Benefits of jasmonate-dependent defenses against vertebrate herbivores in nature
- Max Planck Institute for Chemical Ecology, Germany
- University of Bern, Switzerland
- Max-Planck Institute for Chemical Ecology, Germany
- Washington State University, United States
- Institut de Génétique, Environment et Protection des Plantes, France
Figures
Figure 1 with 4 supplements
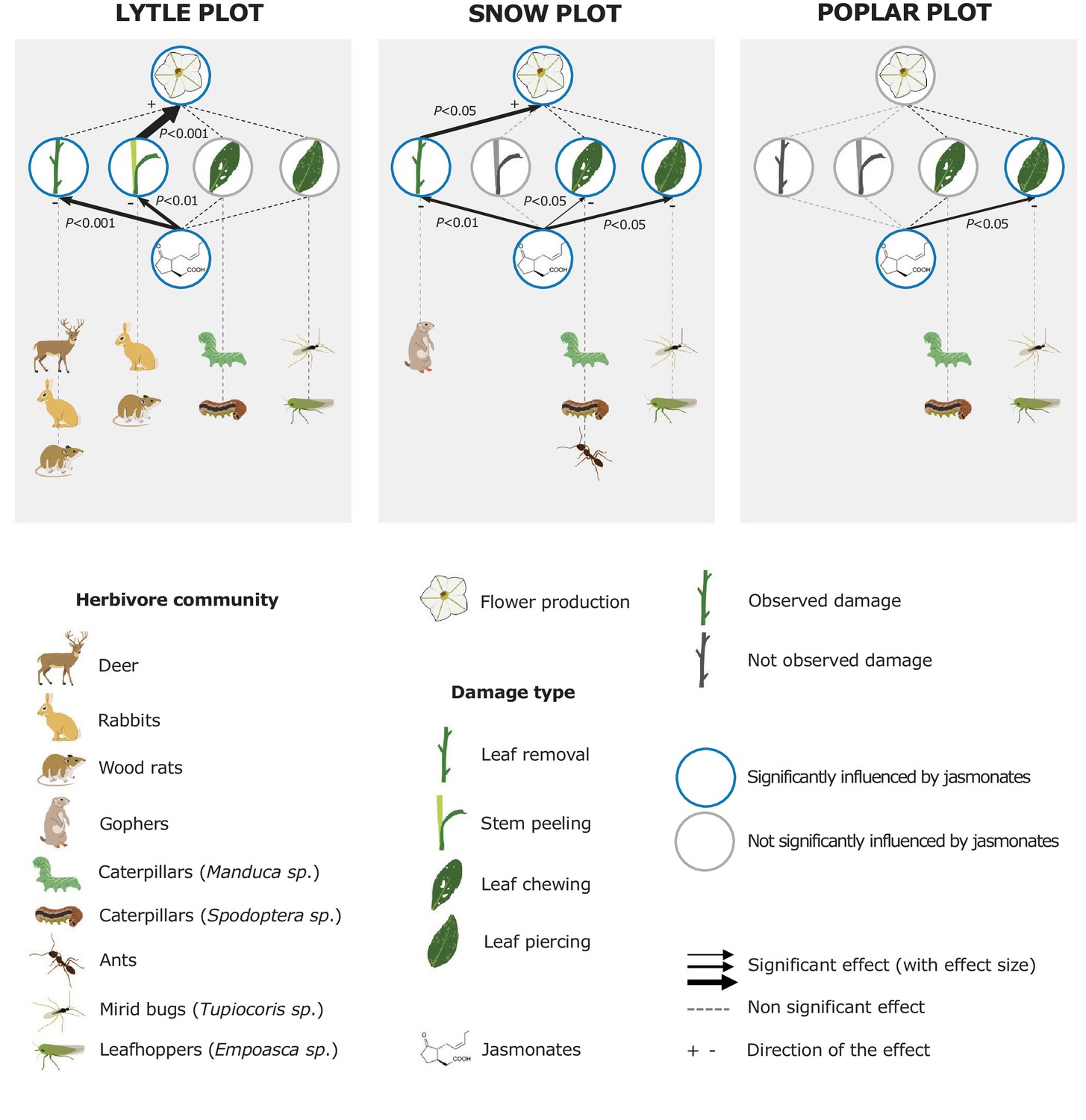
Jasmonate-deficiency reduces flower production by increasing vertebrate damage in nature.
Effects of jasmonate deficiency on vertebrate and invertebrate damage by damage type and effect of damage type on flower production across three experimental plots ('Lytle', 'Poplar' and 'Snow') are shown (n = 12–19). Solid lines indicate significant effects of jasmonate-deficiency on herbivore damaage patterns and flower production. The herbivores responsible for the different damage types were identified based on field observations and characteristic feeding patterns. Jasmonate-deficiency increases damage by vertebrate and arthropod herbivores, but only vertebrate damage leads to a reduction of flower production as a strong predictor of plant reproductive potential.
-
Figure 1—source data 1
Parameters used to determine the presence of different herbivores in the three experimental populations.
- https://doi.org/10.7554/eLife.13720.004
-
Figure 1—source data 2
Quantification and herbivore association of the different damage types observed in the three experimental populations.
- https://doi.org/10.7554/eLife.13720.005
-
Figure 1—source data 3
Leaf-herbivore damage screen and fitness measurements.
- https://doi.org/10.7554/eLife.13720.006
Figure 1—figure supplement 1
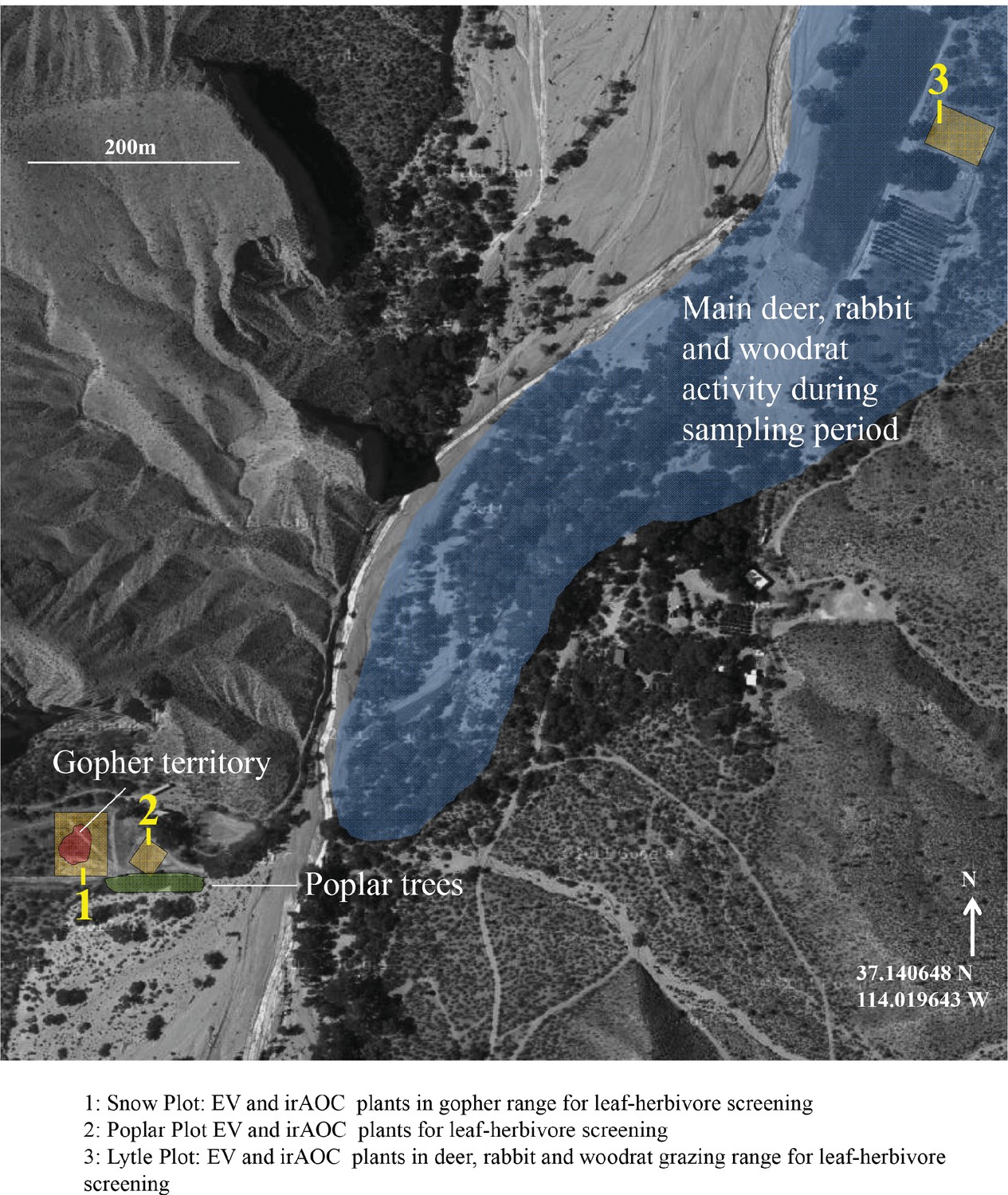
Overview of the different experimental N. attenuata populations used in the present study.
Satellite picture from googlemaps.com.
Figure 1—figure supplement 2
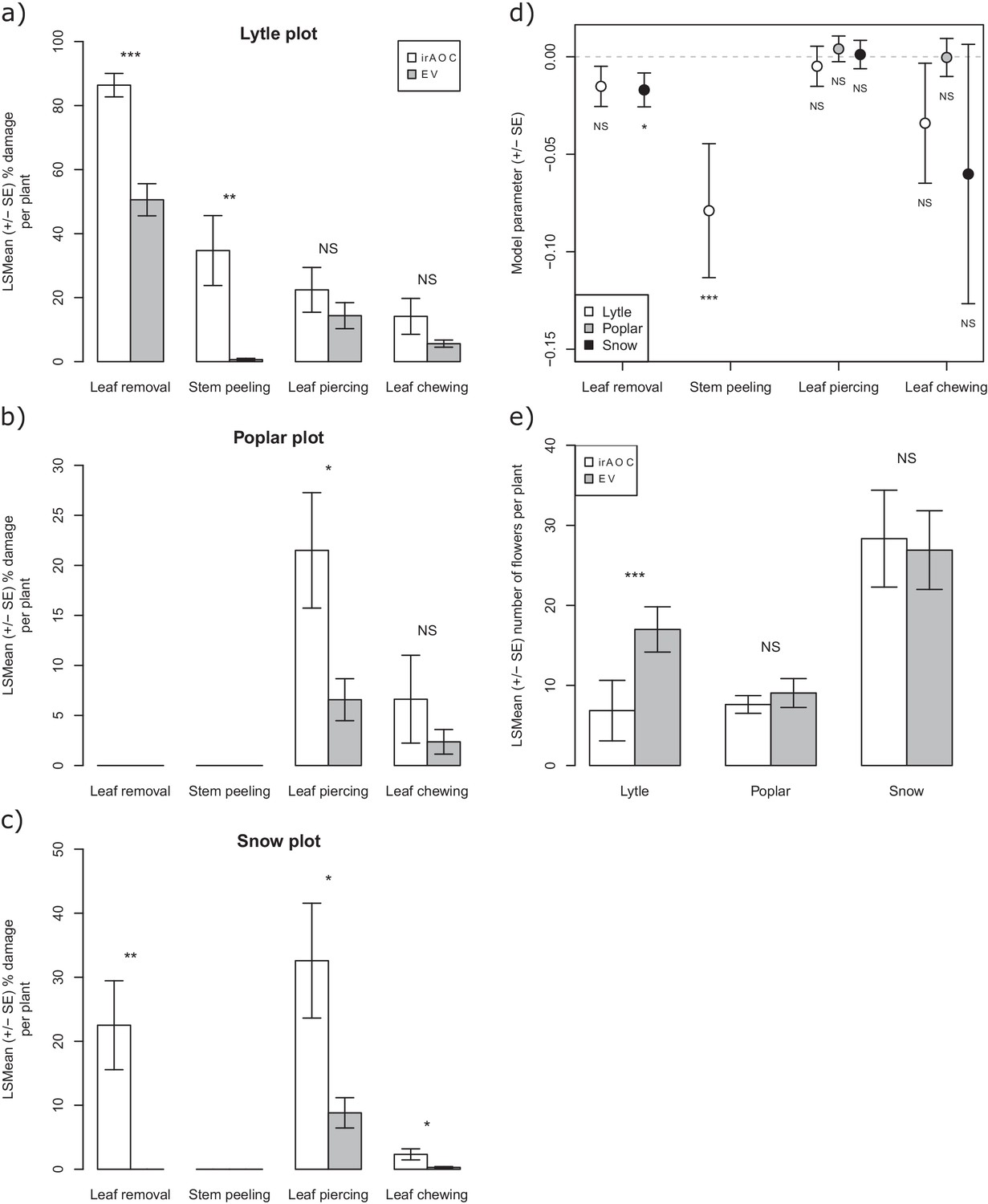
Detailed results of the analysis of the overall effect of jasmonate signaling and herbivore damage on N. attenuata flower production in the field.
Mean herbivore damage in the Lyttle (a), Poplar (b) and Snow (c) plot. Effect of type of damage on plant fitness (d). Mean flower production in the different plots (e). Asterisks indicate significant differences (*p<0.05; **p<0.01; ***p<0.001). N. S: not significant.
Figure 1—figure supplement 3

Photographic evidence of herbivore damage and the associated herbivores in the Lytle plot.
Mule deer and cottontail rabbits were regularly encountered in the vicinity of the Lytle plot (Top, white arrows). Bottom left: Typical EV plant in the Lytle plot, with partial defoliation. Bottom right: IrAOC plant that was completely defoliated and stem-peeled by vertebrate browsing. Stem peeling is typically caused by mountain cottontail rabbits (Sylvilagus nuttallii), black-tailed jackrabbits (Lepus californicus) and woodrats (Neotoma spp.). Cottontail rabbit picture by Pavan Kumar.
Figure 1—figure supplement 4

Photographic evidence of herbivore damage and the associated herbivores in the Snow plot.
Several gophers were observed within the snow plot (Top, white arrow). Bottom left: Typical EV plant in the snow plot. Bottom right: IrAOC plant that was partially defoliated by animal browsing. Gopher picture by Arne Weinhold.
Figure 2 with 1 supplement
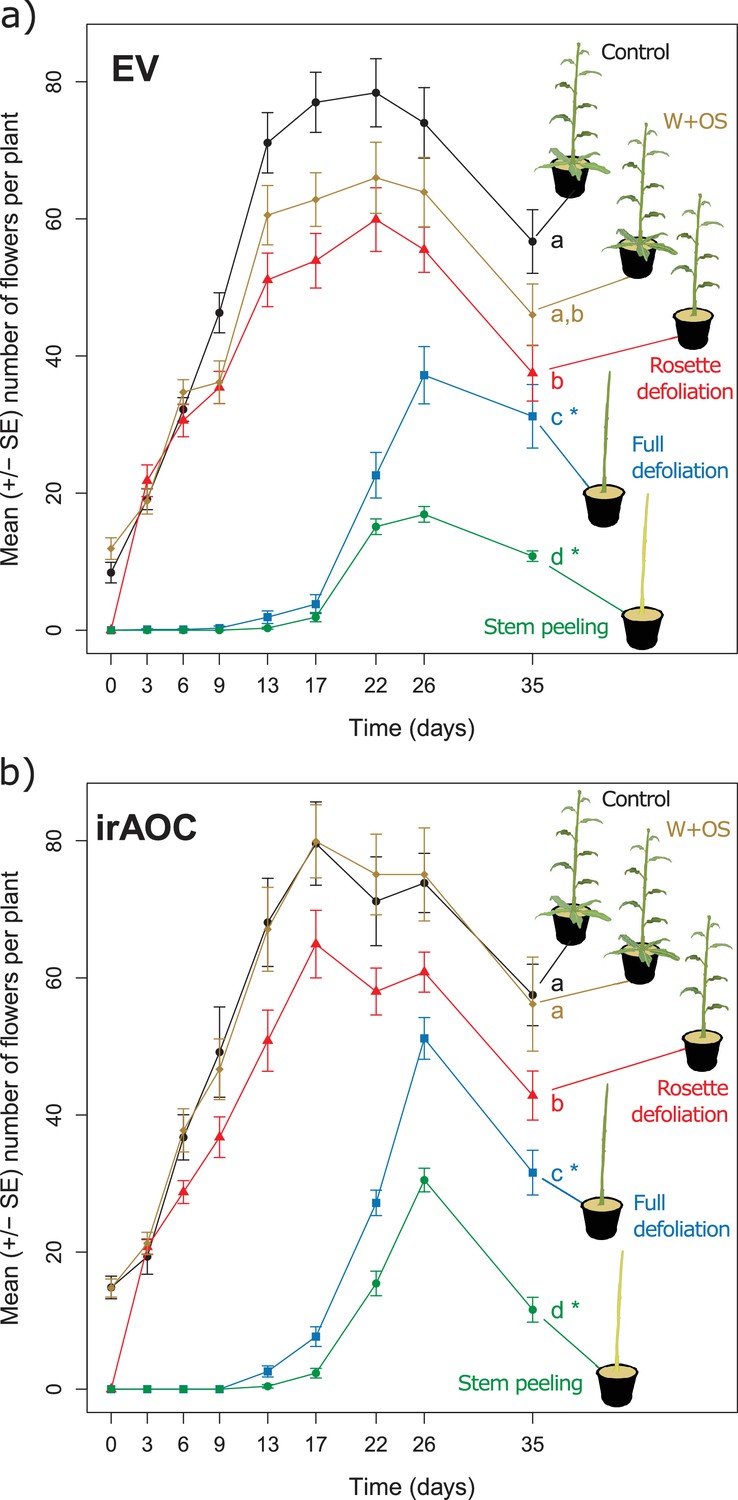
Simulated vertebrate attack strongly reduces plant flower production.
Average (±SE) flower production of wild type (a) and jasmonate deficient irAOC plants (b) following different simulated herbivory treatments (n =10–12). Different letters indicate significant differences between treatments within genotypes (p<0.05). Asterisks indicate significant differences between genotypes within treatments (*p<0.05). W+OS: Wounding + application of Manduca sexta oral secretions.
-
Figure 2—source data 1
Fitness costs of induction and defoliation in the glasshouse.
- https://doi.org/10.7554/eLife.13720.012
Figure 2—figure supplement 1
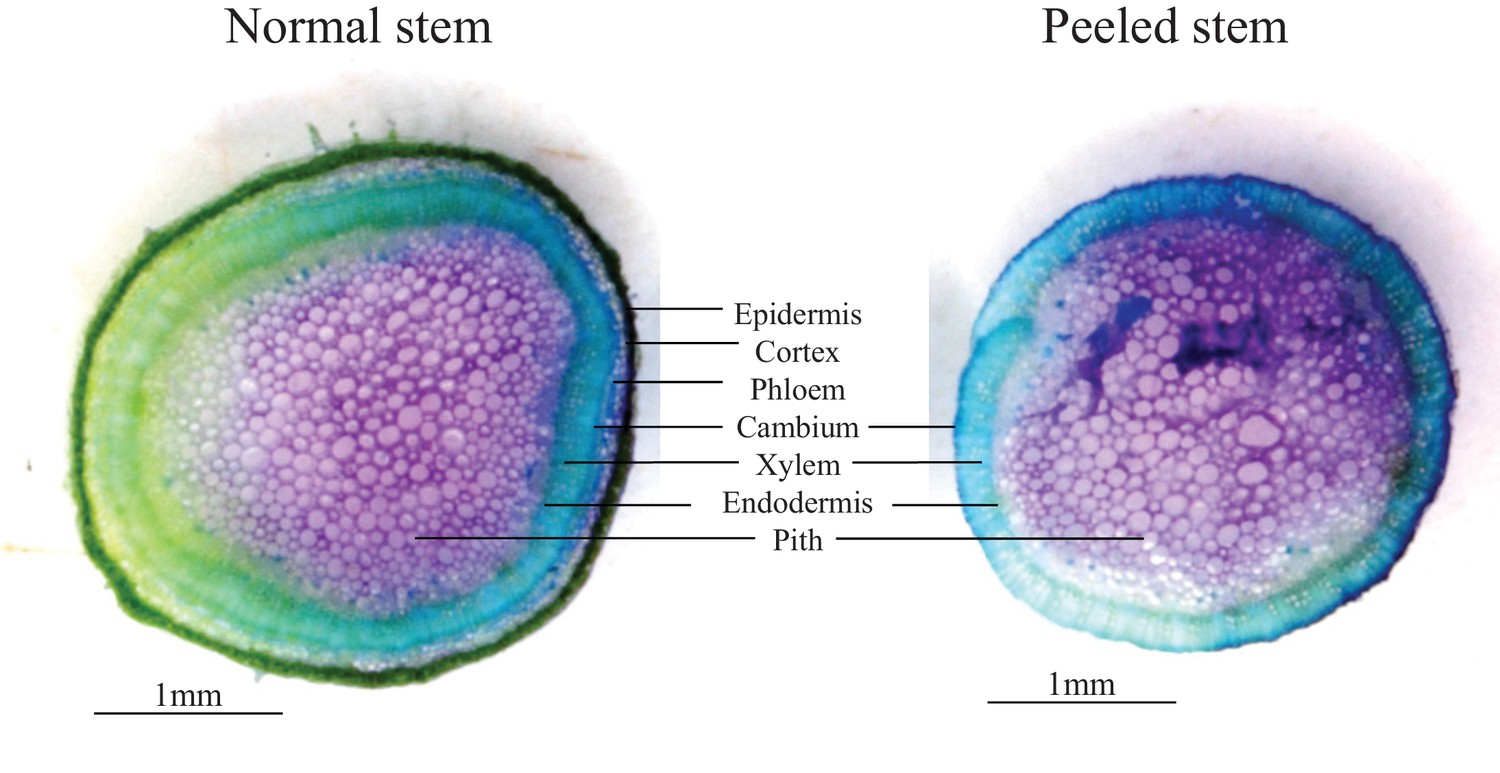
Toluene-blue staining of stem-peeled EV plants at the early bolting stage.
Experimental stem peeling as a mimick of feeding patterns observed in the Lytle plot removed epidermis, cortex and phloem cells from the plant. The remaining tissue consisted of xylem and pith cells.
Figure 3 with 1 supplement

Herbivory-induced jasmonates increase secondary metabolites and decrease sugars in leaves and stems.
Results of a redundancy analysis of the metabolic profiles in the leaves (a) and in the stems (b) of wild type (EV) and jasmonate-deficient (irAOC) plants are shown (n = 3–5). Insets depict average (±SE) concentrations of metabolites explaining most of the variation between induced EV plants and the other treatment*genotype combinations (correlation coefficients > |0.8|). Letters indicate significant differences between treatments within genotypes (p<0.05). Asterisks indicate significant differences between genotypes within treatments (p<0.05). Control: intact plants; W+W: wounded and water-treated plants; W+OS: wounded and M. sexta oral secretions-treated plants; DTGs: diterpene glycosides.
-
Figure 3—source data 1
Plant primary and secondary metabolite concentrations.
- https://doi.org/10.7554/eLife.13720.015
Figure 3—figure supplement 1
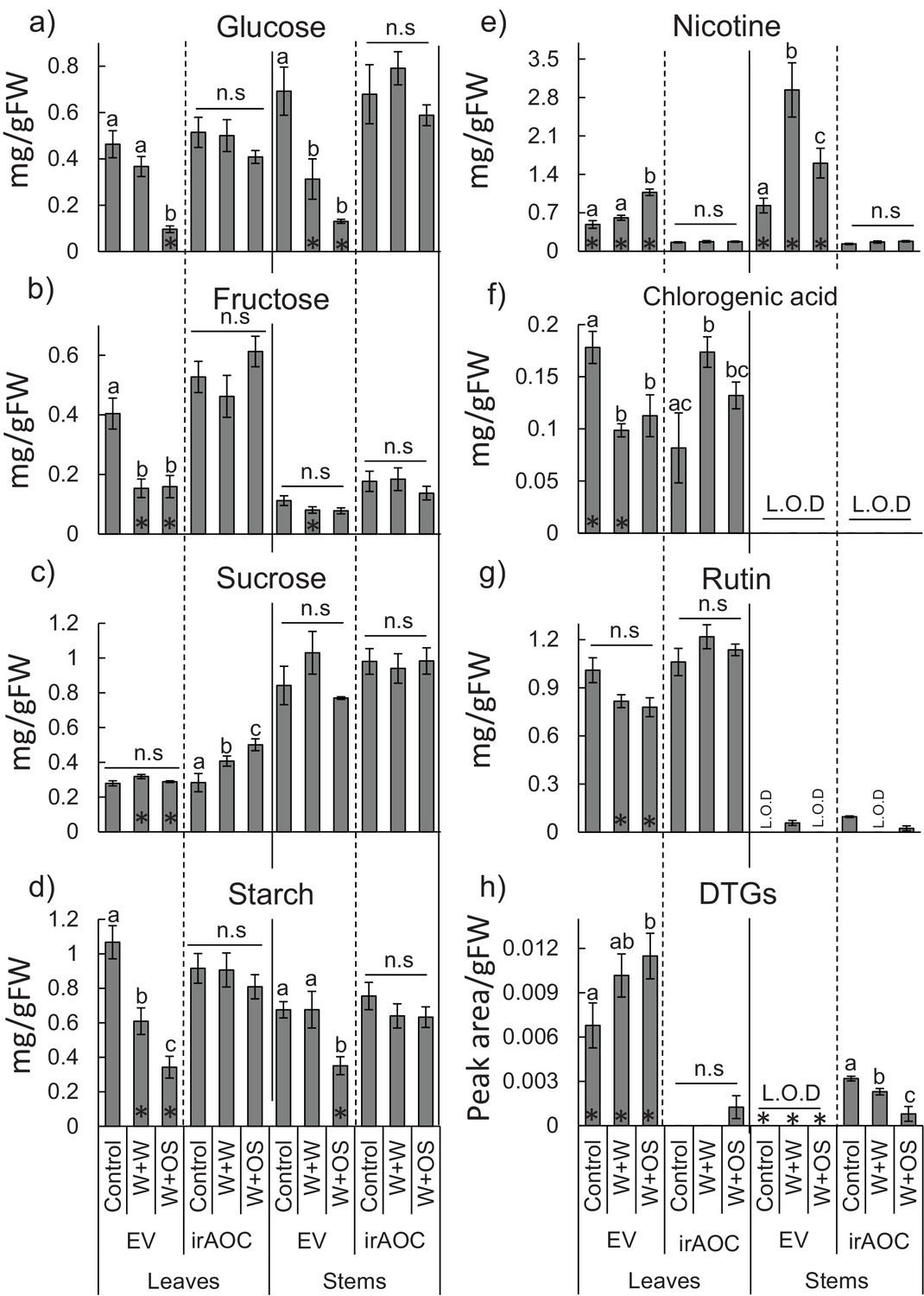
Secondary metabolite profiles in leaves and stems of N. attenuata in response to simulated herbivore attack.
Mean (±SE) glucose (a), fructose (b), sucrose (c), starch (d) nicotine (e), chlorogenic acid (f), rutin (g) and diterpene glycosides (DTGs) (h) (n=5). Different letters indicate significant differences between treatments within genotype and tissue (p<0.05). n.s: not significant. Asterisks indicate significant differences between genotypes within treatment (*p<0.05). L.O.D: limit of detection. Control: intact plants; W+W: wounded and water treated plants; W+OS: wounded and M. sexta-oral secretions treated plants.
Figure 4 with 4 supplements
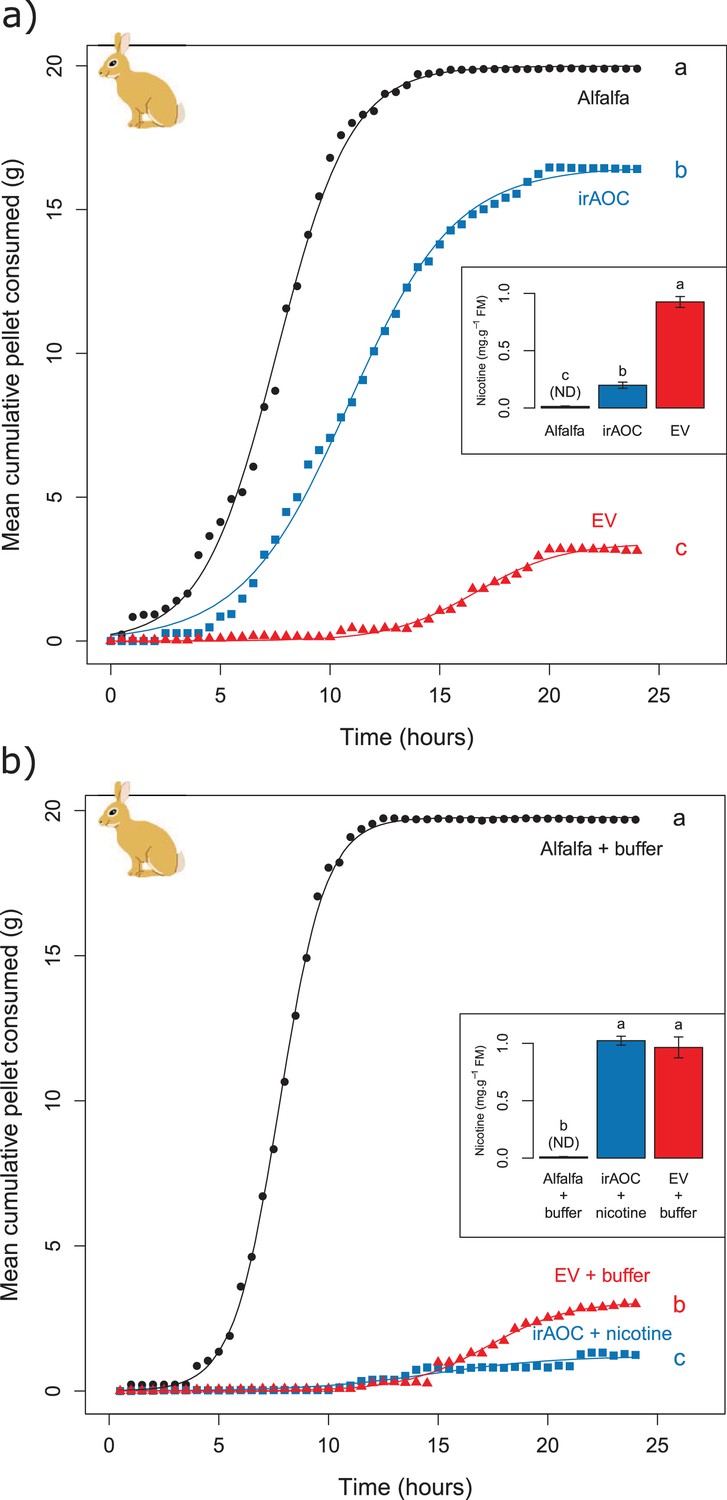
Jasmonate-dependent nicotine determines feeding preferences of a native vertebrate herbivore.
Average amounts of food consumption by cottontail rabbits in a choice experiment. Choice experiments were carried out with EV, irAOC and alfalfa pellets (a) (n = 8) or with EV, irAOC+nicotine and alfalfa pellets (b) (n = 9). Letters indicate significant differences in curve asymptotes tested based on asymptote confidence intervals of logistic models (p<0.001). Insets: nicotine contents of the different pellets. Letters indicate significant differences (p<0.05). N.D: not detected.
-
Figure 4—source data 1
Impact of drying on secondary metabolite profiles.
- https://doi.org/10.7554/eLife.13720.018
-
Figure 4—source data 2
Nicotine concentrations in pellets.
- https://doi.org/10.7554/eLife.13720.019
-
Figure 4—source data 3
Results of choice experiment (repetition).
- https://doi.org/10.7554/eLife.13720.020
-
Figure 4—source data 4
Results of nicotine complementation experiment (repetition).
- https://doi.org/10.7554/eLife.13720.021
Figure 4—figure supplement 1

Experimental setup for feeding preference assays for cottontail rabbits (Sylvilagus nuttallii).
https://doi.org/10.7554/eLife.13720.022
Figure 4—figure supplement 2
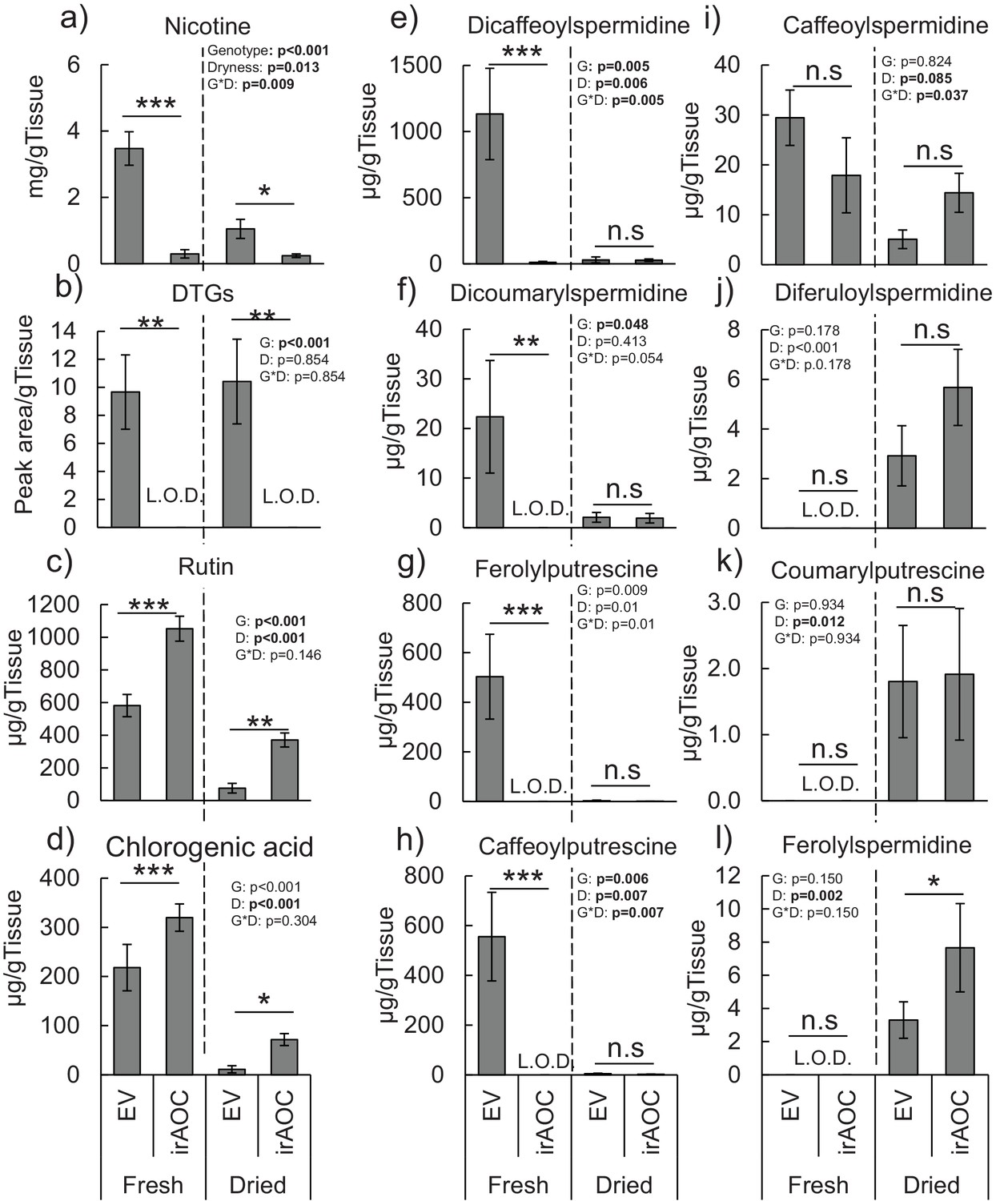
Secondary metabolite profiles in the leaves of N. attenuata before and after drying.
Mean concentrations (±SE) of nicotine (a), diterpeneglycosides (DTGs) (b), rutin (c), chlorogenic acid (d) dicaffeoylspermidine (e), dicoumarylspermidine (f), ferolylputrescine (g), caffeoylputrescine (h), caffeoylspermidine (i), diferuloylspermidine (j), coumarylputrescine (k) and ferolylspermidine (l) (n = 5). Asterisks indicate significant differences between genotypes within water content status (*p<0.05; **p<0.01; ***p<0.001). N. S: not significant. L.O.D: limit of detection.
Figure 4—figure supplement 3
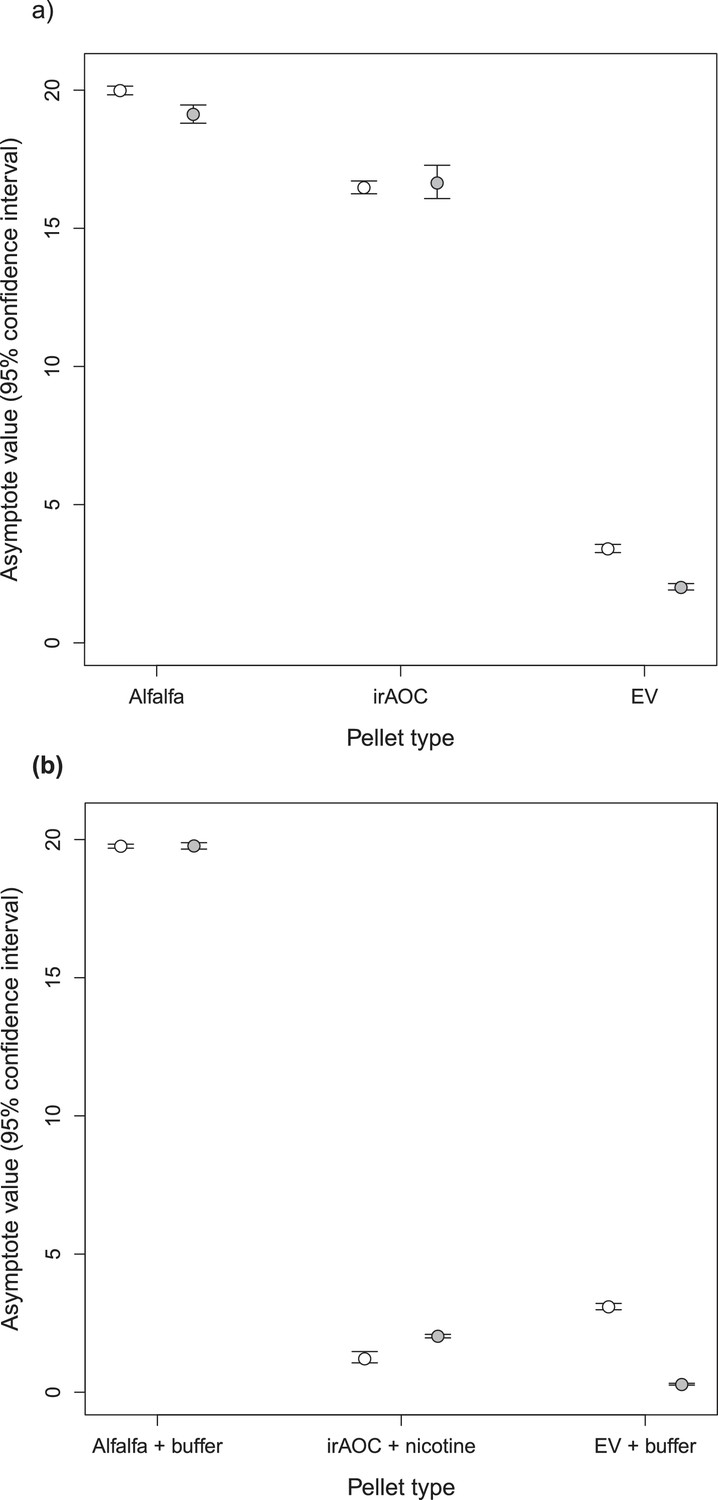
Asymptotes and confidence intervals of the feeding preference assays for cottontail rabbits (Sylvilagus nuttallii).
https://doi.org/10.7554/eLife.13720.024
Figure 4—figure supplement 4
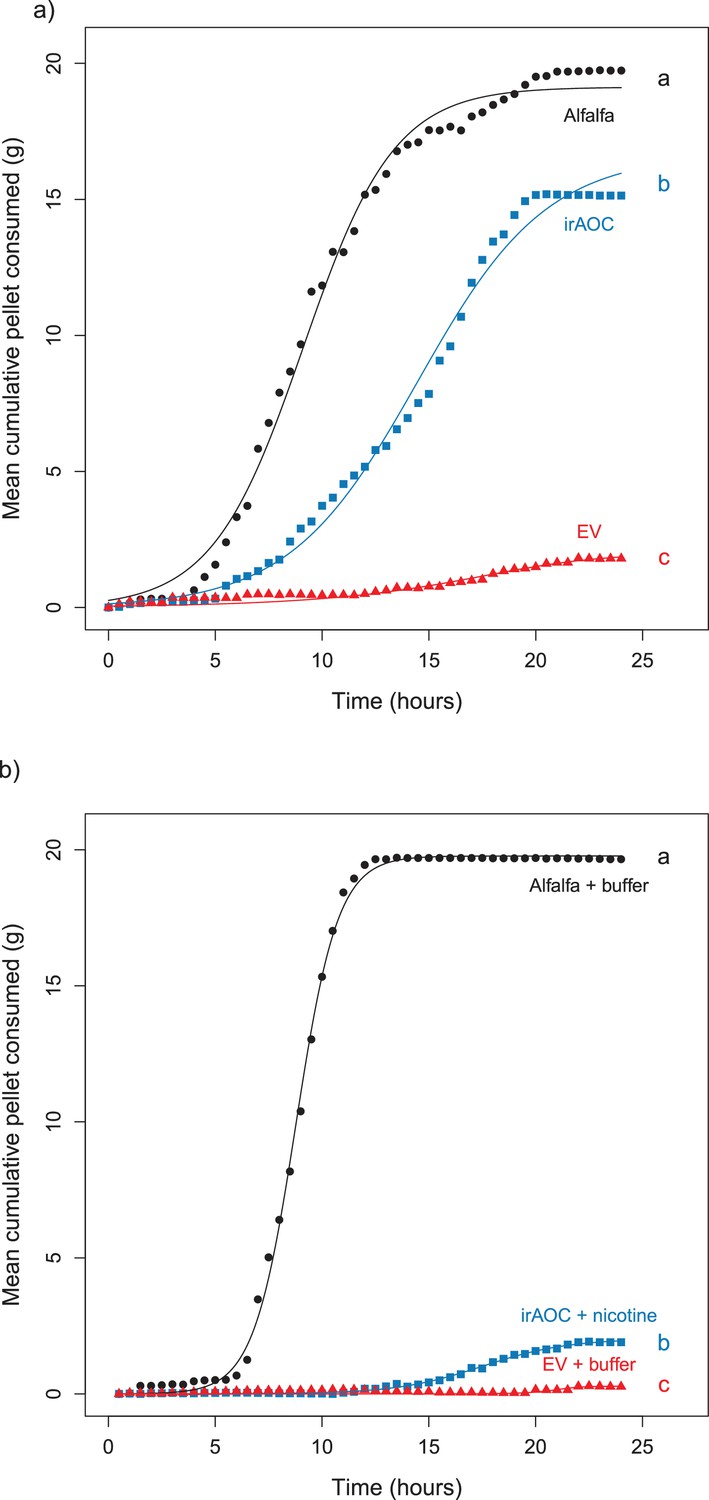
Results of the repetition of the feeding preference assays for cottontail rabbits (Sylvilagus nuttallii).
Choice experiments were carried out with EV, irAOC and alfalfa pellets (a) (n = 8) or with EV, irAOC+nicotine and alfalfa pellets (b) (n = 9). Letters indicate significant differences in curve asymptotes tested based on asymptote confidence intervals of logistic models (p<0.001). Insets: nicotine contents of the different pellets. Letters indicate significant differences (p<0.05). N.D: not detected.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Benefits of jasmonate-dependent defenses against vertebrate herbivores in nature
eLife 5:e13720.
https://doi.org/10.7554/eLife.13720




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}