Exposure Therapy: Enhancing fear extinction
Gradually reducing a source of fear during extinction treatments may weaken negative memories in the long term.
- Department of Psychological Sciences, Purdue University West Lafayette, United States
- Purdue Institute for Integrative Neuroscience, Purdue University West Lafayette, United States
- Center for Neurobiology of Stress Resilience and Psychiatric Disorders, Rosalind Franklin University of Medicine and Science, United States
- Discipline of Physiology and Biophysics, Chicago Medical School, Rosalind Franklin University of Medicine and Science, United States
Memories of aversive or negative events are central to the survival of animals. However, fear learning and memory can become a challenge in people with certain conditions, such as specific phobias, anxiety or PTSD. Such individuals often avoid certain activities or situations, or start to transfer their fear to new stimuli. This avoidance strategy may help reduce fear in the short term, but but it can increase distress in the long term.
One method to alleviate fear is exposure therapy, in which individuals are subjected to objects or situations they are afraid of in a safe environment, without experiencing harm (Craske et al., 2018). For example, for someone with a fear of dogs, exposure-based therapy might include looking at pictures of dogs. This will initially produce a fear response that can include physiological changes, such as perspiration and elevated heart rate, and psychological changes, such as an increase in subjective fear. After several exposures to these stimuli, both physiological and psychological indices of fear will decline.
While this ‘extinction’ method is highly effective in minimizing fear in the short term, fear reduced through this method is highly susceptible to relapse. Even patients and subjects showing no signs of fear at the end of therapy are likely to experience a rebound after a certain amount of time or when they are re-exposed to the distressing stimulus (Bouton et al., 2021; Vervliet et al., 2013).
Decades of research suggests that this re-appearance of symptoms happens because extinction creates a new inhibitory memory that competes with – rather than updates – the original fear memory. Recent learning theories suggest that learned events are assigned a ‘prediction error’: if the prediction error is large (i.e., the new experience differs greatly from the past one) a new association will be formed. But if the prediction error is small (and the new experience is similar to the past one), the existing memory will be updated or overwritten with new information. Since fear- and anxiety-based disorders are on the rise, identifying intervention and treatment strategies to mitigate fear in a long term, relapse-resistant manner is therefore of upmost importance (Jacob et al., 2021; Siegel et al., 2022).
Now, in eLife, Nathan Holmes and colleagues – including Nicholas Kennedy as first author – report evidence that supports this recent learning theory (Kennedy et al., 2024). Kennedy et al. used a classical conditioning protocol in which they conditioned rats to fear a tone (conditional stimulus) by pairing its presentation with a brief electric shock applied to the foot (unconditional stimulus). Following this training, rats responded with freezing to the tone alone.
The researchers then used two different procedures to reduce the rats’ fear. The first group received a ‘standard’ extinction procedure during which the rats were exposed to several presentations of the sound only. The second group received a new ‘gradual’ extinction procedure, during which the tone was paired with a gradually weakened shock voltage: from 0.8 to 0.4, 0.2, and 0.1 mA. While both the traditional and gradual extinction lessened the fear response, the gradual procedure was able to also reduce relapse.
Relapse was tested at different time points following extinction treatment. When animals were re-exposed to the tone two weeks after the extinction procedures, the traditional group showed an increase in fear (e.g., more freezing) while the gradual extinction group showed less fear. The gradual extinction group was also less likely to relapse even when the animals received a brief foot shock without sound (known as ‘reinstatement’) after the extinction treatment. However, both groups were similarly susceptible to relapse if there was a change in environmental conditions from conditioning to extinction, for example, changing the physical chamber or waiting a long time between conditioning and extinction (Figure 1). Thus, the efficacy of gradual extinction relies on similar contextual parameters during conditioning and extinction to optimally minimize relapse.
Figure 1
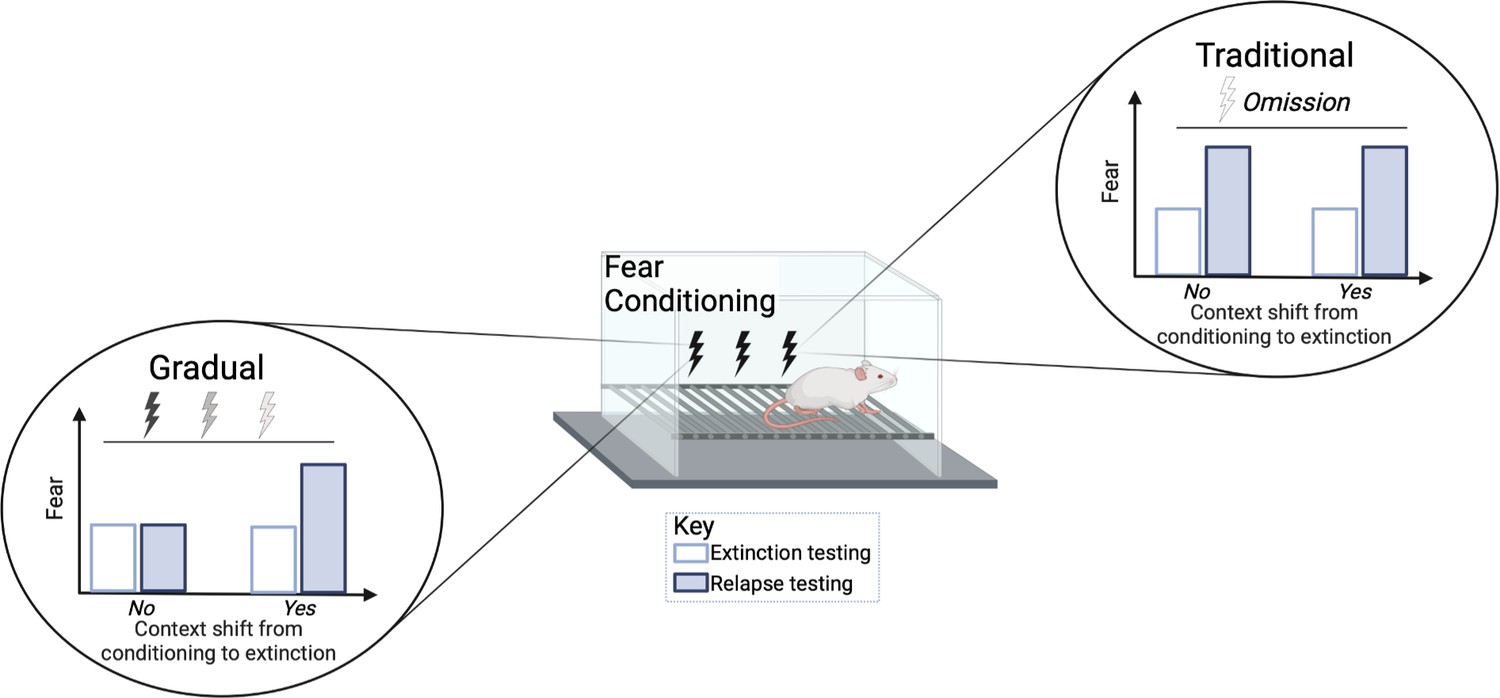
Comparison of standard versus gradual extinction protocols.
Kennedy et al. tested two different extinction procedures that reduce learned fear in rats following a classical fear conditioning protocol. The standard or traditional group (right) was exposed to a conditional stimulus (tone) only, without receiving any further electric shocks, while the gradual group (left) received a tone followed by electric shocks of a gradually reducedvoltage. In the traditional group, the fear response was initially reduced (white bar) but increased again during relapse (blue bar); this occurred regardless of whether the environmental conditions were the same (marked as no) or different (marked as yes) between extinction and conditioning. However, gradual extinction reduced relapse, but only when the environmental conditions between conditioning and extinction remained similar.
© 2024, BioRender Inc. Figure 1 was created using BioRender, and is published under a CC BY-NC-ND license. Further reproductions must adhere to the terms of this license.
Collectively, these results add to a growing literature that suggests manipulating the intensity of an unconditional stimulus (such as an electric shock) following training can have a substantial advantage over standard extinction treatments in reducing fear long-term. Kennedy et al. provide a clear explanation for these results that supports recent learning theories: too much variation between the original training shock and what occurs during extinction (as experienced in the standard group) is more likely to create a new inhibitory memory. On the other hand, a smaller change (as is experienced in the gradual extinction group) is more likely to weaken aspects of the original memory and is thus able to reduce fear long term.
These results present a promising first step in increasing the efficacy of extinction-based treatments. A next step will be to test the neural mechanisms that contribute to this effect to determine if and how this process differs from standard extinction practices, and if it could in fact be more similar to other processes relevant for updating memory, such as reconsolidation.
References
-
State-of-the-art and future directions for extinction as a translational model for fear and anxietyPhilosophical Transactions of the Royal Society of London. Series B, Biological Sciences 373:20170025.https://doi.org/10.1098/rstb.2017.0025
-
Impact of the coronavirus 2019 (COVID-19) pandemic on anxiety diagnosis in general practices in GermanyJournal of Psychiatric Research 143:528–533.https://doi.org/10.1016/j.jpsychires.2020.11.029
-
Nothing to fear but fear itself: a mechanistic test of unconscious exposureBiological Psychiatry 91:294–302.https://doi.org/10.1016/j.biopsych.2021.08.022
-
Fear extinction and relapse: state of the artAnnual Review of Clinical Psychology 9:215–248.https://doi.org/10.1146/annurev-clinpsy-050212-185542
Article and author information
Author details
Publication history
- Version of Record published: April 15, 2024 (version 1)
Copyright
© 2024, Trask and Ferrara
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 457
- views
-
- 46
- downloads
-
- 0
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Exposure Therapy: Enhancing fear extinction
eLife 13:e97633.
https://doi.org/10.7554/eLife.97633
Further reading
-
- Neuroscience
The fragile X syndrome (FXS) represents the most prevalent form of inherited intellectual disability and is the first monogenic cause of autism spectrum disorder. FXS results from the absence of the RNA-binding protein FMRP (fragile X messenger ribonucleoprotein). Neuronal migration is an essential step of brain development allowing displacement of neurons from their germinal niches to their final integration site. The precise role of FMRP in neuronal migration remains largely unexplored. Using live imaging of postnatal rostral migratory stream (RMS) neurons in Fmr1-null mice, we observed that the absence of FMRP leads to delayed neuronal migration and altered trajectory, associated with defects of centrosomal movement. RNA-interference-induced knockdown of Fmr1 shows that these migratory defects are cell-autonomous. Notably, the primary Fmrp mRNA target implicated in these migratory defects is microtubule-associated protein 1B (MAP1B). Knocking down MAP1B expression effectively rescued most of the observed migratory defects. Finally, we elucidate the molecular mechanisms at play by demonstrating that the absence of FMRP induces defects in the cage of microtubules surrounding the nucleus of migrating neurons, which is rescued by MAP1B knockdown. Our findings reveal a novel neurodevelopmental role for FMRP in collaboration with MAP1B, jointly orchestrating neuronal migration by influencing the microtubular cytoskeleton.
-
- Biochemistry and Chemical Biology
- Neuroscience
In most mammals, conspecific chemosensory communication relies on semiochemical release within complex bodily secretions and subsequent stimulus detection by the vomeronasal organ (VNO). Urine, a rich source of ethologically relevant chemosignals, conveys detailed information about sex, social hierarchy, health, and reproductive state, which becomes accessible to a conspecific via vomeronasal sampling. So far, however, numerous aspects of social chemosignaling along the vomeronasal pathway remain unclear. Moreover, since virtually all research on vomeronasal physiology is based on secretions derived from inbred laboratory mice, it remains uncertain whether such stimuli provide a true representation of potentially more relevant cues found in the wild. Here, we combine a robust low-noise VNO activity assay with comparative molecular profiling of sex- and strain-specific mouse urine samples from two inbred laboratory strains as well as from wild mice. With comprehensive molecular portraits of these secretions, VNO activity analysis now enables us to (i) assess whether and, if so, how much sex/strain-selective ‘raw’ chemical information in urine is accessible via vomeronasal sampling; (ii) identify which chemicals exhibit sufficient discriminatory power to signal an animal’s sex, strain, or both; (iii) determine the extent to which wild mouse secretions are unique; and (iv) analyze whether vomeronasal response profiles differ between strains. We report both sex- and, in particular, strain-selective VNO representations of chemical information. Within the urinary ‘secretome’, both volatile compounds and proteins exhibit sufficient discriminative power to provide sex- and strain-specific molecular fingerprints. While total protein amount is substantially enriched in male urine, females secrete a larger variety at overall comparatively low concentrations. Surprisingly, the molecular spectrum of wild mouse urine does not dramatically exceed that of inbred strains. Finally, vomeronasal response profiles differ between C57BL/6 and BALB/c animals, with particularly disparate representations of female semiochemicals.




{kind=link}