Abstract
Dopamine release in the ventromedial striatum (VMS) both invigorates actions and encodes reward-related information, yet how these functions are integrated remains under active debate. To investigate this further, we designed four different versions of a rat Go/No-go task, where we systematically manipulated response requirements, temporal task demands, and controllability of reward pursuit. Dopamine release increased reliably during action initiation (Go) but was delayed during action suppression (No-go), and was insensitive to augmented response demands or controllability. Following response completion, dopamine rose gradually until animals arrived at the reward location, irrespective of reward-delivery timing, prior action demands, or controllability. This proximity dopamine-signal was exaggerated after animals exhibited Pavlovian consummatory behavior during No-go trials, revealing a motivational signal component. Together, these findings indicate that in reward contexts, VMS-dopamine signals successively integrate the invigoration of action initiation with the continuous estimation of spatial – but not temporal – proximity to rewards.
Introduction
Appropriate, situation-dependent behavior in everyday life requires both motor control and information about upcoming rewards. A key neuromodulator involved in these processes is dopamine. Within the striatum, the cardinal projection target of midbrain dopaminergic neurons, it is thought that dopamine release encodes a reward-prediction error (RPE) that signals the discrepancy between expected and obtained outcomes in a given situation (Schultz et al., 1997). This teaching signal supports goal-directed behavior (Dickinson and Balleine, 1994), and invigorates actions toward appetitive rewards (Coddington and Dudman, 2018; da Silva et al., 2018; Howe et al., 2013; Howe and Dombeck, 2016; Nicola, 2007; Panigrahi et al., 2015). Although these functions have been widely studied independently in the ventromedial striatum (VMS), how motivation and movement are integrated in the VMS remains unclear.
While the role of VMS dopamine in signaling reward-related information is well established (Bayer and Glimcher, 2005; Schultz et al., 1997; Starkweather and Uchida, 2021), its contribution to the volition of reward pursuit is less understood. Recent studies have shown that striatal dopamine differentially responds to reward contingencies across instrumental-and Pavlovian-conditioning tasks (Goedhoop et al., 2023; Hamid et al., 2021). Furthermore, midbrain single-neuron activity differs when actions for rewards are self– or cue-initiated (Romo and Schultz, 1990). Together, this indicates that reward-related VMS dopamine signals may be modulated by the controllability of reward seeking.
Various studies have demonstrated a gradual increase in VMS dopamine release as animals approach a known reward location (Hamid et al., 2016; Howe et al., 2013; Kim et al., 2020; Mohebi et al., 2019). This gradual increase scales with spatial proximity to distant rewards (Howe et al., 2013; Kim et al., 2020). This prolonged signal has been suggested to sustain motivational drive for rewards (Howe et al., 2013), reinforcing preceding actions that resulted in reward (Hamid et al., 2016; Niv and Schoenbaum, 2008). However, it is not known whether this gradual increase is modulated by the controllability to pursue rewards, or whether the signal remains consistent across different action requirements for rewards.
Here, we developed a series of Go/No-go task variants to elucidate how VMS dopamine integrates action selection, controllability of reward pursuit, and reward approach. Rats were trained to initiate (Go) or suppress (No-go) actions for rewards in response to an action-cue, and trial starts were either self– or cue-initiated. In another task variant, we added ‘Free’ trials where no overt action was required, increased trial demands, and delayed reward delivery, to test the effects of effort and separate signals associated with action initiation and reward approach. We hypothesized that dopamine signaling dynamics during action execution would differ between self-initiated and cue-initiated task variants, but that the gradual increase in VMS dopamine during reward approach would generalize across contingencies.
By combining real-time measurements of dopamine release together with behavioral tracking, we replicate previous findings showing the importance of movement for the expression of reward-related dopamine dynamics (Syed et al., 2016). We additionally demonstrate that specifically the approach to the reward location, regardless of the preceding required action, increases VMS dopamine release that peaks upon magazine arrival. Furthermore, we show that dopamine-release amplitude during action did not differ between varying strategies of action suppression, but instead strongly differed during reward approach.
Results
Success rates in Self- and Cue-initiated Go/No-go(/Free) task variants
To investigate the role of VMS dopamine in reward processing and motor control, we implanted rats with fast-scan cyclic voltammetry (FSCV) microelectrodes (n = 27; Fig. 2a) and trained them in four variants of the task: 1) a “short-task variant”, in which rats Self-initiated trials, 2) a “short-task variant” in which trials were Cue-initiated (altered controllability of reward pursuit), 3) a “long-task variant” of the Self-initiated task, in which the required duration of rat response action was increased (altered difficulty), and 4) a “long-task variant” of the Cue-initiated task. There were two trial types in the short-task variants of the task: depending on the (auditory) action cue, rats were required to press a lever (Go) or nose poke (No-go) to obtain reward. In the long-task variants, we introduced a third trial type: in addition to Go and No-go action-cues and corresponding required responses, a third (auditory) cue signaled that reward would be delivered without requiring an overt action (Free trials).
Self-initiated Go/No-go (“short task”)
In the Self-initiated Go/No-go task, similarly used previously by Syed et al. 2016), trials were initiated by an (uncued) nose poke (Fig. 1a,b). Our task design ensured that the time of reward delivery following action-cue onset was matched across trial types. After animals achieved proficiency in the task (see Methods), we measured behavioral performance and real-time dopamine release. Animals could initiate a trial by entering the nose-poke port after the intertrial interval (ITI); importantly, rats spent <20% of the ITI time inside the nose-poke port (Supp. Fig. 1), thus in between trials they mostly left the port. Reward success rates were significantly higher when animals had to initiate actions (Go trials) as compared to trials where the animals had to suppress actions (No-go trials) (Fig. 1c; Wilcoxon matched-pairs test, W = 35, p = 0.039).
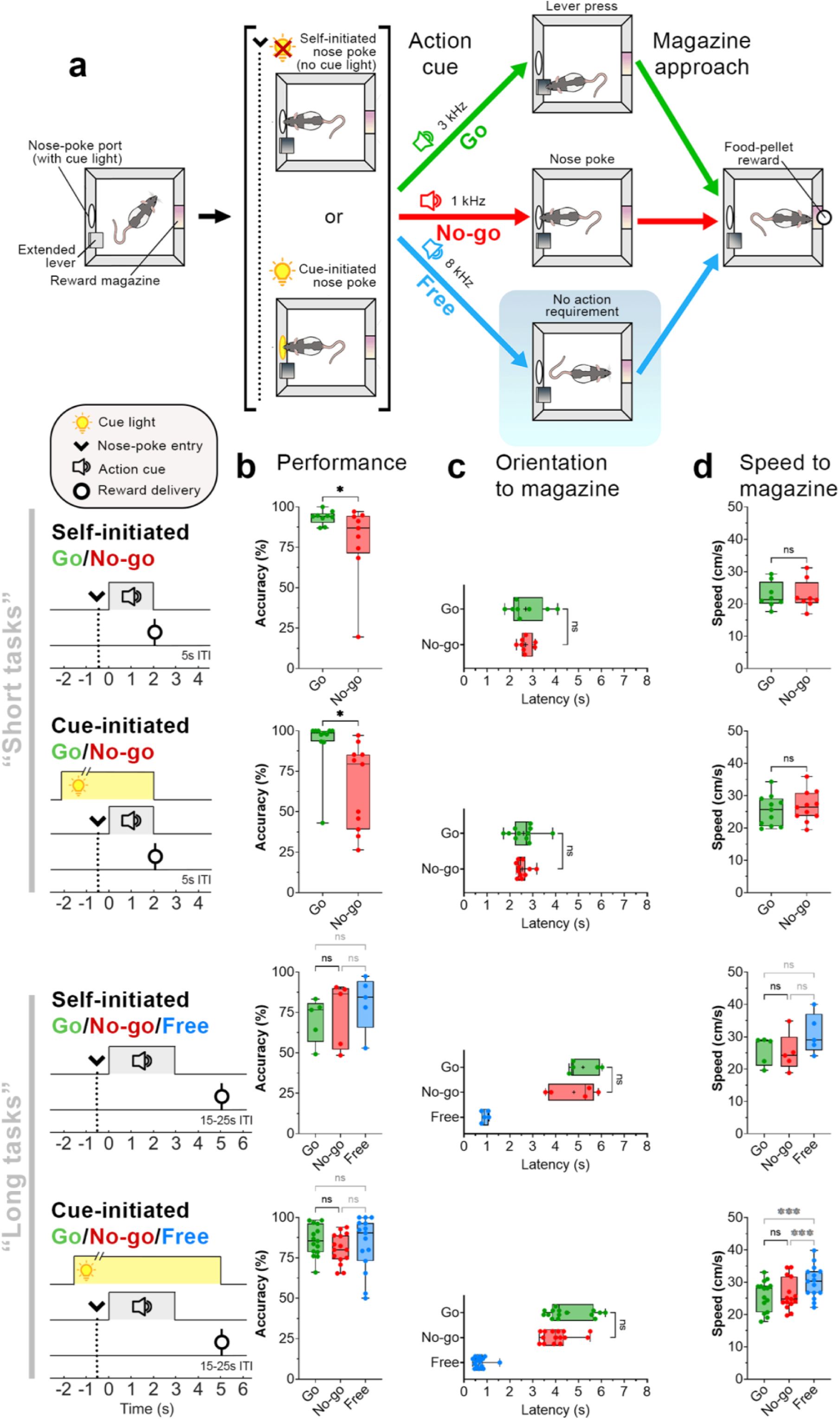
Go and No-go performance is similar across task variants.
a) Schematic of the Go/No-go (short) behavioral task. Rats initiated trials by entering a nose-poke port (arrow, dotted line), either voluntarily (Self-initiated) or following nose-poke light illumination (Cue-initiated). After staying in the nose-poke port for 0.5s, an auditory stimulus (action cue) instructed rats to either initiate action (lever presses: ‘Go’, green) or remain in the nose-poke port (‘No-go’, red). In the Go/No-go/Free (long) task, an additional action cue was presented that did not require an action for rewards (‘Free’, blue). Following Free cue onset, rats could approach the reward magazine immediately. b) Schematic of task timeline. For all trials, reward-delivery latency (relative to action cue onset) was similar. c) Success rates across Go/No-go task variants. d) Latency to orient toward the reward magazine following action cue-onset. e) Speed of movement toward the reward magazine after orientation. Significance: * p < 0.05, ** p < 0.01, *** p < 0.001, Wilcoxon matched-pairs signed rank or post hoc Dunn’s tests.
Cue-initiated Go/No-go (“short task”)
The activity of midbrain dopaminergic neuronal activity differs depending on the controllability of reward pursuit (Romo and Schultz, 1990). To evaluate how VMS dopamine dynamics are affected by diminished control over trial initiation, we additionally trained our rats on a Cue-initiated task variant in which trials could only be initiated by a cued nose poke (Fig 1a,b; see Methods): rats promptly responded to the cue (nose-poke light illumination) to initiate trials (1.68 ± 0.28 s; Supp. Fig. 2), indicating that they understood the contingency. Similar to the Self-initiated variant, rats trained in the Cue-initiated task showed significantly higher success rates on Go trials as compared to No-go trials (Fig. 1b, Cue-initiated Go/No-go; W = 54, p = 0.014).
Self– and Cue-initiated Go/No-go/Free (“long task”)
Previous research has shown that spatial proximity of subjects relative to the reward location is reflected in VMS dopamine concentration, and dopamine responses may differ depending on whether rewards are contingent on a subject’s behavior (i.e., Pavlovian vs. instrumental responding for rewards; Goedhoop et al., 2023; Hamid et al., 2021). Furthermore, VMS dopamine may be influenced by effort (Walton and Bouret, 2019). To investigate the influence of these factors, we trained rats on the long-task variants that included ‘Free’ trials and increased Go and No-go trial requirements (Go: 3s repeated pressing, 6.52 ± 0.32 presses, vs. short 2 presses, 1.80 ± 0.03s; No-go: 3s vs. short 2s nose-poke hold; see Methods, Fig. 1a). Reward-delivery timing in Free trials matched the timing of long-task Go and No-go trials (Fig. 1a). In this Cue-initiated variant, rats promptly responded to the cue (0.95 ± 0.04 s, Supp. Fig. 1), indicating that they understood the contingency. Success rates were not significantly different across Go/No-go/Free trial types for both Self-initiated (Fig. 1b; Kruskal-Wallis test H(2) = 0.400, p = 0.954) and Cue-initiated (H(2) = 1.458, p = 0.482) variants.
Reward approach behaviors
To determine whether action initiation (Go) and suppression (No-go) affected motivational drive to approach the reward location, we tracked behavioral performance using DeepLabCut (Mathis et al., 2018). In the short Self- and Cue-initiated Go/No-go task variants, the latency to orient to the reward magazine (Fig. 1c; Self-initiated: W = 0, p > 0.999; Cue-initiated: W = 6, p = 0.831), approach speed to the magazine (Fig. 1d; Self-initiated: W = –5, p = 0.773, Cue-initiated: W = –28, p = 0.240), and latency to arrive at the magazine after action-cue onset (Supp. Fig. 3; Self-initiated: W = –4, p = 0.844; Cue-initiated: W = 2, p = 0.966) did not differ between Go/No-go trial types.
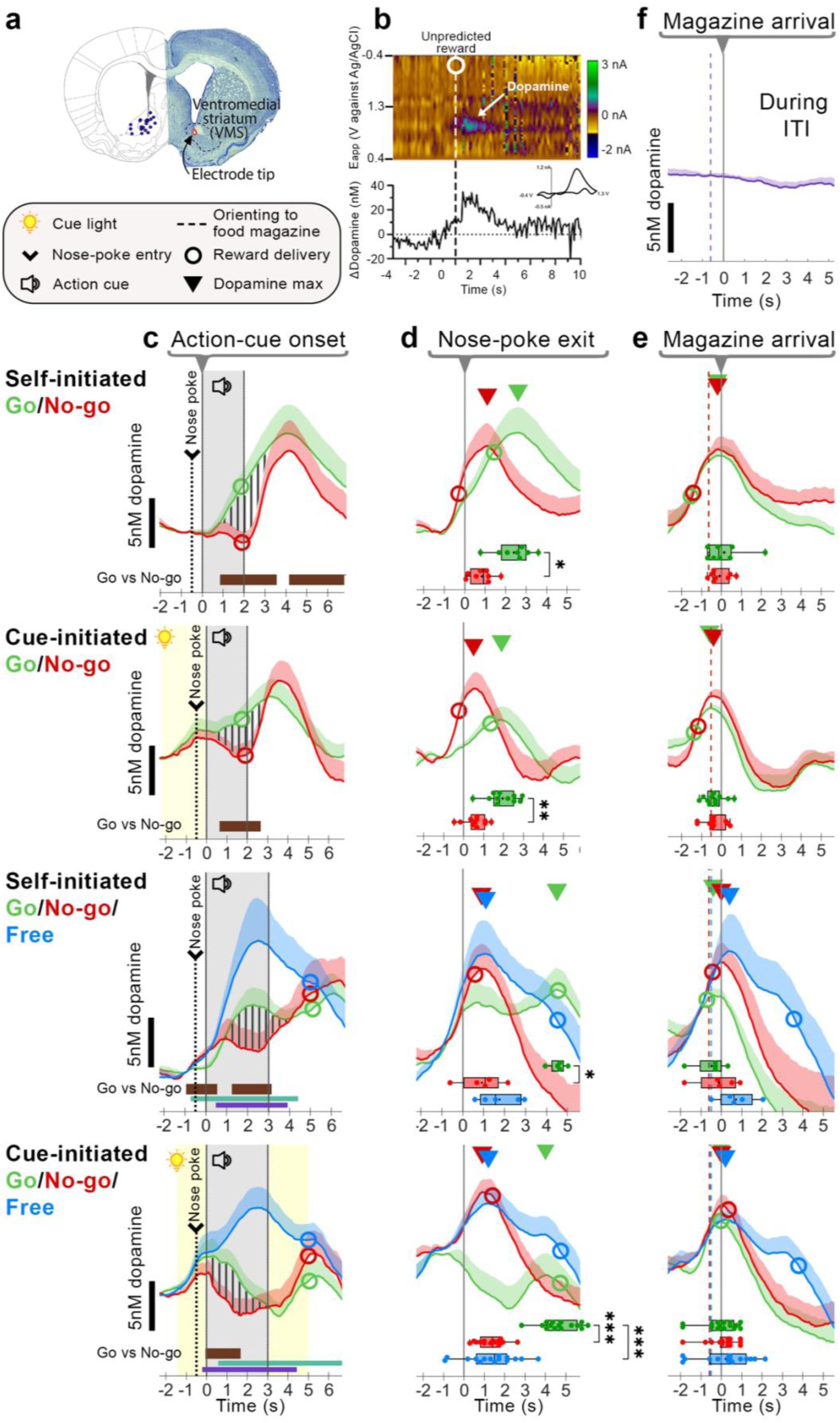
VMS dopamine encodes both action initiation and reward-magazine arrival irrespective of action contingency.
a) Histological verification of electrode placements in VMS (n = 27). b) Representative color plot and corresponding dopamine trace in response to an unpredicted pellet. The dotted line and circle indicate pellet delivery. Inset: Accompanying cyclic voltammogram confirms dopamine detection. c-e) Average dopamine concentration (nM; mean + SEM) for Go (green), No-go (red), and free (blue) trials. Triangles denote the average latency of peak dopamine relative to different alignment points. c) Traces aligned to action-cue onset. Vertical stripes highlight differences in Go vs. No-go dopamine. Horizontal bars indicate significant within-subjects differences between trial types (Brown: Go vs. No-go, Aqua: Free vs. Go, Purple: Free vs. No-go). d) Traces aligned to nose-poke exit; e) magazine arrival. Box plots depict latency of peak dopamine release (± 2s) around magazine arrival per animal. f) Dopamine release during the inter-trial interval (ITI) as rats approached the reward magazine from the opposite wall, aligned to magazine arrival. Boxplot statistical significance: * p < 0.05, ** p < 0.01, *** p < 0.001, Wilcoxon matched-pairs signed rank or post-hoc Dunn’s tests.

Classified No-go behavior reveals VMS dopamine release following action-cue offset in ‘Biting’ trials.
a) Left: Agglomerative hierarchical clustering of No-go trial behaviors using Ward’s linkage and Euclidean distance. A three-cluster solution (dotted grey line) was selected resulting in a balanced distribution of trials across clusters and sessions. These clusters corresponded to three distinct behavioral strategies. ‘Biting’ – defined as biting of the nose-poke port wall; ‘Digging’ – digging movements in the nose-poke hole; ‘Calm’ – all other no-go trials. b) A supervised decision-tree classifier labeled trials to one of three behavioral groups: Digging, Biting, and Calm. Average dopamine concentration of each classified group is depicted (nM; mean + SEM). c) Maximum dopamine concentration (nM) following action-cue offset (+2s) was highest in Biting trials as compared to other No-go trials. ‘After action-cue offset: No-go (trials)’: Individual No-go trials classified by No-go behavior. Solid lines depict the median. ‘After action-cue offset: No-go (rats)’ and ‘Unpredicted pellets (rats)’: Rats classified based on their predominant No-go strategy. Solid lines depict the mean. Significance: * p < 0.05, ** p < 0.01, *** p < 0.001, post hoc Dunn’s tests.
To assess whether altered controllability of reward pursuit, increased task effort, and the addition of Free trials or affected the core comparison between action initiation (Go) and suppression (No-go), we examined behavioral performance in the Go/No-go/Free task variants. Trial type significantly influenced latency to orient to the reward magazine in both Go/No-go/Free task variants (Fig. 1c; Self-initiated: H(2) = 7.600, p = 0.024; Cue-initiated: H(2) = 24.13, p < 0.001). Post hoc Dunn’s test revealed orienting latencies did not significantly differ between Go and No-go trials (all p > 0.05, see Fig. 1c), but that Free trials significantly differed from Go and No-go trials (Self-initiated Go vs. Free: p = 0.034; Cue-initiated: p < 0.001, No-go vs. Free: p = 0.002), which is consistent with Free trials requiring no action as compared to Go (action initiation) and No-go trials (action suppression). However, although the speed to approach the reward magazine was significantly different for the Cue-initiated Go/No-go/Free task (Fig. 1d; H(2) = 19.73, p < 0.001), but not for the Self-initiated task (H(2) = 2.800, p = 0.367), post hoc Dunn’s test revealed that rats moved at similar speeds between Go and No-go trials, but were significantly faster on Free trials as compared to Go and No-go trials (see Fig. 1d; p < 0.01), suggestive of increased vigor for Free trials despite matched reward delivery times. In addition, although the latency to arrive at the reward magazine was significantly different between trial types for Self- (H(2) = 7.600, p = 0.024) and Cue-initiated Go/No-go/Free tasks (H(2) = 22.80, p < 0.001), post hoc Dunn’s test revealed no difference in arrival latencies between Go and No-go trials, but earlier arrival for Free trials (Supp. Fig. 3). Together, these results indicate that Free trials were associated with earlier approach to the reward location, but the motivational drive to approach was comparable between Go and No-go trials, and is not affected by reward controllability (self– vs. cue-initiated) nor increasing task effort (short vs. long-task variant).
VMS dopamine release encodes movement and reward-related information
VMS dopamine recordings (Fig. 2a,b) showed a rapid increase following presentation of the Go-action cue but no equivalent increase in response to the No-go-action cue (Fig. 2c): Within-subject time-series comparisons(Jean-Richard-dit-Bressel et al., 2022) of Go and No-go trials (Fig. 2c, ‘Self-initiated Go/No-go’, vertically striped area) confirmed significant differences (p < 0.05; Fig. 2c, thick horizontal bar insets). Despite differences in task requirements such as controllability to attain reward (i.e., Self- vs. Cue-initiated) and increased effort for Go and No-go trials in the Go/No-go/Free task (i.e., long-task variant, see Methods, Fig. 1a), within-subject comparisons for all task variants showed a consistent, significant increase in dopamine release on Go trials compared to No-go trials (p < 0.05; Fig. 2c Self- and Cue-initiated Go/No-go/Free task, thick horizontal bar insets). In addition, dopamine release was highest for Free trials as compared to Go and No-go trials (p < 0.05; Fig. 2c thin horizontal bar insets). Our data suggest that the increase in VMS dopamine release not only reflects reward-predictive cues but also action initiation. In addition, this difference is unaffected by controllability of reward pursuit and increased trial demands.
Since the rise in dopamine release for Go trials occurs before rises in No-go trials (when traces were aligned to action-cue onset), we sought to clarify whether this difference may be due to an earlier moment of action initiation (nose-poke exit) for Go trials as compared to No-go trials. Therefore, we aligned traces to the time when animals departed the nose-poke port (Fig. 2d). Although dopamine release after nose-poke port-exit for Go and No-go trials rapidly increased for the Self-initiated Go/No-go task, this was less obvious for the other task variants (Fig. 2d). Analysis of peak dopamine release (Fig. 2d, triangles) differed between trial types: the increase in dopamine release during Go trials did not align with nose-poke port exit (Fig. 2d). Furthermore, the latency to peak dopamine was significantly different between trials, where peak dopamine release was consistently delayed for Go trials (Fig. 2d, boxplot insets; Self-initiated Go/No-go: W = 34, p = 0.016; Cue-initiated Go/No-go: W = 64, p = 0.002; Self-initiated Go/No-go/Free: H(2) = 8.40, p = 0.008; Cue-initiated Go/No-go/Free: H(2) = 23.33, p < 0.001). Overall, our data suggests that differences in Go vs. No-go dopamine cannot be attributed to merely the moment of action initiation (nose-poke departure).
Previous research has shown that VMS dopamine can not only encode RPE and movement, but can also reflect a subject’s proximity to the expected reward location (Hamid et al., 2016; Howe et al., 2013; Mohebi et al., 2019). We therefore realigned traces to the moment animals arrived at the reward magazine (Fig. 2e). Across all trial types and sessions, dopamine release consistently peaked around the time of magazine arrival (Fig. 2e, insets; Self-initiated Go/No-go: W = 7, p = 0.672; Cue-initiated Go/No-go: W = 3, p = 0.916; Self-initiated Go/No-go/Free: H(2) = 4.526, p = 0.111; Cue-initiated Go/No-go/Free: H(2) = 4.204, p < 0.122). Importantly, the timing of peak dopamine (Fig. 2e, Self- and Cue-initiated Go/No-go/Free, triangles) occurred independently of reward delivery: in Free trials, average reward delivery latency (Fig. 2e, hollow circles) was delayed following magazine arrival by up to 3s. In addition, rats oriented to the magazine at similar times, across trial-types and task variants, relative to magazine arrival (Fig. 2e; on average 0.6s from orientation to magazine arrival; vertical dashed line). To determine whether peak dopamine reflected any approach to the reward magazine, or required information about an upcoming reward (e.g., action cues), we analyzed changes in dopamine release during unprompted magazine entries in the Self-initiated Go/No-go/Free task (Fig. 2f, During ITI). Our findings show that unprompted magazine entries during the ITI did not elicit changes in VMS dopamine release; this suggests that increases in VMS dopamine require task-relevant features such as knowledge of an upcoming reward and movement towards the reward magazine, where VMS dopamine peaks upon arrival at the expected reward location.
Classified No-go behaviors reveal differences in dopamine signaling (365)
During No-go trials, rats had to suppress actions by remaining in the nose-poke port. However, we observed individual differences in behavioral strategies in performing ‘action suppression’. We therefore classified No-go trials into three behavioral strategies (Biting (purple), Digging (teal), Calm (maroon); Fig. 3a) using a decision-tree classifier based on No-go behavior (Supp. Fig. 4). Dopamine concentrations during the action-cue period revealed no differences between classifications (Fig. 3b, Supp. Fig. 5). This result indicates that behavioral strategy during No-go trials is unrelated to dopamine release during the action-cue period, as they demonstrate similar dopamine release for ‘action suppression’ despite varying No-go behavioral strategies.
However, maximum dopamine concentration measured after action-cue offset (Fig. 3c) significantly differed between strategies for No-go trials in all task variants (Self-initiated Go/No-go: H(2) = 34.15, p < 0.001; Cue-initiated Go/No-go: H(2) = 16.24, p < 0.001; Self-initiated Go/No-go/Free: H(2) = 22.54, p < 0.001; Cue-initiated Go/No-go/Free: H(2) = 37.48, p < 0.001). Post hoc comparisons showed that dopamine levels were consistently higher for Biting as compared to Calm trials (Fig. 3c, ‘After action-cue offset: No-go trials’, all p < 0.001). Dopamine was also higher for Biting as compared to Digging in two task types (Fig. 3c). These findings suggest that while dopamine release during the action-cue period is unaffected by action-suppression strategy, differences emerged during reward approach and may reflect differences in motivational drive.
Regrouping animals based on their predominant No-go behavioral strategy revealed a trend in peak dopamine that resembled individual trial data (Fig. 3c, ‘No-go (rats)‘ and ‘No-go (trials)‘, respectively). Therefore, we grouped animals based on their predominant action-suppression strategy to determine whether Biting animals also showed elevated dopamine in response to other reward-related information. However, peak dopamine release in response to an unpredicted pellet did not follow this pattern and instead, they were similar across all groups (Fig. 3c, ‘Unpredicted pellets (rats)’). Comparably, grouped rats did not show differences in peak dopamine for Go and Free trials following action-cue onset (Supp. Fig. 5). These results suggest that the elevated dopamine response during reward approach in Biting animals is specific to No-go trials.
Discussion
We examined how VMS dopamine integrates motivation and reward-related information during action initiation and suppression in four Go/No-go task variants. In our task, action initiation occurred at two distinct points after trial start: 1) when rats began lever pressing (Go), and 2) when rats walked to the reward magazine, either without action requirement (Free) or after successful trial completion (Go and No-go). By contrast, action suppression was unique to No-go trials, where animals were required to stay in the nose-poke port. Dopamine release during action initiation (Go) was consistently higher than during action suppression (No-go); this difference was unaffected by controllability of reward pursuit (Self-vs. Cue-initiated). Additionally, VMS dopamine signaled spatial proximity to the reward, but was insensitive to the timing of reward delivery, controllability, and the preceding action requirement. These findings indicate that in a reward context, VMS dopamine is not only associated with action execution but continuously tracks spatial, but not temporal, proximity to rewards. Unexpectedly, consummatory Pavlovian behavior in the form of (wall) biting during action suppression led to markedly larger dopamine release during subsequent reward-magazine approach than other behaviors. Together, our findings indicate that VMS dopamine sequentially integrates reward-driven action initiation with the gradual increase in spatial reward proximity.
Dopamine reflects action initiation but not controllability or effort
VMS dopamine has been associated with the invigoration and initiation of reward-seeking behavior (Ikemoto and Panksepp, 1999; Nicola, 2007) as demonstrated through both bulk measurements of VMS dopamine release (Collins et al., 2016; du Hoffmann and Nicola, 2014; Hamid et al., 2016; Mohebi et al., 2019; Nicola, 2010; Phillips et al., 2003; Roitman et al., 2004; Syed et al., 2016; Wassum et al., 2012) and the activity patterns of individual midbrain dopamine neurons (Coddington and Dudman, 2018; Jin and Costa, 2010). Across all four task variants, we found that VMS dopamine release was consistently higher during action initiation (Go) than during action suppression (No-go). Thus, by extending the pioneering work by Syed and colleagues (Syed et al., 2016), our findings provide compelling evidence that striatal dopamine release encodes reward-driven action initiation.
In addition to action initiation, recent work suggests that striatal dopamine dynamics are shaped by the controllability of reward contingencies (Goedhoop et al., 2023; Hamid et al., 2021). For instance, the anticipation of operant action initiation during reward-cue presentation is accompanied by more sustained VMS dopamine release compared to cues that signal reward delivery without action requirement (Goedhoop et al., 2023). Similarly, early midbrain dopamine recordings showed increased responses to externally triggered actions (i.e., Cue-initiated) than to Self-initiated actions for rewards (Romo and Schultz, 1990). In contrast, our data indicate that manipulations of controllability in reward pursuit (i.e., Self- vs. Cue-initiated trials) did not affect the contrast between Go and No-go dopamine dynamics during action execution, but instead only shifted dopamine release to an earlier time point in the trial, presumably triggering earlier positive RPEs in Cue-initiated trials compared to Self-initiated trials (Fig. 2a). However, we cannot exclude the possibility that dopamine would respond to a more extreme manipulation of controllability, since animals retained some volitional control over when to pursue rewards in all our task versions – either by periodically checking the nose-poke port in the Self-initiated task, or by deciding when to respond to nose-poke illumination in Cue-initiated tasks. Thus, our results indicate a prominent role for VMS dopamine in reward-related action (Coddington and Dudman, 2018; Ikemoto and Panksepp, 1999; Nicola, 2007), but less sensitivity for discriminating between voluntary and externally cued action.
Previous studies have shown that dopamine signals are influenced by the effort required to obtain rewards(Walton and Bouret, 2019). We tested this by varying response demands: short and long Go trials requiring either two lever presses or repeated pressing for three seconds, respectively. No-go trials required matching two or three seconds of action suppression, respectively (Fig 1b). However, such effort manipulations had no detectable impact on action-related dopamine: the relative contrast in VMS dopamine in Go versus No-go trials did not differ between short and long task variants, suggesting that bulk VMS dopamine does not scale with response duration or physical effort, consistent with some previous work(Gan et al., 2010).
Dopamine release reflects spatial proximity
Beyond physical effort, the long-task variants imposed a two-second reward delay after action-cue offset, permitting clearer separation of dopamine signaling during action-cue and reward-approach epochs. Critically, peak dopamine responses across all four task variants, could not be explained solely by action initiation, as the latency to peak dopamine differed between trial types when aligned to nose-poke exit (Fig. 2d, triangles), diverging from previous reports(Syed et al., 2016). Instead, dopamine release during the long-task variant revealed a gradual increase and consistent peak upon arrival at the reward magazine, irrespective of whether rewards were received immediately or were delayed (Fig. 2d). Importantly, we did not observe an increase in VMS dopamine when animals approached and checked the magazine between trials, indicating the necessity of reward expectation linked to trial performance (Fig. 2e). In addition, the signal cannot be attributed to orientation to the reward-magazine – if orientation were the cause, dopamine would not be expected to increase while animals maintain a constant orientation when moving toward the reward magazine. Overall, this finding is consistent with several studies reporting a gradual rise in VMS dopamine, frequently referred to as ramps, as animals actively (Farrell et al., 2022; Hamid et al., 2016; Howe et al., 2013; Krausz et al., 2023; Mohebi et al., 2019) or passively (Farrell et al., 2022; Guru et al., 2020; Kim et al., 2020; Mikhael et al., 2022) move toward reward. This phenomenon was particularly pronounced during Free trials, where dopamine peaked when rats arrived at the reward magazine up to five seconds before reward delivery (Fig. 2c,d Go/No-go/Free, circles). Together, our data suggest that while VMS dopamine initially facilitates the initiation of reward-driven action, it subsequently tracks spatial reward information and may serve to sustain motivational drive during reward approach.
The gradual increase in VMS dopamine during reward approach, peaking upon arrival, has been suggested to reflect ongoing state estimation under uncertainty (Farrell et al., 2022; Gershman and Uchida, 2019; Kim et al., 2020; Mikhael et al., 2022). Alternatively, this gradual increase, or ramp, has been suggested to reflect an ‘internal model’ that continuously updates reward-proximity estimates as animals progress toward the goal (Farrell et al., 2022; Guru et al., 2020). Our findings support the proximity account. In our Go/No-go task, rats traversed the same familiar, relatively short path to the magazine in thousands of trials, minimizing state uncertainty and reducing the need for ongoing state estimation updates. Instead, our findings suggest that VMS dopamine maintained a continuous estimate of spatial reward proximity: arrival at the reward magazine following trial completion served as the most reliable predictor of reward and coincided with peak dopamine release, consistent with previous findings (Hamid et al., 2016; Howe et al., 2013; Mohebi et al., 2019). While our results clearly demonstrate dopamine signaling related to spatial proximity to rewards, our experimental design cannot definitively distinguish whether VMS dopamine encodes RPE computations or spatial proximity to sustain motivated approach behavior. More modeling and experimental work is needed to test these predictions (but see Gershman et al., 2024; Lerner et al., 2021)).
Dopamine dynamics are linked to motivated action
Our findings reveal that VMS dopamine follows a consistent temporal pattern across trials: dopamine increases when animals initiate reward-directed actions, then continuously tracks spatial reward proximity. Critically, the magnitude of this proximity signal was modulated by the following trial-type-specific behavioral indicators of motivational engagement, and was particularly evident in Go/No-go/Free task versions: animals could perform lever presses (Go trials) and nose-poke hold times (No-go trials) up to the end of the action-cue epoch, or beyond until delayed reward delivery (Fig. 1a). Animals that completed lever-press requirements faster, exhibited an earlier magazine approach and a concomitant increase in dopamine, whereas slower completion was associated with a delayed dopamine (Supp. Fig 7). We identified a distinct behavioral phenotype in No-go trials: consummatory Biting responses directed at the wall around the nose-poke device during action suppression were associated with shorter nose-poke hold latencies (Supp. Fig. 6) and an approximately two-fold increase in dopamine during subsequent reward approach, as compared to non-biting strategies across all task variants (Fig. 3b). Notably, the increased dopamine signal associated with Biting was specific to reward approach following action-suppression, as there was no difference in dopamine related to other events (e.g., unpredicted pellet delivery; Fig. 3c) or other trial types (Supp. Fig. 5) using this classification.
Although Biting responses, which resembled consummatory behaviors typically directed toward rewards, shared similarities with so-called ‘sign-tracking’ behavior (Flagel et al., 2009; Hearst and Jenkins, 1974), several features distinguish our observations from sign tracking. First, sign tracking is promoted when reward-predictive cues and reward location are in spatial proximity (Christie, 1996; Flagel et al., 2009). However, in our experimental setup, reward-related manipulanda (i.e., levers, nose-poke port) and reward magazine, were positioned on opposing walls of an operant chamber exceeding standard size (Fig. 1a). Second, it has been reported that rats do not readily sign-track to nose-poke ports or auditory cues (Beckmann and Chow, 2015; Meyer et al., 2014), both critical for No-go trials in our paradigm. Third, prior work demonstrated enhanced VMS dopamine during Pavlovian behavior towards the cue (Flagel et al., 2011, 2009), whereas we observed amplified dopamine after action suppression (and after the cue presentation), during reward approach.
These distinctions suggest that Biting may reflect elevated Pavlovian drive and/or difficulty suppressing reward-directed action, likely due to enhanced motivation (Robinson et al., 2014), rather than sign tracking. Accordingly, our findings point at inter-individual differences in behavioral strategy, analogous to sign and goal-tracking behaviors (Flagel et al., 2011; Robinson et al., 2014). Animals exhibiting heightened task engagement or Palovian drive showed amplified dopamine responses to identical changes in spatial reward proximity, suggesting that encoding of spatial-reward proximity is dynamically modulated by motivational state.
A potential explanation for differences in VMS dopamine may be attributed to Pavlovian biases, wherein reward-associated cues promote approach over suppression, typically resulting in higher Go than No-go performance accuracy. We indeed observed this behavioral asymmetry in our standard Go/No-go task, however, it disappeared when Free trials were included (Fig. 1c). However, VMS dopamine continued to differentiate between Go and No-go trials despite the absence of this phenomenon (Fig. 2c). Therefore, our data show that Pavlovian bias cannot account for the observed differences in dopamine dynamics.
Conclusions
Our study indicates that VMS dopamine is not influenced by behavioral controllability or effort, but instead supports the initiation of reward-seeking actions and the tracking of spatial proximity to rewards, thereby bridging two traditionally separate fields investigating the roles of dopamine in movement and in reward processing, respectively. Building on previous reports that individual dopaminergic neurons multiplex sensory, motor and cognitive information (Engelhard et al., 2019; Kremer et al., 2020), our bulk measurements suggest this multiplexing is extended to the broader population level. These findings position population-level VMS dopamine signals as a unifying mechanism that integrates motivated action and reward. We propose that within this framework, VMS dopamine links motor and motivational processes to ensure that actions are maintained until reward receipt.
Methods
Animals
At the start of the experiment, adult male Long-Evans rats (250-350g, Janvier Labs) were individually housed on a reversed 12 h light/dark cycle (lights off from 08:00 to 20:00) with controlled temperature and humidity. Water was available ad libitum. Following surgical procedures and one week of recovery, rats were food-restricted to 85% of their free-feeding body weight. All animal procedures were performed during the dark phase (between 11:00-19:00) in accordance with Dutch and European law, and approved by the Animal Experimentation Committee of the Royal Netherlands Academy of Arts and Sciences. A total of 27 rats had at least one functional and histologically verified recording electrode and were used in the experiments described in the following sections.
Stereotaxic surgery
Stereotaxic surgery was performed as previously described(Willuhn et al., 2012). Rats were given an analgesic (Metacam, 1-2 mg/kg, s.c.), anesthetized with isoflurane (2-3% maintenance), and placed in a stereotactic frame. Body temperature was maintained at 37°C during surgery using a heating pad and monitored with a probe. The scalp was shaved, disinfected with 70% alcohol, and a midline incision was made. The incision site was treated with lidocaine (100 mg/ml) and the periosteum gently removed to expose the cranium. Holes were drilled for 3-4 anchor surgical screws, two custom-made carbon-fiber microelectrodes(Clark et al., 2010) unilaterally (right hemisphere) targeting the VMS (A-P: +1.2, M-L: +1.5, D-V: –7.1 mm, relative to Bregma(Paxinos and Watson, 2006), and an Ag/AgCl reference electrode that was positioned separately in the forebrain. Electrodes were secured and anchored to the surgical screws with dental acrylic cement. Following surgery, rats were subcutaneously injected with 2 mL saline and placed in a temperature-controlled cabinet until they exhibited mobility. Following implantation of the electrodes, rats were individually housed and were left to recover for at least one week.
Behavioral procedures
Three days before starting behavioral training in the operant boxes, 10-15 sugar pellets/day were introduced into the home cage. Behavioral experiments were performed in modified operant boxes (32 x 30 x 29 cm, MedAssociates Inc.), equipped with a nose-poke hole with an integrated cue-light, house light, auditory-tone generator, food-reward magazine, metal-check grid floor, and retractable levers (Fig. 1a). Operant boxes were interfaced with a fast-scan cyclic voltammetry (FSCV) setup for dopamine recordings of awake and freely-moving animals and a video camera to survey behavior. The operant boxes were stored in individual Faraday cages insulated with sound-absorbing polyurethane foam and ventilated by a fan. Before commencing training or recordings in the operant boxes, rats were habituated to the room for at least 10 min.
Self-initiated task variant
Self-initiated Go/No-go task (“short”; n = 9): Using a behavioral paradigm inspired by Syed and colleagues(Syed et al., 2016), rats initiated a trial by entering the nose-poke port for at least 0.5 s. One of two action (auditory) cues were played, instructing rats to either move towards and press on a unilaterally extended lever at least twice within 5 s (‘Go’; 3 kHz) or to remain in the nose-poke hole for another 1.7-1.9 s (‘No-go’; 1 kHz). After five correct Go trials, the contralateral lever extended and the opposing lever retracted. Successful trial completion was marked by the offset of the action cue and sugar-pellet dispensed into the reward-magazine. During training and recording sessions, trials were separated with a fixed intertrial interval (ITI) of 5 s.
Self-initiated Go/No-go/Free task (“long”; n = 5): Go and No-go trial requirements were increased, together with the inclusion of a third trial type (long-task variant). During training, Go trials required an initial lever press within 1.7 s of action-cue onset and a subsequent lever press within the next 1.5 s. During recording days, a correct Go trial occurred when rats made the first lever press between 2.7 to 3.2 s after cue onset. For No-go trials, rats had to stay in the nose-poke hole for at least 2.7 to 3 s after cue onset. Following successful training of the extended Go and No-go trials, a “Free” trial was introduced (8 kHz tone). Rats were permitted to do anything but nose poke or lever press during the Free cue; most rats walked directly to the food-magazine following cue onset. Rewards were delivered with a delay of 2 s following successful trial completion. Trials were separated with a variable ITI between 15-25 s.
Failure to perform the trial’s required action resulted in house-light illumination for 5 s (i.e., incorrect trial). During training (i.e., non-recording days), the trial type was repeated to a maximum of three times or until the trial was correctly completed, whichever came first. During dopamine-recording days, trial type repetition did not occur. Success rates for each trial type was calculated by determining the percentage of correct/(correct+incorrect) trials, with incorrect trials only included if the error was made after action-cue onset.
Failure to hold the snout in the nose-poke port during the first 0.5 s before action-cue onset resulted in an immediate house-light illumination for 5 s and presentation of another ITI.
Cue-initiated task variant
The structure of Cue-initiated trials was similar to that of Self-initiated trials, except that they could only be initiated following the illumination of the nose-poke light. The nose-poke light stayed illuminated for up to 15 s until initiation, and remained illuminated until the end of the trial. All other trial requirements were identical, except that the house light illuminated for 5 s and a new ITI began when rats made a nose poke or lever press during the ITI, or if they did not perform a nose-poke within 15 s of nose-poke light illumination at the start of the trial. A total of n = 11 and n = 15 were included in the Cue-initiated Go/No-go and Cue-initiated Go/No-go/Free tasks, respectively.
Real-time dopamine recordings and analysis
Simultaneous dopamine and video recordings were carried out when rats were able to perform with >60% success rate for each trial type on at least two consecutive training sessions. On recording days, the trial types were counterbalanced. Within a session, Go left, Go right, and No-go trials were presented with 33% probability each, without replacement. For sessions with Free trials, trials were presented with a 25% chance without replacement.
FSCV measurement and analysis
Fast-scan cyclic voltammetry (FSCV) at chronically implanted carbon-fiber microelectrodes(Clark et al., 2010) was used to record rapid changes in extracellular dopamine concentration. Prior to recording, microelectrodes were connected to a head-mounted voltammetric amplifier interfaced with a PC-driven data acquisition and analysis system (National Instruments) through an electrical commutator(Clark et al., 2010). Voltammetric scans were repeated every 100 ms (i.e., 10 Hz sampling rate). An alternating potential at the carbon-fiber electrode tip was applied from –0.4V versus Ag/AgCl to +1.3V (anodic sweep) and back to –0.4V (cathodic sweep) at 400V/s (total scan time of 8.5ms), and held at –0.4V between scans. Dopamine traces were isolated from the voltammetric signal by chemometric analysis using a standard training set, based on electrically stimulated dopamine release detected with chronically implanted electrodes(Clark et al., 2010). All data were smoothed with a 10-point median filter and baseline (set at 1s before event of interest, with the exception of 2.5 s before action-cue onset for Cue-initiated Go/No-go and Go/No-go/Free tasks) subtraction was performed on a trial-by-trial basis prior to analysis.
Histological verification of recording sites
Following completion of behavioral and recording experiments, animals were terminally anesthetized using sodium pentobarbitone. An electrolytic lesion was performed by running a current on the microelectrodes to mark the FSCV recording sites. Rats were transcardially perfused with saline (0.9% NaCl, w/v) and paraformaldehyde (4% in PBS, w/v). Whole brains were collected and postfixed in Parafresh for 24 h, cryoprotected in sucrose solution (30% in PBS w/v), and cryosectioned coronally (40 μm). Slices were stained with cresyl violet. All animals included in this study had electrodes positioned in the ventromedial striatum (VMS).
DeepLabCut pose estimation
For detailed quantification of behavior across each dataset, we employed DeepLabCut (DLC) to estimate body part coordinates of each animal(Mathis et al., 2018). A total of 1500 manually labelled frames from various videos were used to train a ResNet-50 based neural-network.
Labelled body parts included the snout, neck, four back-points, and tail-base. A reference frame from each video was labelled manually to determine the coordinates of the arena floor. Body coordinates were then transformed from pixels into cm using the ‘fitgeotform2d’ function in MATLAB. Custom analyses were developed in Python to filter DLC coordinates and to analyze behavioral features. Specifically, likelihood cut-off scores for each body part were set at 0.9, and linear interpolation was applied to fill in gaps in the data. Data then underwent Gaussian smoothing to reduce noise. This smoothing was performed using a Gaussian window with a SD calculated to capture approximately 99% of the Gaussian probability density function. The smoothing process was applied within a sliding window of 9 frames (29.97 fps) corresponding to approximately 300 milliseconds.
DeepLabCut behavior analysis
Orientation towards the magazine was determined by calculating the angle between the vector from the neck to the snout, and extending this vector to the wall containing the magazine-panel. Frames of animals were labelled as oriented when this vector crossed the length of the magazine-panel. Magazine arrival was determined as the moment the neck was < 2 cm from the magazine-panel. Movement speed was calculated as the distance from initial orientation to the magazine divided by the elapsed time.
To analyze magazine approach during the ITI, we used DLC-derived coordinates to identify instances where animals were at least 25 cm away from the reward magazine and subsequently moved towards the magazine within 2s (similar to reward approach following a successful trial). This was performed in the Self-initiated Go/No-go/Free task, as animals were permitted to interact with the nose-poke hole and lever during the ITI before orienting and approaching the magazine. Only the second approach and subsequent attempts were included in the analysis. Dopamine traces were then analyzed for these specific moments.
Characterization of No-go behaviors
Behavioral strategies employed by rats during No-go trials were manually scored by an experimenter to systematically assess four main features: vigor (scored 0 to 2), and binary indicators for turning, digging, and biting. To characterize vigor, this was based on a scale of 0 to 2: 0 indicating that the animals were not moving, 1 indicating some energy in their movement, and 2 indicating vigorous movement. Turning was defined as the rat rotating the axis of its body while maintaining their snouts in the nose-poke port. Digging was defined as the repeated placement of their paw(s) into the nose-poke hole. Biting was defined by biting the wall of the nose-poke port.
Statistical Analysis
Data were processed using custom scripts written in MATLAB and Python, and only correct trials were included. Behavioral data were analyzed using GraphPad Prism (v 10.3.1). Due to violations of normality assumptions, a non-parametric Wilcoxon matched-pairs signed rank test was used for pairwise comparisons (trial type) within each task. Kruskal-Wallis tests were used to examine differences across three trial types, and statistical significance was followed-up with post hoc pairwise comparisons using Dunn’s test. All tests were two-tailed and statistical significance was set at p < 0.05.
Dopamine time-series analysis
Dopamine traces within each recording session were directly compared against each other by bootstrapping the within-subject difference trace (e.g., mean Go trace – mean No-go trace within-subject (Jean-Richard-dit-Bressel et al., 2022)). Bootstrapped means were obtained by randomly resampling from subject mean dopamine traces with replacement (1000 iterations). CI limits were derived from 2.5 and 97.5 percentiles of bootstrap distribution, expanded by a factor of 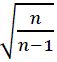
Clustering No-go behaviors and regrouping animals
Prior to clustering, manually scored trial features were normalized. Averages of characterized No-go scores from each animal across all sessions were then clustered into three groups using agglomerative hierarchical clustering with Ward’s linkage method and Euclidean distance as the metric. A decision-tree classifier was implemented in MATLAB to classify labelled trials into their respective groups, and digging and biting were key features identified for group classification (Supp.Fig.4). For Go/No-go tasks, average dopamine was calculated within a 2s window following action-cue onset; and for Go/No-go/Free tasks, within 3s after action-cue onset. Similarly, to determine maximum dopamine release following action-cue offset, values were determined within 2s for Go/No-go tasks and 3s for Go/No-go/Free tasks. Individual trial data were statistically compared using Kruskal-Wallis tests and statistical significance followed-up with post hoc Dunn’s test.
An additional analysis was performed in which animals were regrouped based on their predominant action suppression style within the task (i.e., Biting, Digging or Calm). Regrouped animals were used to compare average and peak dopamine responses across trial types and in response to unpredicted pellets. Due to limited sample sizes within certain subgroups, the resulting data were qualified.
Acknowledgements
We would like to thank members of the Willuhn laboratory for productive discussions. We thank R. Hamelink and A.S.C. França, for providing technical assistance; M.R. Kandroodi for advice on the hierarchical analysis; L. Fellinger, F. Veen, and T. Arbab for input on the manuscript; H.E.M. den Ouden for constructive feedback. This research was supported by a VICI grant from the Netherlands Organization for Scientific Research VI.C.222.052 awarded to I.W. Funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 101066959 was awarded to E.Z.P.
Additional information
Data availability
Behavioral and voltammetry data have been deposited at https://osf.io/wcf2d/ and are publicly available. The code for the analyses presented in this paper is openly accessible at https://osf.io/wcf2d/.
Author contributions
E.Z.P and I.W designed the experiments; E.Z.P, N.B., G.H., L.M., and P.G. performed experiments; N.B., and G.Z., processed video data, E.Z.P. pre-processed and analyzed all data; I.W., L.M., G.H., and P.G assisted with data analysis; E.Z.P. and I.W. wrote and edited the manuscript.
Funding
Netherlands Organization for Scientific Research (VI.C.222.052)
Ingo Willuhn
EC | Horizon Europe | Excellent Science | HORIZON EUROPE Marie Sklodowska-Curie Actions (MSCA)
https://doi.org/10.3030/101066959
Eugenia Z Poh
Additional files
References
- 1.Midbrain dopamine neurons encode a quantitative reward prediction error signalNeuron 47:129–141Google Scholar
- 2.Isolating the incentive salience of reward-associated stimuli: value, choice, and persistenceLearn Mem 22:116–127Google Scholar
- 3.Spatial contiguity facilitates Pavlovian conditioningPsychon Bull Rev 3:357–359Google Scholar
- 4.The timing of action determines reward prediction signals in identified midbrain dopamine neuronsNat Neurosci 21:1563–1573https://doi.org/10.1038/s41593-018-0245-7Google Scholar
- 5.Dynamic mesolimbic dopamine signaling during action sequence learning and expectation violationSci Rep 6:20231https://doi.org/10.1038/srep20231Google Scholar
- 6.Dopamine neuron activity before action initiation gates and invigorates future movementsNature 554:244–248https://doi.org/10.1038/nature25457Google Scholar
- 7.Motivational control of goal-directed actionAnim Learn Behav 22:1–18Google Scholar
- 8.Dopamine Invigorates Reward Seeking by Promoting Cue-Evoked Excitation in the Nucleus AccumbensJ Neurosci 34:14349https://doi.org/10.1523/JNEUROSCI.3492-14.2014Google Scholar
- 9.Specialized coding of sensory, motor and cognitive variables in VTA dopamine neuronsNature 570:509–513https://doi.org/10.1038/s41586-019-1261-9Google Scholar
- 10.Midbrain dopamine neurons signal phasic and ramping reward prediction error during goal-directed navigationCell Rep 41Google Scholar
- 11.Individual differences in the attribution of incentive salience to reward-related cues: Implications for addictionNeuropharmacology 56:139–148Google Scholar
- 12.A selective role for dopamine in stimulus–reward learningNature 469:53–57Google Scholar
- 13.Dissociable cost and benefit encoding of future rewards by mesolimbic dopamineNat Neurosci 13:25–27https://doi.org/10.1038/nn.2460Google Scholar
- 14.Explaining dopamine through prediction errors and beyondNat Neurosci 27:1645–1655Google Scholar
- 15.Believing in dopamineNat Rev Neurosci 20:703–714Google Scholar
- 16.Anticipation of Appetitive Operant Action Induces Sustained Dopamine Release in the Nucleus AccumbensJ Neurosci 43:3922https://doi.org/10.1523/JNEUROSCI.1527-22.2023Google Scholar
- 17.Ramping activity in midbrain dopamine neurons signifies the use of a cognitive mapbioRxiv :2020.05.21.108886https://doi.org/10.1101/2020.05.21.108886Google Scholar
- 18.Wave-like dopamine dynamics as a mechanism for spatiotemporal credit assignmentCell 184:2733–2749https://doi.org/10.1016/j.cell.2021.03.046Google Scholar
- 19.Mesolimbic dopamine signals the value of workNat Neurosci 19:117–126https://doi.org/10.1038/nn.4173Google Scholar
- 20.Sign-tracking: The stimulus-reinforcer relation and directed actionNo Title Google Scholar
- 21.Rapid signalling in distinct dopaminergic axons during locomotion and rewardNature 535:505–510https://doi.org/10.1038/nature18942Google Scholar
- 22.Prolonged dopamine signalling in striatum signals proximity and value of distant rewardsNature 500:575–579https://doi.org/10.1038/nature12475Google Scholar
- 23.The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seekingBrain Res Rev 31:6–41Google Scholar
- 24.Instrumental aversion coding in the basolateral amygdala and its reversion by a benzodiazepineNeuropsychopharmacology 47:1199–1209Google Scholar
- 25.Start/stop signals emerge in nigrostriatal circuits during sequence learningNature 466:457–462https://doi.org/10.1038/nature09263Google Scholar
- 26.A Unified Framework for Dopamine Signals across TimescalesCell 183:1600–1616https://doi.org/10.1016/j.cell.2020.11.013Google Scholar
- 27.Dual credit assignment processes underlie dopamine signals in a complex spatial environmentNeuron 111:3465–3478https://doi.org/10.1016/j.neuron.2023.07.017Google Scholar
- 28.Context-dependent multiplexing by individual VTA dopamine neuronsJ Neurosci 40:7489–7509Google Scholar
- 29.Dopamine, Updated: Reward Prediction Error and Beyond. Neurobiol Learn Plast 67:123–130https://doi.org/10.1016/j.conb.2020.10.012Google Scholar
- 30.DeepLabCut: markerless pose estimation of user-defined body parts with deep learningNat Neurosci 21:1281–1289https://doi.org/10.1038/s41593-018-0209-yGoogle Scholar
- 31.The form of a conditioned stimulus can influence the degree to which it acquires incentive motivational propertiesPloS One 9:e98163Google Scholar
- 32.The role of state uncertainty in the dynamics of dopamineCurr Biol 32:1077–1087https://doi.org/10.1016/j.cub.2022.01.025Google Scholar
- 33.Dissociable dopamine dynamics for learning and motivationNature 570:65–70https://doi.org/10.1038/s41586-019-1235-yGoogle Scholar
- 34.The flexible approach hypothesis: unification of effort and cue-responding hypotheses for the role of nucleus accumbens dopamine in the activation of reward-seeking behaviorJ Neurosci 30:16585–16600Google Scholar
- 35.The nucleus accumbens as part of a basal ganglia action selection circuitPsychopharmacology 191:521–550https://doi.org/10.1007/s00213-006-0510-4Google Scholar
- 36.Dialogues on prediction errorsTrends Cogn Sci 12:265–272Google Scholar
- 37.Dopamine Is Required for the Neural Representation and Control of Movement VigorCell 162:1418–1430https://doi.org/10.1016/j.cell.2015.08.014Google Scholar
- 38.Subsecond dopamine release promotes cocaine seekingNature 422:614–618Google Scholar
- 39.On the motivational properties of reward cues: Individual differencesNIDA 40th Anniv Issue 76:450–459https://doi.org/10.1016/j.neuropharm.2013.05.040Google Scholar
- 40.Dopamine operates as a subsecond modulator of food seekingJ Neurosci 24:1265–1271Google Scholar
- 41.Dopamine neurons of the monkey midbrain: contingencies of responses to active touch during self-initiated arm movementsJ Neurophysiol 63:592–606https://doi.org/10.1152/jn.1990.63.3.592Google Scholar
- 42.A Neural Substrate of Prediction and RewardScience 275:1593–1599Google Scholar
- 43.Dopamine signals as temporal difference errors: recent advances. Curr Opin NeurobiolNeurobiology of Learning and Plasticity 67:95–105https://doi.org/10.1016/j.conb.2020.08.014Google Scholar
- 44.Action initiation shapes mesolimbic dopamine encoding of future rewardsNat Neurosci 19:34–36https://doi.org/10.1038/nn.4187Google Scholar
- 45.What is the relationship between dopamine and effort?Trends Neurosci 42:79–91Google Scholar
- 46.Phasic Mesolimbic Dopamine Signaling Precedes and Predicts Performance of a Self-Initiated Action Sequence TaskBiol Psychiatry 71:846–854https://doi.org/10.1016/j.biopsych.2011.12.019Google Scholar
- 47.Chronic microsensors for longitudinal, subsecond dopamine detection in behaving animalsNat Methods 7:126–129https://doi.org/10.1038/nmeth.1412Google Scholar
- 48.Analyzing event-related transients: confidence intervals, permutation tests, and consecutive thresholdsFront Mol Neurosci 13:14Google Scholar
- 49.Instrumental aversion coding in the basolateral amygdala and its reversion by a benzodiazepineNeuropsychopharmacology 47:1199–1209Google Scholar
- 50.DeepLabCut: markerless pose estimation of user-defined body parts with deep learningNat Neurosci 21:1281–1289https://doi.org/10.1038/s41593-018-0209-yGoogle Scholar
- 51.The Rat Brain in Stereotaxic Coordinates: Hard Cover EditionElsevier Google Scholar
- 52.Action initiation shapes mesolimbic dopamine encoding of future rewardsNat Neurosci 19:34–36https://doi.org/10.1038/nn.4187Google Scholar
- 53.Hierarchical recruitment of phasic dopamine signaling in the striatum during the progression of cocaine useProc Natl Acad Sci 109:20703–20708https://doi.org/10.1073/pnas.1213460109Google Scholar
Article and author information
Author information
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.109868. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026, Poh et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
- views
- 473
- downloads
- 31
- citations
- 0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.



