Climate-mediated cooperation promotes niche expansion in burying beetles
- Biodiversity Research Center, Academia Sinica, Taiwan
- National Taiwan University, Taiwan
- Columbia University, United States
- National University of Tainan, Taiwan
Figures
Figure 1
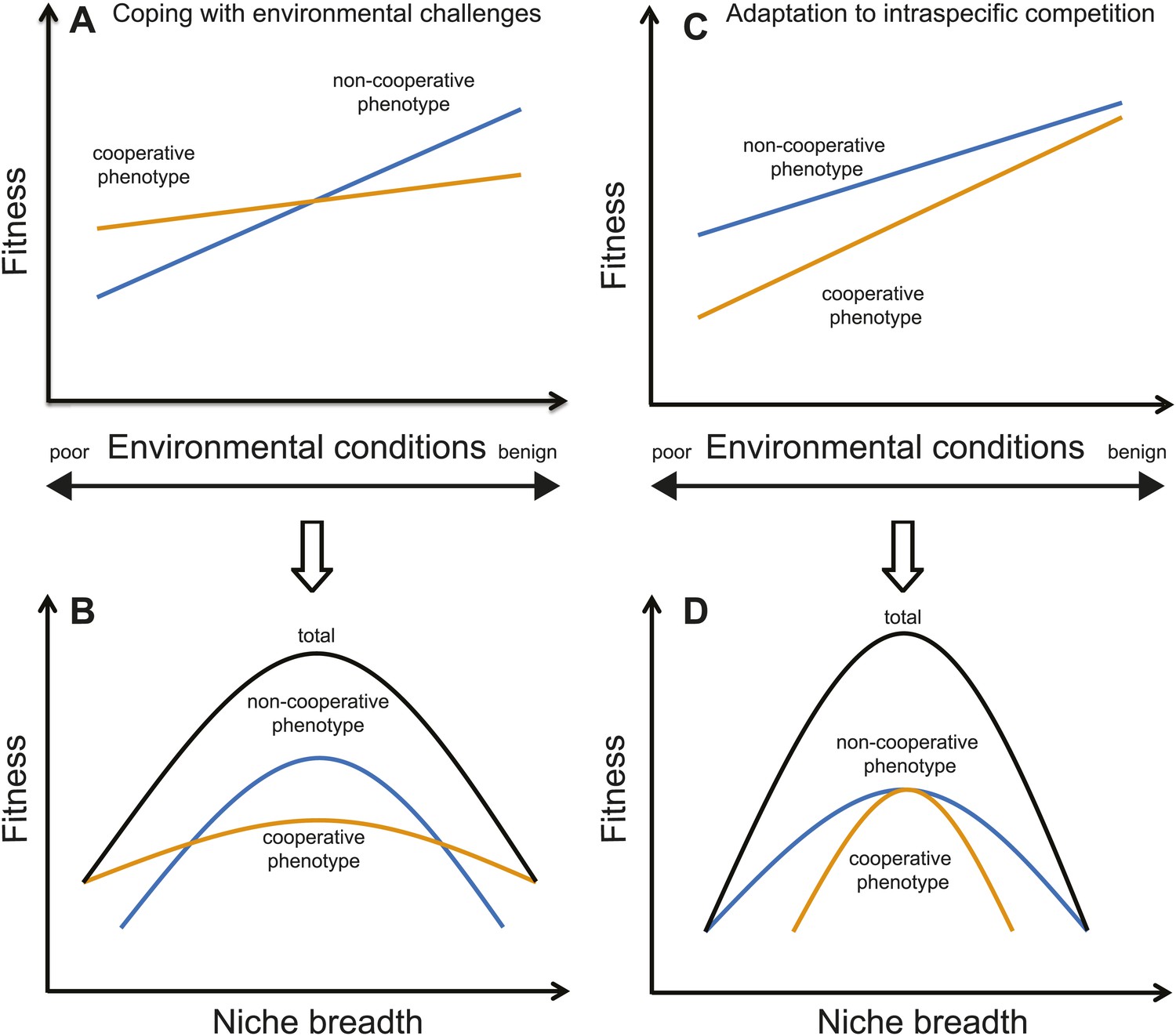
Illustration of two different causes of sociality, and their ecological consequences (i.e., niche breadth).
(A) If cooperation is for coping with harsh environments or interspecific competition, cooperative phenotypes (i.e., forming groups; orange lines) will have higher fitness than non-cooperative phenotypes in poor environments or when the pressure of interspecific competition is high. However, non-cooperative phenotypes (i.e., being solitary; blue lines) could have higher fitness in favorable environments because there are few benefits of cooperating. (B) Under such a scenario, a species' total niche breadth (black lines) is expanded due to the cooperative phenotype because a social species' total niche breadth equals to the sum of the cooperative and non-cooperative phenotypes. (C) In contrast, if cooperation is the best-of-a-bad-job strategy as a response to intraspecific challenges, the per capita reproductive success will be lower in groups than solitary pairs. This scenario often occurs when grouping occurs because of a lack of critical resources, such as when breeding territories are limited in many cooperatively breeding birds (Emlen, 1982). Therefore, cooperative phenotypes do not necessarily have higher fitness than non-cooperative phenotypes in either poor or favorable environments. (D) As a consequence, cooperative phenotypes will have little influence on the total niche breadth of a species when cooperation is a response to intraspecific challenges. Note that the trade-offs between specialist and generalist strategies occur only in the case of coping with environmental challenges or interspecific competition, and not in the case of adaptation to intraspecific competition.
Figure 2
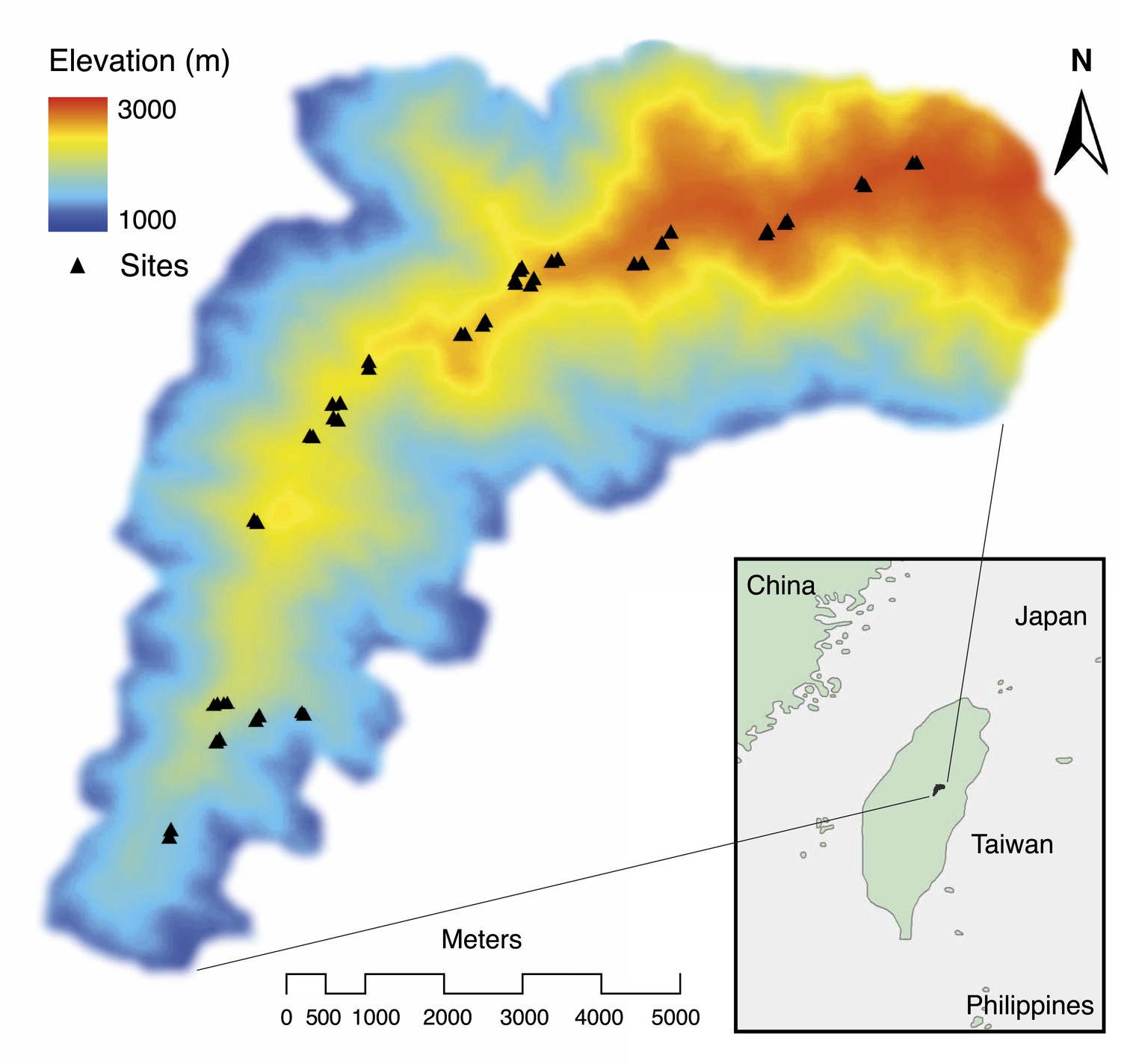
Spatial distribution of study sites (black triangles) along an elevational gradient in Nantou, Taiwan (24°5' N, 121°10' E).
https://doi.org/10.7554/eLife.02440.005
Figure 3
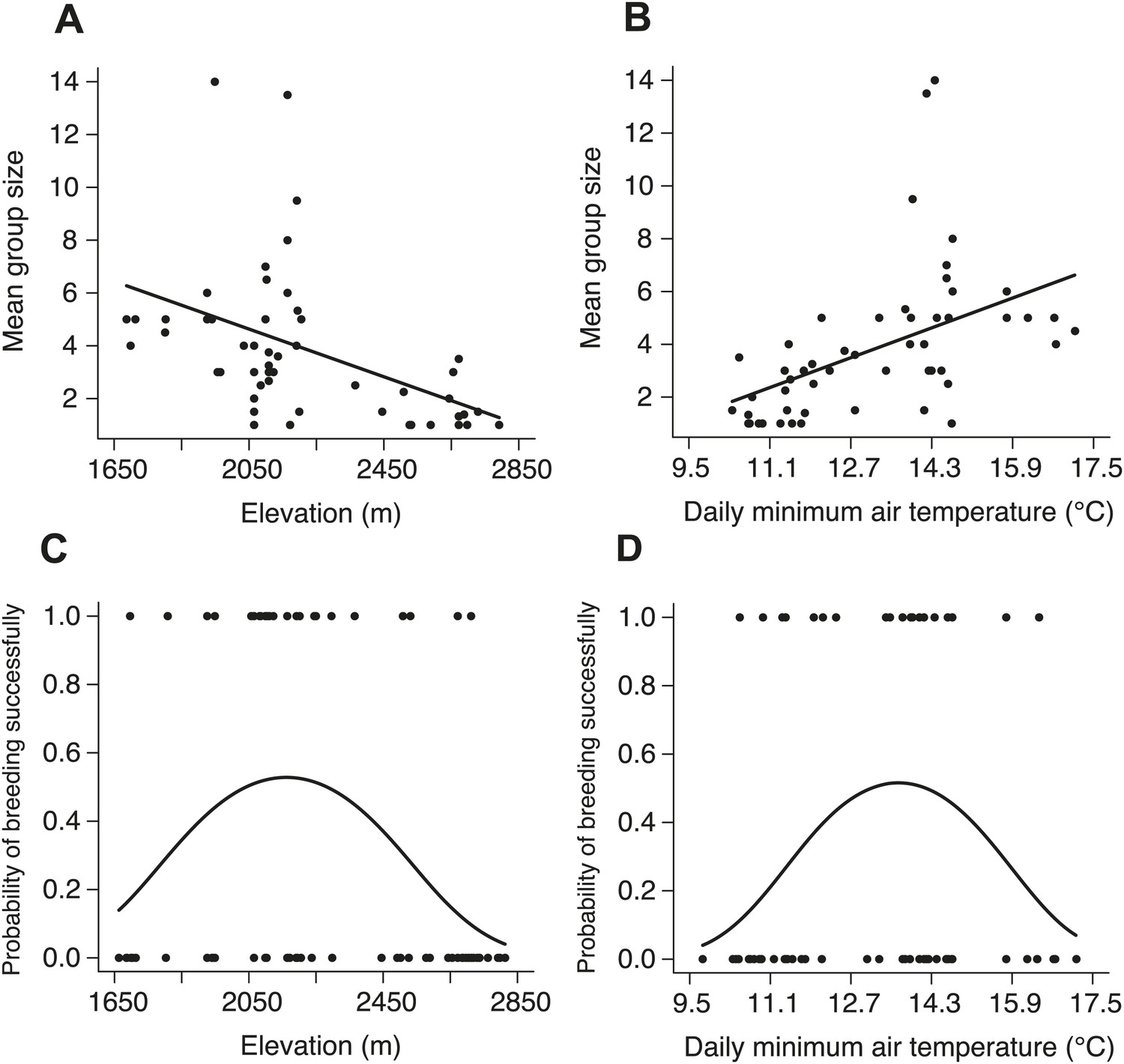
Natural patterns of group size and the probability of breeding successfully in relation to elevation and daily minimum air temperature.
Mean group size in natural populations decreased with (A) increasing elevation (χ²1 = 16.26, p<0.001, n = 54) and (B) daily minimum air temperature (χ²1 = 15.26, p<0.001, n = 53). The probability of breeding successfully in natural populations varied unimodally along (C) the elevational (χ²2 = 8.68, p=0.013, n = 70) and (D) daily minimum air temperature gradients (χ²2 = 6.37, p=0.041, n = 66).
Figure 4
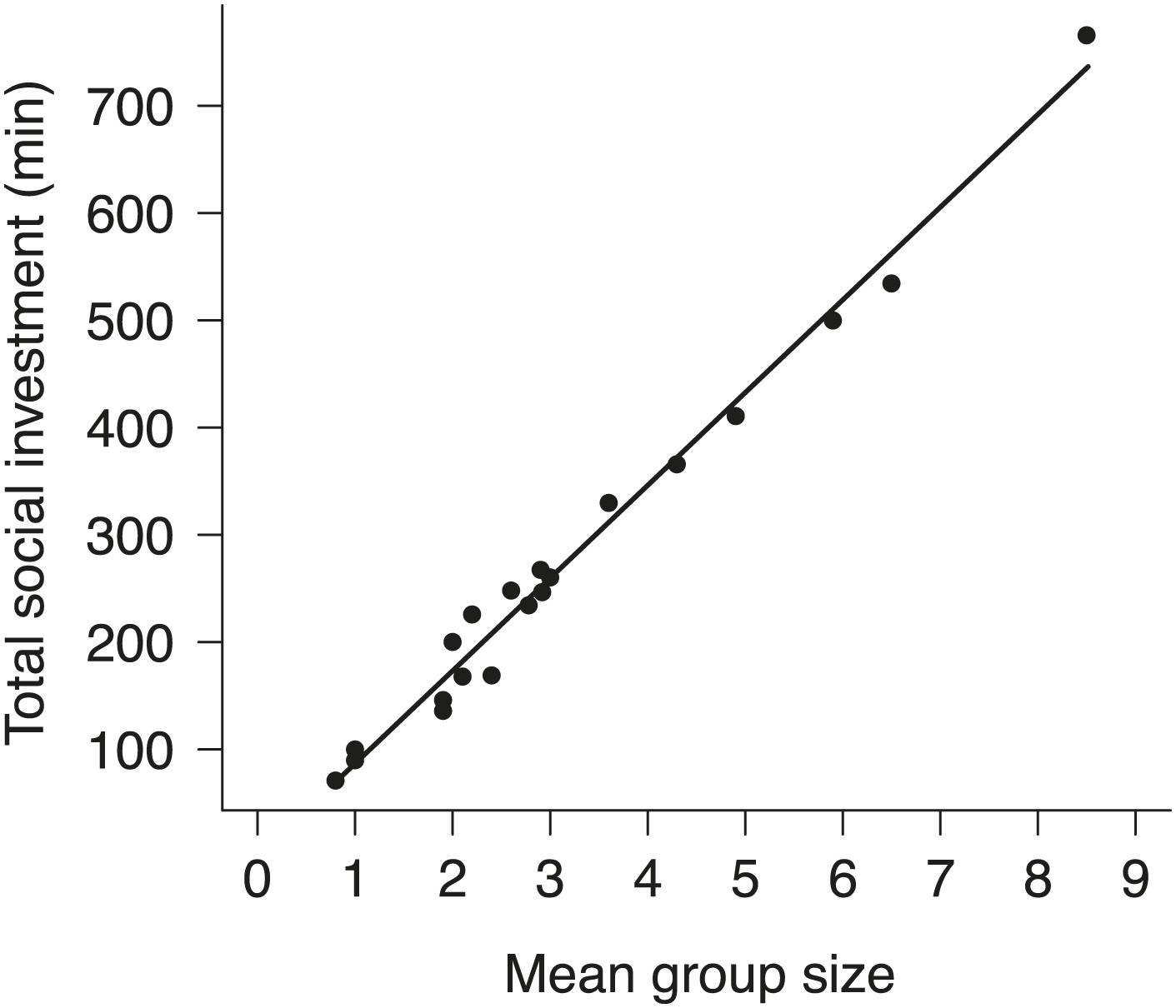
The relationship between group size and total investment in cooperative carcass processing in natural groups.
Total social investment (minutes, min) in cooperative carcass processing increased with the increasing group size (χ²1 = 1681.10, p<0.001, n = 21).
Figure 5
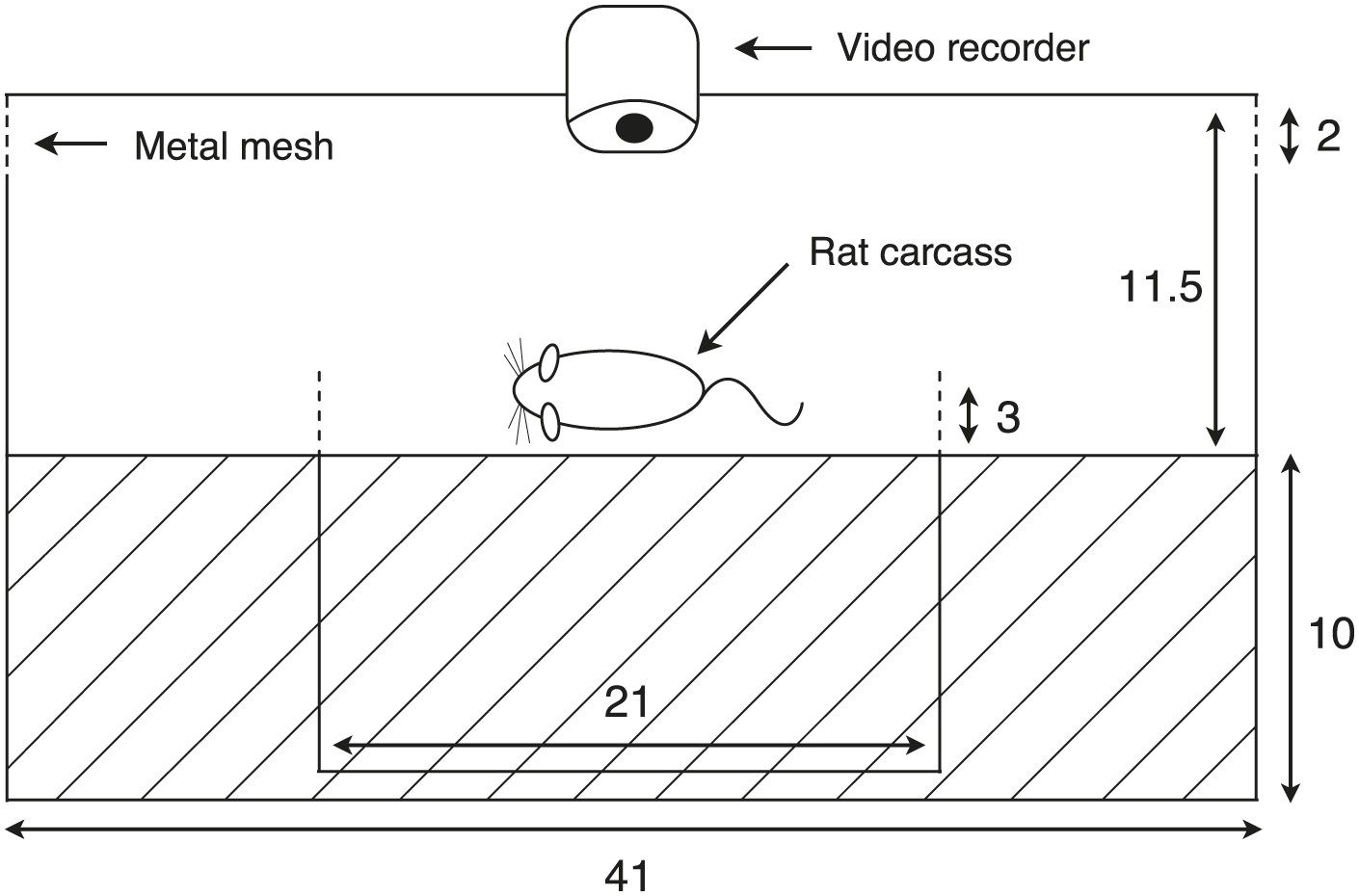
Diagram of the experimental container.
The apparatus consisted of a larger plastic container to isolate the carcass from scavengers, but beetles and flies were allowed to move freely between the chamber and the outside environment. A smaller container with a rat carcass was provided for burial in the center of the larger container. The entire burial process and behavioral assays were recorded with a video-recorder. Dashed lines represent places connected to long pipes, which allowed beetles to leave the box. Cross hatching indicates the soil layer inside the chamber (unit: cm). Our manipulation successfully created different mean group sizes even after some free-living beetles entered the chambers and some experimental beetles left (t58 = 15.08, p<0.001).
Figure 6
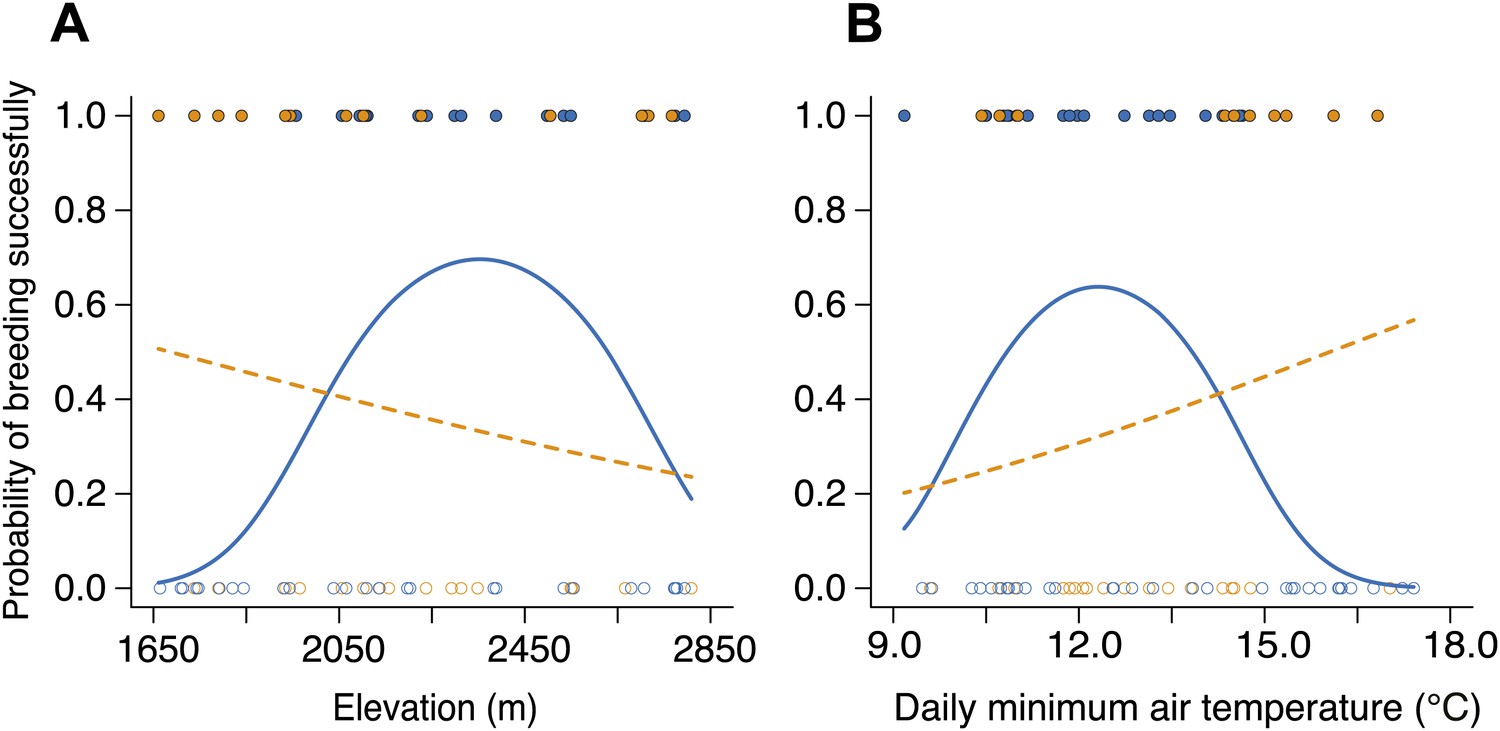
Reproductive success varied with group size along elevational and temperature gradients.
(A) The probability of breeding successfully for small (blue circles, solid line) and large groups (orange circles, dashed line) varied differently along the elevational gradient (group size × elevation interaction, χ²2 = 10.56, p=0.005, n = 92; for large groups, χ²2 = 3.19, p=0.20, n = 39; for small groups, χ²2 = 7.66, p=0.022, n = 53), with large groups having higher breeding success than small groups at lower elevations (χ²1 = 5.60, p=0.018, n = 26), but small groups having marginally higher breeding success than larger groups at intermediate elevations (χ²1 = 3.51, p=0.061, n = 53). There was no significant difference in breeding success between small and larger groups at high elevations (χ²1 = 0.04, p=0.84, n = 13). (B) The probability of breeding successfully for small and large groups also varied differently along the daily minimum air temperature gradient (group size × temperature interaction, χ²2 = 7.28, p=0.026, n = 92; for large groups, χ²2 = 1.55, p=0.46, n = 39; for small groups, χ²2 = 6.15, p=0.046, n = 53), with large groups showing higher breeding success than small groups at higher temperatures (χ²1 = 5.60, p=0.018, n = 26), but small groups having marginally higher breeding success than small groups at intermediate temperatures (χ²1 = 3.46, p=0.063, n = 53). Again, there was no significance in breeding success between small and larger groups at high temperatures (χ²1 = 0.0001, p=0.99, n = 13). Open circles indicate failed breeding attempts and closed circles indicate successful breeding events. Solid lines denote predicted relationships from GLMMs, whereas dashed lines denote statistically non-significant relationships.
Figure 7
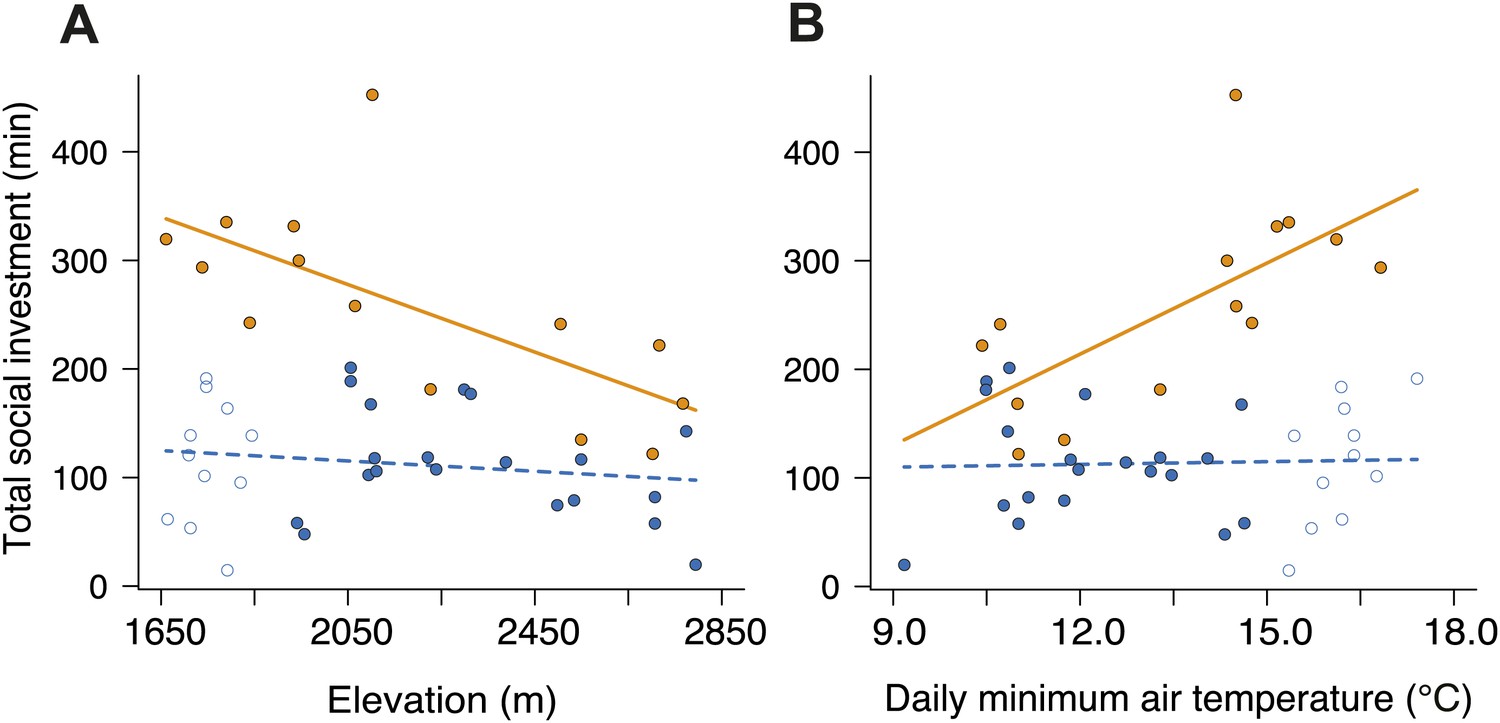
Investment in cooperative carcass processing along the elevational and temperature gradients.
Investment (minutes, min) in large (closed orange circles, solid line) and small groups (closed blue circles, successful trials; open blue circles, failed trials; dashed line) varied along the (A) elevational (group size × elevation interaction, χ²1 = 7.65, p=0.006, n = 45) and (B) daily minimum air temperature gradients (group size × temperature interaction, χ²1 = 9.90, p=0.002, n = 45) such that investment in large groups decreased with (A) increasing elevation (χ²1 = 10.30, p=0.001, n = 14) and (B) decreasing daily minimum temperature (χ²1 = 9.93, p=0.002, n = 14). There was no relationship between cooperative carcass processing and (A) elevation (χ²1 = 0.80, p=0.37, n = 31) or (B) daily minimum air temperature (χ²1 = 0.04, p=0.84, n = 31) in small groups. Solid lines denote predicted relationships from GLMs, whereas dashed lines denote statistically non-significant relationships.
Figure 8
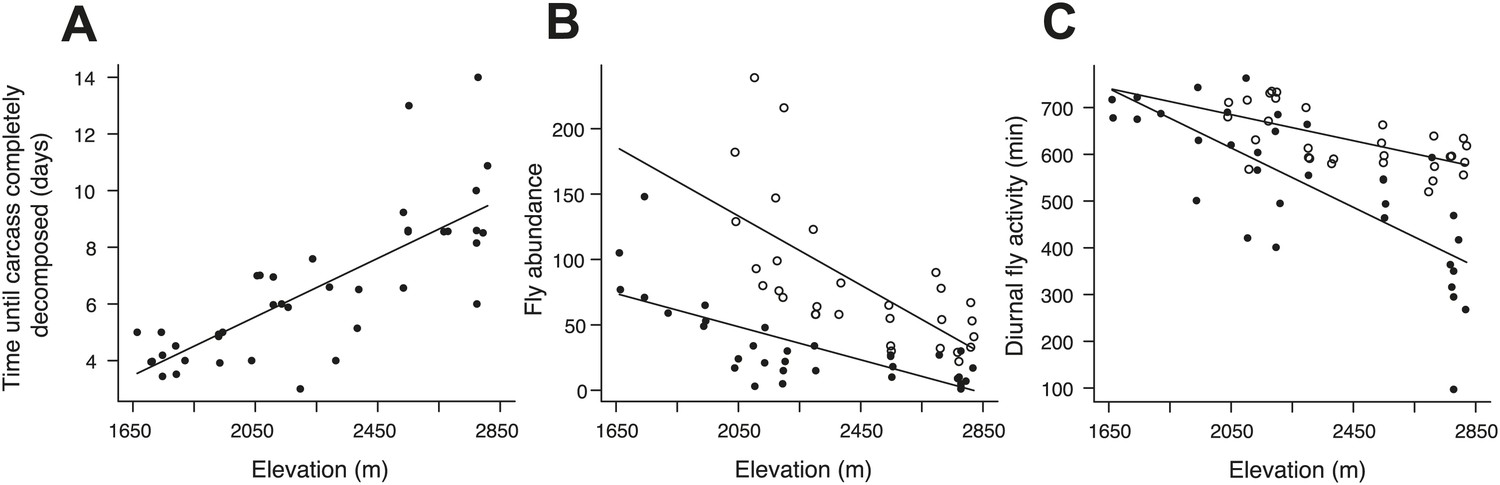
Carcass decomposition rates and the effect of experimentally heating carcasses on fly abundance and activity along the elevational gradient.
(A) The time until the carcass was completely consumed by carrion-feeding insects increased with increasing elevation (χ²1 = 50.87, p<0.001, n = 40). The control treatments (closed circles) represent the natural patterns of fly abundance and activity on carcasses. (B) Fly abundance decreased with increasing elevation (χ²1 = 21.49, p<0.001, n = 33), but heated carcass treatments (open circles) showed higher fly abundance than controls (closed circles) (χ²1 = 42.65, p<0.001, n = 55). (C) Diurnal fly activity decreased with increasing elevation (χ²1 = 39.90, p<0.001, n = 33), but flies were more active on heated carcass treatments than on controls (χ²1 = 29.85, p<0.001, n = 55). Solid lines denote predicted relationships from GLMs.
Figure 9
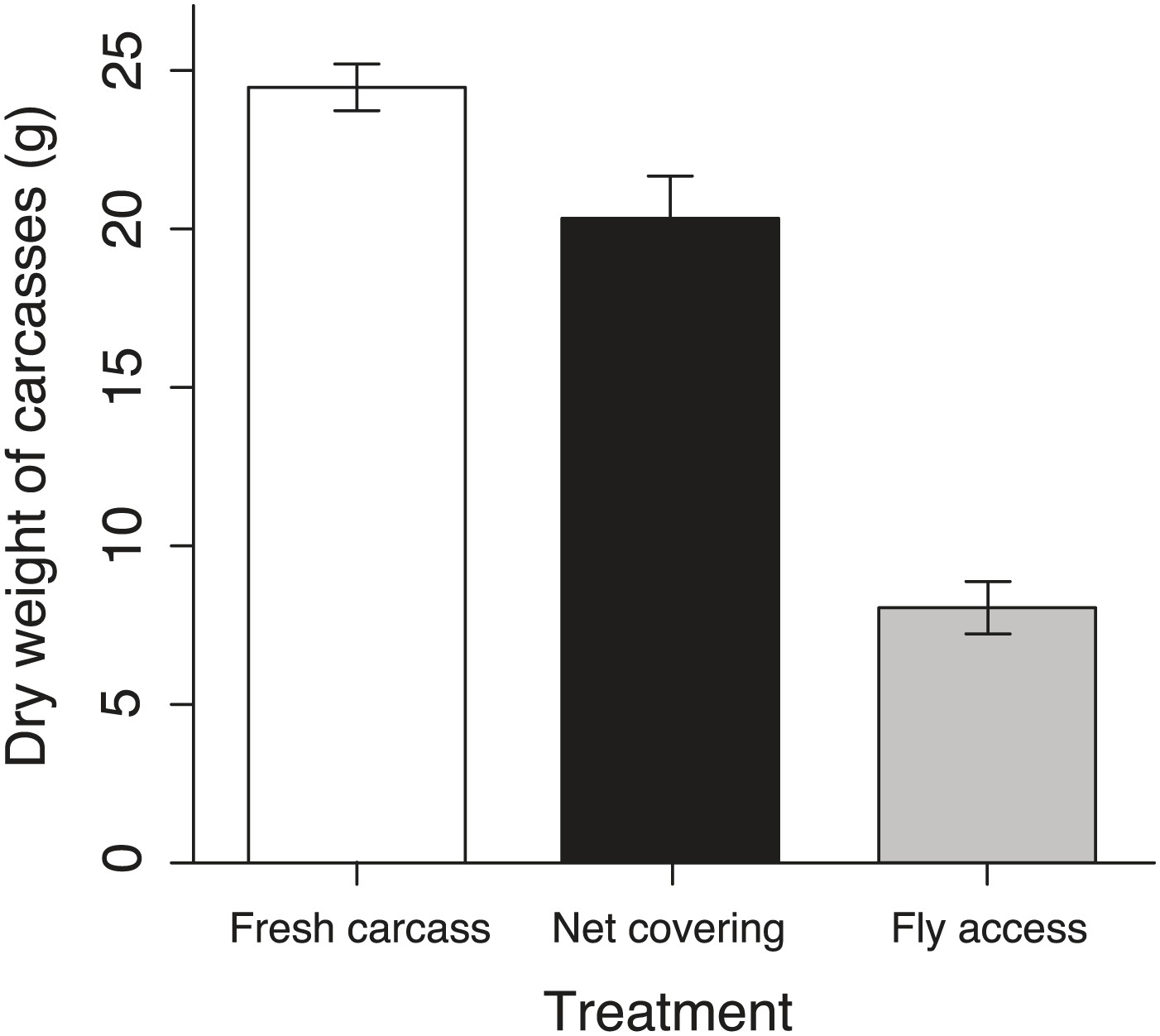
Remaining carcasses dry weight after exposure in different treatments.
The mean ± SE remaining carcass dry weights in the fresh carcass controls (white column, n = 9) were significantly heavier than those in the net covering (black column, n = 9) and fly access treatments (grey column, n = 9) (χ²2 = 145.66, p<0.001, n = 27).
Figure 10
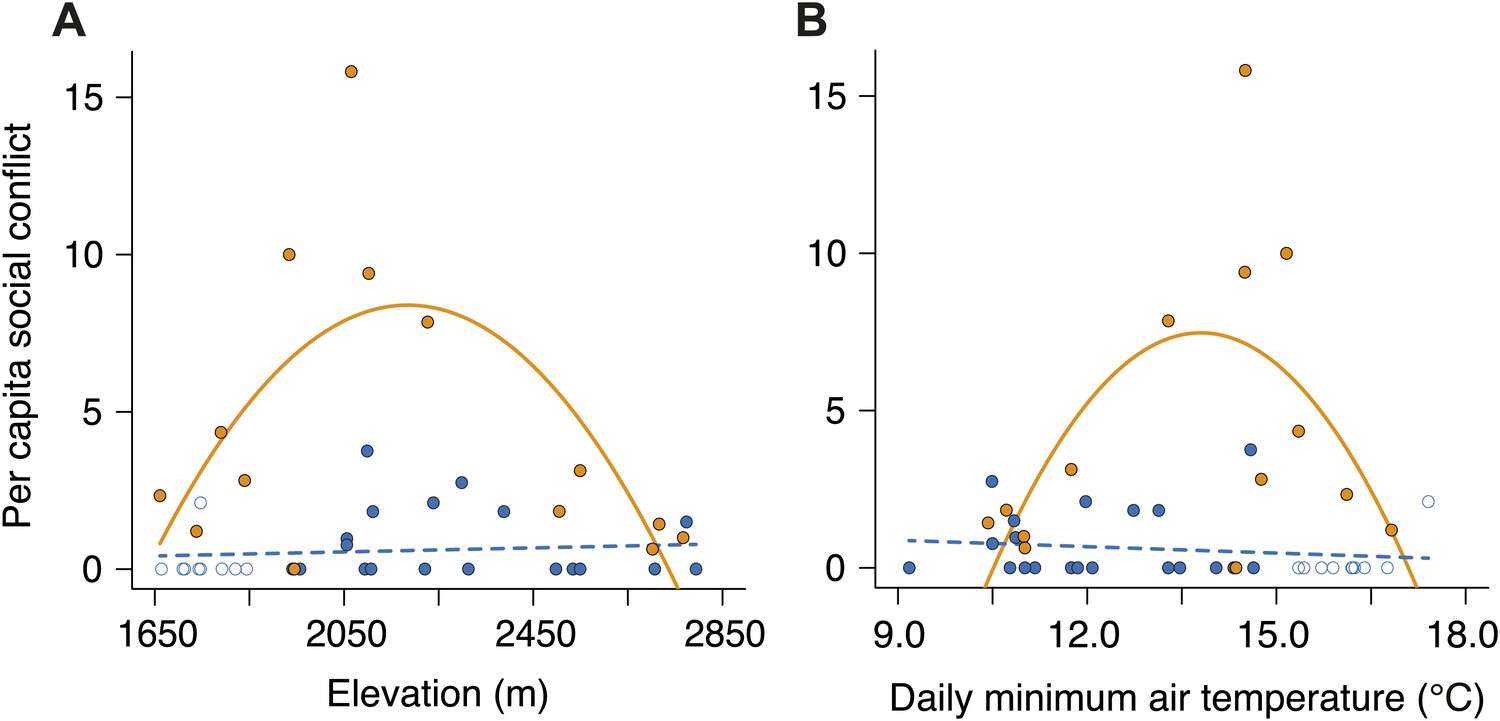
Per capita social conflict in small and large groups along the elevational and temperature gradients.
Patterns of per capita social conflict differed between small (closed blue circles, successful trials; open blue circles, failed trials; dashed line) and large groups (closed orange circles, solid line) along gradients of (A) elevation (group size × elevation interaction, χ²2 = 14.73, p<0.001, n = 45) and (B) daily minimum air temperature (group size × temperature interaction, χ²2 = 13.98, p<0.001, n = 45). In large groups, per capita social conflict varied unimodally with elevation (χ²2 = 9.11, p=0.011, n = 14) and daily minimum air temperature (χ²2 = 6.17, p=0.046, n = 14), peaking at intermediate elevations and temperatures. However, in small groups, per capital social conflict did not vary with elevation (χ²2 = 4.37, p=0.11, n = 31) or daily minimum air temperature (χ²2 = 0.73, p=0.70, n = 31).
Figure 11
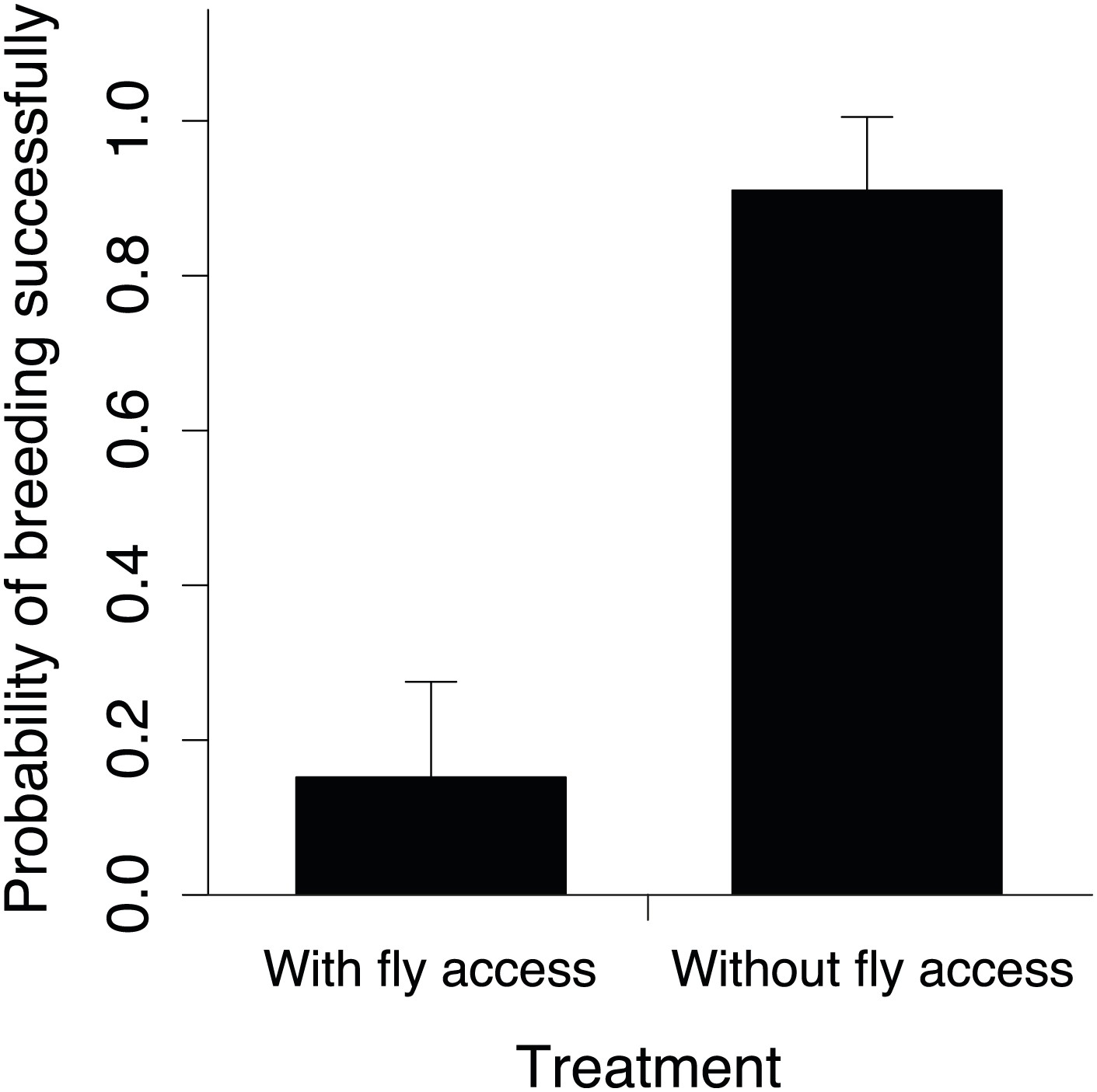
The probability of breeding successfully in relation to fly accessibility.
Mean ± SE probability of breeding successfully (GLMM fitted values) in small groups was higher when flies were excluded from carcasses (n = 18) than when they had access to carcasses (n = 23) along the elevational gradient (χ²1 = 12.06, p<0.001, n = 41).
Figure 12
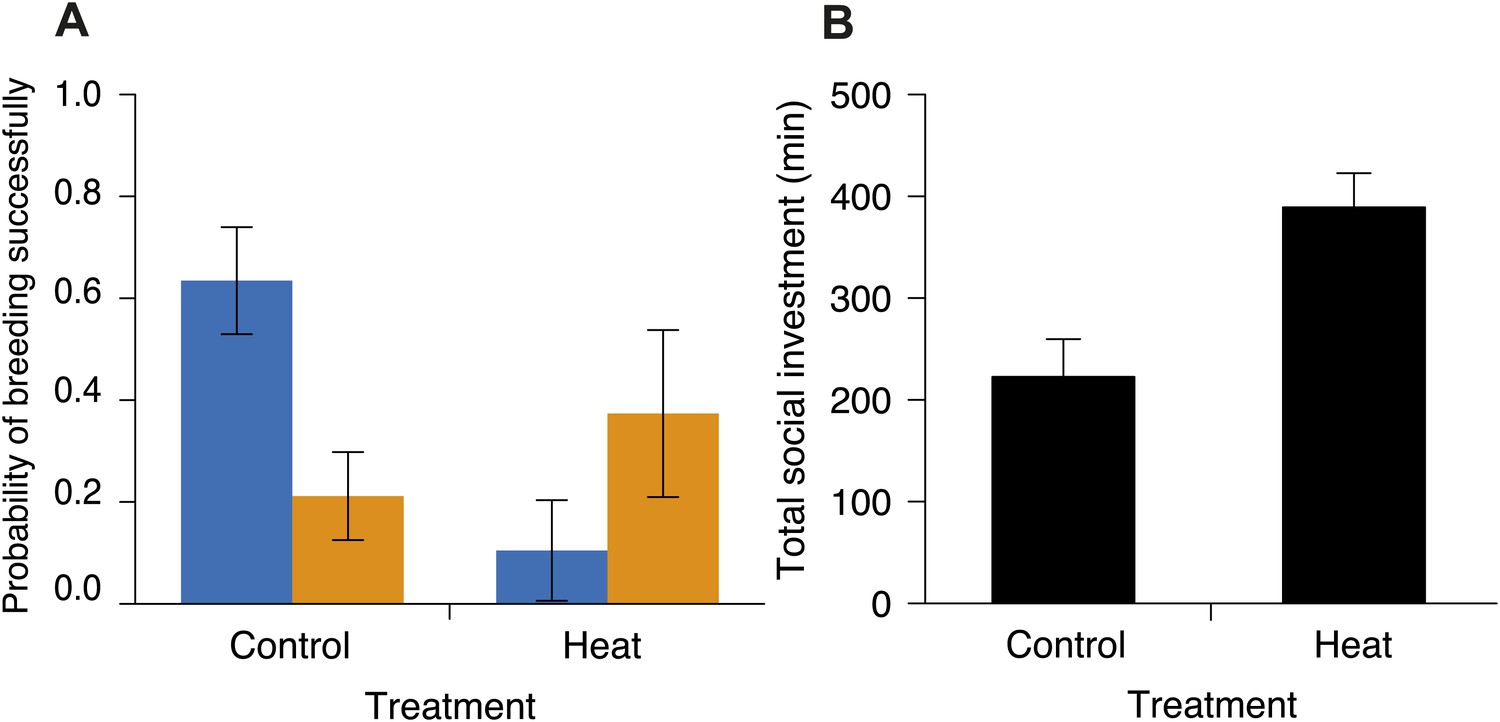
Investment in cooperative carcass processing in control and heated carcass treatments along the elevational gradient.
Heating carcasses differentially affected the breeding success of small and large groups when controlling for elevation (χ²1 = 6.55, p=0.010, n = 116). (A) Mean ± SE probability of breeding successfully (GLMM fitted values) for large (orange columns) and small (blue columns) groups of burying beetles in control and heated carcass treatments. Heating carcasses reduced the probability of breeding successfully in small groups (χ²1 = 5.99, p=0.014, n = 68), but not in large groups (χ²1 = 0.98, p=0.32, n = 48). (B) Mean ± SE total investment (minutes, min) in cooperative carcass processing was higher in heated carcass than control treatments (χ²1 = 12.67, p<0.001, n = 16).
Videos
Video 1
Social investment, Large group, August 15, 2011.
https://doi.org/10.7554/eLife.02440.016
Video 2
Social conflict, Large group, July 3 2013.
https://doi.org/10.7554/eLife.02440.017Tables
Table 1
Identification and abundance of carrion-feeding insects collected on rat carcasses from June to August 2011.
| Order | Percentage (%) | Family | Frequency |
|---|---|---|---|
| Coleoptera | 6.18 | Hydraenidae | 6 |
| Leiodidae | 11 | ||
| Ptiliidae | 6 | ||
| Silphidae | 9 | ||
| Diptera | 91.89 | Anthomyiidae | 7 |
| Calliphoridae | 117 | ||
| Carnidae | 2 | ||
| Drosophilidae | 33 | ||
| Fanniidae | 67 | ||
| Muscidae | 63 | ||
| Mycetophilidae | 1 | ||
| Phoridae | 103 | ||
| Psychodidae | 11 | ||
| Sarcophagidae | 7 | ||
| Sciaridae | 5 | ||
| Sphaeroceridae | 60 | ||
| Hymenoptera | 1.74 | Formicidae | 8 |
| Vespidae | 1 | ||
| Lepidoptera | 0.19 | Tortricidae | 1 |
| Total | 100 | 19 families | 518 |
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Climate-mediated cooperation promotes niche expansion in burying beetles
eLife 3:e02440.
https://doi.org/10.7554/eLife.02440




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}