of glycine mischarging? Furthermore, based on structural considerations and evolutionary substitution patterns of alanine, where it is replaced more by serine than by glycine (Betts et al., 2003), it is to be expected that substitution of glycine for alanine is more detrimental than substitution of serine.
Figure 2 with 1 supplement see all
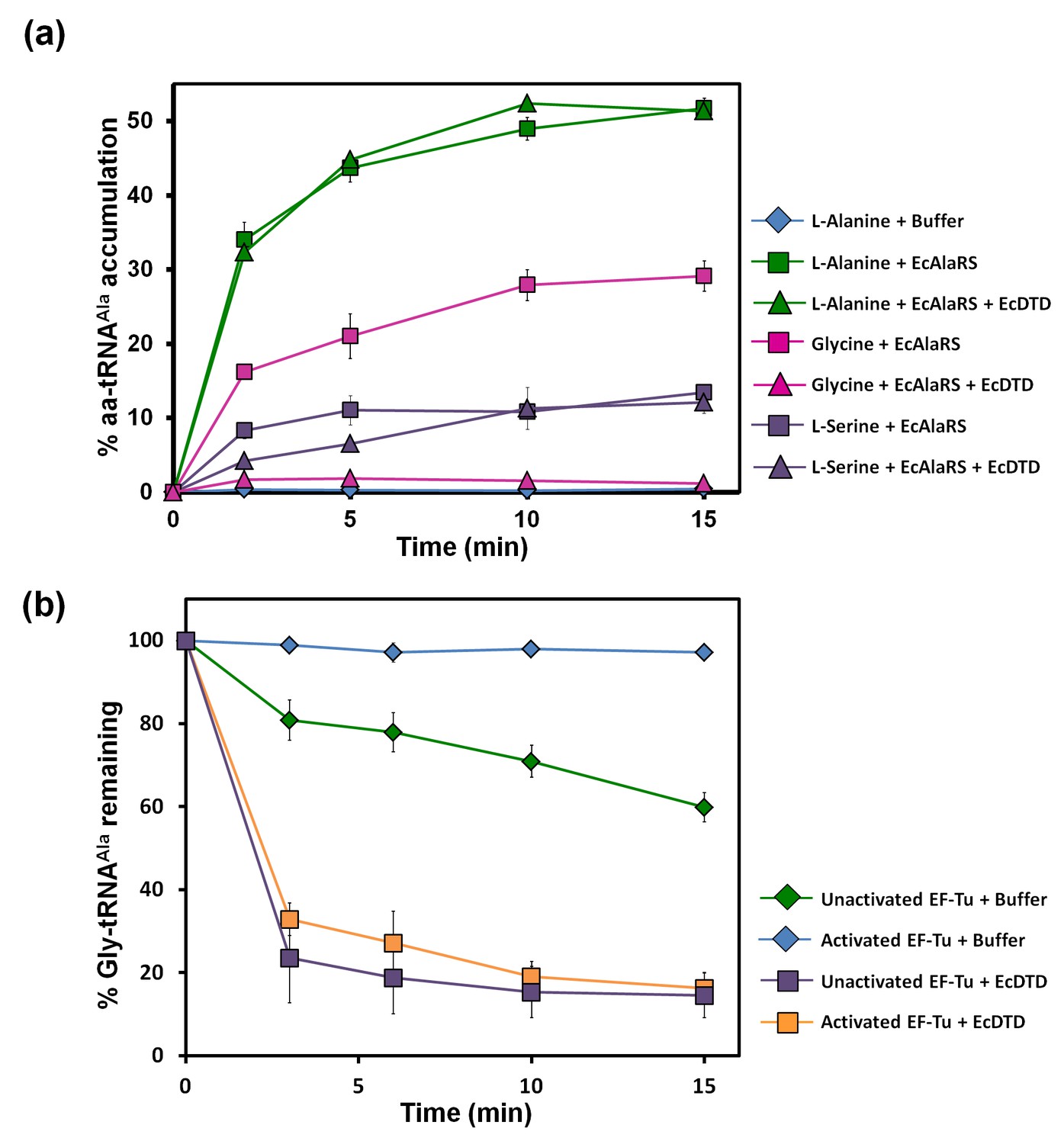
Misacylation of tRNAAla with glycine by AlaRS and its prevention/rectification by DTD.
(a) Aminoacylation of tRNAAla by EcAlaRS in the presence of activated EF-Tu: L-alanine (green square), L-alanine and 10 pM EcDTD (green triangle), glycine (pink square), glycine and 10 pM EcDTD (pink triangle), L-serine (purple square), L-serine and 10 pM EcDTD (purple triangle). No enzyme control (blue diamonds) reaction had all the components of the reaction (with L-alanine) except for EcAlaRS. (b) Deacylation of Gly-tRNAAla in the presence of unactivated EF-Tu (green diamond), activated EF-Tu (blue diamond), 5 pM EcDTD and unactivated EF-Tu (purple square), 5 pM EcDTD and activated EF-Tu (orange square). Error bars indicate one standard deviation from the mean of triplicate readings.
-
Figure 2—source data 1
Misacylation of tRNAAla and deacylation of Gly-tRNAAla in the presence of EF-Tu.
- https://doi.org/10.7554/eLife.24001.005
DTD effectively decouples glycine mischarged on tRNAAla
The leads for the solution to this puzzle came when, surprisingly, we found that the activity of DTD on Gly-tRNAAla was ~1000-fold more than that on Gly-tRNAGly (as discussed later). Moreover, although the ratio of activated EF-Tu to DTD in our assays (viz., ~200 nM to 5 or 10 pM) was much higher than the cellular ratio (viz., ~200:1) (Li et al., 2014), DTD could easily edit Gly-tRNAAla in the presence of EF-Tu (Figure 2b). This was unlike the case of Gly-tRNAGly, wherein EF-Tu showed significant protection of the cognate achiral substrate from DTD (Routh et al., 2016). The deacylation of Gly-tRNAAla by DTD was so striking that ~20,000 times more AlaRS (which is the only universally occurring editing factor for Gly-tRNAAla) as compared to DTD was required for similar kind of deacylation under identical conditions (Figures 2b and 3b). In addition, unlike DTD, AlaRS showed a significant decrease in deacylation activity on Gly-tRNAAla when tested in the presence of EF-Tu (Figure 3a,b). Moreover, our assays demonstrated that DTD is not only efficient in eliminating Gly-tRNAAla despite the presence of EF-Tu (Figure 2b) but can also very effectively prevent the accumulation of Gly-tRNAAla during aminoacylation by AlaRS in the presence of EF-Tu (Figure 2a).
Figure 3

DTD has higher activity than AlaRS for the editing of Gly-tRNAAla.
(a) Deacylation of Gly-tRNAAla in the presence of unactivated EF-Tu: buffer (blue diamond), 10 nM EcAlaRS (red circle), 50 nM EcAlaRS (green circle), 100 nM EcAlaRS (purple circle), 500 nM EcAlaRS (pink circle). (b) Deacylation of Gly-tRNAAla in the presence of activated EF-Tu: buffer (blue diamond), 10 nM EcAlaRS (red square), 50 nM EcAlaRS (green square), 100 nM EcAlaRS (purple square), 500 nM EcAlaRS (pink square). (c) Deacylation of Gly-tRNAAla by EcDTD and increasing concentration of EcAlaRS: buffer (blue diamond), 10 pM EcDTD (orange diamond), 10 pM EcDTD and 10 nM EcAlaRS (red triangle), 10 pM EcDTD and 50 nM EcAlaRS (green triangle), 10 pM EcDTD and 100 nM EcAlaRS (purple triangle), 10 pM EcDTD and 500 nM EcAlaRS (pink triangle). Error bars indicate one standard deviation from the mean of triplicate readings.
-
Figure 3—source data 1
Deacylation of Gly-tRNAAla in the presence of unactivated and activated EF-Tu.
- https://doi.org/10.7554/eLife.24001.008
DTD has significantly higher activity than AlaRS for clearing mischarged Gly-tRNAAla
Considering the high activity of DTD on Gly-tRNAAla, we probed the relative efficiencies of DTD and AlaRS in editing Gly-tRNAAla. To this end, we performed competition assays involving AlaRS, DTD and EF-Tu. In both aminoacylation and deacylation conditions, we found that DTD deacylated Gly-tRNAAla even in the presence of EF-Tu and AlaRS at just 10 pM concentration of DTD (Figures 2a and 3c). Considering the cellular ratio of DTD to AlaRS (viz, ~1:5) (Li et al., 2014) and their relative activities on Gly-tRNAAla, it is evident that when DTD is present, it will eliminate Gly-tRNAAla more efficiently than AlaRS if the non-cognate achiral substrate is released in solution from the synthetase. In this context, it is important to note that compared to Class I synthetases, enzymes belonging to Class II, which include AlaRS, have been shown to have faster product release rates (Zhang et al., 2006). Hence, Class II aaRSs require resampling of the released mischarged product to edit the cytosolic pool of mischarged tRNAs (Ling et al., 2009). This makes even more sense as regards AlaRS, since our structural analysis of AlaRS in complex with tRNAAla (PDB id: 3WQY) suggests that it would be very difficult for the CCA-arm at the 3′ end of tRNAAla harboring the non-cognate amino acid to flip from the aminoacylation site to the editing site without undergoing major conformational changes (Naganuma et al., 2014). Such a dynamics would naturally facilitate a faster release of the misacylated product in solution, implying that a significant fraction of Gly-tRNAAla and Ser-tRNAAla is released from AlaRS prior to their recapture for proofreading by the cis-editing domain. Moreover, our own data, in which we observed significant accumulation of Gly-tRNAAla in the presence of EF-Tu (Figure 2a), corroborate the aforementioned aspects of Class II synthetases in general and AlaRS in particular. Overall, the above data suggest that the problem of glycine mischarging by AlaRS would have been so detrimental that a highly efficient factor like DTD was required to be employed for this function in addition to AlaRS editing domain.
Editing-defective AlaRS and DTD knockout in E. coli
Earlier in vivo studies had shown that AlaRS editing defect causes glycine toxicity only at very high levels of glycine supplementation (80 mM) as opposed to serine which causes toxicity at significantly lower levels (2.5 mM) (Beebe et al., 2003). It is worth noting that these studies were carried out in strains harboring DTD, hence explaining the need for supplementation with more glycine to show toxicity. To check if the absence of DTD makes E. coli susceptible to glycine, we generated an E. coli strain in which dtd (the gene encoding DTD) was knocked out in the background of editing-defective AlaRS. To create a strain that was completely devoid of AlaRS editing activity, the genomic copy of AlaRS gene (alaS) was knocked out and a triple-mutant AlaRS (viz., T567F/S587W/C666F) was expressed from a plasmid. The editing site mutations were designed on the basis of a structural (homology) model of E. coli AlaRS cis-editing domain that was generated using the structure of Archaeoglobus fulgidus AlaRS (PDB id: 2ZTG) as a template. This model was then superimposed on Pyrococcus horikoshii AlaX complexed with serine (PDB id: 1WNU) (the best substrate-mimicking complex for AlaRS and AlaX available so far) (Sokabe et al., 2005). Three residues in the proposed editing site (Beebe et al., 2003; Sokabe et al., 2005) were supplanted by bulkier residues to occlude the pocket and prevent substrate binding (Figure 4a,b). The triple-mutant was found to be inactive on both Ser-tRNAAla and Gly-tRNAAla even when the protein concentration was increased to 1500-fold that of wild-type AlaRS (Figure 4c,d). It is worth mentioning here that the previously known editing-defective mutants of AlaRS (C666A and C666A/Q584H) (Beebe et al., 2003), when checked for deacylation activity on both Ser-tRNAAla and Gly-tRNAAla, were found to show significant activity at just 10-fold higher concentration of the enzyme (Figure 4c,d). Thus, to completely abrogate AlaRS editing activity and to see the effect of editing from only DTD, we chose to use AlaRS triple-mutant for our cell-based toxicity studies.
Figure 4
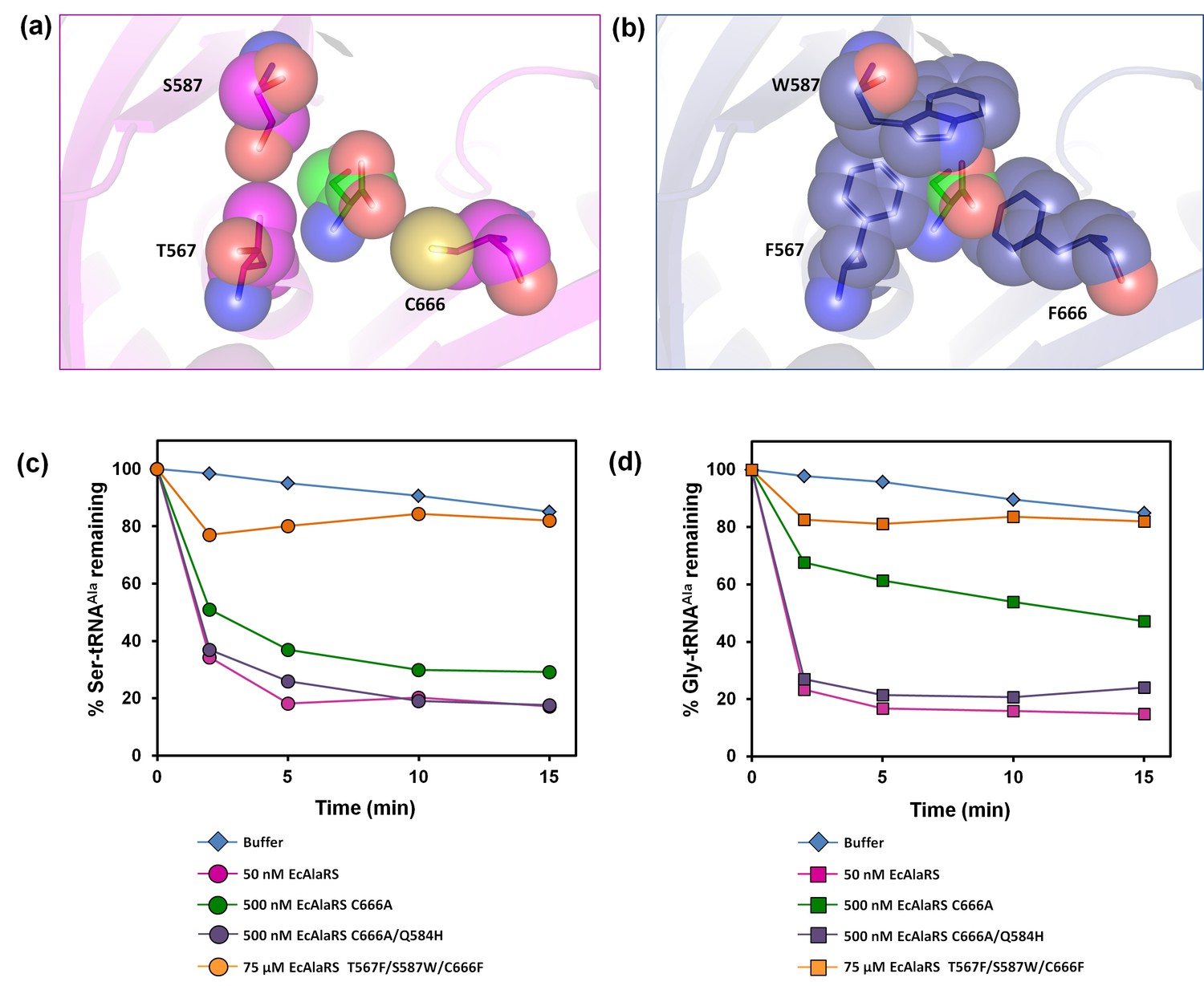
E. coli AlaRS editing site mutants.
Homology model of E. coli AlaRS depicting serine (green sticks/spheres) in the editing site. E. coli AlaRS cis-editing domain was modeled using A. fulgidus AlaRS (PDB id: 2ZTG) as a template, whereas the position and orientation of serine in the model corresponds to that observed in serine-bound P. horikoshii AlaX structure (PDB id: 1WNU). (a) In the wild-type enzyme, residues selected for mutagenesis are represented as megenta sticks/spheres, showing an open pocket for substrate binding. (b) In AlaRS T567F/S587W/C666F, the mutated bulkier residues are depicted as blue sticks/spheres, showing occlusion of the pocket to prevent substrate binding. (c) Deacylation of Ser-tRNAAla by buffer (blue diamond), 50 nM EcAlaRS (pink circle), 500 nM EcAlaRS C666A (green circle), 500 nM EcAlaRS C666A/Q584H (purple circle), 75 µM EcAlaRS T567F/S587W/C666F (orange circle). (d) Deacylation of Gly-tRNAAla by buffer (blue diamond), 50 nM EcAlaRS (pink square), 500 nM EcAlaRS C666A (green square), 500 nM EcAlaRS C666A/Q584H (purple square), 75 µM EcAlaRS T567F/S587W/C666F (orange square).
-
Figure 4—source data 1
Deacylation of Ser-tRNAAla and Gly-tRNAAla by E. coli AlaRS editing site mutants.
- https://doi.org/10.7554/eLife.24001.010
DTD prevents glycine toxicity in E. coli
Cellular toxicity studies using spot dilution assays and growth curve analysis with the DTD knockout strain in the background of AlaRS editing defect showed some toxicity even without any amino acid supplementation, and the toxicity increased with glycine supplementation as low as 3 mM. At 10 mM of glycine supplementation, the cells showed severe growth defect (Figure 5a). To check whether this was specifically due to mischarging caused by AlaRS, toxicity experiments were carried out in the presence of alanine, since the latter is expected to compete for the AlaRS aminoacylation site during charging of tRNAAla. It was observed that alanine supplementation completely recovered the toxicity caused by glycine and rescued the growth completely (Figure 5b,c,d). Moreover, to rule out any non-specific effects due to amino acid supplementation, histidine was used as a negative control, and it was found that it failed to rescue the cells from glycine toxicity (Figure 5c,d). Furthermore, DTD was found to be totally inactive on Ser-tRNAAla in our biochemical assays which confirmed that the toxicity observed in the DTD-lacking cells in the background of AlaRS editing defect was not due to serine mischarging on tRNAAla by AlaRS (Figure 5—figure supplement 1). The above observation is expected since it has been shown earlier that DTD’s chiral proofreading site rejects even L-alanine, the smallest L-chiral substrate (Routh et al., 2016). Taken together, these experiments established that DTD acts as a key cellular factor that edits glycine mischarged on tRNAAla by AlaRS. However, no toxicity of glycine supplementation was observed in E. coli MG1655 ∆dtd strain in the background of wild-type AlaRS (Figure 5—figure supplement 2). This indicates that under normal growth conditions, AlaRS editing is sufficient for the cell to survive. The errors produced because of the absence of DTD are possibly tolerated by E. coli under laboratory conditions, as has been noted in several other cases of editing function of aaRSs (Reynolds et al., 2010a, 2010b). The real implications of editing defects are only recently being appreciated in some specific growth conditions like oxidative stress, oxygen deprivation, starvation/nutrient limiting conditions etc. (Bullwinkle et al., 2014; Cvetesic et al., 2014; Kermgard et al., 2017).
Figure 5 with 2 supplements see all
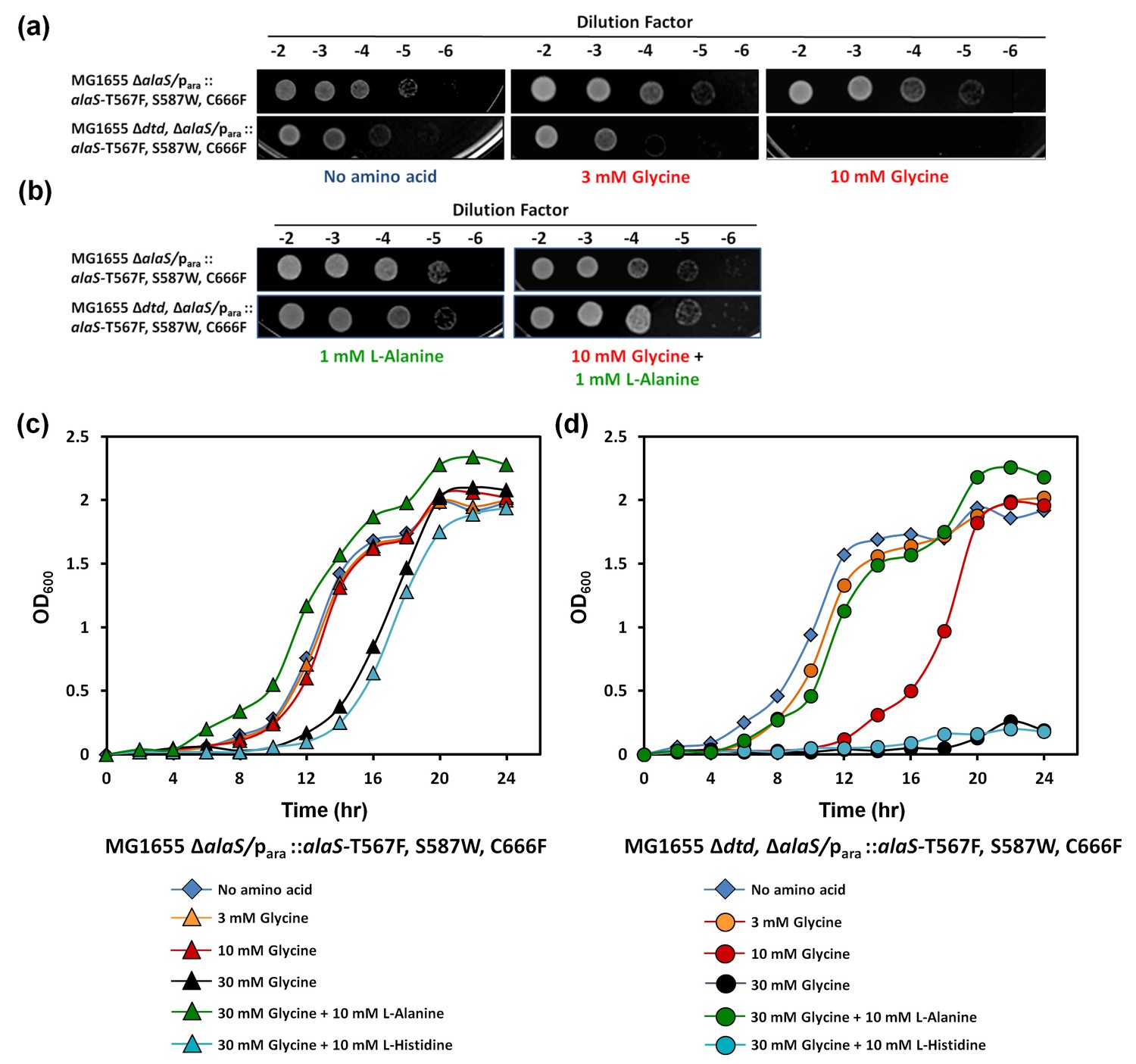
DTD knockout causes pronounced glycine toxicity in E. coli.
Spot dilution assay of E. coli MG1655 ∆alaS/para: : alaS-T567F, S587W, C666F compared with that of E. coli MG1655 ∆dtd, ∆alaS/para:: alaS-T567F, S587W, C666F (a) in the presence of no amino acid, 3 mM glycine, or 10 mM glycine, and (b) in the presence of 1 mM L-alanine, or 10 mM glycine and 1 mM L-alanine. (c) Growth curve of E. coli MG1655 ∆alaS/para: : alaS-T567F, S587W, C666F supplemented with no amino acid (blue diamond), 3 mM glycine (orange triangle), 10 mM glycine (red triangle), 30 mM glycine (black triangle), 30 mM glycine and 10 mM L-alanine (green triangle), 30 mM glycine and 10 mM L-histidine (cyan triangle) (d) Growth curve of E. coli MG1655 ∆dtd, ∆alaS/para: : alaS-T567F, S587W, C666F supplemented with no amino acid (blue diamond), 3 mM glycine (orange circle), 10 mM glycine (red circle), 30 mM glycine (black circle), 30 mM glycine and 10 mM L-alanine (green circle), 30 mM glycine and 10 mM L-histidine (cyan circle).
-
Figure 5—source data 1
Growth curves of E. coli MG1655 with and without dtd knockout in AlaRS editing-defective background.
- https://doi.org/10.7554/eLife.24001.012
tRNAAla-specific G3•U70 wobble base pair acts as a positive determinant for DTD
Significantly high (~1000-fold higher) activity of DTD on Gly-tRNAAla when compared to that on Gly-tRNAGly (Figure 6d, Figure 6—figure supplement 1) indicated that tRNAAla must have some positive determinants for DTD. Since G3•U70 is unique to and one of the major identity elements of tRNAAla across all life forms (Hou and Schimmel, 1988; 1989; McClain and Foss, 1988; Ripmaster et al., 1995; Shiba et al., 1995), we envisaged that DTD could be positively selecting tRNAAla using the wobble base pair. To test this hypothesis, we transplanted G3•U70 at the same position in tRNAGly. We found that DTD had more than 10-fold increased activity on Gly-tRNAGly harboring G3•U70 as compared to the wild-type achiral cognate substrate (Figure 6a,c). To strengthen this hypothesis, we substituted the G3•C70 in the wild-type Gly-tRNAGly with the other Watson-Crick base pair, that is A3•U70. This substitution did not cause any increase in the activity of DTD (Figure 6b). This clearly suggests that the G3•U70 wobble base pair, which is a universally conserved feature of tRNAAla, acts as a positive determinant for DTD. It also suggests the existence of other features in tRNAAla distinct from tRNAGly that accounts for the higher activity of DTD on tRNAAla when compared to tRNAGly, and this aspect requires further exploration. Since DTD is expected to act on all D-aminoacyl-tRNAs, it was assumed that there is no specificity code for its action on tRNAs. The identity determinant–switching experiment resulting in higher activity suggests for the first time an underlying tRNA-based code for DTD action.
Figure 6 with 1 supplement see all

DTD positively selects the tRNA acceptor stem element G3•U70.
(a) Deacylation of Gly-tRNAGly by buffer (blue diamond), 5 nM EcDTD (purple square), 50 nM EcDTD (pink circle). (b) Deacylation of Gly-tRNAGly A3•U70 by buffer (blue diamond), 5 nM EcDTD (purple square), 50 nM EcDTD (pink circle). (c) Deacylation of Gly-tRNAGly G3•U70 by buffer (blue diamond), 500 pM EcDTD (green triangle), 5 nM EcDTD (purple square), 50 nM EcDTD (pink circle). (d) Deacylation of Gly-tRNAAla by buffer (blue diamond), 5 pM EcDTD (orange diamond). Error bars indicate one standard deviation from the mean of triplicate readings.
-
Figure 6—source data 1
Deacylation of Gly-tRNAGly mutants and Gly-tRNAAla by DTD.
- https://doi.org/10.7554/eLife.24001.016
DTD is recruited throughout bacteria and eukaryotes for Gly-tRNAAla removal
DTD’s ubiquitous presence in bacteria and eukaryotes prompted us to test whether its role in clearing mischarged Gly-tRNAAla, in addition to its role in proofreading D-aminoacyl-tRNAs, is conserved in these two domains of life. It is important to investigate this aspect because we found significant differences in residues—which are believed to interact with the acceptor stem of tRNA—around the chiral proofreading site of DTD. To this end, we first superimposed crystal structures of DTD from various organisms and manually docked tRNAAla (tRNAAla was taken from PDB id: 3WQY) on the superposed structures. Since DTD is not expected to establish contacts beyond the acceptor stem of tRNA, we took into consideration only those residues of DTD that were within 6 Å from the 3′-terminal CCA-arm of tRNA (Figure 7—figure supplement 1b), and looked for their conservation/variation using structure-based multiple sequence alignment (Figure 7—figure supplement 1a). Notably, only 7 out of 20 residues in the selected region are invariant across bacterial and eukaryotic DTDs (Figure 7—figure supplement 1).
Given the significant differences observed in a region around the active site of DTD which is likely to interact with tRNA, it is not unreasonable to anticipate that they may affect DTD’s activity on Gly-tRNAAla. To ascertain this, we tested DTDs from different eukaryotes—Leishmania major (LmDTD), Drosophila melanogaster (DmDTD) and Danio rerio (DrDTD)—spanning the entire spectrum of unicellular, invertebrate and vertebrate species, in addition to DTDs from Plasmodium falciparum (PfDTD) and E. coli (EcDTD). These DTDs were tested on glycine mischarged on both E. coli tRNAAla and D. melanogaster tRNAAla (Figure 7). All these DTDs were found to act effectively on both bacterial and eukaryotic tRNAs at 10 pM of DTD concentration. This indicates that in spite of the cross-species differences in DTD and tRNAAla, DTD’s activity on Gly-tRNAAla is most likely conserved throughout bacteria and eukaryotes.
Figure 7 with 1 supplement see all

DTD edits Gly-tRNAAla across bacteria and eukaryotes.
(a) Deacylation of E. coli Gly-tRNAAla by buffer (blue diamond), 10 pM EcDTD (red square), 10 pM PfDTD (green triangle), 10 pM LmDTD (purple cross), 10 pM DmDTD (cyan star), 10 pM DrDTD (orange circle). (b) Deacylation of D. melanogaster Gly-tRNAAla by buffer (blue diamond), 10 pM EcDTD (red square), 10 pM PfDTD (green triangle), 10 pM LmDTD (purple cross), 10 pM DmDTD (cyan star), 10 pM DrDTD (orange circle). Error bars indicate one standard deviation from the mean of triplicate readings.
-
Figure 7—source data 1
Deacylation of E. coli Gly-tRNAAla and D. melanogaster Gly-tRNAAla by bacterial and eukaryotic DTDs.
- https://doi.org/10.7554/eLife.24001.019
Discussion
This study provides an unprecedented solution to a fundamental and long-standing puzzle by elucidating a hitherto unknown and physiologically important function of DTD, which was till now implicated only in enforcing homochirality during translation of the genetic code. The discovery of DTD as a key cellular factor for the elimination of Gly-tRNAAla provides an elegant explanation as to why glycine mischarging by AlaRS was not encountered or considered as a cellular hazard in all the previous studies. So far, only serine mischarging on tRNAAla by AlaRS was believed to be the major threat to the cell, since cells harboring editing-defective AlaRS would show toxicity only to low levels of serine but not glycine (Beebe et al., 2003; Lee et al., 2006). However, this observation seemed puzzling for two reasons. Firstly, glycine misactivation by AlaRS is known to occur at about twice the rate of serine misactivation (Tsui and Fersht, 1981). Secondly and more importantly, it is unlikely that a defect in the proofreading domain that edits both serine and glycine would cause toxicity only due to serine but not glycine. Moreover, for a protein’s structure and function, the substitution of glycine for alanine is expectedly more subversive than the substitution of serine for alanine (Betts et al., 2003). Our study thus brings forth the criticality of glycine mischarging problem, which was largely overlooked and underappreciated prior to this work. It also reveals that nature was forced to devise and retain throughout evolution a key checkpoint in the form of DTD that is more efficient than even AlaRS’s proofreading function to tackle this problem (Figure 8). However, for E. coli under laboratory conditions, knockout of DTD in AlaRS editing-proficient background did not cause toxicity on glycine supplementation. Very likely, errors caused due to defect in proofreading by DTD knockout are tolerated by E. coli, as noted in several cases of other proofreading deficiencies in E. coli (Reynolds et al ., 2010a, 2010b). Moreover, defects in error correction are known to manifest in toxic effects only in special growth conditions like oxidative stress, oxygen deprivation, starvation etc. (Bullwinkle et al., 2014; Cvetesic et al., 2014; Kermgard et al., 2017).
Figure 8

DTD doubles as a key factor to uncouple glycine mischarged on tRNAAla.
In the cell, aminoacylation by aaRSs leads to the formation of different aa-tRNAs, of which L-aa-tRNAs (left extreme) are not acted upon by DTD, while D-aa-tRNAs (right extreme) are effectively decoupled in the presence or absence of EF-Tu, thereby enforcing homochirality. Glycylated tRNAs are acted upon by DTD (centre) but EF-Tu offers protection to the cognate Gly-tRNAGly to prevent its misediting, while the mischarged/non-cognate Gly-tRNAAla is efficiently cleared even in the presence of EF-Tu. Thick connecting arrows indicate the cellular scenario, wherein both DTD and EF-Tu are present.
The study also highlights the necessity of keeping DTD’s active site design intact during the course of evolution, probably because removal of the mischarged Gly-tRNAAla species from the cellular pool took precedence over DTD’s unwarranted activity on Gly-tRNAGly. Nevertheless, the glycine ‘misediting paradox’ was effectively resolved by safeguarding the cognate achiral substrate using EF-Tu as well as keeping the cellular levels of DTD low and tightly regulated (Routh et al., 2016). Thus, what seemed to be an apparent flaw in the architecture of DTD’s active site proved to be a necessity. Moreover, the dual activity of DTD on both achiral and D-chiral substrates depicts it as a plausible ‘connecting link’ or ‘bridging factor’ between D-chirality–based and the canonical L-chirality–based proofreading during protein biosynthesis (Figure 8). This view gains support from the fact that a DTD-like fold appended to archaeal threonyl-tRNA synthetase (ThrRS) as the N-terminal editing domain (NTD) is specific for editing L-serine mischarged on tRNAThr (Ahmad et al., 2015; Dwivedi et al., 2005; Hussain et al., 2006, 2010). It is also worth noting here that the DTD-like fold present in two functional contexts—as NTD in archaea, and as DTD in bacteria and eukaryotes—operates majorly through main chain-mediated contacts for substrate recognition and performs catalysis through RNA, suggesting its primordial origins (Ahmad et al., 2013, 2015; Routh et al., 2016).
Glycine mischarging by AlaRS is inevitable and is a classic case of error made by the aminoacylation site of aaRS, whereas serine mischarging is an offshoot of amino group selection for alanine (Guo et al., 2009). DTD’s significantly higher activity on Gly-tRNAAla as compared to AlaRS suggests that wherever and whenever present, DTD plays the major role in clearing the non-cognate achiral substrate from the cellular pool. This probably helps the cell to overcome the double-discrimination problem that is encountered by AlaRS in all extant organisms. In the primordial scenario, DTD could have been primarily employed as a glycine-removing factor. This activity of DTD could have aided the formation of relatively rigid and stable peptide/protein scaffolds by precluding glycine misincorporation, since the latter would have been detrimental to their stability.
Another interesting facet of DTD that has emerged from this study is its ability to specifically recognize G3•U70 in the acceptor arm of tRNAAla. This wobble base pair is a unique and major identity determinant of tRNAAla which marks it for both aminoacylation and deacylation by AlaRS from bacteria to humans (Beebe et al., 2008; Hou and Schimmel, 1988; McClain and Foss, 1988). The specificity of AlaRS for G3•U70 is so robust that incorporation of this base pair into other tRNAs or minihelices converts non-alanine-accepting tRNAs to be recognized and charged by AlaRS (Musier-Forsyth and Schimmel, 1999). This primordial mode of recognition was proposed to be an acceptor stem-based genetic code that could have been operational since the pre-tRNA era. The G3•U70-based selection of tRNAAla by DTD clearly indicates towards the role of DTD in editing Gly-tRNAAla even before the recruitment of AlaRS editing domains, which have primarily evolved to remove serine mischarged on tRNAAla (Novoa et al., 2015). Furthermore, modulation of DTD’s activity depending on tRNA elements is counter-intuitive as DTD is expected to act on multiple tRNAs with comparable efficiencies. Hence, the present work has unveiled a completely new aspect of DTD’s aminoacyl-tRNA recognition code in which the role of the amino acid as well as the tRNA component needs to be looked at separately. Interestingly, a recognition code exists for EF-Tu, wherein three successive base pairs in the T-stem of tRNA thermodynamically compensate for the differential binding affinity of EF-Tu toward different amino acids (LaRiviere et al., 2001; Roy et al., 2007; Sanderson and Uhlenbeck, 2007b; Schrader et al., 2009). Thus, it becomes important to understand how DTD treats multiple aminoacyl-tRNAs using its own recognition code.
It is interesting to note that in certain cellular contexts, glycine as well as some D-amino acids can be present in relatively high concentrations, wherein these amino acids play important physiological roles. For example, in neuronal tissues, D-serine and D-aspartate along with glycine are abundant and act as neurotransmitters/neuromodulators (Hashimoto and Oka, 1997; Snyder and Kim, 2000). In such instances, especially in neuronal tissues, DTD’s function and its corresponding up-regulation (Zheng et al., 2009) suggest an all-pervasive requirement of this protein from a primordial domain involved in perpetuation of homochirality to current-day proofreader in physiological context. It has been established that even a mild compromise in AlaRS editing for Ser-tRNAAla causes severe pathological conditions, such as neurodegeneration and cardiomyopathy in mouse (Lee et al., 2006; Liu et al., 2014). In this regard, the role and regulation of DTD in various cellular contexts, and more importantly in higher eukaryotes, will be an important aspect which needs to be probed to gain newer insights into DTD’s physiological significance.
Materials and methods
Cloning, expression and protein purification
Request a detailed protocolDTD genes from the genome of E. coli and cDNAs of P. falciparum, L. major, D. melanogaster (fruit fly) and D. rerio (zebrafish) were cloned, and the proteins were expressed and purified as described previously (Ahmad et al., 2013; Routh et al., 2016). The gene (alaS) encoding E. coli AlaRS (EcAlaRS) cloned in pET-26b vector was a gift from Prof. William H. McClain (University of Wisconsin-Madison, USA). An N-terminal 6X His-tagged (N-His) construct was made for EcAlaRS in pET-28b using restriction-free cloning method (van den Ent and Löwe, 2006). The plasmid containing EcAlaRS gene was transformed in E. coli BL21(DE3) cells for protein overexpression. The N-His-tagged EcAlaRS protein was purified by a two-step protocol involving Ni-NTA affinity chromatography and size exclusion chromatography (SEC). The cells were lysed in lysis buffer containing 50 mM Tris–HCl pH 8.0, 150 mM NaCl, 5 mM 2-Mercaptoethanol (β-ME), and 10 mM imidazole. The same buffer was used to pre-equilibrate Ni-NTA column on which the cell lysate was loaded. After loading, the column was first washed with lysis buffer followed by wash buffer containing 50 mM Tris–HCl pH 8.0, 150 mM NaCl, 5 mM β-ME, and 30 mM imidazole. Protein was eluted with elution buffer containing 50 mM Tris–HCl pH 8.0, 150 mM NaCl, 5 mM β-ME, and 250 mM imidazole. The fractions containing protein of interest were pooled, concentrated and subjected to SEC purification using Superdex-200 in a buffer containing 100 mM Tris–HCl pH 8.0, 300 mM NaCl, and 10 mM β-ME. Finally, the fractions containing purified protein were pooled, concentrated and mixed with equal volume of 100% glycerol before storing as aliquots at −30°C for further use.
The gene (tufA) encoding EF-Tu was PCR-amplified from the genomic DNA of Thermus thermophilus, and cloned in pET-28b using restriction-free cloning method (van den Ent and Löwe, 2006). The N-His-tagged EF-Tu protein from T. thermophilus was was then overexpressed in E. coli BL21(DE3) cells and purified by a two-step method involving Ni-NTA affinity chromatography and SEC. The overall purification protocol remained very similar to the one described above except for changes in the buffer composition used in both steps. For Ni-NTA affinity chromatography, all the buffers contained 50 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) titrated with potassium hydroxide (KOH), that is, HEPES-KOH, pH 7.5, 500 mM NaCl, 10 mM magnesium chloride (MgCl2), 5% glycerol, 5 mM β-ME, and 100 µM guanosine-5′-diphosphate (GDP). Additionally, lysis, wash and elution buffers contained 10 mM, 30 mM and 250 mM imidazole, respectively. Following affinity chromatography, SEC was carried out using Sephadex G-200 and a buffer containing 50 mM HEPES-KOH pH 7.5, 500 mM ammonium chloride (NH4Cl), 20 mM MgCl2, 10% glycerol, 5 mM dithiothreitol (DTT), and 100 µM GDP. Finally, the purified protein was processed and stored at −30°C as described above. All protein purification steps from cell lysis onwards were carried out on ice or at 4°C.
The mutants were generated using QuickChange XL Site-directed kit (Stratagene, La Jolla, CA).
Biochemical assays
Request a detailed protocolE. coli tRNAAla was charged with alanine, serine and glycine by EcAlaRS C666A mutant as described by Pasman et al. (2011). The same protocol was followed to charge D. melanogaster tRNAAla with glycine. E. coli tRNAGly was charged with glycine by T. thermophilus GlyRS as described by Routh et al. (2016). Deacylation assays with AlaRS and DTD were carried out as described by Pasman et al. (2011) and Ahmad et al. (2013). EF-Tu activation was carried out as described by Routh et al. (2016). It is to be noted that considering only 10–15% efficiency of EF-Tu activation reaction (Cvetesic et al., 2013; Sanderson and Uhlenbeck, 2007a), the effective (activated) EF-Tu concentration in our assay conditions was in the range of 200–300 nM when the total EF-Tu concentration used was 2 µM. Aminoacylation competition assays were performed in a solution of 100 mM HEPES pH 7.2, 2.5 mM DTT, 2 mM adenosine-5′-triphosphate (ATP), and 200 mM amino acid (alanine/serine/glycine) with 2 μM (total concentration) EF-Tu, 100 nM tRNAAla, 100 nM EcAlaRS and 10 pM EcDTD. These assays were performed at 37°C and were tracked for 15 min. Deacylation competition assays were also carried out in a solution of 100 mM HEPES pH 7.2, and 2.5 mM DTT with 2 μM (total concentration) EF-Tu, 200 nM aa-tRNAAla and varying concentrations of EcAlaRS and EcDTD. Unless otherwise stated, the tRNAs used in the assays were from E. coli. Every data point denotes the mean of three independent readings. Error bars represent one standard deviation from the mean.
Strain constructions
Construction of ΔalaS deletion mutant
Request a detailed protocolWild-type E. coli alaS (WT) gene and editing-defective alaS triple-mutant T567F/S587W/C666F (TM) were cloned into SacI/HindIII sites of pBAD33 vector (CamR, pACYC origin) under the control of an arabinose-inducible promoter. E. coli MG1655 strain (RRID: SCR_002804) was transformed with pBAD33 WT-AlaRS and TM-AlaRS, and the transformants were selected on LB-agar plate containing chloramphenicol (20 µg ml−1 final concentration) at 37°C. These strains were used for making ΔalaS::Kan deletion using P1 phage-mediated transductions (Miller, 1992). P1 phage for alaS knockout was prepared from a Keio collection deletion mutant JW 2667, which had a duplication of alaS (Baba et al., 2006). The presence of the deletion was confirmed by PCR amplification and sequencing the junctions of the deletion–insertion.
Construction of Δdtd∆alaS deletion mutant
Request a detailed protocolMG1655 Δdtd::Kan was generated using P1 lysate from Keio collection (JW 3858–2). Marker-less Δdtd was generated by flipping out the antibiotic resistance (Kan) marker by transforming with plasmid pCP20 (Datsenko and Wanner, 2000). This mutant strain was transformed with pBAD33 WT-AlaRS and TM-AlaRS. ΔalaS::Kan was introduced into these strains to generate Δdtd∆alaS. Deletion mutants were selected on LB-agar plate containing kanamycin (25 µg ml−1 final concentration), chloramphenicol (20 µg ml−1 final concentration) and 0.4% (w/v) L-arabinose.
Viability assays
Request a detailed protocolViability assays were performed with deletion mutant strains of ΔalaS and Δdtd∆alaS in minimal medium (Miller, 1992). Relevant cultures were grown until OD600 reached 0.6 and were 10-fold serially diluted (10−2, 10−3, 10−4, 10−5 and 10−6). Of each serially diluted sample, 3 μl was spotted on minimal agar plates containing 0.002% L-arabinose, 0.2% maltose as carbon source, glycine (3 mM or 10 mM) and/or L-alanine (1 mM). The plates were incubated at 37°C for 20–36 hr.
For growth curves, primary cultures were grown in LB medium containing 0.0002% L-arabinose, Kanamycin and Chloramphenicol at 37°C until OD600 reached 1.0. Subsequently, 2% inoculum was used to initiate 15 ml secondary culture in 1X minimal medium containing 0.2% maltose as carbon source and 0.0002% L-arabinose. The secondary culture was grown at 37°C to obtain a cell density (OD600) of ~0.6. These cultures were again grown in 1X minimal medium with/without amino acids (glycine, L-alanine and L-histidine) of the indicated concentrations. The growth was monitored at every 2-hr interval. All the experiments were done in triplicates.
References
-
Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collectionMolecular Systems Biology 2:2006.0008.https://doi.org/10.1038/msb4100050
-
Bioinformatics for GeneticistsAmino acid properties and consequences of subsitutions, Bioinformatics for Geneticists, Wiley.
-
D-Tyrosyl RNA: formation, hydrolysis and utilization for protein synthesisJournal of Molecular Biology 26:39–54.https://doi.org/10.1016/0022-2836(67)90259-8
-
The physiological target for LeuRS translational quality control is norvalineThe EMBO Journal 33:1639–1653.https://doi.org/10.15252/embj.201488199
-
A D-amino acid editing module coupled to the translational apparatus in archaeaNature Structural & Molecular Biology 12:556–557.https://doi.org/10.1038/nsmb943
-
Structural analyses clarify the complex control of mistranslation by tRNA synthetasesCurrent Opinion in Structural Biology 22:119–126.https://doi.org/10.1016/j.sbi.2011.11.008
-
Free D-aspartate and D-serine in the mammalian brain and peripheryProgress in Neurobiology 52:325–353.https://doi.org/10.1016/S0301-0082(97)00019-1
-
Aminoacyl-tRNA synthesisAnnual Review of Biochemistry 69:617–650.https://doi.org/10.1146/annurev.biochem.69.1.617
-
Amino acid toxicities of Escherichia coli that are prevented by leucyl-tRNA synthetase amino acid editingJournal of Bacteriology 189:8765–8768.https://doi.org/10.1128/JB.01215-07
-
BookA Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia Coli and Related BacteriaCold Spring: Cold Spring Harbor Laboratory.
-
Multiple Quality Control Pathways Limit Non-protein amino acid use by yeast cytoplasmic Phenylalanyl-tRNA synthetaseJournal of Biological Chemistry 291:15796–15805.https://doi.org/10.1074/jbc.M116.726828
-
Mistranslation of the genetic codeFEBS Letters 588:4305–4310.https://doi.org/10.1016/j.febslet.2014.08.035
-
Atomic determinants for aminoacylation of RNA minihelices and relationship to Genetic CodeAccounts of Chemical Research 32:368–375.https://doi.org/10.1021/ar970148w
-
Genetic code ambiguity. cell viability related to the severity of editing defects in mutant tRNA synthetasesJournal of Biological Chemistry 277:45729–45733.https://doi.org/10.1074/jbc.M208093200
-
Ancestral AlaX editing enzymes for control of genetic code fidelity are not tRNA-specificJournal of Biological Chemistry 290:10495–10503.https://doi.org/10.1074/jbc.M115.640060
-
Structural insights into translational fidelityAnnual Review of Biochemistry 74:129–177.https://doi.org/10.1146/annurev.biochem.74.061903.155440
-
Cellular mechanisms that control mistranslationNature Reviews Microbiology 8:849–856.https://doi.org/10.1038/nrmicro2472
-
Structural elements defining elongation factor Tu mediated suppression of Codon ambiguityNucleic Acids Research 35:3420–3430.https://doi.org/10.1093/nar/gkm211
-
Directed mutagenesis identifies amino acid residues involved in elongation factor Tu binding to yeast Phe-tRNA PheJournal of Molecular Biology 368:119–130.https://doi.org/10.1016/j.jmb.2007.01.075
-
Understanding the sequence specificity of tRNA binding to elongation factor tu using tRNA mutagenesisJournal of Molecular Biology 386:1255–1264.https://doi.org/10.1016/j.jmb.2009.01.021
-
D-amino acids as putative neurotransmitters: focus on D-serineNeurochemical Research 25:553–560.https://doi.org/10.1023/A:1007586314648
-
Metabolism of D-aminoacyl-tRNAs in Escherichia coli and Saccharomyces cerevisiae cellsJournal of Biological Chemistry 275:32535–32542.https://doi.org/10.1074/jbc.M005166200
-
Functional characterization of the D-Tyr-tRNATyr deacylase fromJournal of Biological Chemistry 274:19109–19114.https://doi.org/10.1074/jbc.274.27.19109
-
RF cloning: a restriction-free method for inserting target genes into plasmidsJournal of Biochemical and Biophysical Methods 67:67–74.https://doi.org/10.1016/j.jbbm.2005.12.008
-
Distinct kinetic mechanisms of the two classes of Aminoacyl-tRNA synthetasesJournal of Molecular Biology 361:300–311.https://doi.org/10.1016/j.jmb.2006.06.015
Article and author information
Author details
Komal Ishwar Pawar

Rajan Sankaranarayanan
Funding
Council of Scientific and Industrial Research (12th Five Year Plan Project BSC0113)
- Rajan Sankaranarayanan
Science and Engineering Research Board (JC Bose Fellowship)
- Rajan Sankaranarayanan
Department of Biotechnology, Ministry of Science and Technology (Centre of Excellence)
- Rajan Sankaranarayanan
Council of Scientific and Industrial Research (Research Fellowship)
- Komal Ishwar Pawar
- Satya Brata Routh
Department of Biotechnology, Ministry of Science and Technology (Research Associateship)
- Katta Suma
Department of Biotechnology, Ministry of Science and Technology (INSPIRE Fellowship)
- Santosh Kumar Kuncha
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
The authors acknowledge Dr. Manjula Reddy and L. Sai Sree for their help with cell-based toxicity assays. We thank NBRP (Japan): E. coli for Keio collection mutants. KIP and SBR thank Council of Scientific and Industrial Research (CSIR), India, KS thanks DBT-RA Programme, India, and SKK thanks DST-INSPIRE, India, for research fellowships. RS acknowledges funding from 12th Five Year Plan Project BSC0113 of CSIR, India, JC Bose Fellowship of SERB, India and Centre of Excellence Project, DBT, India. The funding agencies had no role in study design, analysis, decision to publish or preparation of the manuscript.
Copyright
© 2017, Pawar et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,726
- views
-
- 415
- downloads
-
- 39
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 39
- citations for umbrella DOI https://doi.org/10.7554/eLife.24001
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Role of D-aminoacyl-tRNA deacylase beyond chiral proofreading as a cellular defense against glycine mischarging by AlaRS
eLife 6:e24001.
https://doi.org/10.7554/eLife.24001




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}