Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorGáspár JékelyHeidelberg University, Heidelberg, Germany
- Senior EditorAlbert CardonaUniversity of Cambridge, Cambridge, United Kingdom
Reviewer #1 (Public review):
Summary:
The manuscript characterizes a functional peptidergic system in the echinoderm Apostichopus japonicus that is related to the widely conserved family of calcitonin/diuretic hormone 31 (CT/DH31) peptides in bilaterian animals. In vitro analysis of receptor-ligand interactions, using multiple receptor activation assays, identifies three cognate receptors for two CT-like peptides in the sea cucumber, which stimulate cAMP, calcium, and ERK signaling. Only one of these receptors clusters within the family of calcitonin and calcitonin-like receptors (CTR/CLR) in bilaterian animals, whereas two other receptors cluster with invertebrate pigment dispersing factor receptors (PDFRs). In addition, this study sheds light on the expression and in vivo functions of CT-like peptides in A. japonicus, by quantitative real-time PCR, immunohistochemistry, pharmacological experiments on body wall muscle and intestine preparations, and peptide injection and RNAi knockdown experiments. This reveals a conserved function of CT-like peptides as muscle relaxants and growth regulators in A. japonicus.
Strengths:
This work combines both in vitro and in vivo functional assays to identify a CT-like peptidergic system in an economically relevant echinoderm species, the sea cucumber A. japonicus. A major strength of the study is that it identifies three G protein-coupled receptors for AjCT-like peptides, one related to the CTR/CLR family and two related to the PDFR family. A similar finding was previously reported for the CT-related peptide DH31 in Drosophila melanogaster that activates both CT-type and PDF-type receptors. Here, the authors expand this observation to a deuterostomian animal, which suggests that receptor promiscuity is a more general feature of the CT/DH31 peptide family and that CT/DH31-like peptides may activate both CT-type and PDF-type receptors in other animals as well.
Besides the identification of receptor-ligand pairs, the downstream signaling pathways of AjCT receptors have been characterized, revealing broad and in some cases receptor-specific effects on cAMP, calcium, and ERK signaling.
Functional characterization of the CT-related peptide system in heterologous cells is complemented with ex vivo and in vivo experiments. First, peptide injection and RNAi knockdown experiments establish transcriptional regulation of all three identified receptors in response to changing AjCT peptide levels. Second, ex vivo experiments reveal a conserved role for the two CT-like peptides as muscle relaxants, which have differential effects on body wall muscle and intestine preparations. Finally, peptide injection and knockdown experiments uncover a growth-promoting role for one CT-like peptide (AjCT2). Injection of AjCT2 at high concentration, or long-term knockdown of the AjCT precursor, affects diverse growth-related parameters including weight gain rate, specific growth rate, and transcript levels of growth-regulating transcription factors. The authors also reveal a growth-promoting function for the PDFR-like receptor AjPDFR2, suggesting that this receptor mediates the effects of AjCT2 on growth.
Weaknesses:
The authors present a more detailed phylogenetic analysis in the revised version, including a larger number of species. But some clusters in the analysis are not well supported because they have only low bootstrap values. This makes it difficult to interpret the clustering in some parts of the tree.
Expression of CT-like peptides was investigated both at transcript and protein level, but insight into the expression of the three peptide receptors is limited. This makes it difficult to understand the mechanism underlying the (different) functions of the two CT-like peptides in vivo. The authors identify differences in signal transduction cascades activated by each peptide, which might underpin distinct functions, but these differences were established only in heterologous cells.
The authors show overlapping phenotypes for a long-term knockdown of the AjCT precursor and the AjPDFR2 receptor, suggesting that the growth-regulating functions of AjCT2 are mediated by this receptor pathway. However, it remains unclear whether this mechanism underpins the growth-regulating function of AjCT2, until further in vivo evidence for this ligand-receptor interaction is presented. For example, the authors could investigate whether knockdown of AjPDFR2 attenuates the effects of AjCT2 peptide injection. In addition, a functional PDF system in this species remains uncharacterized, and a potential role of PDF-like peptides in growth regulation has not yet been investigated in A. japonicus. Therefore, it also remains unclear whether the ability of CT-like peptides to activate PDFRs is an evolutionary ancient property of this peptide family or whether this is an example of convergent evolution in some protostomian (Drosophila) and deuterostomian (sea cucumber) species.
Reviewer #2 (Public review):
Summary:
The authors show that A. japonicus calcitonins (AjCT1 and AjCT2) activate not only the calcitonin/calcitonin-like receptor, but they also activate the two "PDF receptors", ex vivo. They also explore secondary messenger pathways that are recruited following receptor activation. They determine the source of CT1 and CT2 using qPCR and in situ hybridization and finally test the effects of these peptides on tissue contractions, feeding and growth. This study provides solid evidence that CT1 and CT2 act as ligands for calcitonin receptors; however, evidence supporting cross-talk between CT peptides and "PDF receptors" is weak.
Strengths:
This is the first study to report pharmacological characterization of CT receptors in an echinoderm. Multiple lines of evidence in cell culture (receptor internalization and secondary messenger pathways) support this conclusion.
Weaknesses:
The authors claim that A. japonicus CTs activate "PDF" receptors and suggest that this cross-talk is evolutionary ancient since similar phenomenon also exists in the fly Drosophila melanogaster. These conclusions are not fully supported. The authors perform phylogenetic analysis to show that the two "PDF" receptors form an independent clade. The bootstrap support is quite low in a lot of instances, especially for the deuterostomian and protostomian PDFR clades which is below 30. With such low support, it is unclear if the clade comprising deuterostomian "PDFR" is in fact PDFRs and not another receptor type whose endogenous ligand (besides CT) remains to be discovered.
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1 (Public review):
Weaknesses:
(1). Analysis of transcript expression is limited to the CT-peptide encoding gene, while no gene expression analysis was attempted for the three identified receptors. Differences in the activation of downstream signaling pathways between the three receptors are also questionable due to unclarities in the statistical analysis and variation in the control and experimental data in heterologous assays. Together, this makes it difficult to propose a mechanism underlying differences in the functions of the two CT-like peptides in muscle control and growth regulation.
We appreciate the reviewer's rigorous critique. The manuscript has been comprehensively revised as follows:
(1) For the expression analysis of the three identified receptors, the updated results are presented in Figure 5, with the detailed descriptions in Results section 2.4 (line 287-290) and Materials and Methods section 4.5 (line 767).
(2) For the statistical tests and methodological clarity, statistical tests were indeed performed for all experiments. However, we acknowledge that the original labeling methods required enhanced methodological clarity, and we apologize for any confusion caused. All figures have been revised to improve the visibility of differences, and statistical test information has been added to both the figure legends and the Materials and methods section “4.10 Statistical Analysis” (line 900-910).
(3) For the variation in the control and experimental data, the minor observed variations in control conditions across experiments primarily arise from two methodological factors: 1) Each experimental set used cells transfected with distinct receptor subtypes (e.g., AjPDFR1 vs. AjPDFR2), inherently introducing baseline variability due to differential receptor expression profiles. 2) Independent cell culture batches were employed for replicate experiments to ensure biological reproducibility. Importantly, these minor variations did not compromise the statistical significance of downstream signaling differences (p < 0.01 for all comparative analyses). Therefore, differences in the activation of downstream signaling pathways between the three receptors are reliable.
(2) The authors also suggest a putative orexigenic role for the CT-like peptidergic system in feeding behavior. This effect is not well supported by the experimental data provided, as no detailed analysis of feeding behavior was carried out (only indirect measurements were performed that could be influenced by other peptidergic effects, such as on muscle relaxation) and no statistically significant differences were reported in these assays.
Thank you for the reviewer’s valuable comments. Our revised manuscript now includes the following multidimensional analyses to strengthen evidence of the orexigenic role of AjCT2: Firstly, in sea cucumbers, the mass of remaining bait is a common indicator of feeding condition. After long-term AjCT2 injection, this value was significantly decreased in comparison with control group during phase V (Figure 8A-figure supplement 1), which indicates that AjCT2 promotes feeding in A. japonicus. Correspondingly, in long-term loss-of-function experiments (newly added in the revised manuscript), the remaining bait in the siAjCTP1/2-1 group was significantly increased in comparison with siNC group form phase II to IV (Figure 10B). The detailed descriptions of these supplementary experiments have been added to Results Section 2.6 (lines 390-396) and Materials and Methods Section 4.9 (line 879-888).
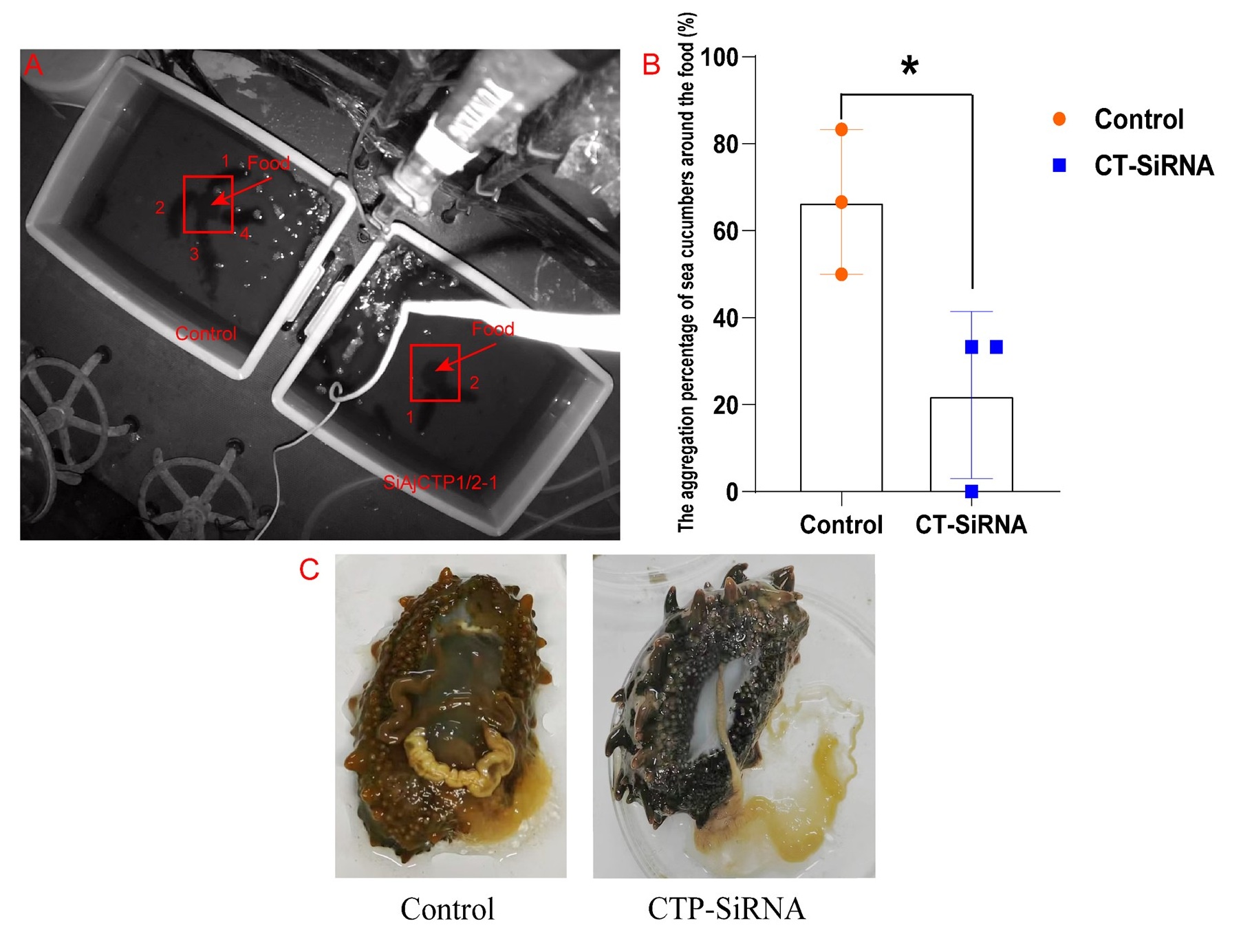
Secondly, after 24 days of continuous injections of siAjCTP1/2-1, we monitored the feeding behavior of these sea cucumbers over three consecutive days. Each day, we removed residual bait and feces, then repositioned fresh food at the tank center. We calculated the aggregation percentage (AP) of sea cucumbers around the food during the feeding peak (2:00-4:00) each day, which is the most reliable indicator of feeding behavior in this species. The results showed that the AP in siAjCTP1/2-1 group was significantly lower than that in control group. Post-dissection observations revealed reduced intestinal food content and significant intestinal degeneration in the siAjCTP1/2-1 group (The figure has been added below). These results indicate that long-term functional loss of AjCT2 reduces food intake and influences the feeding behavior of A. japonicus.
In response to the comment regarding “No statistically significant differences were reported in these assays”, we have modified the figures to clearly visualize the differences and added statistical test details in both the figure legends and the Materials and methodssection “4.10 Statistical analysis” (lines 900–910).
Author response image 1.
The feeding behavior of A. japonicus after long-term loss-of-function of AjCT2. (A) A record of feeding behavior. The red arrow refers to the food and the red box represents the feeding area. The numbers in the figure represent individuals entering into the feeding area. (B) The aggregation percentage (AP) of sea cucumbers around the food during the feeding peak (2:00-4:00) (n=3 days). (C) The degenerated intestine of sea cucumber after 24 days of siAjCTP1/2-1 injection. Data in the graph represent the mean ± standard deviation. *Significant differences between groups (p < 0.05). Control: siNC injection group; CT-SiRNA: siAjCTP1/2 injection group.
(3) Overall, details regarding statistical analyses are not (clearly) specified in the manuscript, and there are several instances where statements are not supported by literature evidence.
Thank you for the reviewer’s comments. Again, we sincerely apologize for the confusion caused. To clarify, statistical tests were performed for all experiments. However, the original labeling may have been somewhat messy. We have revised all figures to enhance the visibility of differences and provided detailed statistical test information in both the figure legends and the Materials and Methods section titled “4.10 Statistical Analysis” (lines 900–910). Additionally, we have supplemented the revised manuscript with further literature evidence to support our statements: (1) citation to Furuya et al. (2000), Johnson et al. (2005), Jékely (2013) and Mirabeau et al. (2013) have been added to clarify the foundation studies on DH31 and DH31 receptors in invertebrates (line 73-74); (2) Conzelmann et al. (2013) and Furuya et al. (2000) were cited to validate the present of two different types of CT-related peptides in protostomes: CT-type peptides (with an N-terminal disulphide bridge) and DH31-type peptides (lacking this feature) (line 78-79); (3) Johnson et al. (2005) was referenced to support the dual ligand-receptor interactions of DH31 in Drosophila, specifically its binding to both CG17415 (a CTR/CLR-related protein) and CG13758 (the PDF receptor) (line 94); (4) Johnson et al. (2005) and Goda et al. (2019) were cited to reinforce the functional significance of dual DH31 receptor pathways in Drosophila, as extensively studied in prior research (line 95-97).
Reviewer #2 (Public review):
Weaknesses:
(1) The authors claim that A. japonicus CTs activate "PDF" receptors and suggest that this cross-talk is evolutionarily ancient since a similar phenomenon also exists in the fly Drosophila melanogaster. These conclusions are not fully supported for several reasons. The authors perform phylogenetic analysis to show that the two "PDF" receptors form an independent clade. This clade is sister to the clade comprising CT receptors. This phylogenetic analysis suffers from several issues. Firstly, the phylogenies lack bootstrap support. Secondly, the resolution of the phylogeny is poor because representative members from diverse phyla have not been included. For instance, insect or other protostomian PDF receptors have not been included so how can the authors distinguish between "PDF" receptors or another group of CT receptors? Thirdly, no in vivo evidence has been presented to support that CT can activate "PDF" receptors in vivo.
We thank the reviewers for their constructive comments. As suggested, we expanded our taxon sampling to include more representative members across diverse phyla and reanalyzed the phylogenetic relationships (including bootstrap tests) in Figure 1C. The revised analysis revealed two distinct clades: one containing CTR/CLR-type receptors and the other PDF-type receptors. Specifically, AjCTR clustered within the CTR/CLR-type receptor group, while AjPDFR1 and AjPDFR2 were placed in the PDF-type receptor clade. The full species names for all taxa were provided in the Supplementary Table 2.
To provide in vivo evidence supporting CT-mediated activation of "PDF" receptors, we conducted the following experiments: Firstly, we confirmed that AjPDFR1 and AjPDFR2 were the functional receptors of AjCT1 and AjCT2 (Figure 2, 3 and 4). Secondly, injection of AjCT2 and siAjCTP1/2-1 in vivo induced corresponding changes in AjPDFR1 and AjPDFR2 expression levels in the intestine (Figure 8C, 9A, 9B and 9C).
(2) The source of CT which mediates the effects on longitudinal muscles and intestine is unclear. Is it autocrine or paracrine signaling by CT from the same tissue or is it long-range hormonal signaling?
Thank you for this feedback. We have now analysed CT-type neuropeptide expression in A. japonicus using immunohistochemistry with the antiserum to the A. rubens CT-type peptde ArCT, which has previously been shown to cross-react with CT-type neuropeptides in other echinoderms (Aleotti et al., 2022). We have added related descriptions in the following sections: Results (section 2.4, line 299-336), Discussion (section 3.3, line 545-554) and Materials and methods (section 4.6, line 785-817). Consistent with this previous finding, the ArCT antiserum labelled neuronal cells and fibers in the central and peripheral nervous system and in the digestive system of A. japonicus (Figure 6). The specificity of immunostaining was confirmed by performing pre-absorption tests with the ArCT antigen peptide (Figure 6-figure supplement 1). The detection of immunostaining in the innervation of the intestine is consistent with PCR results and the relaxing effect of AjCT2 on intestine preparations. Interestingly, no immunostaining was observed in longitudinal muscle, which is inconsistent with the detection of AjCT1/2 transcripts in this tissue. This may reflect differences in the sensitivity of the methods employed to detect transcripts (PCR) and mature peptide (immunohistochemistry). The absence of ArCT-like immunoreactivity in the longitudinal muscles suggests that AjCT1 and AjCT2 may exert relaxing effects on this tissue in vivo via hormonal signaling mechanisms. However, because AjCT1/2 expression in the longitudinal muscles may be below the detection threshold of the ArCT antibodies, we can’t rule out the possibility that AjCT1/2 are released within the longitudinal muscles physiologically.
(3) Pharmacology experiments showing the effects of CT1 and CT2 on ACh-induced contractions were performed. Sample traces have been provided but no traces with ACh alone have been included. How long do ACh-induced contractions persist? These controls are necessary to differentiate between the eventual decay of ACh effects and relaxation induced by CT1 and CT2. The traces also do not reflect the results portrayed in dose-response curves. For instance, in Figure 6B, maximum relaxation is reported for 10-6M. Yet, the trace hardly shows any difference before and after the addition of 10-6M peptide. The maximum effect in the trace appears to be after the addition of 10-8M peptide.
Thank you for the reviewer’s comments. As requested, we have included representative traces of ACh-induced contraction of longitudinal muscle and intestinal preparations (Figure 7—figure supplement 1B and 1C). Notably, the positive control (ACh) maintained contraction effects for at least 15 minutes, consistent with its known pharmacological properties. Regarding Figure 7B (previous Figure 6B), the trace illustrates the cumulative effects of successive neuropeptide treatments at increasing concentrations. A gradual reduction in response amplitude was observed at the highest peptide concentration, likely reflecting receptor desensitization, a phenomenon previously reported for neuropeptide Y and oxytocin (Tsurumaki et al., 2003; Arrowsmith and Wray, 2014). These results are now explicitly described in the Results Section 2.5 (lines 340-345 and 348-352) and discussed in Section 3.3 (lines 569-574). In response to the reviewer’s suggestion, we further tested the pharmacological effects of AjCT2 at 10⁻⁶ M. As shown in Figure 7—figure supplement 1A, this concentration induced maximal relaxation, confirming its dose-dependent efficacy.
(4) I am unsure how differences in wet mass indicate feeding and growth differences since no justification has been provided. Couldn't wet mass also be influenced by differences in osmotic balance, a key function of calcitonin-like peptides in protostomian invertebrates? The statistical comparisons have not been included in Figure 7B.
We appreciate the reviewer's insightful comments. We fully concur that wet mass constitutes an inadequate indicator for evaluating feeding and growth variations. Consequently, we reassessed A. japonicus growth parameters using two established metrics: weight gain rate (WGR) and specific growth rate (SGR), to delineate differences between experimental and control groups. Notably, the high-concentration AjCT2 injection group exhibited statistically significant increases in both WGR and SGR relative to controls (Figure 8A). This demonstrates a putative physiological role of AjCT2 signaling in enhancing feeding efficiency and growth performance in A. japonicus. Detailed methodologies are provided in the Materials and methods Section 4.8 (lines 847-851), with corresponding results presented in the Results Section 2.6 (lines 370-375). Besides, Cong et al., (2024) reported holotocin-induced osmoregulatory function in A. japonicus, manifested by significant wet weight elevation and body bloating. However, our AjCT2 intervention showed no such phenotypic alterations, suggesting that AjCT2 likely does not participate in osmotic balance regulation, at least under these experimental conditions. Crucially, the observed WGR and SGR enhancements following AjCT2 administration was not caused by osmoregulatory effects.
(5) While the authors succeeded in knocking down CT, the physiological effects of reduced CT signaling were not examined.
Thank you for the reviewer’s comment. We have supplemented the experiments to investigate the physiological effects of long-term reduced CT signaling following the reviewer’s suggestions, including measuring the dry weight of remaining bait and excrement, calculating the weight gain rate and specific growth rate, and testing the expression levels of three growth factors (AjMegf6, AjGDF-8 and AjIgf) to further assess AjCT2’s role in feeding and growth. The results demonstrated that weight gain rate and specific growth rate in the siAjCTP1/2-1 group were significantly decreased (As shown in Figure 10A). Correspondingly, except in phase I, the siAjCTP1/2-1 group exhibited a significant increase in remaining bait and a decrease in excrement during phases II-VI (Figure 10B). Furthermore, the growth inhibitory factor AjGDF-8 was significantly up-regulated and the growth promoting factor AjMegf6 was significantly down-regulated in siAjCTP1/2-1 group (Figure 10C). These findings further support the potential physiological role of AjCT2 signaling in promoting feeding and growth in A. japonicus. The added results are presented in Figure 10, with related descriptions in Section 2.6 (Results, lines 390-396), Section 3.4 (Discussion, line 597-603) and Section 4.9 (Materials and Methods, lines 879-888).
Reviewer #1 (Recommendations for the authors):
(1) The abstract states that loss-of-function tests (RNAi knockdown) reveal a potential physiological role for AjCT2 signaling in promoting feeding and growth in A. japonicus. However, RNAi knockdown was only followed by analysis of transcript expression of CT-like receptors and not by the assessment of feeding or growth.
Thank you for this helpful feedback. In the revised manuscript, we have supplemented the experiments to investigate the physiological effects of long-term reduced CT signaling, as suggested by the reviewer. These include measuring the dry weight of remaining bait and excrement, calculating the weight gain rate and specific growth rate, and testing the expression levels of the three growth factors (AjMegf6, AjGDF-8 and AjIgf) to further assess the function of AjCT2 on feeding and growth in A. japonicus. The results are as follows:
(1) The weight gain rate and specific growth rate in the siAjCTP1/2-1 group were significantly decreased (As shown in Figure 10A).
(2) Correspondingly, except for the phase I, the siAjCTP1/2-1 group had significantly increased remaining bait and decreased excrement during phases II-VI (Figure 10B).
(3) The growth inhibitory factor AjGDF-8 was significantly up-regulated, while the growth promoting factor AjMegf6 was significantly down-regulated in the siAjCTP1/2-1 group (Figure 10C).
These findings further support the potential physiological role of AjCT2 signaling in promoting feeding and growth in A. japonicus. We have incorporated these results into Figure 10 and added related descriptions in the following sections: Results (section 2.6, line 390-396), Discussion (section 3.4, line 597-603) and Materials and methods (section 4.9, line 879-888).
Regarding the original statement in the abstract “Furthermore, in vivo pharmacological experiments and loss-of-function tests revealed a potential physiological role for AjCT2 signaling in promoting feeding and growth in A. japonicus.” This sentence effectively summarizes our findings. Therefore, we have retained it in the revised manuscript while supplementing the missing experimental details as requested.
(2) Information on the statistical tests that were performed is lacking for most experiments. It is recommended to include this information in the figure legends, in addition to the methods section. Details on the phylogenetic analysis (parameters and statistics used) and calculation of half maximal effective concentrations (calculation methods and confidence intervals) also need to be included in the manuscript.
Thank you for this constructive feedback. As the reviewer suggested, statistical test information has been incorporated into both the figure legends and the “4.10 Statistical Analysis” subsection of the Materials and methods (lines 900-910). Specifically:
(1)Phylogenetic analysis details (parameters and statistical approaches) are now provided in the Materials and methods section 4.2 (line 675-682);
(2) Bootstrap test results supporting the phylogenetic trees have been added to Figure 1B and 1C;
(3)Half-maximal effective concentration (EC₅₀) calculations, including methodologies and confidence intervals, are documented in both the Figure 2B legend and the “4.10 Statistical Analysis” section (lines 900-910).
(3) In some figures (e.g. Figure 5A, 7A), the n number indicated does not match the number of data points shown in the figure panel. It is not clear what n represents here. In Figure 6B, an x-axis label is missing. In some figure legends (e.g. Figure 4 - Figure Supplement 1), the error bars and significance levels are not defined.
We apologize for this error; we have corrected all quantity errors related to "n" in the manuscript’ figure legends. And also, the x-axis label was added in Figure 7B (previous Figure 6B), error bars and significance levels were defined in all figure legends clearly
(4) It would be useful to explain what the difference is between the Cre and SRE luciferase assay and why these two assays were used to study receptor-activated signaling cascades. The source of the synthetic peptides is mentioned, but it is recommended to also state the purity of the synthetic peptides.
Thank you for the valuable comments. As stated in the introduction (line 66-69)- “binding of CT to CTR in the absence of RAMPs can activate signaling via several downstream pathways, including cAMP accumulation, Ca2+ mobilization, and ERK activation.” Based on this established mechanism, we selected cAMP and Ca²⁺ signaling pathways as biomarkers for studying receptor-activated cascades, with the following experimental rationale: CRE-Luc Reporter System functions as a cAMP response element detector and SRE-Luc Reporter System serves as an intracellular Ca²⁺ level indicator. In CRE-Luc detection, when the receptor is activated by a ligand, it couples with Gαs protein to activate the cAMP/PKA signaling pathway. The accumulation of cAMP can lead to the phosphorylation of PKA, and then enhance the transcription of CRE-containing genes. Therefore, significant increase in CRE-Luc activity directly correlates with cAMP accumulation. Similarly, SRE-Luc activity reflects dynamic changes in intracellular Ca2+ levels. We have added the explanation of this part in the materials and methods section 4.4 (line 715-721). The purity of the synthetic peptides was >95%, and we have also added this information in section 4.4 (line 715) according to the reviewer’s suggestion.
(5) In Figure 3B, it is difficult to see receptor internalization in response to the application of synthetic CT-like peptides, and a control condition (without peptide application) is lacking.
Thank you for the reviewer’s comment. The control condition (without peptide application) was added in Figure 3-figure supplement 1, which shows the localization of pEGFP-N1/receptors in the cell membrane. Upon stimulation with synthetic CT-like peptides (Materials and methods section 2.3), the receptors exhibit clear internalization into the cytoplasm, as visualized in Figure 3B through comparative analysis.
(6) Differences in the activation of downstream signaling cascades between the three receptors are questionable because there is substantial variation in the experimental data and control conditions in different experiments (for example, in Figures 3A and 4A). To better represent this variation, it is recommended to plot individual data points onto the bar graphs in all figures and to nuance the interpretation of putative differences in downstream signaling of different receptors. Differences in the physiological roles of CT-like peptides may be explained by various mechanisms, including differences in peptide/receptor expression or in the potency of peptides to activate different receptors in vivo. It would be useful to elaborate on these different explanations in the discussion.
We appreciate the reviewer's critical assessment. The observed variations in control conditions across experiments (e.g., Figures 3A & 4A) primarily arise from two methodological factors: ① Each experimental set used cells transfected with distinct receptor subtypes (e.g., AjPDFR1 vs. AjPDFR2), inherently introducing baseline variability due to differential receptor expression profiles. ② Independent cell culture batches were employed for replicate experiments to ensure biological reproducibility. Importantly, these minor variations did not compromise the statistical significance of downstream signaling differences (p < 0.01 for all comparative analyses). And according to the reviewer’s suggestion, we have plotted individual data points onto the bar graphs in all figures.
And also, according to the reviewer’s suggestion, we have expanded the discussion on receptor-specific signaling cascades in Section 3.4 (lines 589-609). Key findings include: In vivo pharmacological assays demonstrated that only high concentrations of AjCT2 significantly enhanced feeding and growth rates in A. japonicus. In contrast, neither a low concentration of AjCT2 nor any concentration of AjCT1 (low or high) induced detectable effects. Furthermore, long-term knockdown of AjCTP1/2 further validated the essential role of AjCT2 in regulating feeding and growth in this species. To elucidate the receptor mediating AjCT2’s feeding- and growth-promoting effects, we selected AjPDFR2 based on its distinct activation profile: AjCT2 selectively activated AjPDFR2, inducing downstream ERK1/2 phosphorylation, whereas AjCT1 exhibited no activity toward this receptor. Given this receptor specificity, we performed AjPDFR2 knockdown experiments, which revealed phenotypic changes consistent with those in AjCTP1/2 knockdown animals, including significantly reduced WGR and SGR, alongside increased remaining bait accumulation and diminished excrement output compared to control. Collectively, these results support a model wherein AjCT2 promotes feeding and growth in A. japonicus via AjPDFR2-dependent activation of the cAMP/PKA/ERK1/2 and Gαq/Ca²⁺/PKC/ERK1/2 cascades. Considering the inherent complexity of neuropeptide signaling systems, which involve multiple GPCR subtypes coupled to diverse signaling cascades, ligands bound to the same receptor may activate distinct G protein subforms within a single cell (Møller et al., 2003; Mendel et al., 2020). Receptor activation modes may be modulated by structural polymorphisms or binding site diversity (Wong et al., 2000; Changeux, 2010), as well as by the differential efficacy of peptides in activating receptors in vivo.
(7) For the peptide injection experiments, it is recommended to explain the different animal groups in the results section. In addition, injection in the control condition seems to have a small effect on the wet weight. Therefore, it would be useful to compare control-injected and peptide-injected groups after injection.
Thank you for the reviewer’s comments. We have provided an expanded explanation of the animal group classifications in Section 2.6 (lines 367–375). We fully agree that a comparative analysis between the experimental and control groups post-injection is essential. However, since wet weight measurement is suboptimal for demonstrating feeding and growth variations, we re-evaluated the data using two validated metrics: weight gain rate (WGR) and specific growth rate (SGR) of A. japonicus. The results revealed that the high-concentration AjCT2 injection group exhibited significantly elevated weight gain rate and specific growth rate compared to the control group, suggesting a potential role of AjCT2 signaling in promoting feeding and growth in A. japonicus. These results are presented in Figure 8A, with detailed descriptions in Results Section 2.6 (lines 370–375) and methodology in Materials and Methods Section 4.8 (lines 847-851).
(8) Regarding the RNAi knockdown experiments, it is not clear from the methods section what the siNC control exactly is, and how the interference rate is calculated.
Thank you for this comment. The siNC control was siRNA which does not target any genes in A. japonicus, with interference rates quantified through the 2-ΔΔCT method to assess siRNA inhibition efficiency. These methodological details have been incorporated into Materials and Methods Section 4.9 (lines 866–867 and 874-876) for enhanced clarity.
Reviewer #2 (Recommendations for the authors):
(1) Both the phylogenies are missing bootstrap tests. Please include this analysis. The phylogenetic analyses should also include other Family B ligands and receptors from both vertebrates and invertebrates because it is widely assumed that PDF is related to VIP given their shared roles in circadian clock and gut regulation. Therefore, this analysis needs to be more comprehensive than currently presented. Drosophila melanogaster receptors have also been excluded in spite of the Drosophila PDFR exhibiting ligand promiscuity. The legend should also include the full species names of the various taxa (or modify the figure to include full names) instead of referring to another table. The supplementary table was not available to this reviewer.
Thank you for the reviewer’s constructive comments. According to the reviewer’s suggestion, we have incorporated the VIPRs and Drosophila melanogaster receptors into the comparative analysis and reanalyzed the phylogenies in Figure 1C, and both phylogenies included bootstrap tests (Figure 1B, 1C) in the revised manuscript. The full species names of the various taxa are listed in supplementary tables 1 and 2 in the revised manuscript.
(2) Expression data indicate that AjCTP1/2 is expressed in both the longitudinal muscles and intestine. What are the cell types that express AjCTP1/2? Given that the authors show an effect of CT1 and CT2 on both of these tissues, it would be important to know whether this is local regulation (paracrine or autocrine) vs long-distance hormonal control by the nervous system. This can be addressed by performing in situ hybridization or immunohistochemistry of CT (using Asterias rubens CT antibody: https://doi.org/10.3389/fnins.2018.00382) on these tissues.
Thank you for this feedback. We have now analysed CT-type neuropeptide expression in A. japonicus using immunohistochemistry with the antiserum to the A. rubens CT-type peptde ArCT, which has previously been shown to cross-react with CT-type neuropeptides in other echinoderms (Aleotti et al., 2022). We have added related descriptions in the following sections: Results (section 2.4, line 299-336), Discussion (section 3.3, line 545-554) and Materials and methods (section 4.6, line 785-817). Consistent with this previous finding, the ArCT antiserum labelled neuronal cells and fibers in the central and peripheral nervous system and in the digestive system of A. japonicus (Figure 6). The specificity of immunostaining was confirmed by performing pre-absorption tests with the ArCT antigen peptide (Figure 6-figure supplement 1). The detection of immunostaining in the innervation of the intestine is consistent with PCR results and the relaxing effect of AjCT2 on intestine preparations. Interestingly, no immunostaining was observed in longitudinal muscle, which is inconsistent with the detection of AjCT1/2 transcripts in this tissue. This may reflect differences in the sensitivity of the methods employed to detect transcripts (PCR) and mature peptide (immunohistochemistry). The absence of ArCT-like immunoreactivity in the longitudinal muscles suggests that AjCT1 and AjCT2 may exert relaxing effects on this tissue in vivo via hormonal signaling mechanisms. However, because AjCT1/2 expression in the longitudinal muscles may be below the detection threshold of the ArCT antibodies, we can’t rule out the possibility that AjCT1/2 are released within the longitudinal muscles physiologically.
(3) While Drosophila DH31 can activate both PDF and DH31 receptors, the EC50 values differ drastically. Importantly, there is an independent gene encoding PDF which is a more sensitive ligand for the PDF receptor. This is in stark contrast to the situation presented here where the authors have yet to identify the PDF gene in their system. Outside Drosophila this cross signaling between the two systems has not been observed in any species. Based on this, I would argue that the ability of CTs to activate PDFR is not an evolutionary ancient property but rather an example of convergent evolution if supported by more evidence.
We sincerely appreciate the reviewers' insightful comments. We agree that we cannot rule out the possibilty that ability of CT-type peptides to activate PDF-type receptors in Drosophila and A. japonicus has arisen independently. Therefore, we have modified the text in the discussion accordingly so that this alternative explanation for the effects of CT-type peptides on PDF-type receptors is also presented: “Alternatively, the ability of CT-type neuropeptides to act as ligands for PDF-type receptors in D. melanogaster and A. japonicus may have evolved independently. Further studies on a wider variety of both protostome (e.g. molluscs, annelids) and deuterostome taxa (e.g. other echinoderms, hemichordates) are needed to address this issue.”
(4) AjCT1 and CT2 can activate the two PDF receptors ex vivo. However, their EC50 values are larger and the responses are lower compared to those seen for the CT receptor. Similar cross-talk between closely related peptide families is often observed in ex vivo systems (see: https://doi.org/10.1016/j.bbrc.2010.11.089 , https://doi.org/10.1073/pnas.162276199 , https://doi.org/10.1093/molbev/mst269 and others). However, very few signaling systems exhibit this type of cross-talk in vivo. Without any in vivo evidence, I suspect that the more likely possibility is that the bona fide endogenous ligand for PDF receptors remains to be discovered. The authors could, however, perform peptide and receptor knockdown experiments and show overlap in phenotypes following CT knockdown and PDFR knockdown to support their claim.
We sincerely appreciate the reviewers' insightful critique. According to the reviewer’s suggestion, we have supplemented CTP and AjPDFR2 knockdown experiments, and measured the dry weight of remaining bait and excrement, as well as calculating the weight gain rate and specific growth rate in response to phenotypic changes. The results showed that weight gain rate and specific growth rate in experimental groups were significantly decreased respectively (As shown in Figure 10A and 11B), Correspondingly, except for the I phase, the siAjCTP1/2-1 group had significantly increased remaining bait and decreased excrement in II-VI phases (Figure 10B), the remaining bait weight was significantly increased in siAjPDFR2-1 group (except during phase I), while the weight of excrement was significantly decreased in phase V and VI (Figure 11C). Therefore, AjCT and AjPDFR2 knockdown experiments showed overlap in phenotypes, providing evidence that AjCT does act as an endogenous ligand for PDFR. These results were added in Figure 10 and Figure 11. The related description was added in the results section 2.6 (line 390-396), section 2.7 (line 427-439) and the materials and methods section 4.9 (line 879-898). We acknowledge, however, that other peptides, in addition AjCT1 and AjCT2, may also act as ligands for AjPDFR1 and AjPDFR2 in vivo and on-going studies in the Chen (OUC) and Elphick (QMUL) labs are attempting to address this issue
(5) Why are receptor transcripts upregulated following peptide injection? Usually, increased ligand levels/signaling result in a compensatory decrease in receptor levels. These negative feedback loops maintain optimum signaling levels. Since the authors have successfully implemented RNAi for this CT precursor, what are the phenotypes on growth and feeding?
We thank the reviewers for raising these critical points. Our responses are structured as follows: Firstly, our findings align with established mechanisms of neuropeptide-induced receptor modulation (Please check the reference Tiptanavattana et al. 2022). Secondly, based on the reviewer’s suggestion, we have supplemented the experiments to detect the phenotype variations on growth and feeding based on long-term reduced CT signaling, including measuring the dry weight of remaining bait and excrement, calculating the weight gain rate and specific growth rate, as well as testing the expression levels of the three growth factors (AjMegf6, AjGDF-8 and AjIgf). The results showed that weight gain rate and specific growth rate in siAjCTP1/2-1 group were significantly decreased (As shown in Figure 10A), Correspondingly, except for the I phase, the siAjCTP1/2-1 group had more remaining bait and less excrement in II-VI phases (Figure 10B). Furthermore, the growth inhibitory factor AjGDF-8 was significantly up-regulated and the growth promoting factors AjMegf6 were significantly down-regulated in siAjCTP1/2-1 group (Figure 10C). We have added these results in Figure 10, with detailed description in the results section 2.6 (line 390-396) and in the materials and methods section 4.9 (line 879-888). And after long-term continuous injections of siAjCTP1/2-1, we further recorded the feeding behavior of these sea cucumbers for three consecutive days. The remaining bait and feces were cleaned and the food was re-placed in the middle of the tank each day. We calculated the aggregation percentage (AP) of sea cucumbers around the food during the peak feeding period (2:00-4:00) each day, which is the best indicator for sea cucumber feeding behavior detecting. The results showed that the AP in siAjCTP1/2-1 group was significantly lower than that in control group. After dissection, we also found the intestines of siAjCTP1/2-1 group had less food and significantly degenerated (see author response image 1). All these results supported that long-term functional loss of AjCT2 negatively influence the feeding and growth of A. japonicus.
Other comments:
(6) What criteria do the authors use to classify some proteins as "type", some as "like" and others as "related"? In my opinion, DH31 could be referred to as CT-like or CT-type. Please use one term for clarity unless there is a scientific explanation behind this terminology.
Thank you for the reviewer’s comment. If you look at the paper by Cai et al. (2018) you will see in Figure 14 that CT-type peptides and DH31-type peptides are paralogous, probably due to a gene duplication in the common ancestor of the protostomes. The CT-related peptides in protostomes that have a disulphide bridge we would describe as CT-type because they have conserved a feature that is found in CT-type peptides in deuterostomes. Whereas the DH31 peptides we would describe as CT-like. But there is not a formal rule on this. It is possible the duplication event that gave rise to DH31 and CT-type peptides occurred in the common ancestor of the Bilateria but DH31-type signaling was lost in deuterostomes. On the other hand, if the gene duplication that gave rise to DH31-type peptides and CT-type peptides in protostomes did occur in a common ancestor of the protostomes, then DH31 and CT-type peptides in protostomes could be described as co-orthologs of CT-type peptides in deuterostomes. In this case, both CT peptides and DH31 peptides in protostomes could be described as CT-type. Here is a useful link for explanation of terms: https://omabrowser.org/oma/type/
(7) Was genomic DNA removal step performed before cDNA synthesis for qRT-PCR?
Thank you for the reviewer’s comment. The genomic DNA removal step was performed before cDNA synthesis for qRT-PCR and we have added the information in the section 4.5 (line 774-776).
(8) Line 70: The presence of calcitonin-like peptides (DH31) and DH31 receptors in invertebrates was discovered long before the discoveries by Jekely 2013 and Mirabeau and Joly 2013. Please credit these original studies: https://pubmed.ncbi.nlm.nih.gov/10841553/ and https://pubmed.ncbi.nlm.nih.gov/15781884/.
Thank you for the reviewer’s comment. We have credited these original studies in the revised manuscript.
(9) Lines 72-74: Please cite https://pubmed.ncbi.nlm.nih.gov/24359412/.
Thank you for the reviewer’s comment. We have cited it in the revised manuscript.
(10) Line 87: Please cite https://pubmed.ncbi.nlm.nih.gov/15781884/.
Thank you for the reviewer’s comment. We have cited it in the revised manuscript.
(11) Lines 89-91: The functional significance of DH31 signalling to PDFR in Drosophila is known. See: https://pubmed.ncbi.nlm.nih.gov/15781884/ and https://pubmed.ncbi.nlm.nih.gov/30696873/. There are several studies that have shown the functions of DH31 signalling via DH31R.
Thank you for the reviewer’s comment. We have corrected it and added all this studies in the revised manuscript.
(12) Figure 1 Supplement 1: The tertiary models for CT1 and CT2 look completely different. This prediction is not in line with both ligands activating the same receptor.
Thank you for the reviewer’s comment. We have deleted this supplementary figure.
(13) Figure 1 Supplement 3 legend: Please add panel labels next to the corresponding receptor.
Thank you for the reviewer’s comment. We have added panel labels next to the corresponding receptors as you suggested.
(14) Figure 2: What does CO refer to?
Thank you for the reviewer’s comment. CO (Control) refers to the stimulation of HEK293T transfected cells with serum-free DMEM, and we have added the detailed information in Figure 2 legend (line 251-252).
(15) Figure 3: Due to the low magnification of the cells, it is difficult to see the localization of the receptor. It would also be more appropriate to use a membrane marker rather than DAPI which does not label the cytoplasm or membrane where the receptor can be found.
we appreciate the reviewer's insightful comment regarding the experimental controls. The baseline receptor localization data under non-stimulated conditions are presented in Figure 3—figure supplement 1, demonstrating constitutive membrane distribution of pEGFP-N1-tagged receptors. Upon stimulation with synthetic CT-like peptides, qualitative imaging analysis revealed significant ligand-induced receptor internalization into the cytoplasm (Figure 3B).
(16) Figure 9: Please include PDF precursor and receptor as separate columns. Also, Drosophila CT/DH31 receptors have been characterized.
Thank you for the reviewer’s comment. We have added PDF precursor, predicted peptides and receptors as separate columns in the revised manuscript Figure 12. And also, we corrected the error summary of Drosophila CT/DH31 receptors according to your suggestions.
(17) Table 1: It is not very clear why there are multiple columns for ERK1/2 with different outcomes.
Thank you for the reviewer’s comment. Although the cAMP/PKA or Gαq/Ca2+/PKC signaling is activated after ligand binding to receptors, the downstream ERK1/2 cascade is not necessarily activated. Therefore, we counted the activation status of cAMP/PKA and its downstream ERK1/2 cascade, and Gαq/Ca2+/PKC and its downstream cascade in Table 1 respectively. We have optimized Table1 to make it clearer in the revised manuscript.



