Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorMegan CareyChampalimaud Foundation, Lisbon, Portugal
- Senior EditorJoshua GoldUniversity of Pennsylvania, Philadelphia, United States of America
Reviewer #1 (Public review):
In this manuscript, Hinojosa and colleagues analysed the changes in V1 visual responses induced by locomotion in head-fixed mice using two-photon calcium imaging. The authors observe that locomotion strongly increases the visual responses of V1 excitatory neurons that exhibit sensitizing responses to visual stimuli. Also, there is an increased response in VIP interneurons, and to a lesser extent, PV interneurons and SST interneurons (non-significant). The authors used a model fitted with data presented in the manuscript, as well as previous knowledge on cortical connectivity among different neuron types. The model suggests that the major component of the increased responses during locomotion is an increase in excitatory drive from external inputs (feedforward, feedback and modulatory), most importantly onto VIP interneurons and excitatory neurons. However, the excitatory drive of local excitatory neurons onto other surrounding excitatory and inhibitory cells is reduced.
The manuscript is well presented and represents a valuable analysis of how locomotion modulates the activity of different subtypes of cortical neurons. However, major issues should be addressed to strengthen the results.
Major issues:
(1) Speed and mismatch between locomotion and visual stimulation.
The authors do not clearly describe the definition of locomotion versus the resting state. The speed should, by itself, have an impact on neuronal responses, especially at the onset of locomotion. Several published studies show that the mismatch between a visual stimulus and the speed of the animal induces specific responses in V1, both in excitatory and subtypes of inhibitory neurons. The authors should address these points upfront in the manuscript, since it is likely a major variable explaining their results
(2) Use of deconvolution with MLSpike.
Some results (Figure 2) exclusively depend on the deconvolution of calcium signals into spikes (since the initial peak is not seen in calcium transients). The authors should validate this result either with electrophysiological recordings or with the use of another deconvolution method (e.g. CASCADE), emphasising the limitations of this approach and the limitations of the time resolution of calcium imaging.
(3) The manuscript is centred around a specific increase in visual responses in sensitizing neurons during locomotion, both in the fraction of responsive neurons and response magnitudes.
It is hard to tell whether this difference is due to a greater scaling effect of locomotion, a difference in responses during the resting state, or both. The manuscript should further explore and discuss the differences in responses between sensitizing and depressing neurons, both during the resting state and locomotion. Adding metrics and direct comparisons of the magnitudes of fast responses, slow responses, and time integrals between sensitizing and depressing neurons in resting and locomotion states would help to clarify this. Same for fractions of responsive neurons of each type in each condition. E.g., the slow phase is harder to judge from the plots, but the DeltaF/F integral shown in Figure 1G seems to suggest the difference in response magnitude between sensitizing and depressing neurons is largest in locomotion state, rather than resting state. How do these integrals look for inferred firing rates shown in Figure 2?
(4) There is something counterintuitive about how the changes in inhibition onto sensitizing and depressing neurons during locomotion explain the reported activity changes.
Sensitizers receive reduced SST input and increased PV input during locomotion. If SSTs depress and PVs sensitize (and this is the main reason why sensitizers, which receive dominant input from SSTs sensitize, and vice-versa), how is it possible that this switch does not alter the sensitizing or depressing nature of these neurons' responses in locomotion? Are these changes insufficient to flip the dominant SST-PV drive? Figure 6D-E seems to show there is a flip, at least for sensitizers. How do authors explain this? Do authors think this is related to the narrowing of the adaptive index distribution shown in Figure 1C?
(5) Presentation of the experimental data and the model.
The manuscript introduces the results of interneuron recordings during the description of the model. Similarly, the results of optogenetic manipulations are presented inside the model's description. It would be clearer to present all experimental data first and introduce the model later, fitting it to all experimental evidence previously presented.
Reviewer #2 (Public review):
This is an interesting paper with important results. The authors, working in V1, have previously, in a 2022 paper, defined sensitizing and depressing excitatory (E) cells as those whose response increases or decreases, respectively, across the 10 seconds of showing a drifting grating stimulus. They showed that sensitizing E cells are dominantly inhibited by SST inhibitory cells, which are dominantly depressing, and that depressing E cells are dominantly inhibited by PV inhibitory cells, which are very largely sensitizing. It's been well established that locomotion greatly increases E-cell firing rates in V1 compared to rest, but much remains to be worked out as to the mechanism. Here, they find that locomotion increases the responses of the sensitizing E cells much more than depressing cells. They develop a model of changes in synaptic weights between rest and locomotion to account for the changes. One reason that sensitizers are increased more by locomotion than depressors is that PV cells, which more strongly inhibit depressors, have increased firing for locomotion, whereas SST cells, which more strongly inhibit sensitizers, don't change their firing rates with locomotion. However, in the mode,l a complex array of postulated changes in connection strengths is also involved.
I have, though, a number of concerns: with the model, with the lack of proper discussion of connection to some previous works, and with an overall unclear and confusing presentation and certain controls that should be done.
In the model, they postulate that synapses within the 6-cell-type network - sensitizing, intermediate, and depressing E cells, and PV, SST, and VIP I cells - and from three sources of external input to each of the six types all change between rest and locomotion (except that connections between the E cells don't depend on their types). There are a lot of degrees of freedom, and this makes interpretation of the results difficult. I would have liked to have seen more efforts to constrain the degrees of freedom. For example, there seems to be very little difference between the three E cell types in any of the three types of external input received. Why not constrain them all to get the same external input and see if it significantly affects model fit? Or what if synapses from the three types of external input are left unchanged, and only change their strengths between rest and locomotion? How well could this do? During optimization, why not constrain the changes between rest and locomotion, for example, by putting an L1 penalty on the changes or the relative changes, trying to force them to be sparse, and see whether there are roughly equally good fits? And then, if the main changes are in a small set of synapses, can the authors isolate changes to that small set and do roughly equally well? What about looking at the principal components of the weight changes across models, to isolate patterns of change that are most important?
In terms of comparing to previous works, when optogenetic manipulations of SST and PV are done to test various hypotheses, I would like to see some discussion of what is already known from the authors' 2022 paper and what they are adding or testing that wasn't known or tested from that paper. And Dipoppa et al (2018) also found weight changes to account for the difference between rest and locomotion. They were looking at a fixed point of responses of neurons across retinotopic space to stimuli of various sizes with only one E-cell type, whereas they are accounting for trajectories across time considering 3 E-cell subtypes but without variation in stimuli or retinotopic position of neurons, so the efforts are somewhat different, but still, it would be good to see a bit more discussion of what is in agreement or in contradiction in the conclusions.
In terms of presentation and controls, I have many concerns, which include:
(1) The main result is that sensitizers increase their responses with locomotion ~2X (for dF/F) or about 3.5X (for spikes) more than depressors. But there are other differences between sensitizers and depressors, for example sensitizers have smaller initial stimulus responses at rest, and depressors have larger. What if cells were divided into tertiles by initial stimulus response at rest? Would the authors see the same differences in the effects of locomotion? If so, can they establish whether the difference is really attached to the adaptation properties rather than to, for example, the initial responses, for example, by comparing the regression of response increase against AI vs the regression of response increase against initial resting response? And there might be other controls to be done for other features in which sensitizers and depressors differ.
(2) Lines 103 and following: the authors refer to a "second notable change" which is the narrower distribution of adaptive effects, but I think this is trivial. The adaptive index is AI=(R1-R2)/(R1+R2), where R1 is response 0.5-2.5s after stimulus onset and R2 over 8-10s. But if the change is additive, as suggested by the dF/F figures (and I believe the distributions of AI here are based on dF/F measurements) -- adding the same constant to R1 and R2 will shrink |AI| without changing the sign of AI. So this would seem to just be a signature of a change that is primarily additive rather than multiplicative.
Also, if the authors do decide that they are going to focus on spikes after showing the raw dF/F, then this analysis should be repeated for spikes.
(3) Figure 2, F is supposed to be D minus E, but it doesn't look like it. For example, the initial response under locomotion is very similar in sensitizers and depressors, so the initial difference in F should be small, but it's not; and at rest, depressors initially have larger responses than sensitizers, whereas later depressors have smaller responses than sensitizers, yet the difference at rest is positive at all times. Something seems wrong here.
Reviewer #3 (Public review):
This study aimed to understand the depressing and sensitizing effects of adaptation in mice visual cortex during different behavioral states: locomotion and stationary. There is an impressive characterisation of the responses in different cortical cell types and with different optogenetic manipulations to the inhibitory populations. These form a very interesting dataset to understand the effects of the state on the circuits and gain insight into the mechanisms. This data is then used to constrain a model of the responses. Unfortunately, the model appears to be too flexible, and it was difficult to interpret the insights gained from the different model fits.
Strengths:
The data is impressive. There is a characterisation of responses of PCs and VIP, SST and PV interneurons. Additionally, there is the characterisation of some responses to specific optogenetic manipulations, VIP inactivation, SST or PV activation or inactivation. These data will help develop a good insight into the system. The principle of using the optigenetic manipulations to constrain model parameters is very interesting.
Weaknesses:
Many of the analyses have some concerns in the methodology used, which we list in detail below. Further, the model used to gain insight into the mechanism appears overly complicated and seems hard to gain clear insights from.
Major concerns:
(1) Key concern is the usage of dF/F signals for all analyses, especially when comparing responses.
1a) Figure 1G: Comparison of sensitisers and depressors. It is important to consider what the baseline rates are when making these comparisons, especially when comparing the degree of effects between different cell types. For example, if baseline rates for sensitizers were overall higher, it would mean the difference in gain of response would be lower, and could affect the results in the opposite direction of what is claimed. One option to account for this would be to z-score the overall responses, using the same normalization for locomotion and rest. We also suggest plotting differences in sensitisers, intermediates, and depressors as a function of firing rate. Matching for firing rate across each PC categorization and calculating delta AI for each matched firing rate bin.
1b) Figure 2A-F: The above is an even more significant issue when it comes to estimating spiking rates. The methods do not state how dF/F is calculated. If these are based on using the pre-stim as the reference, the algorithms for spike rate used might not be appropriate if this were used. Using pre-stimulus referencing could result in the estimate going into the wrong range in the calculation of the spike rate.
1c) In both cases above, it could be a problem if baseline firing rates are different between cell types, or states (locomotion/stationary). The latter is established to have effects on many cell types measured, and so needs to be accounted for very carefully.
1d) It would be informative to see per-neuron comparison for adaptive indices during rest and locomotion states. This could be visualized using a scatter plot with AI-rest vs. AI-locomotion for Figures 1D- 1F and 2J- 2L.
1e) Are neurons more strongly modulated between locomotion and rest, also more likely to experience a shift in AI indices (i.e. delta AI). Is there a correlation between the change in firing rate between behavioral states and Delta AI (Loco-Rest)? If so, is this present for all neuron subtypes (e.g. VIP, SST, and PV)?
1f) Optogenetic inhibition of VIP neurons on average abolished the slow depressive effects of adaptation in SST (Figure 3). The strength and prevalence of this effect are unclear. Perhaps one can perform a bootstrap control and opto AI indices and calculate whether AI was significantly reduced following optogenetics inhibition, and if so, on average, how likely was this to occur for the recorded SST neurons? This is important in knowing that the average effects (Figure 3D) aren't driven by a portion of SST neurons, especially as this is later used to confirm the region of parameter space and affects the subsequent results in Figure 4.
(2) Statistics for the effects. There is a mention of Liner mixed models, but no information is given on the actual models being used and tested. This is particularly for the case of Figure 1G, where there is a composition of effect sizes between different populations. What precise significance test is being used? Are the stats on paired cells when considering locomotion and rest?
(3) Model parameters: It is acknowledged that there is a large range of parameters that can model the responses effectively, up to 11% of initial conditions. At 9000 initial conditions, this is around 1000. The parameter estimates are then considered as the mean of each parameter. This seems like a strange choice for a few different reasons:
3a) A mean solution might not be one of the solutions. Let's say the parameters range over a large dimensional space. They could occupy non-overlapping / discontinuous subspaces. In that case, the mean parameters do not necessarily fall within the solution subspaces. Therefore, this reduction to means might not be valid.
3b) Compare distributions rather than means. There are multiple distributions of parameters between conditions. All stats should be on the comparison of distributions rather than just the means.
(4) Visualizing weight matrices: It is very challenging to interpret the weight matrices. Furthermore, it appears that the stationary and locomotion conditions fit independently, and given the large parameter spaces, it is even harder to interpret. Can the fitting instead be done by fitting on one and using those at the initial conditions for the other state? Figure 7 shows an initiative cartoon, but it is not clear how the matrices in Figures 5 and 6 lead to the summary shown in Figure 7. It is also not clear why the connections between inhibitory neurons are not shown in Figure 7. One option is to perhaps run some kind of dimensionality deduction on the parameter space to better interpret the data. When showing deltaWeights, was the model initialised with 'Rest' weights and allowed to change? It is not obvious what the difference is between 'relative change in connection weights' and 'relative change in synaptic weights'.This needs to be clarified.
4a) Model parameters were reduced differently for locomotion and rest (Figure 4). We suggest evaluating the results for locomotion and rest using the same chi-square value of 3 for both behavioral states (at least in controls).
Author response:
Public Reviews:
Reviewer #1 (Public review):
(1) Speed and mismatch between locomotion and visual stimulation.
The authors do not clearly describe the definition of locomotion versus the resting state. The speed should, by itself, have an impact on neuronal responses, especially at the onset of locomotion. Several published studies show that the mismatch between a visual stimulus and the speed of the animal induces specific responses in V1, both in excitatory and subtypes of inhibitory neurons. The authors should address these points upfront in the manuscript, since it is likely a major variable explaining their results.
We will clarify in the methods that a trial was considered as locomotion when an animal ran at a minimum of 3 cm/s for at least 80% of the 10 s stimulus presentation, and was considered rest when running under 3 cm/s during the same fraction of time. Trials with abrupt changes from locomotion to rest were rare and excluded following these criteria.
Locomotion speed and visuomotor mismatch can influence neuronal responses in V1 but in the large majority of our trials mice either run continuously at a stable speed or remained still
i.e locomotion onsets or offsets did not occur (see Hinojosa et al. 2026 for example running traces). Furthermore, sensitizing and depressing neurons were typically recorded simultaneously within the same field of view, experiencing identical locomotor behaviour. For these reasons, we think it is unlikely that differences in speed or mismatch alone can account for the different increase in amplitude observed between depressors and sensitizers.
To directly address this point and further explore the role of speed on V1 neurons, we will quantify the relationship between running speed and amplitude increase in both PCs and interneurons, and include these analyses in the revised version of the manuscript.
(2) Use of deconvolution with MLSpike.
Some results (Figure 2) exclusively depend on the deconvolution of calcium signals into spikes (since the initial peak is not seen in calcium transients). The authors should validate this result either with electrophysiological recordings or with the use of another deconvolution method (e.g CASCADE), emphasising the limitations of this approach and the limitations of the time resolution of calcium imaging.
A similar initial increase in amplitude followed by fast depression has been observed previously with electrophysiological recordings in V1 (Chance et al., 1998; Jin & Glickfeld, 2020; Varela et al., 1997). We will further validate our results using an alternative spike inference method like CASCADE (Rupprecht et al., 2021), as well as expanding on the limitations of our approach.
(3) The manuscript is centred around a specific increase in visual responses in sensitizing neurons during locomotion, both in the fraction of responsive neurons and response magnitudes.
It is hard to tell whether this difference is due to a greater scaling effect of locomotion, a difference in responses during the resting state, or both. The manuscript should further explore and discuss the differences in responses between sensitizing and depressing neurons, both during the resting state and locomotion. Adding metrics and direct comparisons of the magnitudes of fast responses, slow responses, and time integrals between sensitizing and depressing neurons in resting and locomotion states would help to clarify this. Same for fractions of responsive neurons of each type in each condition. E.g., the slow phase is harder to judge from the plots, but the DeltaF/F integral shown in Figure 1G seems to suggest the difference in response magnitude between sensitizing and depressing neurons is largest in locomotion state, rather than resting state. How do these integrals look for inferred firing rates shown in Figure 2?
We will further explore the response dynamics of adaptive types within the locomotion and resting state, highlighting the differences between calcium signals and inferred spikes. We will then include our findings in the new version.
(4) There is something counterintuitive about how the changes in inhibition onto sensitizing and depressing neurons during locomotion explain the reported activity changes.
Sensitizers receive reduced SST input and increased PV input during locomotion. If SSTs depress and PVs sensitize (and this is the main reason why sensitizers, which receive dominant input from SSTs sensitize, and vice-versa), how is it possible that this switch does not alter the sensitizing or depressing nature of these neurons' responses in locomotion? Are these changes insufficient to flip the dominant SST-PV drive? Figure 6D-E seems to show there is a flip, at least for sensitizers. How do authors explain this? Do authors think this is related to the narrowing of the adaptive index distribution shown in Figure 1C?
This result is only counterintuitive if we consider exclusively the internal connections within V1. The PV:SST ratio changes from 0.9 during rest, dominated by SST induced sensitization, to 1.2, dominated by PV depression. Although adaptation is strongly driven by the opposing inhibition of PV and SST in PCs during locomotion, its origin is more easily explained by an external input (SS) that targets VIPs, PVs and PCs. As a result, when locomotion increases the drive coming from SS input, it injects a source of sensitization that partly balances the decrease in PV:SST ratio, preventing a switch in their adaptive properties which, although reduced, remain sensitizing. We will include these calculations in the revised version.
(5) Presentation of the experimental data and the model.
The manuscript introduces the results of interneuron recordings during the description of the model. Similarly, the results of optogenetic manipulations are presented inside the model's description. It would be clearer to present all experimental data first and introduce the model later, fitting it to all experimental evidence previously presented.
We understand that a clear separation between experimental and modelling results is often preferred in papers that combine these approaches but in our case modelling and experimental data are highly interdependent and we believe that an overlapping presentation make it easier for the reader to appreciate the links. One example is Fig. 2G-L that shows experimental results validating a key feature of the model - the use of average response dynamics for each population of interneuron. Similarly, the results in Fig. 3 validate the use of the VIP response dynamics as the template for the slow modulatory input to layer 2/3. Then the results of optogenetic experiments in Fig. 4 are used to narrow down fits to the model. For these reasons, we have chosen to present experimental results and the model in this more integrated manner.
Reviewer #2 (Public review):
In the model, they postulate that synapses within the 6-cell-type network - sensitizing, intermediate, and depressing E cells, and PV, SST, and VIP I cells - and from three sources of external input to each of the six types all change between rest and locomotion (except that connections between the E cells don't depend on their types). There are a lot of degrees of freedom, and this makes interpretation of the results difficult. I would have liked to have seen more efforts to constrain the degrees of freedom. For example, there seems to be very little difference between the three E cell types in any of the three types of external input received. Why not constrain them all to get the same external input and see if it significantly affects model fit? Or what if synapses from the three types of external input are left unchanged, and only change their strengths between rest and locomotion? How well could this do? During optimization, why not constrain the changes between rest and locomotion, for example, by putting an L1 penalty on the changes or the relative changes, trying to force them to be sparse, and see whether there are roughly equally good fits? And then, if the main changes are in a small set of synapses, can the authors isolate changes to that small set and do roughly equally well? What about looking at the principal components of the weight changes across models, to isolate patterns of change that are most important?
To reduce the number of degrees of freedom and ease interpretation we did limit the model fitting for adaptive subtypes by fixing the PC-PC (𝑤𝑃𝐶_𝑃𝐶) and restricting the external inputs weights (𝑤𝐹𝐹_𝑃𝐶, 𝑤𝑆𝑆_𝑃𝐶, 𝑤𝐹𝐵_𝑃𝐶) to changes of ± 10 %. We will explicitly explain these constrains in the methods and discuss its limitations.
We thank the reviewer for their suggestions of testing different conditions to find those providing the best fit for sensitizing and depressing PCs. We tried an approach similar to that described by Dipoppa et al. 2018 by using the locomotion weights as initial conditions for the rest traces and introducing penalties at later stages. However, the local optimization algorithms failed to reach distant regions of parameter space containing minimum 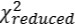 solutions for the rest condition. We finally opted for repeating the same process of initial condition searching for locomotion and rest, making the L1 penalty approach impracticable in our case. We believe this approach is effective because it has both allowed us to describe circuit changes during internal-state transitions (the present paper) and, more recently, it has made a series of predictions about different learning states that have been confirmed by optogenetic tests (Hinojosa et al., 2026). We will nevertheless explore this and other of the reviewer suggestions to further optimize the fitting in the revised manuscript.
solutions for the rest condition. We finally opted for repeating the same process of initial condition searching for locomotion and rest, making the L1 penalty approach impracticable in our case. We believe this approach is effective because it has both allowed us to describe circuit changes during internal-state transitions (the present paper) and, more recently, it has made a series of predictions about different learning states that have been confirmed by optogenetic tests (Hinojosa et al., 2026). We will nevertheless explore this and other of the reviewer suggestions to further optimize the fitting in the revised manuscript.
In terms of comparing to previous works, when optogenetic manipulations of SST and PV are done to test various hypotheses, I would like to see some discussion of what is already known from the authors' 2022 paper and what they are adding or testing that wasn't known or tested from that paper. And Dipoppa et al (2018) also found weight changes to account for the difference between rest and locomotion. They were looking at a fixed point of responses of neurons across retinotopic space to stimuli of various sizes with only one E-cell type, whereas they are accounting for trajectories across time considering 3 E-cell subtypes but without variation in stimuli or retinotopic position of neurons, so the efforts are somewhat different, but still, it would be good to see a bit more discussion of what is in agreement or in contradiction in the conclusions.
Thanks for this prompt. We will add further discussion of this work in light of the Heintz et al. (2022) and Dipoppa et al. (2018) papers.
(1) The main result is that sensitizers increase their responses with locomotion ~2X (for dF/F) or about 3.5X (for spikes) more than depressors. But there are other differences between sensitizers and depressors, for example sensitizers have smaller initial stimulus responses at rest, and depressors have larger. What if cells were divided into tertiles by initial stimulus response at rest? Would the authors see the same differences in the effects of locomotion? If so, can they establish whether the difference is really attached to the adaptation properties rather than to, for example, the initial responses, for example, by comparing the regression of response increase against AI vs the regression of response increase against initial resting response? And there might be other controls to be done for other features in which sensitizers and depressors differ.
We will explore the possibility that initial response influences the increase in amplitude. Preliminary data suggest that initial amplitude is higher in depressors than in sensitizers.
(2) Lines 103 and following: the authors refer to a "second notable change" which is the narrower distribution of adaptive effects, but I think this is trivial. The adaptive index is AI=(R1-R2)/(R1+R2), where R1 is response 0.5-2.5s after stimulus onset and R2 over 8-10s. But if the change is additive, as suggested by the dF/F figures (and I believe the distributions of AI here are based on dF/F measurements) -- adding the same constant to R1 and R2 will shrink |AI| without changing the sign of AI. So this would seem to just be a signature of a change that is primarily additive rather than multiplicative.
Also, if the authors do decide that they are going to focus on spikes after showing the raw dF/F, then this analysis should be repeated for spikes.
We agree with the reviewer and will change the text accordingly to highlight the additive nature of the change in amplitude. We will also show the analysis with spikes (this shows similar results as the calcium data).
(3) Figure 2, F is supposed to be D minus E, but it doesn't look like it. For example, the initial response under locomotion is very similar in sensitizers and depressors, so the initial difference in F should be small, but it's not; and at rest, depressors initially have larger responses than sensitizers, whereas later depressors have smaller responses than sensitizers, yet the difference at rest is positive at all times. Something seems wrong here.
We apologize for the confusion this has caused. Figure 2F does not represent the difference between sensitizing and depressing PCs from panels D and E. Instead, it shows the time-varying difference between locomotion and rest states of sensitizers (blue, in figure 2D) and depressors (green, in figure 2E). Thus, panel F shows within-population modulation by behavioural state, rather than differences between sensitizing and depressing neurons. We will amend the figure legend and main text to explain this point and avoid misinterpretation.
Reviewer #3 (Public review):
(1) Key concern is the usage of dF/F signals for all analyses, especially when comparing responses.
(1a) Figure 1G: Comparison of sensitisers and depressors. It is important to consider what the baseline rates are when making these comparisons, especially when comparing the degree of effects between different cell types. For example, if baseline rates for sensitizers were overall higher, it would mean the difference in gain of response would be lower, and could affect the results in the opposite direction of what is claimed. One option to account for this would be to z-score the overall responses, using the same normalization for locomotion and rest. We also suggest plotting differences in sensitisers, intermediates, and depressors as a function of firing rate. Matching for firing rate across each PC categorization and calculating delta AI for each matched firing rate bin.
(1b) Figure 2A-F: The above is an even more significant issue when it comes to estimating spiking rates. The methods do not state how dF/F is calculated. If these are based on using the pre-stim as the reference, the algorithms for spike rate used might not be appropriate if this were used. Using pre-stimulus referencing could result in the estimate going into the wrong range in the calculation of the spike rate.
(1c) In both cases above, it could be a problem if baseline firing rates are different between cell types, or states (locomotion/stationary). The latter is established to have effects on many cell types measured, and so needs to be account ted for very carefully.
The DF/F0 trace was calculated using the mode of the whole trace as F0. While this approach is less sensitive to biases than subtracting the pre-stimulus, it does not consider noise levels like the z-score suggested by the reviewer. We will, therefore, normalize the calcium traces to z-score to further account for changes in the baseline. Spike inference using MLSpike, however, explicitly models baseline noise and subtracts its effect from that of the spikes calculated from the calcium signal (Deneux et al., 2016). This transformation preserved the difference in amplitude triggered by locomotion between depressing and sensitizing PCs while revealing their similar baseline activity (see Figs. 2D,E and F). These results indicate that the distinct changes in response amplitude between sensitizing and depressing PCs during locomotion are not driven by baseline differences. We will add this explanation to the methods section.
We will also plot the changes in activity with locomotion across cell types as a function of firing rate and add these results to the revised manuscript.
(1d) It would be informative to see per-neuron comparison for adaptive indices during rest and locomotion states. This could be visualized using a scatter plot with AI-rest vs. AI-locomotion for Figures 1D- 1F and 2J- 2L.
(1e) Are neurons more strongly modulated between locomotion and rest, also more likely to experience a shift in AI indices (i.e. delta AI). Is there a correlation between the change in firing rate between behavioral states and Delta AI (Loco-Rest)? If so, is this present for all neuron subtypes (e.g. VIP, SST, and PV)?
Sorting was carried out separately on locomotion and rest data sets to capture the adaptive properties of the network under each condition. When assessing the change in adaptive index in individual cells there was a weak but significant correlation (r = 0.10, p<0.05), probably due to trial to trial stochasticity in the network which has been shown to be present in V1 (Carandini, 2004; Lee et al., 2010). Although adaptation profiles of individual PCs are not fully conserved across rest and locomotion, the observed overlap exceeds that expected by chance, suggesting that stochastic fluctuations modulate an underlying, stable circuit organization. Despite including the stochastic component of the responses, the conclusions hold: sensitizers undergo a larger gain modulation than that of depressors. We will include this analysis and the correlation between change in firing rate and Delta AI in the revised version of the paper.
(1f) Optogenetic inhibition of VIP neurons on average abolished the slow depressive effects of adaptation in SST (Figure 3). The strength and prevalence of this effect are unclear. Perhaps one can perform a bootstrap control and opto AI indices and calculate whether AI was significantly reduced following optogenetics inhibition, and if so, on average, how likely was this to occur for the recorded SST neurons? This is important in knowing that the average effects (Figure 3D) aren't driven by a portion of SST neurons, especially as this is later used to confirm the region of parameter space and affects the subsequent results in Figure 4.
The strength and prevalence of the effect are reflected in the distribution of AI changes across SST neurons, which is centred at AI = -0.3 ± 0.3, indicating a consistent reduction in AI across the population instead of being driven by a small portion of SST neurons. To further clarify this, we will report the proportion of SST neurons showing a reduction in AI and include statistical analyses on the changes.
(2) Statistics for the effects. There is a mention of Liner mixed models, but no information is given on the actual models being used and tested. This is particularly for the case of Figure 1G, where there is a composition of effect sizes between different populations. What precise significance test is being used? Are the stats on paired cells when considering locomotion and rest?
We used Linear mixed models to test for statistical significance between different conditions composed of hundreds of cells from several mice, i.e. nested analysis (cells nested within mice; see (Judd et al., 2017)). For analyses such as Fig. 1G, we considered locomotion state, adaptive type and their interaction (loco’adap) as fixed effects and mouse number as the random effect. The p-values depicted in the legend indicates the interaction between locomotion and adaptive type, i.e. the increase in amplitude during locomotion is significantly different in sensitizers compared to depressors with p < 0.0001. We will revise the method section and figure legends to explicitly describe the model and statistical test used.
(3) Model parameters: It is acknowledged that there is a large range of parameters that can model the responses effectively, up to 11% of initial conditions. At 9000 initial conditions, this is around 1000. The parameter estimates are then considered as the mean of each parameter. This seems like a strange choice for a few different reasons:
(3a) A mean solution might not be one of the solutions. Let's say the parameters range over a large dimensional space. They could occupy non-overlapping / discontinuous subspaces. In that case, the mean parameters do not necessarily fall within the solution subspaces. Therefore, this reduction to means might not be valid.
(3b) Compare distributions rather than means. There are multiple distributions of parameters between conditions. All stats should be on the comparison of distributions rather than just the means.
To test for the presence of subsets of solutions grouped around different parameter values we plotted the distribution of each parameter across all the good solutions found. Most of the weights were a gaussian distribution centred around the mean and, most importantly, none of them had two peaks. Furthermore, after computing the mean weight values we plotted the solutions given by them in the model, and it rendered a good fit as shown in the figures. We will include those distributions in the new version and base the overall comparison on these distributions.
(4) Visualizing weight matrices: It is very challenging to interpret the weight matrices. Furthermore, it appears that the stationary and locomotion conditions fit independently, and given the large parameter spaces, it is even harder to interpret. Can the fitting instead be done by fitting on one and using those at the initial conditions for the other state? Figure 7 shows an initiative cartoon, but it is not clear how the matrices in Figures 5 and 6 lead to the summary shown in Figure 7. It is also not clear why the connections between inhibitory neurons are not shown in Figure 7. One option is to perhaps run some kind of dimensionality deduction on the parameter space to better interpret the data. When showing deltaWeights, was the model initialised with 'Rest' weights and allowed to change? It is not obvious what the difference is between 'relative change in connection weights' and 'relative change in synaptic weights'.This needs to be clarified.
Thanks for raising this concern. We will firstly try to make the weight matrices clearer to interpret.
Regarding the fitting of rest and locomotion conditions, we fitted the locomotion traces first and used those solutions as initial conditions for the rest traces. However, this rendered no good solutions as minimums in the parameter space were too far from the initial starting points. We opted, therefore, for repeating the same process of initial condition searching for locomotion and rest. This approach is less biased in satisfying our aim of finding solutions that fit the data and can explain their dynamics, which are different for each condition. We believe this approach is effective, as not only has it allowed us to describe circuit changes during internal-state transitions but has also made a series of predictions under different learning states that were confirmed by optogenetic tests (Hinojosa et al., 2026).
We simplified Fig. 7 for clarity but we will make it more accurate and explain it more in detail in the legend, including connections between interneurons.
Interpreting high-dimensional parameter spaces can be challenging. In this study, we focused on low-dimensional summaries of the parameter space (e.g., average connection weights and their distributions across populations), which revealed consistent and interpretable differences between sensitizing and depressing neurons. Importantly, our conclusions do not rely on individual parameter values, but rather on systematic differences across populations that are robust across solutions. Additionally, we ran clustering analysis and found that there is no parameter that can be removed. We focused, therefore, on the larger and more robust differences. We will explore additional dimensionality reduction approaches and include these results if they provide further insight beyond the current analyses.
Finally, the change in weights was calculated with equation 4, in which the weight from locomotion and rest, obtained through independent fits, were used to calculate the relative change from rest to locomotion. These were either connection weights (equation 2) which consider the strength of the connection between cell j and i, or synaptic weights (equation 3) which express the weight of individual synapses by dividing connection weights by the number of presynaptic cells and probability of connection. This distinction arises because we used average traces from all the neurons imaged to fit the model, requiring considering the number of cells to know the strength of individual synapses. We will add this explanation in the results and methods sections.
(4a) Model parameters were reduced differently for locomotion and rest (Figure 4). We suggest evaluating the results for locomotion and rest using the same chi-square value of 3 for both behavioral states (at least in controls).
Thank you for this prompt, this is an important point that we tried to resolve during our analysis. We used the reduced chi-square () to evaluate model fits within locomotion and rest condition independently. As defined in equation 12, reduced chi-square is inversely proportional to the standard error of the data which is higher in the rest dataset. As a consequence, setting the same threshold across conditions would not correspond to an equivalent goodness-of-fit criterion, and would impose a disproportionately strict constraint on the condition with lower variability, where deviations between model and data are more heavily penalized. For this reason, we used condition specific thresholds to ensure comparable fit quality relative to the noise level in each condition. In addition, to enable direct comparison across conditions independent of their noise levels, we used the RMSE as a complementary metric.
References
Carandini, M. (2004). Amplification of trial-to-trial response variability by neurons in visual cortex. PLoS Biol, 2(9), E264. https://doi.org/10.1371/journal.pbio.0020264
Chance, F. S., Nelson, S. B., & Abbott, L. F. (1998). Synaptic Depression and the Temporal Response Characteristics of V1 Cells. The Journal of Neuroscience, 18(12), 4785–4799. https://doi.org/10.1523/JNEUROSCI.18-12-04785.1998
Deneux, T., Kaszas, A., Szalay, G., Katona, G., Lakner, T., Grinvald, A., Rózsa, B., & Vanzetta, I. (2016). Accurate spike estimation from noisy calcium signals for ultrafast three-dimensional imaging of large neuronal populations in vivo. Nature Communications, 7(1), 12190. https://doi.org/10.1038/ncomms12190
Dipoppa, M., Ranson, A., Krumin, M., Pachitariu, M., Carandini, M., & Harris, K. D. (2018). Vision and Locomotion Shape the Interactions between Neuron Types in Mouse Visual Cortex. Neuron, 98(3), 602–615.e608. https://doi.org/10.1016/j.neuron.2018.03.037
Heintz, T. G., Hinojosa, A. J., Dominiak, S. E., & Lagnado, L. (2022). Opposite forms of adaptation in mouse visual cortex are controlled by distinct inhibitory microcircuits. Nature Communications, 13(1), 1031. https://doi.org/10.1038/s41467-022-28635-8
Hinojosa, A. J., Dominiak, S. E., Kosiachkin, Y., & Lagnado, L. (2026). Distinct Disinhibitory Circuits Link Short-Term Adaptation to Familiarity and Reward Learning in Visual Cortex. bioRxiv, 2026.2003.2024.713929. https://doi.org/10.64898/2026.03.24.713929
Jin, M., & Glickfeld, L. L. (2020). Magnitude, time course, and specificity of rapid adaptation across mouse visual areas. J Neurophysiol, 124(1), 245–258. https://doi.org/10.1152/jn.00758.2019
Judd, C. M., Westfall, J., & Kenny, D. A. (2017). Experiments with More Than One Random Factor: Designs, Analytic Models, and Statistical Power. Annu Rev Psychol, 68, 601–625. https://doi.org/10.1146/annurev-psych-122414-033702
Lee, J., Kim, H. R., & Lee, C. (2010). Trial-to-trial variability of spike response of V1 and saccadic response time. J Neurophysiol, 104(5), 2556–2572. https://doi.org/10.1152/jn.01040.2009
Rupprecht, P., Carta, S., Hoffmann, A., Echizen, M., Blot, A., Kwan, A. C., Dan, Y., Hofer, S. B., Kitamura, K., Helmchen, F., & Friedrich, R. W. (2021). A database and deep learning toolbox for noise-optimized, generalized spike inference from calcium imaging. Nat Neurosci, 24(9), 1324–1337. https://doi.org/10.1038/s41593-021-00895-5
Varela, J. A., Sen, K., Gibson, J., Fost, J., Abbott, L. F., & Nelson, S. B. (1997). A Quantitative Description of Short-Term Plasticity at Excitatory Synapses in Layer 2/3 of Rat Primary Visual Cortex. The Journal of Neuroscience, 17(20), 7926–7940. https://doi.org/10.1523/JNEUROSCI.17-20-07926.1997



