Sir2 phosphorylation through cAMP-PKA and CK2 signaling inhibits the lifespan extension activity of Sir2 in yeast
- Chungnam National University, Republic of Korea
- Chung-Ang University, Republic of Korea
- Korea Research Institute of Bioscience and Biotechnology, Republic of Korea
Abstract
Silent information regulator 2 (Sir2), an NAD+-dependent protein deacetylase, has been proposed to be a longevity factor that plays important roles in dietary restriction (DR)-mediated lifespan extension. In this study, we show that the Sir2's role for DR-mediated lifespan extension depends on cAMP-PKA and casein kinase 2 (CK2) signaling in yeast. Sir2 partially represses the transcription of lifespan-associated genes, such as PMA1 (encoding an H+-ATPase) and many ribosomal protein genes, through deacetylation of Lys 16 of histone H4 in the promoter regions of these genes. This repression is relieved by Sir2 S473 phosphorylation, which is mediated by active cAMP-PKA and CK2 signaling. Moderate DR increases the replicative lifespan of wild-type yeast but has no effect on that of yeast expressing the Sir2-S473E or S473A allele, suggesting that the effect of Sir2 on DR-mediated lifespan extension is negatively regulated by S473 phosphorylation. Our results demonstrate a mechanism by which Sir2 contributes to lifespan extension.
https://doi.org/10.7554/eLife.09709.001eLife digest
We know that cutting calorie intake through a restricted diet can slow down the aging process and prolong the lives of many organisms ranging from yeast to mammals. Calorie restriction also has protective effects on various age-related diseases including neurodegenerative disorders, cardiovascular disease, and cancer. Many studies suggest that we may mimic the beneficial effects of calorie restriction by controlling the activities of some proteins involved in the aging process.
An enzyme called Sir2 is required for calorie restriction to be able to increase lifespan. This enzyme modifies proteins called histones, which are used to package DNA inside cells. In yeast, Sir2 modifies the histones in such a way that the genes contained in that section of DNA are inactivated (or ‘silenced’). As the yeast cells age, the activity of Sir2 declines, which allows these genes to become active and contribute to the aging process. However, when yeast cells are grown in the presence of little sugar—which mimics caloric restriction—Sir2 is activated and this restores gene silencing.
It is not clear how Sir2's ability to silence these genes contributes to prolonged lifespan. Kang et al. studied the role of Sir2 in yeast and observed that one of the genes that Sir2 inactivates is called PMA1. This gene encodes a protein that is known to restrict the lifespan of yeast cells. Further experiments show that other proteins attach or remove molecules called phosphate groups from Sir2 to regulate its activity. Sir2 is inactivated when a phosphate group is attached, and active in the absence of phosphate. Under a reduced diet, the proteins that add phosphate to Sir2 are inactive, which allows Sir2 to become active and reduce the expression of the PMA1 gene.
These results show that Sir2 fine-tunes the expression of PMA1 and other age-related genes and that the attachment of phosphate groups to Sir2 by other proteins interferes with this regulation. The next challenges will be to identify the proteins responsible for attaching phosphate groups to Sir2, and to find out how they work.
https://doi.org/10.7554/eLife.09709.002Introduction
Sirtuins, a highly conserved family of nicotinamide adenine dinucleotide (NAD)-dependent protein deacetylases, have been implicated as a key metabolic sensor to link dietary restriction (DR) with lifespan extension in yeast, worms, flies, and mice (Longo and Kennedy, 2006). In yeast, silent information regulator 2 (Sir2), the founding member of the sirtuin family, mediates transcriptional silencing at the ribosomal DNA (rDNA) locus, mating type loci and telomeres by deacetylating the acetylated lysine 16 of histone H4 (Rine and Herskowitz, 1987; Hecht et al., 1996; Smith and Boeke, 1997; Imai et al., 2000). Sir2 is thought to affect replicative aging of yeast cells by repressing recombination and instability at the rDNA (Sinclair and Guarente, 1997; Falcon and Aris, 2003; Ganley et al., 2009; Stumpferl et al., 2012; Kwan et al., 2013). In addition, Sir2 may regulate lifespan by deacetylating H4K16 at sub-telomeric regions (Dang et al., 2009) and by asymmetrically segregating damaged proteins and cellular organelles between mother and daughter cells (Aguilaniu et al., 2003; Erjavec and Nystrom, 2007; McFaline-Figueroa et al., 2011). However, whether Sir2 mediates lifespan extension by DR in yeast is still under debate. Despite lots of reports supporting the role of Sir2 in DR-mediated lifespan extension (Lin et al., 2000, 2002; Anderson et al., 2003; Lin et al., 2004), numerous studies challenged the hypothesis (Jiang et al., 2002; Kaeberlein et al., 2004; Fabrizio et al., 2005; Smith et al., 2007). Similarly, in worms and flies, the role of Sir2 in lifespan extension is still controversial (Tissenbaum and Guarente, 2001; Rogina and Helfand, 2004; Lee et al., 2006; Wang and Tissenbaum, 2006; Burnett et al., 2011; Viswanathan and Guarente, 2011; Banerjee et al., 2012; Kanfi et al., 2012).
Here, we propose a new, potentially conserved, molecular mechanism of Sir2 in DR-mediated lifespan extension. Given the potential therapeutic implications of sirtuins, understanding the complex and controversial actions of sirtuins is one of central tasks in the sirtuin biology and aging field. We demonstrate that the phosphorylation of Sir2 at S473, which is regulated through cAMP-PKA and casein kinase 2 (CK2) signaling, determines the role of Sir2 in replicative lifespan (RLS).
Results
Deletion of SIR2 is associated with loss of transcriptional silencing at sub-telomeric regions, sterility in haploids, destabilization of rDNA, and shortened lifespan (Kaeberlein et al., 1999). We observed that sir2Δ mutant cells were more sensitive than the wild-type (WT) strain to high concentrations of NaCl and other monovalent cations, a phenotype not previously associated with loss of Sir2 (Figure 1A and Figure 1—figure supplement 1A–C). No sensitivity to divalent cations or osmotic stress was detected in the sir2Δ mutant (Figure 1—figure supplement 1A). The NaCl sensitivity of the sir2Δ mutant was affected neither by Fob1, which increases the amount of extrachromosomal rDNA circles, nor by pseudodiploid state of the sir2Δ mutant strain (Figure 1—figure supplement 2).
Figure 1 with 3 supplements see all
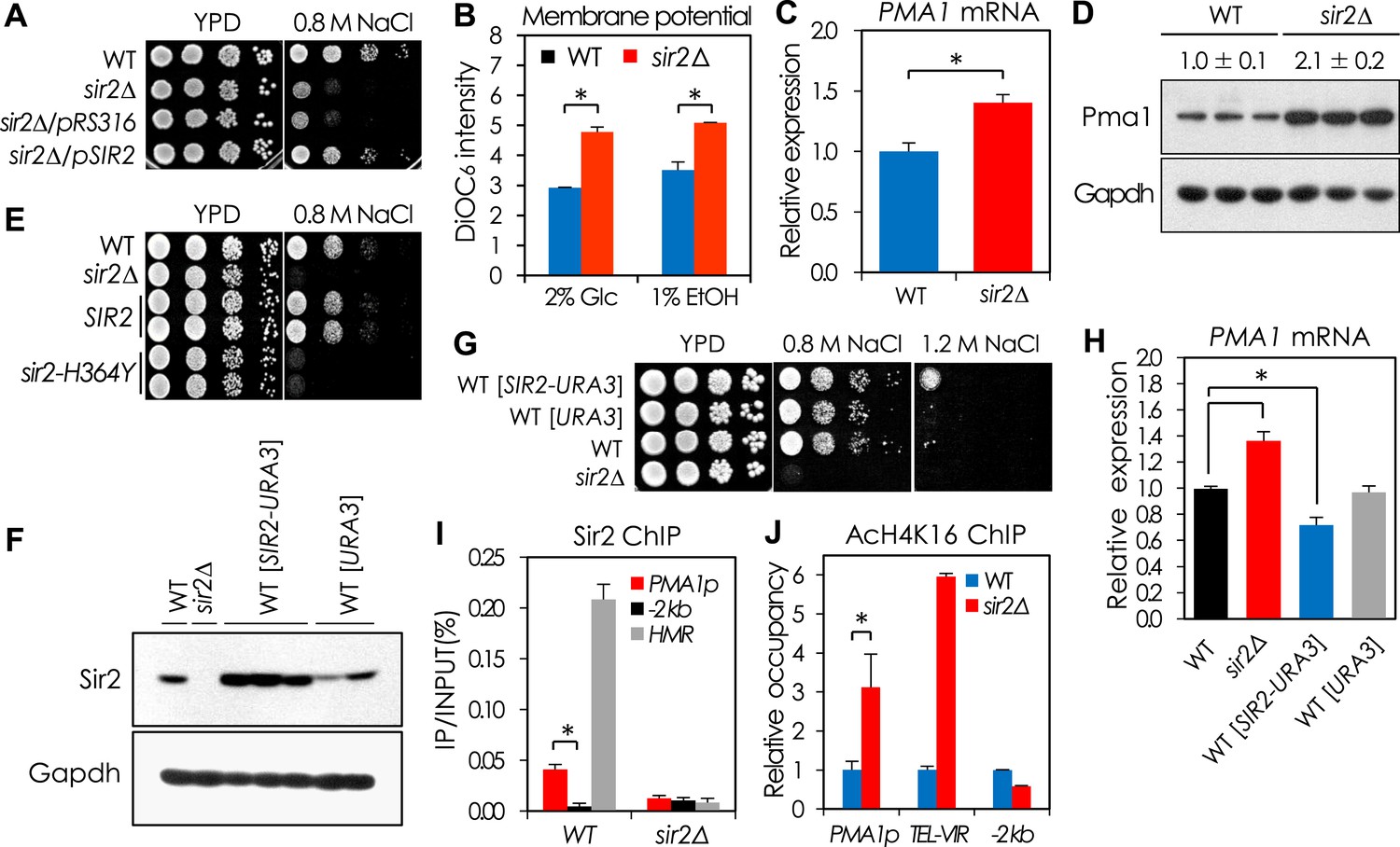
Sir2 negatively regulates PMA1 expression by deacetylating H4K16 in the PMA1 promoter.
(A) NaCl sensitivity of wild-type (WT), sir2∆, and sir2∆ cells expressing SIR2. (B) Plasma membrane potential as indicated by DiOC6 staining of WT and sir2∆ cells grown in glucose or ethanol medium (*p < 0.005). (C) PMA1 mRNA levels in WT and sir2∆ cells measured by qRT-PCR (*p < 0.001). (D) Pma1 protein levels in WT and sir2∆ cells measured by Western blot (WB). (E) NaCl sensitivity of WT, sir2∆, and sir2∆ cells carrying WT SIR2 or the sir2-H364Y mutant allele. (F) Silent information regulator 2 (Sir2) protein levels of the SIR2-overexpressing cells measured by WB. (G) NaCl sensitivity of the SIR2-overexpressing cells. (H) PMA1 mRNA levels of the SIR2-overexpressing cells measured by qRT-PCR (*p < 0.02). (I) Sir2 enrichment at the PMA1 promoter measured by Sir2 ChIP (*p < 0.001). (J) H4K16 acetylation levels at the PMA1 promoter in WT and sir2∆ cells measured by AcH4K16 ChIP (*p < 0.02). Values in (B), (C), (H), (I), and (J) represent the average of at least three independent experiments (±S.D.).
Sir2 negatively regulates PMA1 transcription by deacetylating H4K16
The plasma membrane potential (ΔΨ) is a critical determinant of cation tolerance (Navarre and Goffeau, 2000), and we reasoned that the NaCl sensitivity of the sir2Δ mutant could be due to hyperpolarization of the membrane. Indeed, relative to the WT strain, the sir2Δ mutant was more sensitive to the ΔΨ-dependent antibiotic Hygromycin B (Figure 1—figure supplement 1A) and had a higher ΔΨ (Figure 1B). The high ΔΨ in the sir2Δ mutant could result from at least two distinct mechanisms: decreased activity of the potassium uptake system (Madrid et al., 1998) or increased expression of PMA1, which encodes a plasma membrane H+-ATPase (Serrano et al., 1986). Reduced potassium uptake was unlikely to contribute to the membrane hyperpolarization because potassium chloride did not reverse the NaCl-sensitive phenotype of the sir2Δ mutant (Figure 1—figure supplement 2E). In contrast, we observed elevated levels of both PMA1 mRNA and protein levels in sir2Δ cells, which could not be rescued by expression of the enzymatically inactive sir2-H364Y allele (Tanny et al., 1999) (Figure 1C–E and Figure 1—figure supplement 1D,E). Further, overexpression of Sir2 decreased NaCl sensitivity and Pma1 expression relative to WT cells (Figure 1F–H).
Given its role in transcriptional silencing (Moazed, 2001), we speculated that Sir2 might regulate PMA1 expression by directly deacetylating H4K16 in the PMA1 promoter region. Indeed, Sir2 binding at the PMA1 promoter region was significantly greater than at another DNA fragment 2 kb upstream of the PMA1 promoter, although it was not as great as at other Sir2-regulated sites, specifically Tel-VIR and HMR (Figure 1I). Accordingly, the H4K16 acetylation level in the PMA1 promoter region was substantially higher in the sir2Δ mutant than in the WT strain (Figure 1J). We next tested whether Sas2, a major H4K16 acetyltransferase that antagonizes the effects of Sir2 on telomeric silencing and RLS in yeast (Kimura et al., 2002; Suka et al., 2002; Dang et al., 2009), opposes the Sir2-dependent H4K16 deacetylation in the PMA1 promoter region. A sas2Δ mutant showed lower Pma1 mRNA levels and less H4K16 acetylation in the PMA1 promoter region than WT cells (Heise and et al., 2012), although higher PMA1 expression and H4K16 acetylation in a sir2Δ sas2Δ mutant suggest additional H4K16 acetyltransferases, for example, Esa1 (Clarke et al., 1999; Suka et al., 2001; Chang and Pillus, 2009), replacing for Sas2 in the absence of Sir2 (Figure 1—figure supplement 3). Taken together, these data support the model that Sir2 plays an important role in maintaining the ΔΨ in yeast through regulation of PMA1 expression via deacetylation of H4K16 in the PMA1 promoter, while Sas2 antagonizes this function by acetylating H4K16 at this site.
cAMP-PKA signaling inhibits Sir2 activity for the transcriptional repression of PMA1 through serine 473 phosphorylation
Prior studies have shown that hyperactivation of the cyclic AMP (cAMP)-dependent protein kinase A (PKA) results in sensitivity to cellular stresses (Stanhill et al., 1999) and NaCl (Figure 2—figure supplement 1), and PKA signaling has been proposed to negatively regulate Sir2 in response to glucose availability (Lin et al., 2000). To examine the possibility that PKA is important for regulation of PMA1 by Sir2, we deleted PDE2, encoding a high-affinity cAMP phosphodiesterase, which increased the intracellular cAMP level without affecting Sir2 expression (Figure 2—figure supplement 1D–H). The pde2Δ mutant was as sensitive to NaCl as the sir2Δ mutant and increased the Pma1 mRNA level as high as the sir2Δ mutant (Figure 2A,B). Furthermore, the addition of 8-Bromo-cAMP, a non-hydrolyzable cAMP analog, into the culture medium mimicked the effect of the SIR2 deletion on PMA1 expression (Figure 2C). To examine whether PKA is essential for PMA1 regulation, we deleted TPK1, TPK2, and TPK3, the genes encoding the catalytic subunits of PKA, from the pde2Δ mutant strain. The Pma1 mRNA level in the pde2Δ tpk1/2/3Δ mutant strain was similar to that in WT (Figure 2D), therefore, we concluded that cAMP-PKA signaling controls the ability of Sir2 to regulate PMA1 transcription.
Figure 2 with 3 supplements see all
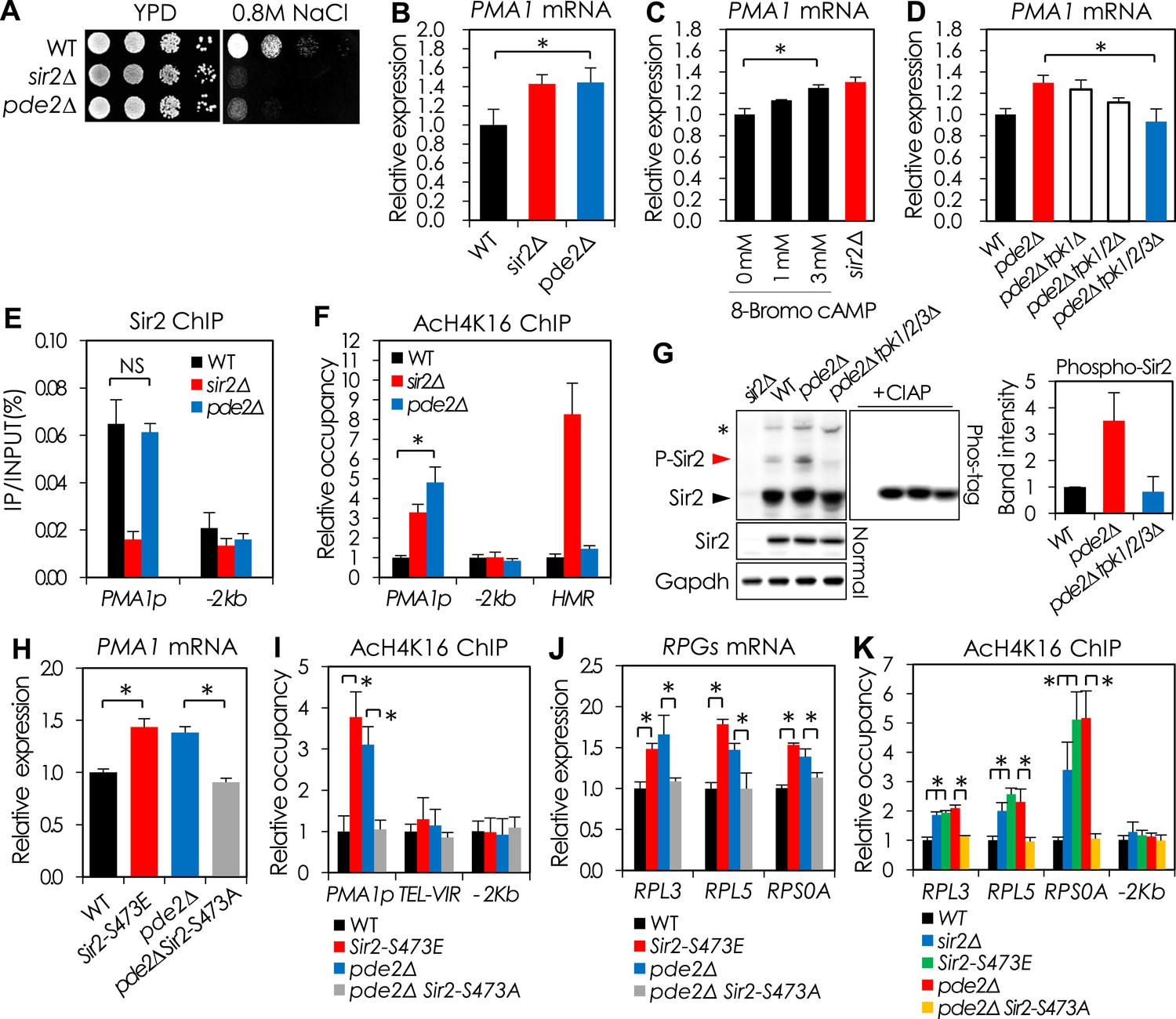
cAMP-PKA signaling inhibits Sir2 activity for the transcriptional repression of PMA1 and RPGs through serine 473 phosphorylation.
(A) Effects of PDE2 deletion on NaCl sensitivity. (B) Effects of PDE2 deletion on PMA1 expression measured by qRT-PCR (*p < 0.01). (C) Effects of 8-Bromo-cAMP addition on PMA1 expression measured by qRT-PCR (*p < 0.01). (D) Effects of TPK deletion on PMA1 expression in pde2∆ cells measured by qRT-PCR (*p < 0.01). (E) Sir2 enrichment at the PMA1 promoter in WT and pde2∆ cells measured by Sir2 ChIP (NS, not significant). (F) H4K16 acetylation levels at the PMA1 promoter in WT, sir2∆, and pde2∆ cells measured by AcH4K16 ChIP (*p < 0.001). (G) Sir2 phosphorylation levels in WT, pde2∆, and pde2∆ tpk1/2/3∆ cells analyzed by Phos-tag SDS-PAGE and WB. Arrowheads indicate cAMP-PKA-dependent phosphorylated (red) and non-phosphorylated (black) Sir2. The asterisk indicates cAMP-PKA-independent phosphorylation of Sir2. (H) Effects of SIR2-S473E or SIR2-S473A on PMA1 expression in WT and pde2∆ cells measured by qRT-PCR (*p < 0.005). (I) Effects of SIR2-S473E or SIR2-S473A on H4K16 acetylation at the PMA1 promoter in WT and pde2∆ cells measured by AcH4K16 ChIP (*p < 0.05). (J) Effects of SIR2-S473E or SIR2-S473A on the expression of ribosomal protein genes (RPGs) in WT and pde2∆ cells measured by qRT-PCR (*p < 0.005). (K) Effects of SIR2-S473E or SIR2-S473A on H4K16 acetylation at the RPG promoters in WT and pde2∆ cells measured by AcH4K16 ChIP (*p < 0.05). Values in (B), (C), (D), (E), (F), (H), (I), (J), and (K) represent the average of at least three independent experiments (±S.D.).
To investigate how cAMP-PKA signaling regulates Sir2 activity in the PMA1 promoter, we measured the H4K16 acetylation level and the amount of Sir2 bound to the PMA1 promoter in the pde2Δ mutant. The Sir2 binding efficiency was unaffected, but the H4K16 acetylation level was increased in the pde2Δ mutant (Figure 2E,F), suggesting that active cAMP-PKA signaling inhibits Sir2 activity but not Sir2-binding efficiency. To investigate how cAMP-PKA signaling inhibits Sir2 activity, we analyzed the phosphorylation of Sir2 in WT, pde2Δ, and pde2Δ tpk1/2/3Δ cells. The pde2Δ cells showed more Sir2 phosphorylation than the WT and pde2Δ tpk1/2/3Δ cells, suggesting that active cAMP-PKA signaling increases Sir2 phosphorylation (Figure 2G).
We examined whether Sir2 S473, a conserved phosphorylation site in all known Sir2 homologs (Gerhart-Hines et al., 2011), is responsible for the regulation of Sir2 deacetylase activity. Mutation of the S473 residue to alanine (Sir2-S473A) abolished the Sir2 phosphorylation induced by cAMP-PKA signaling (Figure 2—figure supplement 2). Moreover, the Pma1 mRNA level increased in WT cells expressing the phospho-mimetic Sir2-S473E protein and decreased in pde2Δ cells expressing the phospho-deficient Sir2-S473A protein, matching the respective levels in WT and pde2Δ cells expressing WT Sir2 (Figure 2H). Consistently, the occupancy of H4K16 acetylation in the PMA1 promoter region was higher in the WT cells expressing Sir2-S473E and lower in the pde2Δ cells expressing Sir2-S473A than that in the cells expressing WT Sir2 (Figure 2I). Those results suggest that Sir2 S473 is phosphorylated in response to cAMP-PKA signaling, and that phosphorylated Sir2 is unable to repress PMA1 transcription.
Sir2 regulates transcription of ribosomal protein genes in a cAMP-PKA-dependent manner
We observed that deletion of SIR2 or PDE2 resulted in cell size increase, which was not associated with rDNA destabilization or pseudodiploid state of the sir2Δ mutant strain (Figure 2—figure supplement 3A,B). Because ribosome biosynthesis rate can affect yeast cell size (Jorgensen et al., 2004) and Sir2 associates with actively transcribed genes (Tsankov et al., 2006; Li et al., 2013), including PMA1 and ribosomal protein genes (RPGs) (Tsankov et al., 2006), we reasoned that Sir2 might regulate the expression of ribosomal proteins. The mRNA levels of many RPGs were higher in sir2Δ and pde2Δ cells than in WT cells (Figure 2—figure supplement 3C). Because the amount of Sir2 bound to the RPL3 and RPL5 promoters was not changed in the pde2Δ mutant (Figure 2—figure supplement 3D), we thought that the cAMP-PKA-dependent phosphorylation of Sir2 is also responsible for the repression of RPGs transcription as shown in the PMA1 regulation. WT cells expressing Sir2-S473E produced more RPG (RPL3, RPL5, and RPS0A) mRNAs than WT cells expressing WT Sir2, and pde2Δ cells expressing Sir2-S473A produced fewer RPG mRNAs than pde2Δ cells expressing WT Sir2 (Figure 2J). Consistent with the increased amounts of RPG mRNAs, the relative occupancy of H4K16 acetylation in the RPG promoters was higher in the sir2Δ, pde2Δ, and SIR2-S473E cells than in the WT cells (Figure 2K). The expression of Sir2-S473E in WT cells, or that of Sir2-S473A in pde2Δ cells, reversed the cell size of each strain (Figure 2—figure supplement 3E). These observations suggest that Sir2 regulates the transcription of many RPGs in a cAMP-PKA-dependent manner.
CKA2 mediates cAMP-PKA-dependent Sir2 phosphorylation
Because the amino acid residues flanking Sir2 S473 do not constitute a PKA consensus motif, other kinases downstream of PKA may be involved in S473 phosphorylation. We screened a collection of yeast kinase–gene deletions consisting of 121 mutant strains (Figure 3—source data 1) and selected 21 kinase mutants that grew better than WT cells in medium containing 0.8 M NaCl. Among these, we focused on CKA2, KSS1, and DBF2, because those three kinases are known to localize in the nucleus and the sensitivity of each of the corresponding deletion mutants to NaCl was increased in the presence of nicotinamide, a Sir2 inhibitor (Figure 3—figure supplement 1A,B). Only the CKA2 deletion decreased NaCl sensitivity and the PMA1 and RPL3 mRNA levels of the pde2Δ mutant (Figure 3A–D and Figure 3—figure supplements 1C–F, 2), and the phospho-mimetic Sir2-S473E protein increased NaCl sensitivity, cell size, and PMA1 and RPL3 mRNA levels of the cka2Δ mutant (Figure 3E–G). The CKA2 deletion reduced the phosphorylation of Sir2, which was significantly increased in the pde2Δ mutant (Figure 3H), and the increased interaction of Cka2 with Sir2 in the pde2Δ mutant was reversed in the absence of the TPK genes (Figure 3I). Furthermore, chromatin immunoprecipitation analysis revealed that Cka2 binding to the PMA1 and RPL3 promoter regions, but not to other Sir2 target sites including Tel-VIR and HMR, was increased in the pde2Δ mutant but decreased in the pde2Δ tpk1/2/3Δ mutant (Figure 3J). Collectively, those results support the hypothesis that Cka2 works downstream of PKA to phosphorylate Sir2 bound to the PMA1 promoter region.
Figure 3 with 2 supplements see all
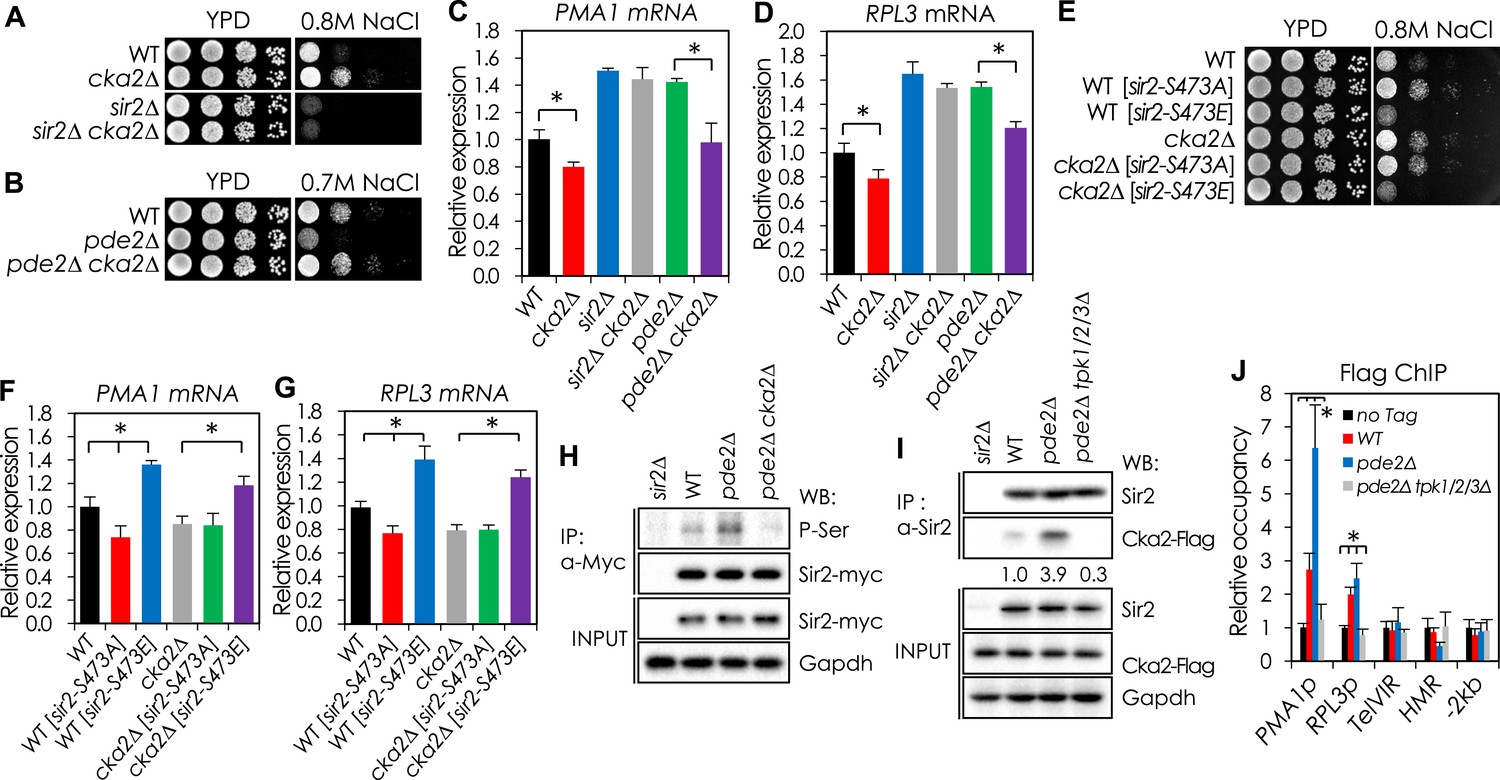
CKA2 mediates cAMP-PKA-dependent Sir2 phosphorylation to regulate the expression of PMA1 and RPGs.
(A, B) Effects of CKA2 deletion on NaCl sensitivity in WT (A), sir2∆ (A), and pde2∆ cells (B). (C, D) Effects of CKA2 deletion on the expression of PMA1 (C) and RPL3 (D) in WT, sir2∆, and pde2∆ cells measured by qRT-PCR (*p < 0.01). (E) Effects of SIR2-S473E or SIR2-S473A on NaCl sensitivity in WT and cka2∆ cells. (F, G) Effects of SIR2-S473E or SIR2-S473A on the expression of PMA1 (F) and RPL3 (G) in WT and cka2∆ cells measured by qRT-PCR (*p < 0.005). (H) Sir2 phosphorylation levels in WT, pde2∆, and pde2∆ cka2∆ cells. Myc-tagged Sir2 proteins were immunoprecipitated (IP) and analyzed by WB as indicated. (I) In vivo Sir2 and Cka2 interaction in WT, pde2∆, and pde2∆ tpk1/2/3∆ cells. Flag-tagged Cka2 proteins (Cka2-Flag) were IP and analyzed by WB. (J) Cka2-Flag enrichment at the PMA1 promoter in WT, pde2∆, and pde2∆ tpk1/2/3∆ cells measured by Flag ChIP (*p < 0.001). Values in (C), (D), (F), (G), and (J) represent the average of at least three independent experiments (±S.D.). NaCl sensitivity of the 121 kinase mutant strains used to identify kinases required for protein kinase A (PKA)-dependent Sir2 phosphorylation is available in the Figure 3—source data 1.
-
Figure 3—source data 1
NaCl sensitivity of the kinase mutant strains.
- https://doi.org/10.7554/eLife.09709.012
Sir2 S473 phosphorylation inhibits DR-mediated lifespan extension by Sir2
Since DR is known to reduce cAMP-PKA activity (Lin et al., 2000) and both PMA1 and RPGs are critical regulators of yeast RLS (Steffen et al., 2008; Ito et al., 2010; Henderson et al., 2014), we investigated the relevance of Sir2 S473 phosphorylation in the DR-mediated lifespan extension. The RLS in 2% glucose medium of the SIR2-S473A and SIR2-S473E cells was about 20% longer and about 10% shorter, respectively, than that of the WT cells (Figure 4A). The short RLS of the pde2Δ cells was increased by the SIR2-S473A allele to equal that of the WT cells in 2% glucose medium (Figure 4B). The SIR2-S473A allele had no effect on the RLS in 0.5% glucose medium (Figure 4C), however, which is in agreement with the PMA1 and RPL3 mRNA levels (Figure 4D,E). In addition, the effect of Sir2 S473 phosphorylation on RLS of yeast cells was independent of rDNA recombination/stability (Figure 4F,G). Collectively, those results suggest that the phosphorylation of Sir2 S473 inhibits DR-mediated lifespan extension by Sir2.
Figure 4

Sir2 S473 phosphorylation inhibits DR-mediated lifespan extension by Sir2.
(A) Replicative lifespan (RLS) of the strains expressing SIR2-S473E or SIR2-S473A measured by micromanipulation. The median lifespan is indicated. p < 0.0001 (WT vs sir2Δ), p = 0.0013 (WT vs sir2-S473E), p = 0.0062 (WT vs sir2-S473A). (B) Effect of SIR2-S473A on the RLS of pde2∆ cells. The median lifespan is indicated. p < 0.0001 (WT vs pde2Δ), p < 0.0001 (pde2Δ vs pde2Δ sir2-S473A). (C) RLS of WT, sir2∆, and strains expressing SIR2-S473E or SIR2-S473A grown under 0.5% glucose conditions. The median lifespan is indicated. p < 0.0001 (WT vs sir2Δ), p = 0.0006 (WT vs sir2-S473E), p = 0.5051 (WT vs sir2-S473A). (D, E) Effects of SIR2-S473A on the expression of PMA1 (D) and RPL3 (E) in cells grown under 2.0%, 0.5%, or 0% glucose conditions measured by qRT-PCR (*p < 0.005). The values represent the average of at least three independent experiments (±S.D.). (F, G) Effects of SIR2-S473E or SIR2-S473A on the RLS of fob1∆ background under 2% (F) and 0.5% (G) glucose conditions. The median lifespan is indicated. p < 0.0001 (fob1∆ vs sir2Δ under 2.0% glucose), p < 0.0001 (fob1∆ vs sir2-S473E under 2.0% glucose), p = 0.0002 (fob1∆ vs sir2-S473A under 2.0% glucose), p < 0.0001 (fob1∆ vs sir2Δ under 0.5% glucose), p < 0.0001 (fob1∆ vs sir2-S473E under 0.5% glucose), p = 0.3221 (fob1∆ vs sir2-S473A under 0.5% glucose). (H) A working model for how Sir2 regulates dietary restriction (DR)-mediated lifespan in yeast.
Discussion
In this study, we propose a new, potentially conserved, molecular mechanism of Sir2 in DR-mediated lifespan extension (Figure 4H). We show that Sir2 is able to increase RLS of yeast cells only under conditions where cAMP-PKA and CK2 signaling is not active. This result suggests that the role of Sir2 in DR-mediated lifespan extension depends on the metabolic status of cells, which is also supported by our previous report that the role of Sir2 in mediating oxidative stress resistance and chronological lifespan is growth-phase dependent (Kang et al., 2014). We speculate that a similar paradigm may exist in higher eukaryotes, although the functions and molecular mechanisms of sirtuins are much more complex, because Sir2 S473 is a conserved phosphorylation site in all known sirtuins (Gerhart-Hines et al., 2011) and numerous papers report the genetic and molecular interaction between cAMP-PKA and CK2 and SirT1 phosphorylation in mammalian cells (Kang et al., 2009; Zschoernig and Mahlknecht, 2009; Gerhart-Hines et al., 2011; Dixit et al., 2012; Park et al., 2012; Lee et al., 2014).
We also demonstrate that Sir2 fine-tunes transcription of the PMA1 and RPGs by deacetylating H4K16 in the promoter. This finding indicates that Sir2-dependent H4K16 deacetylation plays a role in the regulation of actively transcribed genes in addition to the silencing of the rDNA locus, mating type loci, and telomeres in yeast. Considering previous studies showing that Sir2 binds on actively transcribed genes including PMA1 and RPGs in yeast (Tsankov et al., 2006; Li et al., 2013), we expect this result will open a new perspective about the molecular functions of Sir2.
Pma1 protein level increases almost twofold in the absence of Sir2 (Figure 1D and Figure 1—figure supplement 1E). Recent reports indicate that vacuolar acidity is functionally linked with mitochondria and autophagy, which have a central role in the aging process (Hughes and Gottschling, 2012; Ruckenstuhl et al., 2014). And, vacuolar acidity declines in aging yeast cells because of Pma1 accumulation that reduces cytosolic protons (Henderson et al., 2014). It is thought that Pma1 expression affects yeast cell aging by changing intracellular pH, vacuolar pH, amino acid import into vacuole, and mitochondrial function (Hughes and Gottschling, 2012; Henderson et al., 2014). Thus, our study suggests a possibility that Sir2 contributes to DR-mediated lifespan extension at least in part by affecting mitochondrial function through cytoplasmic and vacuolar pH regulation in a context-dependent manner.
Materials and methods
Yeast strains and growth conditions
Request a detailed protocolThe yeast strains used in the study are listed in Supplementary file 1. The experiments were performed using the BY4741 strain, unless otherwise noted. The 10560-2B strain was used to compare and confirm the results obtained with the BY4741 strain. Deletion strains were generated by replacing each open reading frame with URA3 through homologous recombination. To confirm the effects of SIR2 mutations on stress resistance, a centromeric plasmid (pRS316) containing the SIR2 promoter (−1000 to −1), the entire SIR2 gene, and the ADH1 terminator was introduced into a sir2∆ mutant strain. To generate the SIR2 overexpression strains, a fragment that included a SIR2 promoter, an entire SIR2 gene, an ADH1 terminator, and URA3 was integrated at the URA3 locus of WT or pde2∆ strains by homologous recombination. To generate strains expressing a Sir2 protein with no deacetylase activity (sir2-H364Y) or a mutated phosphorylation site (sir2-S473E or sir2-S473A), fragments that included a SIR2 promoter, each mutated SIR2 gene, an ADH1 terminator, and URA3 were integrated at the endogenous SIR2 promoter locus in the sir2∆, pde2∆ sir2∆, or cka2∆ sir2∆ mutant strains by homologous recombination. To facilitate Western blotting and immunoprecipitation experiments, endogenous SIR2, PMA1, and CKA2 were tagged at the C-terminus with 13 MYC-URA3 or FLAG-URA3 fragment by homologous recombination. All strains generated in this study were verified using PCR and/or Western blotting.
Yeast cells were routinely grown in YPD (1% yeast extract, 2% peptone, and 2% glucose) at 30°C. Synthetic complete medium lacking uracil was used for the selection of URA+ strains. When required, transformants were plated onto solid medium containing 5′-fluoroorotic acid (1 mg/ml) to select for the loss of the URA3 marker.
Stress resistance test
Request a detailed protocolYeast cells were grown in YPD medium at 30°C for 1 day and then seeded into 25–50 ml YPD medium at an initial OD600 = 0.2 and incubated to log phase. One milliliter of the cell culture was collected, and the cells were washed with distilled water, diluted, and spotted onto regular YPD or YPD containing various concentrations of chemicals (for cationic stress: 0.15–1.2 M NaCl, 0.2–0.8 M KCl, 0.5 M LiCl, 50 mM CsCl, 5 mM–1 M CaCl2, or 300 μg/ml Hygromycin B; for osmotic stress: 1 M sorbitol or 1 M mannitol; for acidic stress: 50 mM citrate buffer [pH 3.5]). The cells were incubated for 2–3 days at 30°C, and the plates were photographed.
Measurement of plasma membrane potential
Request a detailed protocolThe relative plasma membrane potential of each strain was measured as described previously (Madrid et al., 1998). Briefly, cells were grown in YPD (OD600 < 0.5), harvested, resuspended to OD600 = 0.1 in PBS (Phosphate-buffered saline) buffer, and exposed to 1 nM DiOC6(3) cyanine dye (3,3′-dihexyloxacarbocynine iodide, Molecular Probes, Eugene, OR) for 30 min at 30°C in the dark. The fluorescence values were calculated by flow cytometer (Becton Dickinson, San Jose, CA). All measurements were made at least three times using independent preparations.
Preparation of whole-cell extracts and Western blotting
Request a detailed protocolTotal cell extracts were prepared using the TCA method (Keogh et al., 2006). Proteins were separated on 8–15% SDS-PAGE gels and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA). The membranes were probed with specific antibodies, and immuno-reactivity was detected using enhanced chemiluminescence reagent (Elpis Biotech, Korea). The primary antibodies were anti-Sir2 (1:200, Santa Cruz, Dallas, TX), anti-FLAG (1:1000, Sigma, Saint Louis, MO), anti-Myc (1:1000, Santa Cruz), anti-GAPDH (1:10,000, Acris, Germany), anti-AcH4K16 (1:2000, Upstate, Lake Placid, NY), and anti-H4 (1:0000, Millipore). Band density trace and quantification were determined using ImageJ (National Institutes of Health).
Phos-tag SDS-PAGE and immunoprecipitation
Request a detailed protocolTo detect phosphorylated Sir2 proteins, total cell extracts were separated on 6% SDS-PAGE gels containing 25 mM Phos-tag (Wako, Japan) and 100 mM MnCl2 as recommended by the supplier and analyzed by Western blot (WB) with anti-Sir2 antibody (1:200, Santa Cruz). Band intensities were quantified using ImageJ software (National Institutes of Health). Sir2 phosphorylation levels were calculated by subtracting the up-shifted Sir2 protein levels detected by Phos-tag SDS-PAGE from the total Sir2 protein levels of the same sample detected by normal SDS-PAGE. Alternatively, Sir2-13Myc proteins were immunoprecipitated (IP) using an anti-Myc antibody (1:100, Santa Cruz) and then analyzed by WB with anti-Phosphoserine antibody (1:100, Qiagen, Valencia, CA).
To investigate the interaction between Sir2 and Cka2 in vivo, cells expressing Flag-tagged Cka2 were resuspended in lysis buffer (50 mM HEPES pH 7.5, 140 mM NaCl, 1 mM EDTA, 1% Triton-100, 1 mM PMSF, 1 mM Na3VO4, and 1 mM NaF) and lysed using glass beads with vigorous vortexing. The lysates were IP overnight using anti-Sir2 antibody (1: 100, Santa Cruz) and 20 µl Protein A/G agarose beads (Santa Cruz). The beads were then washed five times with lysis buffer, and the proteins bound to the beads were analyzed by WB with anti-FLAG antibody (1:1000, Sigma).
cAMP extraction and determination
Request a detailed protocolThe method for cAMP extraction using TCA was modified from a protocol for ATP extraction (Gustafsson, 1979). Briefly, 2 × 108 cells (20 ml culture with OD600 = 0.5) were pelleted, washed, and resuspended in 1 ml cold milliQ-water. Metabolites were extracted by adding 1.2 ml TCA (0.5 M) and vigorously vortexing while the samples were kept on ice for 15 min. TCA was removed by ether extraction. The cAMP levels in the extracts were determined using cAMP Direct Immunoassay Kit (Biovision, San Francisco, CA) as recommended by the supplier. The values were normalized to dry cell weight.
RNA isolation, cDNA synthesis, and real-time PCR analysis
Request a detailed protocolTotal RNA was purified with the RNeasy Mini kit (Qiagen) and quantified by measuring the absorbance at 260 nm. From each 0.5 µg RNA sample, cDNA was synthesized using First Strand cDNA synthesis kit (Invitrogen, Carlsbad, CA) according to the manufacturer's recommendations and analyzed by quantitative RT-PCR with the oligonucleotides described in Supplementary file 2. Real-time PCR was performed with SYBR green PCR mix (Bio-Rad) and CFX connect system (Bio-Rad, Hercules, CA). Relative expression levels (normalized to ACT1) were determined using the comparative CT method.
Chromatin immunoprecipitation
Request a detailed protocolAll chromatin immunoprecipitation (ChIP) assays were performed at least in triplicate using independent chromatin preparations. The ChIP assay was carried out essentially as described (Nelson et al., 2006). Briefly, cells were fixed by 1% formaldehyde (Sigma) for 1 hr and quenched in 125 mM glycine. Cells were harvested and lysed using glass beads with vigorous vortexing, and the lysates were sonicated at 4°C for 10 cycles of 1 min on and 1 min off. Then, the supernatant was IP with anti-Sir2 antibody (Santa Cruz), anti-AcH4K16 antibody (Abcam, Cambridge, MA), or anti-FLAG antibody (Sigma) followed by incubation with BSA-coated Protein A/G agarose beads (Santa Cruz). The crosslinks were reversed, and IP DNA was precipitated and purified. Quantitative real-time PCR was performed to amplify specific regions using each oligonucleotide described in Supplementary file 2.
Cell size measurement
Request a detailed protocolCells were harvested at an OD600 ∼0.5, and the average area of the unbudded single cells was measured from microscopic images of approximately 150 cells per sample using ImageJ software (National Institutes of Health).
Screening of the yeast kinase–gene deletion collection
Request a detailed protocolTo identify kinases required for PKA-dependent Sir2 phosphorylation, we screened 121 mutant strains harboring kinase deletions in the BY4741 background for resistance to NaCl. The strains were first grown in 100 µl YPD containing 150 µg/ml G418 (Gibco-BRL, Rockville, MD) in 96-well plates at 30°C with vigorous shaking (240 rpm) for 1 day and then inoculated into 50 µl regular YPD or YPD containing 0.8 M NaCl in 96-well plates using a 96-pin replicator (V&P Scientific, San Diego, CA). The plates were incubated at 30°C with vigorous shaking (240 rpm), and the OD600 of each well was read every 1 hr for 9 hr using a microplate reader (Bio-Rad). The NaCl resistance of each strain was calculated on the basis of growth in YPD containing 0.8 M NaCl relative to that in regular YPD (Figure 3—source data 1). The NaCl-resistant mutants identified during the screening were further tested for Sir2-dependence of the NaCl resistance using a spotting assay with serial dilution on plates containing 5 mM nicotinamide (Sigma).
Yeast lifespan determination
Request a detailed protocolThe RLSs of the yeast strains were determined by micromanipulation as previously described (Kaeberlein et al., 2005) using 50–100 virgin cells grown on standard YPD plates containing 2% or 0.5% glucose. Statistical significance of the difference in the RLS between strains was determined by a two-tailed Wilcoxon rank-sum test (p < 0.05).
References
-
Esa1p is an essential histone acetyltransferase required for cell cycle progressionMolecular and Cellular Biology 19:2515–2526.
-
Sir2p-dependent protein segregation gives rise to a superior reactive oxygen species management in the progeny of Saccharomyces cerevisiaeProceedings of the National Academy of Sciences of USA 104:10877–10881.https://doi.org/10.1073/pnas.0701634104
-
Plasmid accumulation reduces life span in Saccharomyces cerevisiaeThe Journal of Biological Chemistry 278:41607–41617.https://doi.org/10.1074/jbc.M307025200
-
Metabolism of 3 alpha, 7 alpha-dihydroxy-5 beta-cholestanoic acid by rat liver in vivo and in vitroJournal of Lipid Research 20:265–270.
-
Pma1, a P-type proton ATPase, is a determinant of chronological life span in fission yeastThe Journal of Biological Chemistry 285:34616–34620.https://doi.org/10.1074/jbc.M110.175562
-
The Saccharomyces cerevisiae histone H2A variant Htz1 is acetylated by NuA4Genes & Development 20:660–665.https://doi.org/10.1101/gad.1388106
-
Attenuation of beta-amyloid-induced tauopathy via activation of CK2alpha/SIRT1: targeting for cilostazolJournal of Neuroscience Research 92:206–217.https://doi.org/10.1002/jnr.23310
-
Calorie restriction extends yeast life span by lowering the level of NADHGenes & Development 18:12–16.https://doi.org/10.1101/gad.1164804
-
Ectopic potassium uptake in trk1 trk2 mutants of Saccharomyces cerevisiae correlates with a highly hyperpolarized membrane potentialThe Journal of Biological Chemistry 273:14838–14844.https://doi.org/10.1074/jbc.273.24.14838
-
Enzymatic activities of Sir2 and chromatin silencingCurrent Opinion in Cell Biology 13:232–238.https://doi.org/10.1016/S0955-0674(00)00202-7
-
Fast chromatin immunoprecipitation assayNucleic Acids Research 34:e2.https://doi.org/10.1093/nar/gnj004
-
Four genes responsible for a position effect on expression from HML and HMR in Saccharomyces cerevisiaeGenetics 116:9–22.
-
Sir2 mediates longevity in the fly through a pathway related to calorie restrictionProceedings of the National Academy of Sciences of USA 101:15998–16003.https://doi.org/10.1073/pnas.0404184101
-
An unusual form of transcriptional silencing in yeast ribosomal DNAGenes & Development 11:241–254.https://doi.org/10.1101/gad.11.2.241
-
The yeast ras/cyclic AMP pathway induces invasive growth by suppressing the cellular stress responseMolecular and Cellular Biology 19:7529–7538.
-
Natural genetic variation in yeast longevityGenome Research 22:1963–1973.https://doi.org/10.1101/gr.136549.111
-
Overlapping and distinct functions for a Caenorhabditis elegans SIR2 and DAF-16/FOXOMechanisms of Ageing and Development 127:48–56.https://doi.org/10.1016/j.mad.2005.09.005
-
Carboxy-terminal phosphorylation of SIRT1 by protein kinase CK2Biochemical and Biophysical Research Communications 381:372–377.https://doi.org/10.1016/j.bbrc.2009.02.085
Article and author information
Author details
Funding
National Research Foundation of Korea (2010-0013086)
- Jeong-Yoon Kim
- Ki-Sun Kwon
The funder had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Matt Kaeberlein at University of Washington, Brian Kennedy at Buck Institute, and Joon Ho Lee and Cheon Ah Kim at Seoul National University for critical reading of the manuscript and valuable comments and to Won Ki Hur at Seoul National University for providing yeast deletion library. We are grateful to Ji Young Lee and Young Eun Kim for experimental assistance. This research was supported by grants from Korean Research Foundation (2010-0013086) and the Bio & Medical Technology Development Program (2013M3A9B6076413) of National Research Foundation funded by the Korean government (MSIP).
Copyright
© 2015, Kang et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,548
- views
-
- 794
- downloads
-
- 29
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 29
- citations for umbrella DOI https://doi.org/10.7554/eLife.09709
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Sir2 phosphorylation through cAMP-PKA and CK2 signaling inhibits the lifespan extension activity of Sir2 in yeast
eLife 4:e09709.
https://doi.org/10.7554/eLife.09709




{kind=link}
{kind=link}
{kind=link}
{kind=link}