Comment on "Magnetosensitive neurons mediate geomagnetic orientation in Caenorhabditis elegans"
- Research Institute of Molecular Pathology, Austria
Figures
Figure 1
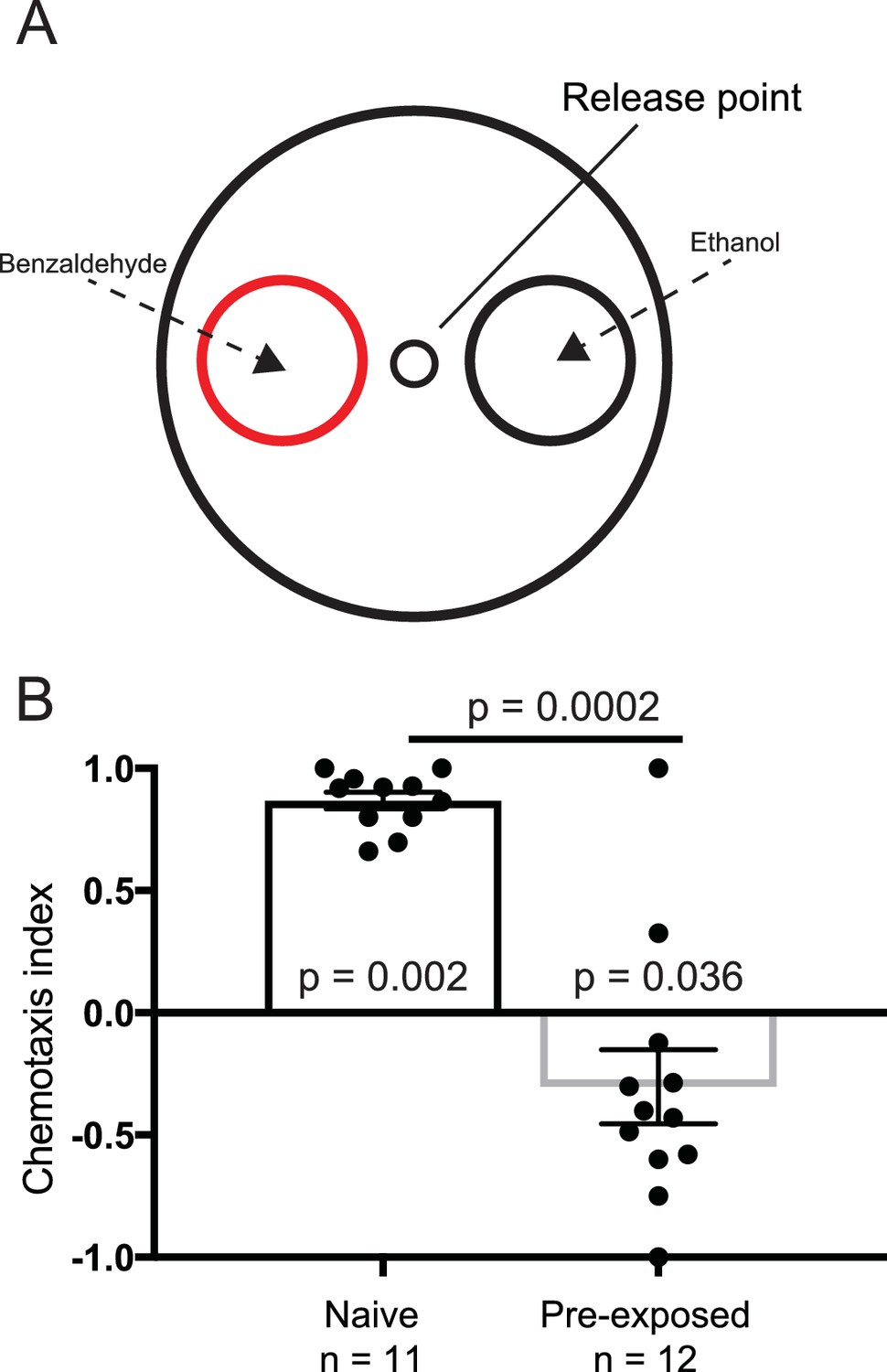
Benzaldehyde control experiment.
(A) Experimental set up for the benzaldehyde-positive control experiments. Worms were placed at the release point and given a choice between 1% benzaldehyde in ethanol, or 100% ethanol. (B) Naive worms preferentially orientated toward the benzaldehyde (n = 11, p=0.002), and away from it if pre-exposed to benzaldehyde (n = 12, p=0.036). Each data point represents the result of one independent test plate.
Figure 2
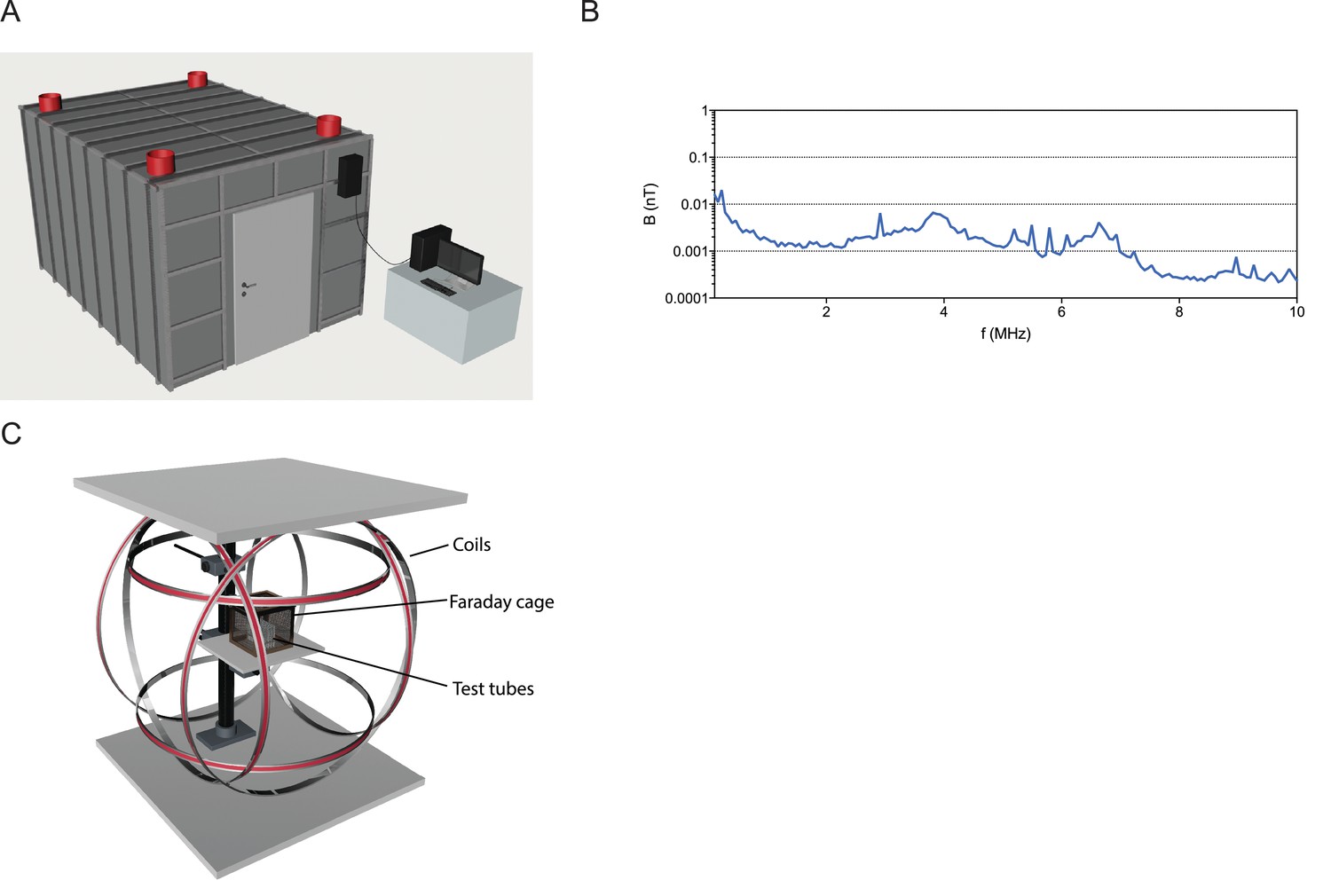
Infrastructure for magnetic experiments.
(A) All experiments were performed within a mu-metal shielded room surrounded by a 5 mm aluminum Faraday cage. DC power sources and the computer driving the Helmholtz coils were located outside this shielded room, and cables into the room were filtered for radio frequencies. (B) Graph showing the radio-frequencies present in the shielded room between 0.1 to 10 MHz are below 0.1 nT, indicative of very low levels of radio frequency contamination. (C) Experimental setup for exposure of worms to magnetic fields. Three pairs of double-wrapped Helmholtz coils surround a plastic stage in the center. Worms were placed on this stage for the vertical burrowing, horizontal plate, and magnetotaxis assays. In the burrowing assay, we surrounded the tubes by an additional small Faraday cage.
Figure 3 with 1 supplement
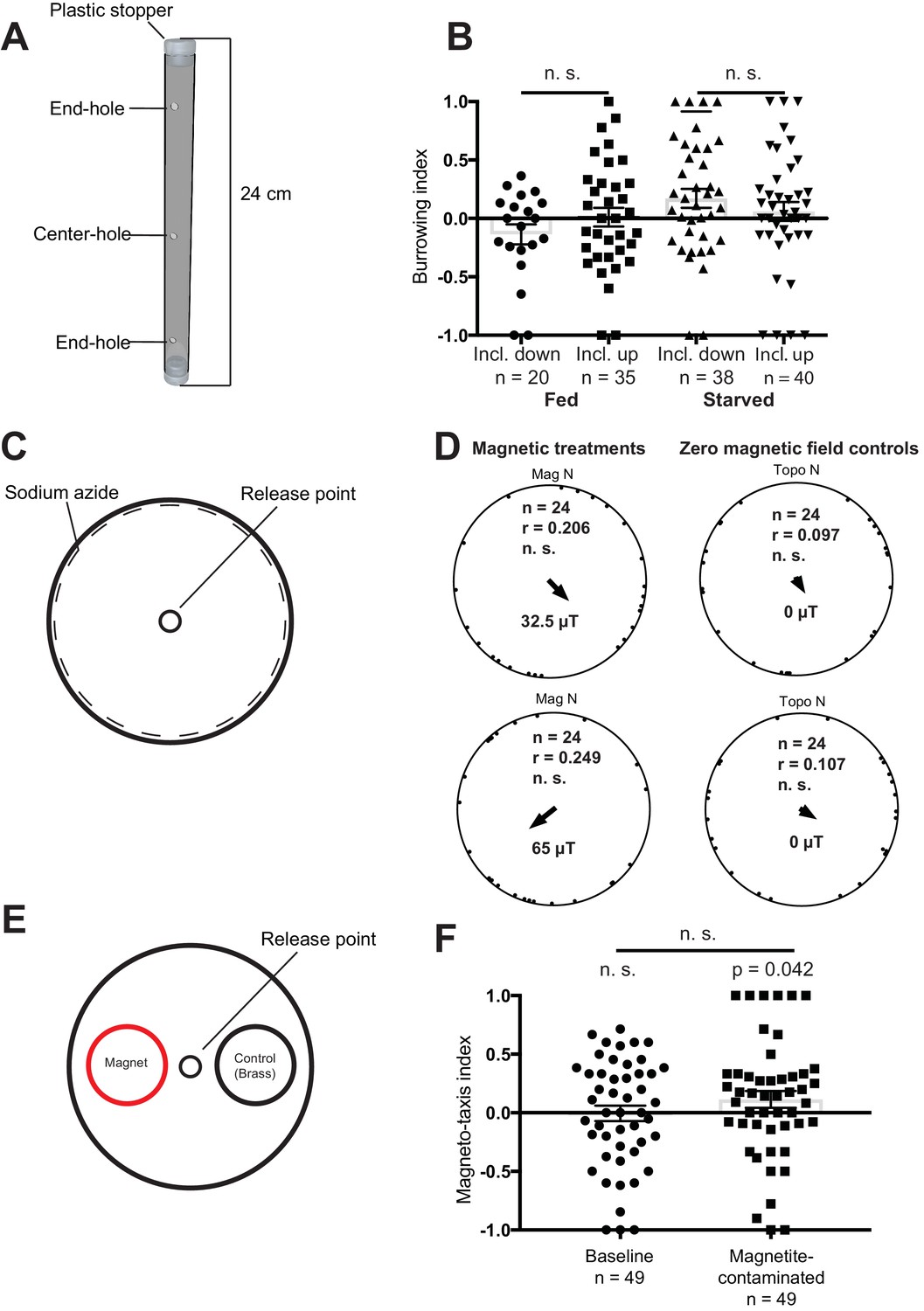
Magnetic assays and results.
(A) Diagram showing the tubes employed for the vertical burrowing assay. Worms were injected in the center hole, and NaN3 in the end-holes to immobilize them. Fed or starved worms were allowed to burrow overnight with the inclination of the magnetic field either up (59.16°) or down (−59.16°). At the conclusion of the test, the worms on either side (3 cm from the end hole) were counted and a preference index calculated. (B) Results for the vertical burrowing assay. We observed no significant difference in the burrowing index when the inclination of the magnetic field was inverted, whether the worms were fed or starved. (C) Set up for the horizontal plate assay. Worms were released in the center of the plate and allowed to move freely for 1 hr before the position and the direction of each worm relative to the center was recorded. Animals were tested in one of four magnetic directions (magnetic north pointing toward either topographic north, east, south, and west), with a field strength of 32.5 µT and 65 µT. Control experiments employed antiparallel currents resulting in a zero magnetic field. We calculated one mean orientation vector for each test plate by calculating the vector sum of all worms from this plate. (D) Results for the horizontal plate assay. We observed no directional preference when worms were exposed to either 32.5 µT or 65 µT magnetic stimuli. Each dot represents the mean worm direction for one plate, while the black arrow showing the direction and length (r) of the mean vector (radius of the circle is 1). Mag N indicates the normalized magnetic north and Topo N the topographic north. (E) Set up for the magneto-taxis assay. Worms were released in the center of a testing plate and could choose between two 3.5 cm diameter circles (goal areas) with a strong magnet (0.29 T) or a brass control underneath. Worms in each of the goal areas were counted and a preference index calculated. (F) We observed no preference for the area above the magnet, unless worms were fed bacteria contaminated with magnetite particles (p = 0.042, n = 49 plates). Error bars show standard error of the means.
Figure 3—figure supplement 1
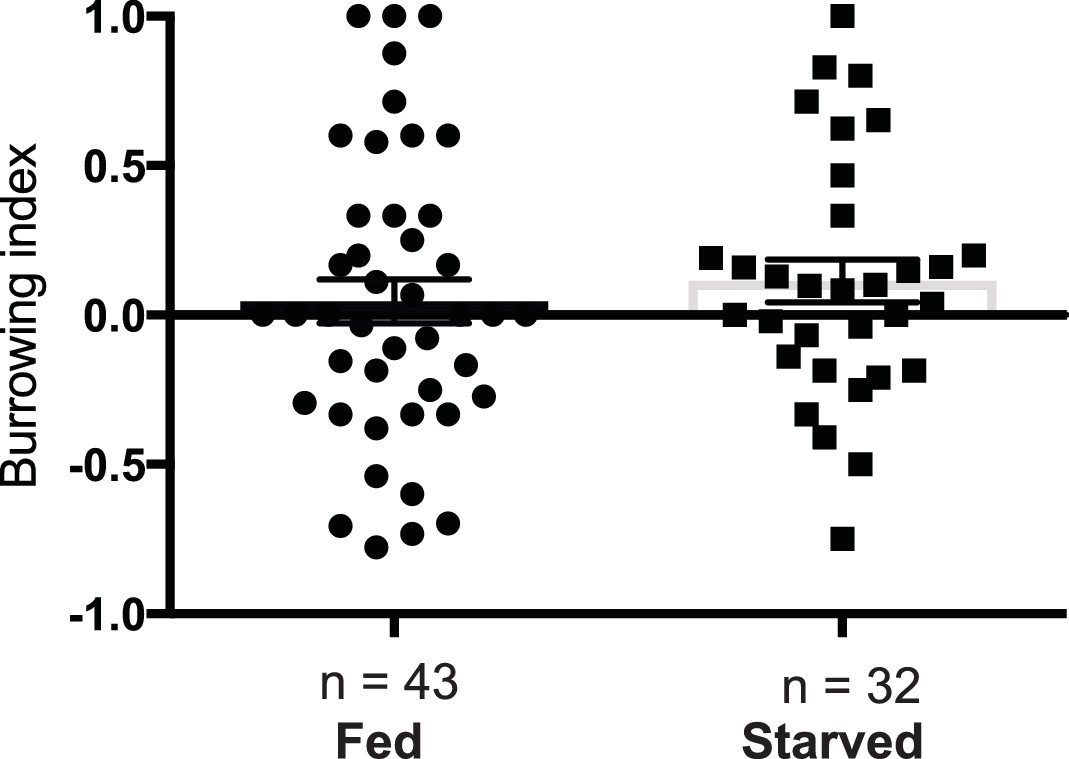
Results of the burrowing assay performed on fed and starved worms in the absence of a magnetic field.
The distribution of starved and fed worms is similar.
Figure 4
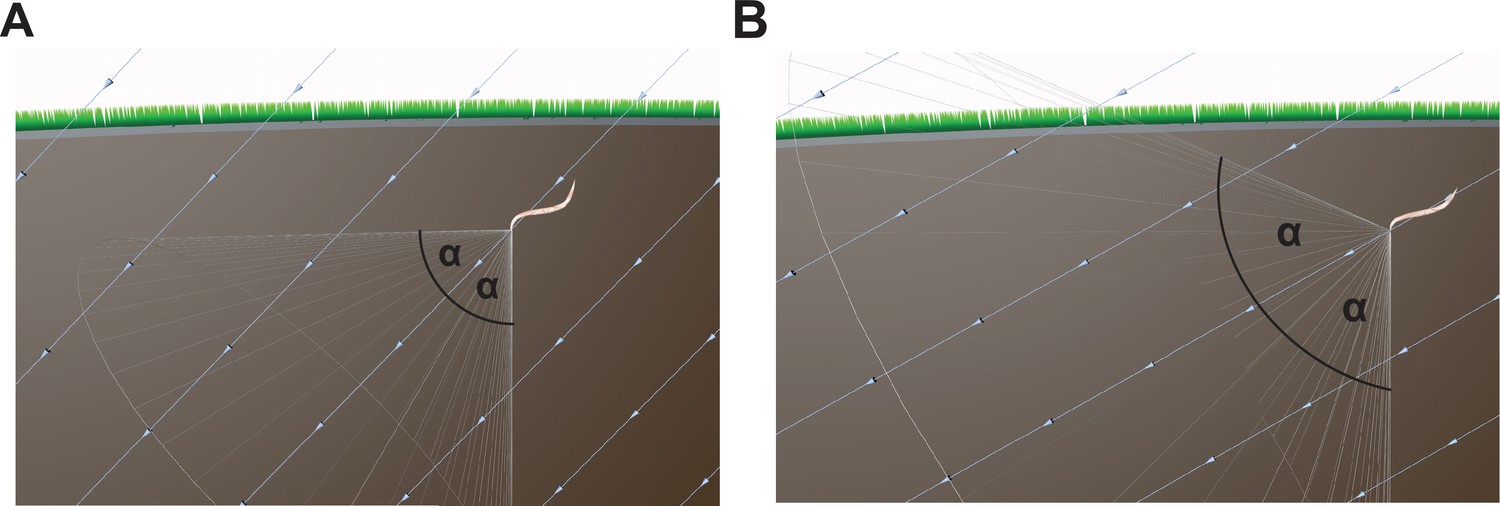
Conceptual issues with the Vidal-Gadea hypothesis.
(A) The hypothesis advanced by Vidal-Gadea and colleagues argues that nematodes exploit the inclination of the Earth's magnetic field to guide vertical movement. They propose that nematodes adopt a correction angle (α, e.g. 45°) relative to the inclination of the field, which varies depending on the latitude. However, if the worms adopt such an angle and take a fixed trajectory this is as likely to result in a worm that travels horizontally as vertically. (B) As the latitude nears the equator the correction angle increases (e.g. 60°), and consequently a worm is just as likely to translate downwards, or at an oblique angle toward the Earth's surface. The light blue lines show the magnetic field vector.
Additional files
-
Supplementary file 1
Detailed summary of statistics used for the chemotaxis as well as the magnetic assays.
- https://doi.org/10.7554/eLife.30187.007
-
Transparent reporting form
- https://doi.org/10.7554/eLife.30187.008
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Comment on "Magnetosensitive neurons mediate geomagnetic orientation in Caenorhabditis elegans"
eLife 7:e30187.
https://doi.org/10.7554/eLife.30187




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}