A beta-glucosidase of an insect herbivore determines both toxicity and deterrence of a dandelion defense metabolite
- Institute of Plant Biology and Biotechnology, University of Muenster, Germany
- Department of Biochemistry, Max-Planck Institute for Chemical Ecology, Germany
- Institute of Plant Sciences, University of Bern, Switzerland
- Department of Bioorganic Chemistry, Max-Planck Institute for Chemical Ecology, Germany
- Research group Biosynthesis/NMR, Max-Planck Institute for Chemical Ecology, Germany
Figures
Figure 1 with 7 supplements
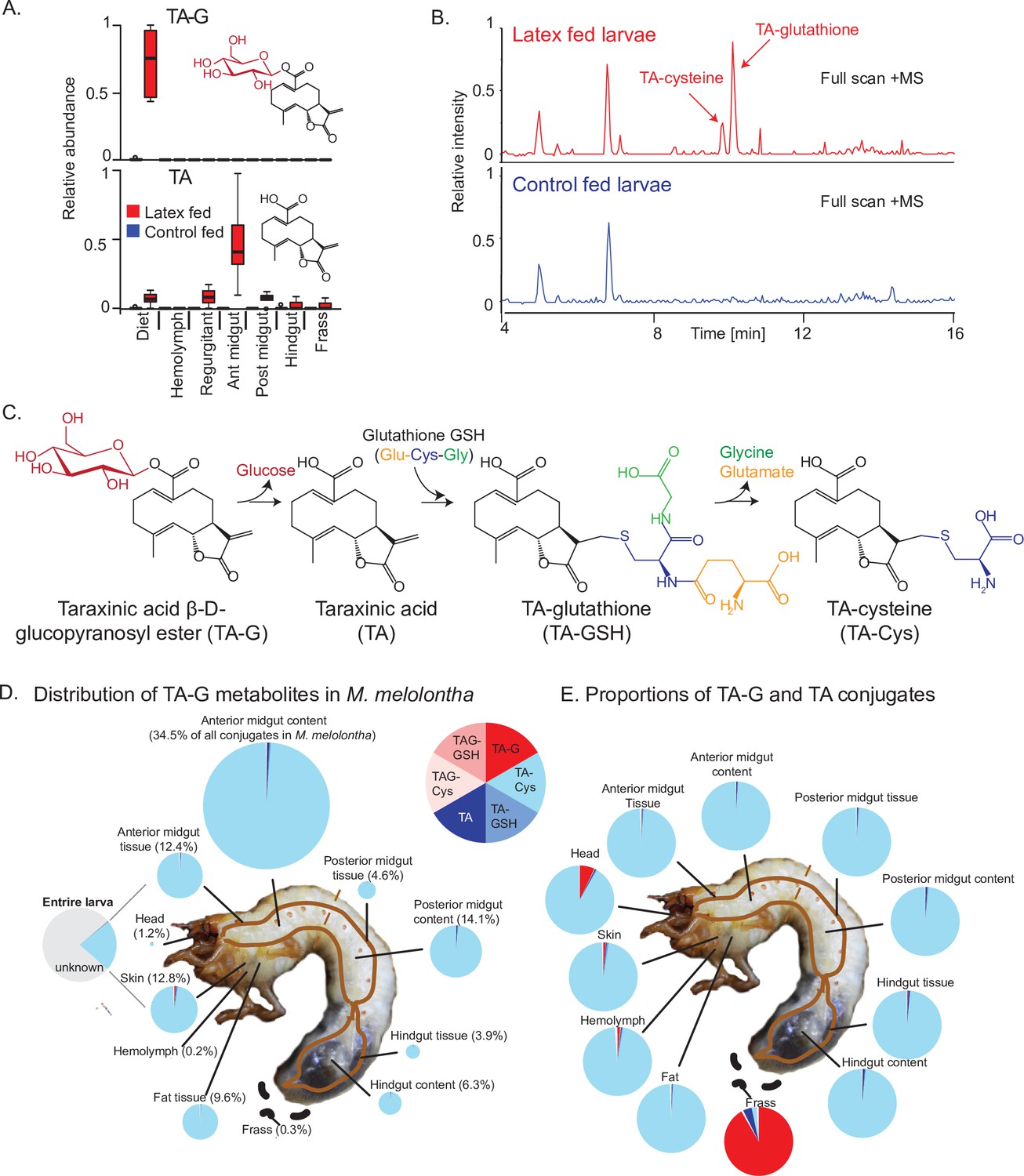
Taraxinic acid β-D-glucopyranosyl ester is rapidly deglucosylated and conjugated to glutathione upon ingestion by Melolontha melolontha.
(A) Relative abundance of taraxinic acid β-D-glucopyranosyl ester (TA-G) and its aglycone taraxinic acid (TA) in diets enriched with Taraxacum officinale latex and in Melolontha melolontha larval gut, hemolymph, and frass after feeding on latex-containing and control diets. Ant = anterior; post = posterior. N = 5. For relative quantification of TA-glutathione (TA-GSH) conjugates in M. melolontha, refer to Figure 1—figure supplement 1. (B) High-pressure liquid chromatography-mass spectrometry (HPLC-MS) full scan (positive mode) of the anterior midgut of M. melolontha larvae fed with latex-containing and control diets. (C) Schematic illustration of proposed TA-G metabolism in M. melolontha. For nuclear magnetic resonance (NMR) analysis of TA-GSH conjugates, refer to Figure 1—figure supplements 2–6. Distribution of the total deglucosylated and conjugated metabolites of TA-G in M. melolontha larvae that consumed 100 µg TA-G within 24 hr. The size of the circles is relative to the total amount of conjugates. Values denote the percentage of metabolites found in each body part and are the mean of eight replicates. For long-term distribution of TA-Cys in M. melolontha, refer to Figure 1—figure supplement 7. (E) Relative proportions of TA-G metabolites in quantities from panel (D). Values denote the mean of eight replicates. Raw data are available in Figure 1—source data 1.
-
Figure 1—source data 1
Source data of main and supplementary figures of Figure 1.
- https://cdn.elifesciences.org/articles/68642/elife-68642-fig1-data1-v1.xlsx
Figure 1—figure supplement 1

Relative quantification of TA-glutathione conjugates in Melolontha melolontha larvae feeding on diets with and without Taraxacum officinale latex.
N = 5. TA = taraxinic acid; GSH = glutathione; Cys = cysteine; Gly = glycine; Glu = glutamate.
Figure 1—figure supplement 2
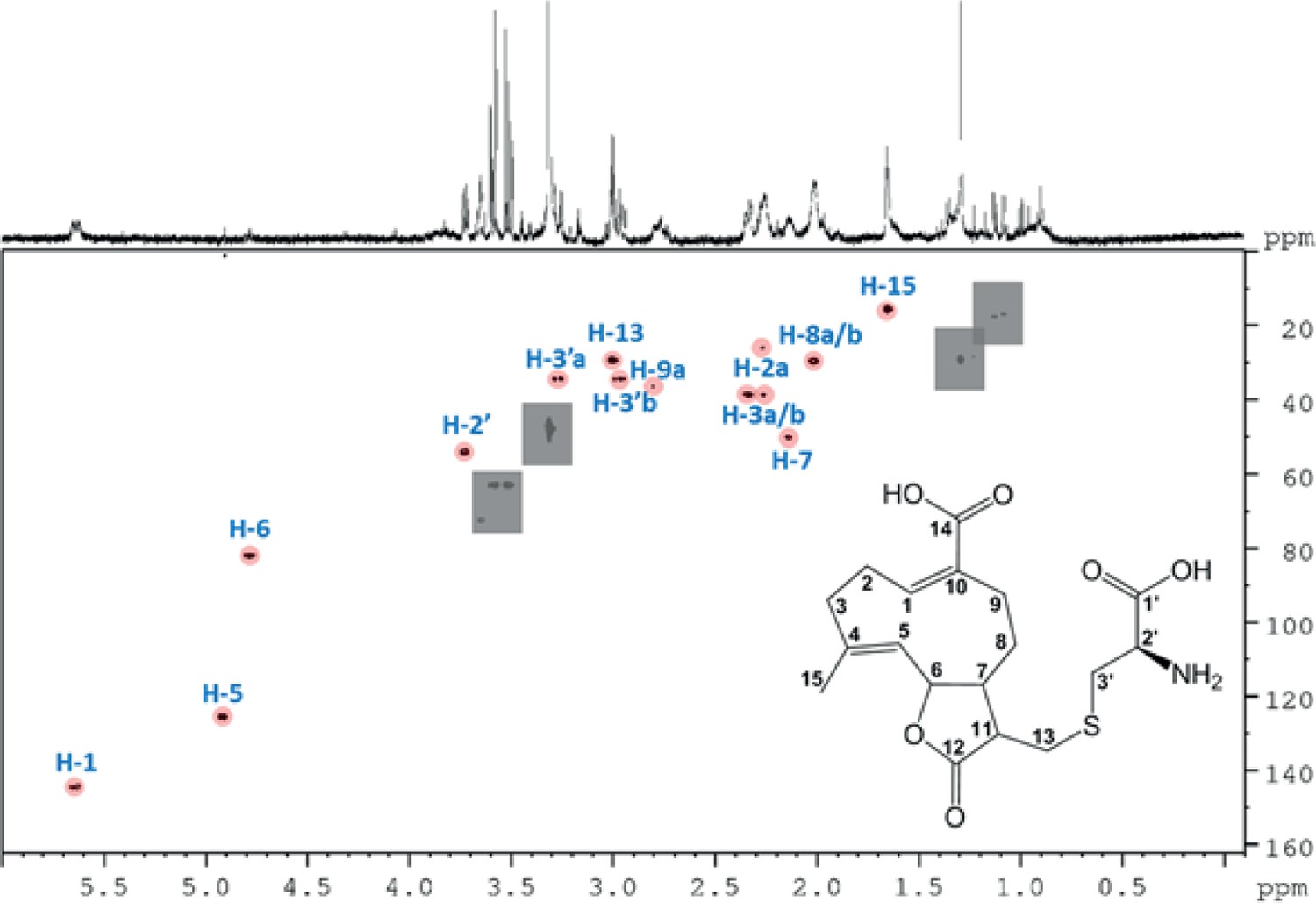
500 MHz 1H-13C HSQC NMR spectrum of the partially purified Melolontha melolontha midgut extract.
Intensity level was adjusted to suppress noise. Gray rectangles mask impurities for clarity. Resonances above noise level are highlighted by pink circles. Missing resonances in the noise level are not considered in the picture. Sample was measured in MeOH-d4.
Figure 1—figure supplement 3

Structure of synthesized TA-G-GSH with chemical shifts (500 MHz, in MeOD-d4).
Figure 1—figure supplement 4
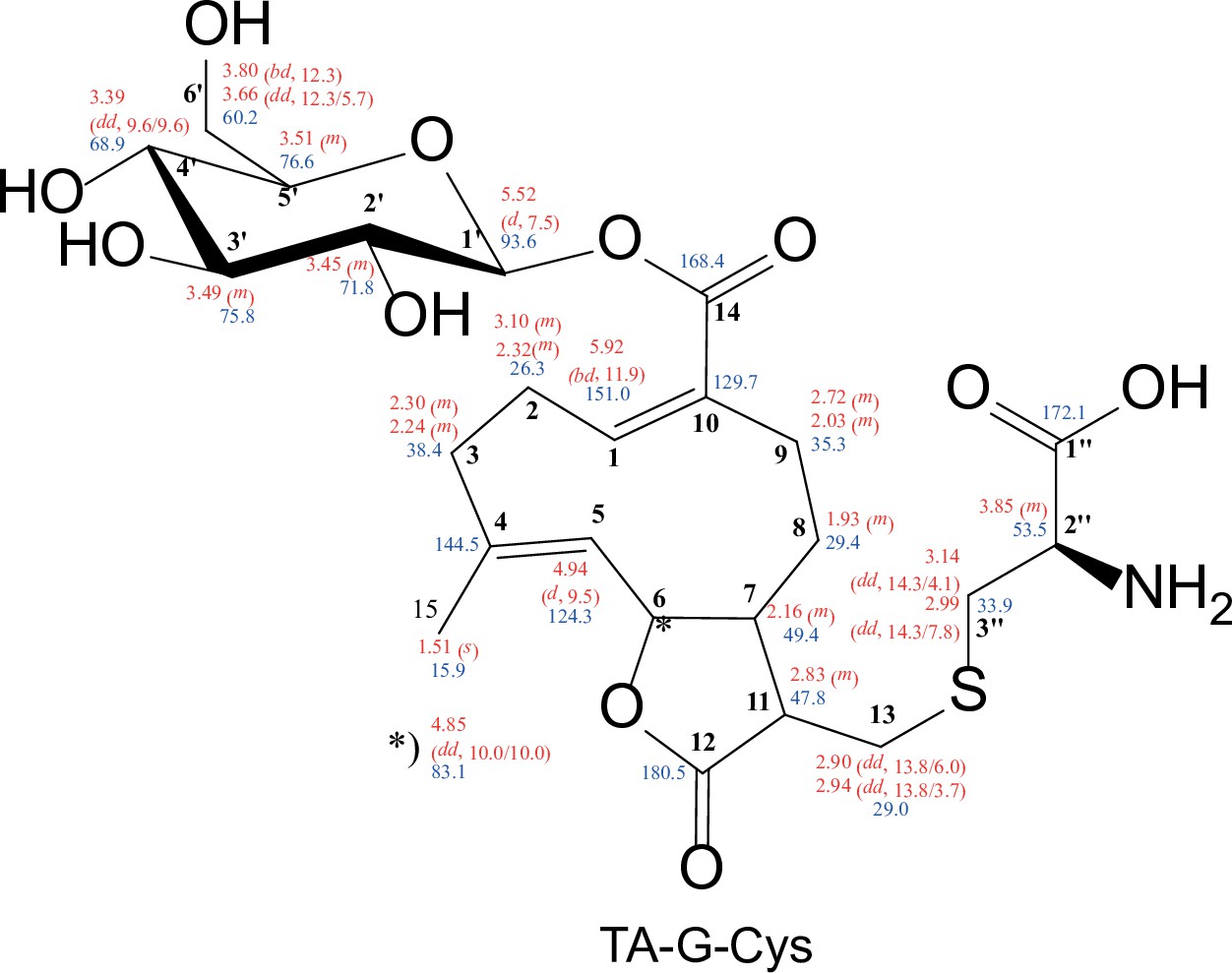
Structure of synthesized TA-G-Cys with chemical shifts (500 MHz, in MeOD-d4).
Figure 1—figure supplement 5
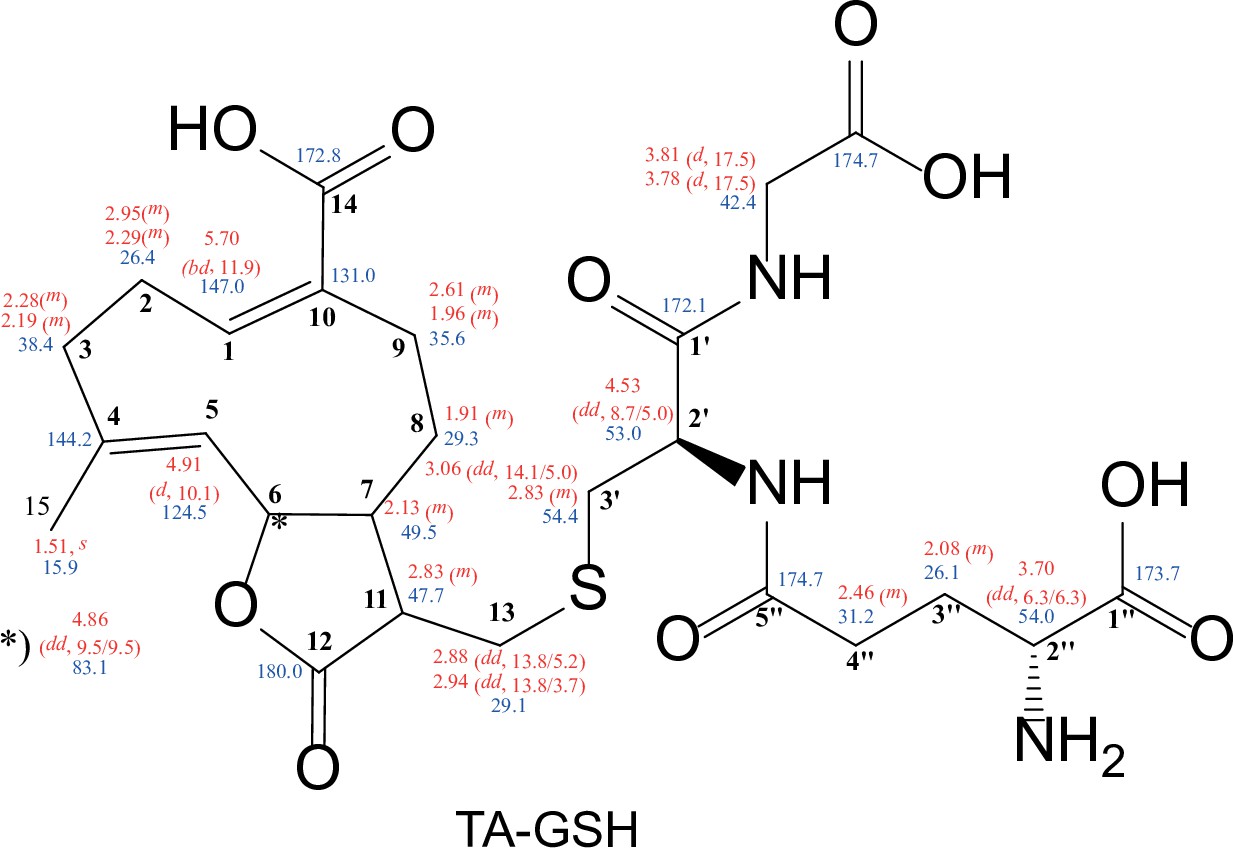
Structure of synthesized TA-GSH with chemical shifts (500 MHz, in MeOD-d4).
Figure 1—figure supplement 6
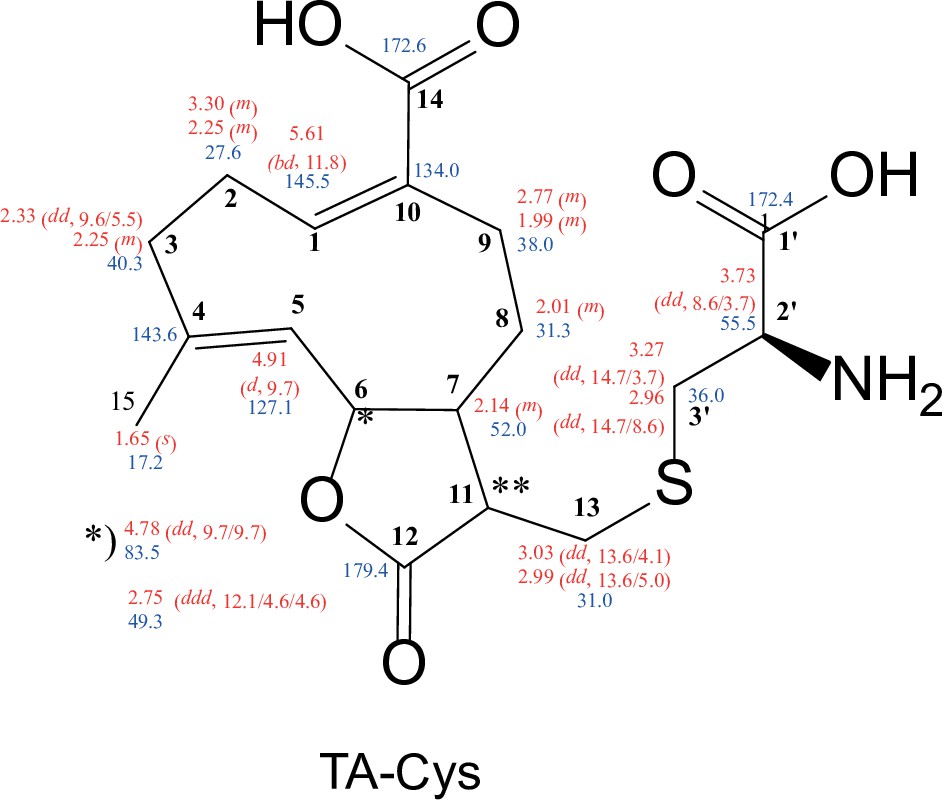
Structure of synthesized TA-Cys with chemical shifts (500 MHz, in MeOD-d4).
Figure 1—figure supplement 7
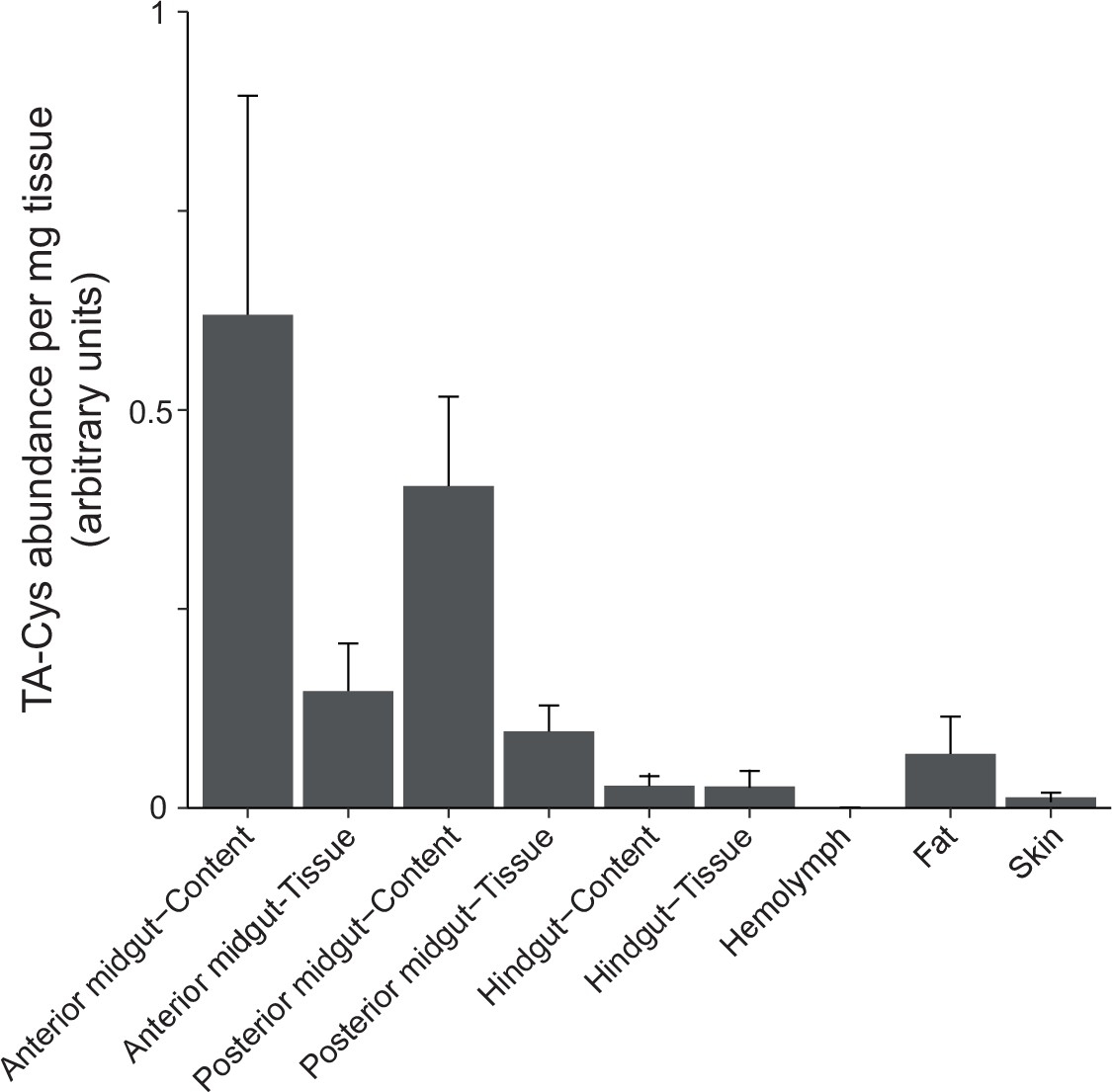
Accumulation of TA-Cys in different body parts of Melolontha melolontha upon feeding for 1 month on Taraxacum officinale plants.
The mean of four to six replicates is displayed.
Figure 2 with 2 supplements
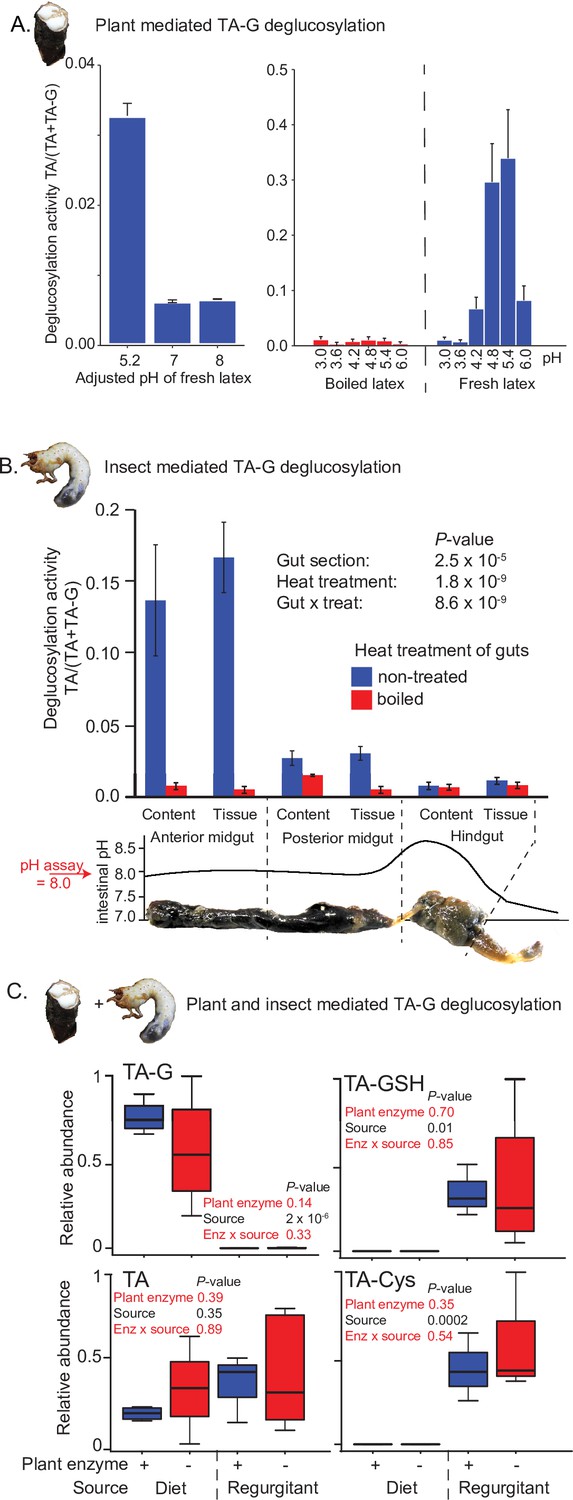
Insect rather than plant enzymes deglucosylate TA-G.
(A) Left and right panels: plant-mediated enzymatic deglucosylation of TA-G at pH 3–8. Taraxacum officinale latex was collected from wounded roots and incubated in buffers adjusted to different pH values. N = 3. (B) Deglucosylation activity of untreated and boiled extracts of Melolontha melolontha gut content and gut tissue incubated at pH 8.0 with boiled latex extracts. The p-values of a two-way analysis of variance (ANOVA) are shown. N = 6. Error bars = SEM. The intestinal pH of M. melolontha is shown for comparative purposes (data from Egert et al., 2005). For in vitro M. melolontha glucosidase inhibition assays, refer to Figure 2—figure supplement 1 - 2. (C) Relative abundance of TA-G and its metabolites in the diet and regurgitant of larvae fed with carrot slices coated with either intact (+) or heat-deactivated (-) T. officinale latex. Heat deactivation of latex did not significantly affect the deglucosylation of TA-G in M. melolontha. p-values refer to two-way ANOVAs. N = 4. TA-G: taraxinic acid β-D-glucopyranosyl ester; TA = taraxinic acid; GSH = glutathione; Cys = cysteine. Peak area was normalized across all treatments based on the maximal value of each metabolite. Raw data are available in Figure 2—source data 1.
-
Figure 2—source data 1
Source data of main and supplementary figures of Figure 2.
- https://cdn.elifesciences.org/articles/68642/elife-68642-fig2-data1-v1.xlsx
Figure 2—figure supplement 1

TA-G deglucosylation activity (TA/(TA + TA-G)) of Melolontha melolontha anterior midgut samples in the presence of either the carboxylesterase inhibitor bis(p-nitrophenyl)phosphate or the α- and β-glucosidase inhibitor castanospermin.
Only the glucosidase inhibitor reduced deglucosylation of taraxinic acid β-D-glucopyranosyl ester (TA-G). N = 6. Error bars denote SEM.
Figure 2—figure supplement 2
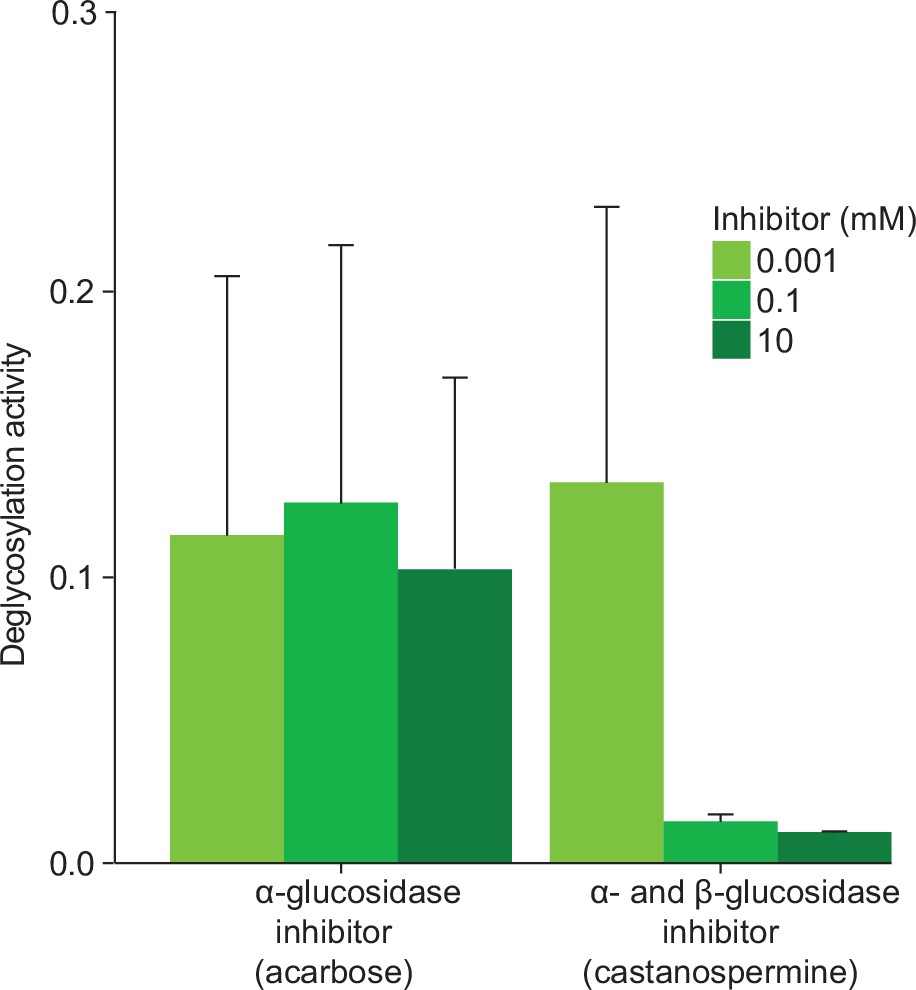
TA-G deglucosylation activity (TA/(TA + TA G)) of Melolontha melolontha anterior midgut samples in the presence of acarbose, an α-glucosidase-specific inhibitor, or castanospermine, an α- and β-glucosidase inhibitor.
Only castanospermine reduced deglucosylation of TA-G. TA-G = taraxinic acid β-D-glucopyranosyl ester; TA = taraxinic acid. N = 3. Standard errors denote SEM.
Figure 3 with 4 supplements

Melolontha melolontha midgut β-glucosidases hydrolyze TA-G and other plant defensive glycosides.
(A) Phylogeny of newly identified Melolontha melolontha β-glucosidases and previously reported β-glucosidases of Tenebrio molitor (Tm bGlc, AF312017.1) and Chrysomela populi (Cp bGlc, KP068701.1), and myrosinases (thioglucosidases) of Phyllotreta striolata (Ps myrosinase, KF377833.1) and Brevicoryne brassicae (Bb myrosinase, AF203780.1) based on amino acid similarities using maximum likelihood method. Bootstrap values (N = 1000) are shown next to each node. Amino acid sequence alignments of the β-glucosidases are shown in Figure 3—figure supplement 1. (B) Heat map of average (n = 3) gene expression levels of M. melolontha β-glucosidases in the anterior and posterior midgut of larvae feeding on diets supplemented with water, taraxinic acid β-D-glucopyranosyl ester (TA-G), or Taraxacum officinale latex-containing diet. FPKM = fragments per kilobase of transcript per million mapped reads. (C) Activity of heterologously expressed M. melolontha β-glucosidases with TA-G, a mixture of maize benzoxazinoids, the salicinoid salicin, 4-methylsulfinylbutyl glucosinolate (4-MSOB), cellobiose, and the fluorogenic substrate 4-methylumbelliferyl-β-D-glucopyranoside (Glc-MU). Glucosidase activities of three consecutive assays with excreted proteins from insect High Five cells were measured. Negative controls (buffer, non-transfected wild-type cells, and cells transfected with green fluorescent protein) did not hydrolyze any defense metabolite. Results from the individual assays are shown in Figure 3—figure supplement 2. For deglycosylation of these compounds by M. melolontha gut protein crude extracts, refer to Figure 3—figure supplement 3. Deglycosylation assays with recombinant Mm_bGlc17 yielded highest aglycone formation; Figure 3—figure supplement 4. Raw data are available in Figure 3—source data 1.
-
Figure 3—source data 1
Source data of main and supplementary figures of Figure 3.
- https://cdn.elifesciences.org/articles/68642/elife-68642-fig3-data1-v1.xlsx
Figure 3—figure supplement 1
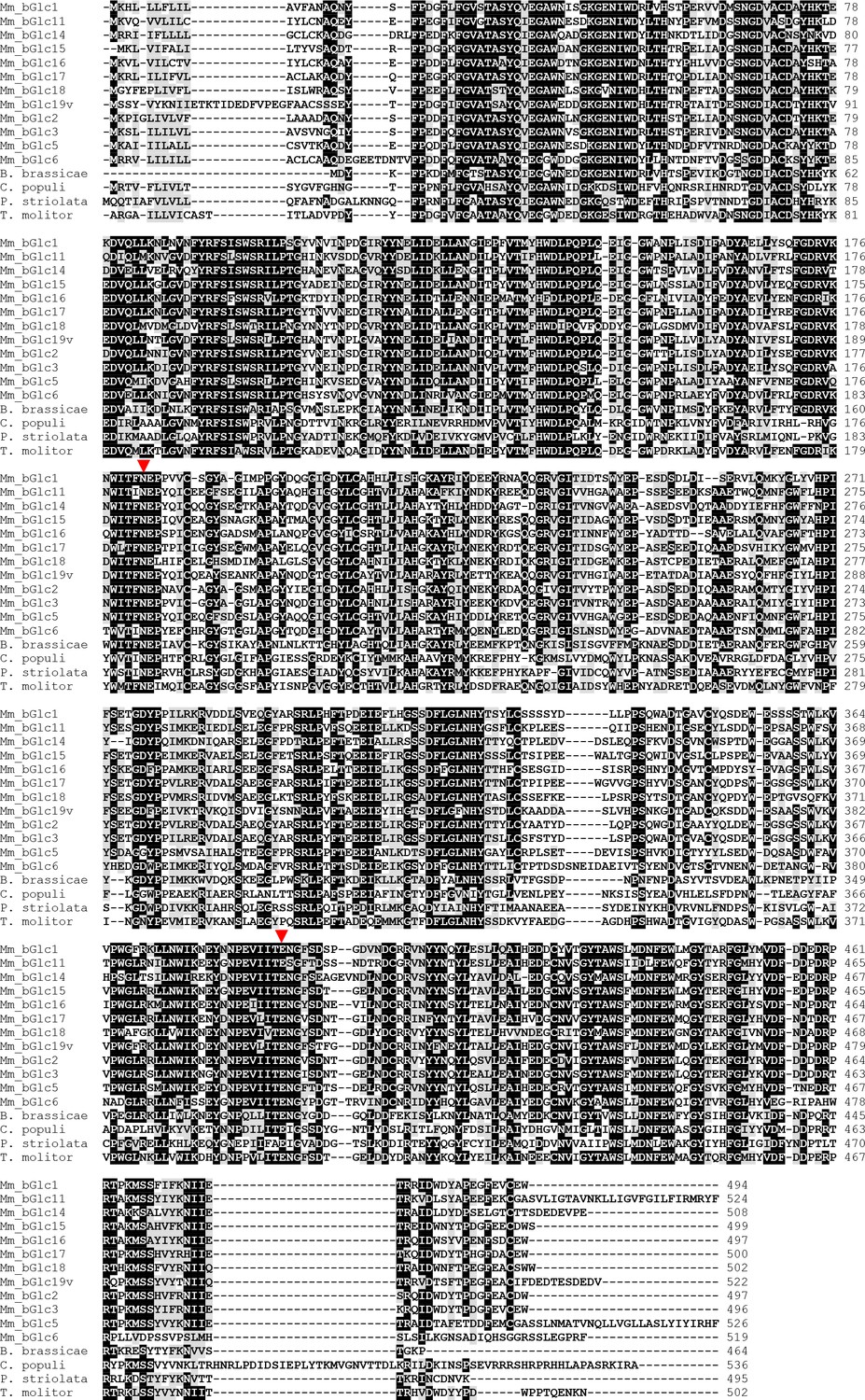
Amino acid sequence alignment of β-glucosidases of Melolontha melolontha and other insect species.
The catalytic glutamates of the glucosidase-characteristic NEP and ITENG motifs are indicated with red triangles.
Figure 3—figure supplement 2
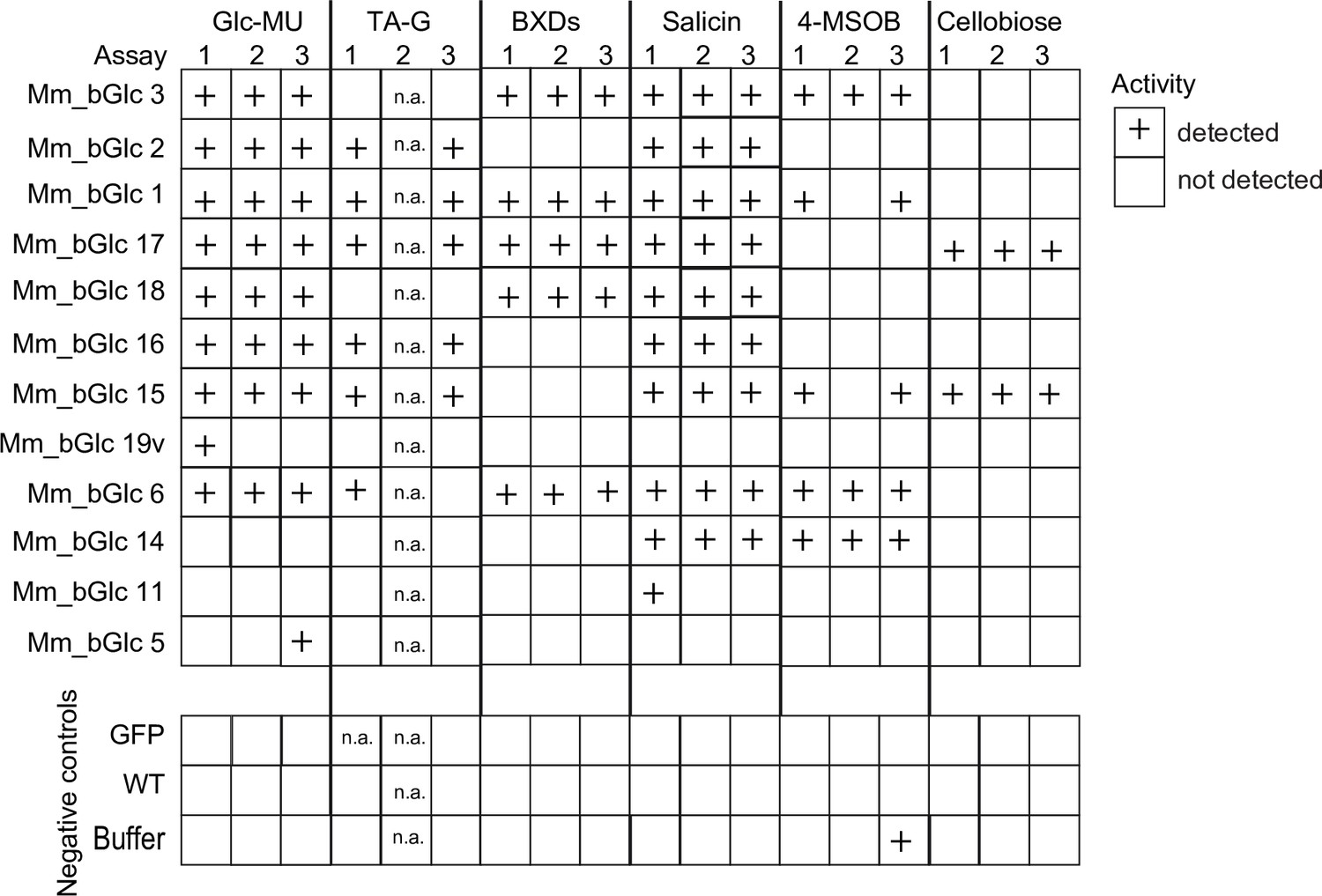
Activity of heterologously expressed Melolontha melolontha β-glucosidases and negative controls (GFP = green fluorescent protein; WT = non-transfected wild type; buffer) toward plant defensive glycosides, cellobiose, and the standard substrate 4-methylumbelliferyl-β-D-glucopyranoside (Glc-MU), which fluoresces upon deglucosylation.
The glucosidase activity of three deglucosylation assays was categorized according to relative activity. Both total protein levels and catalytic activity may contribute to the overall activity. Na = not available.
Figure 3—figure supplement 3
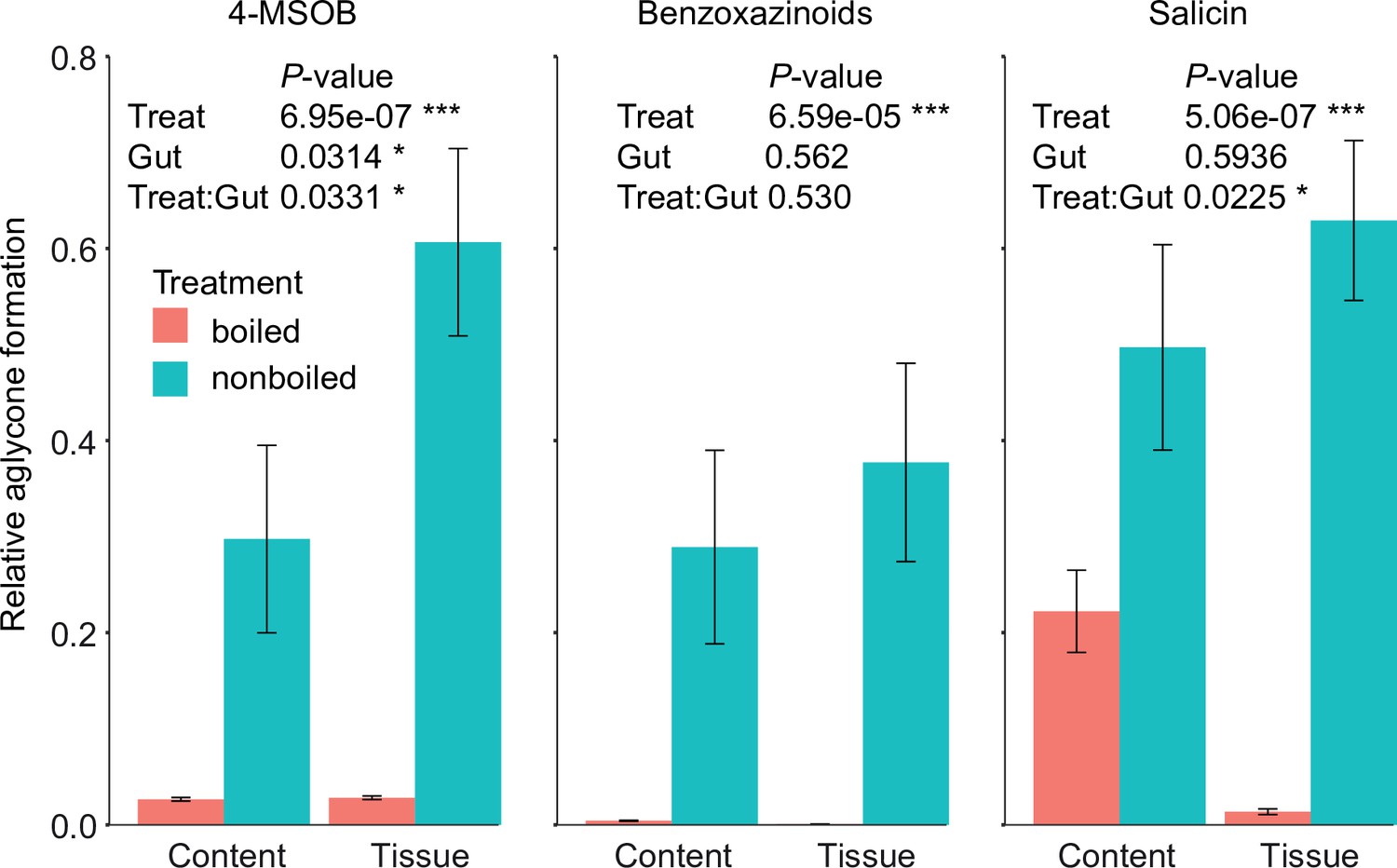
Deglucosylation of defensive glycosides by boiled and non-boiled Melolontha melolontha anterior midgut extracts in vitro.
Aglycone formation was normalized to the maximal peak area of all samples. p-values of two-way analyses of variance (ANOVAs) are shown for each metabolite separately. N = 10. Error bars denote SEM. TA-G = taraxinic acid β-D-glucopyranosyl ester; BXDs = benzoxazinoids; 4-MSOB = 4-methylsulfinylbutyl glucosinolate.
Figure 3—figure supplement 4
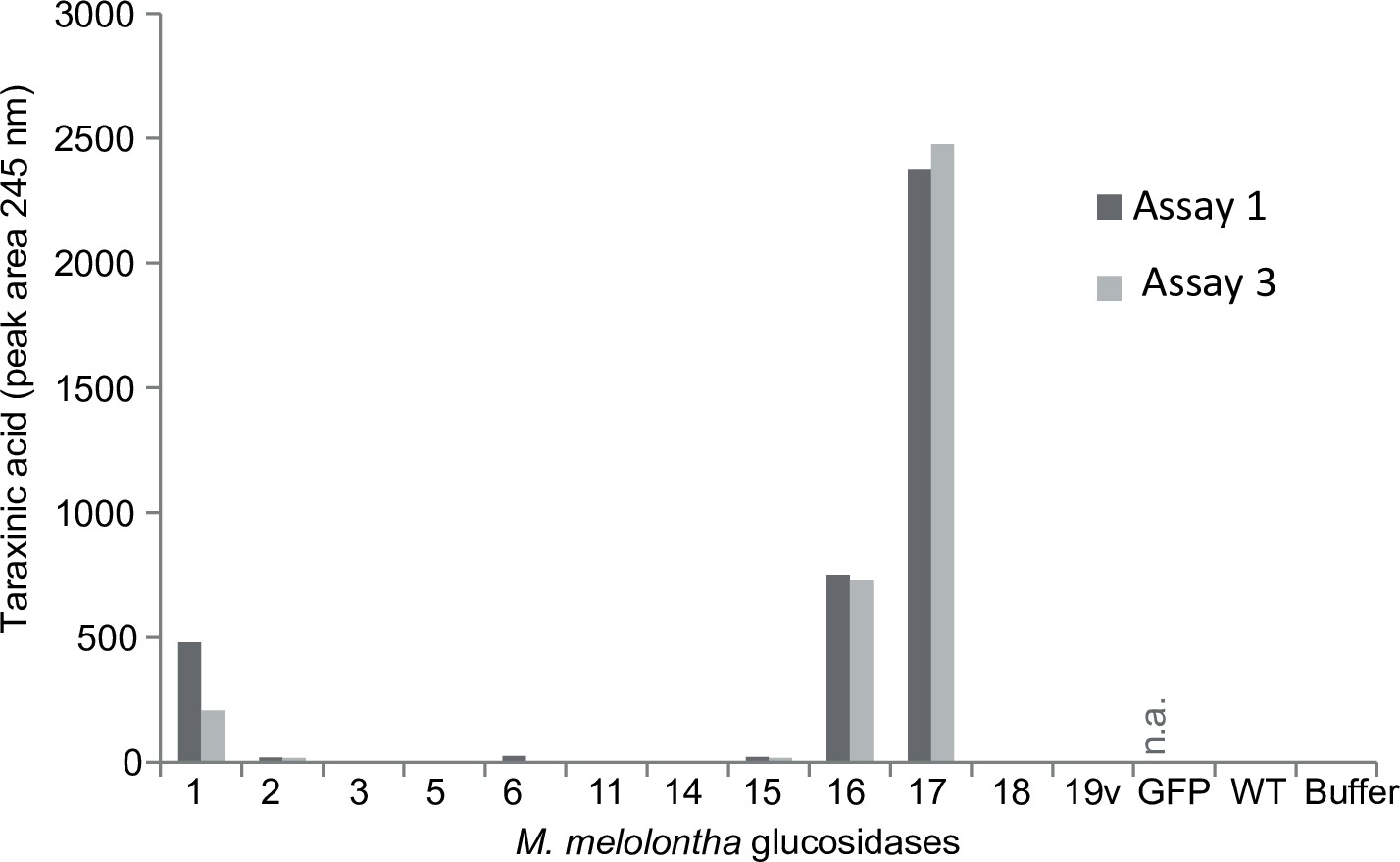
Taraxinic acid aglycone formation of the heterologously expressed Melolontha melolontha β-glucosidases and negative controls (GFP = green fluorescent protein; WT = non-transfected wild type; buffer) of two deglucosylation assays.
Both total protein levels and catalytic activity may contribute to the aglycone formation. Na = not available.
Figure 4 with 6 supplements

Silencing of Mm_bGlc17 reduces TA-G deglucosylation and modifies the impact of TA-G on larval growth and host plant choice.
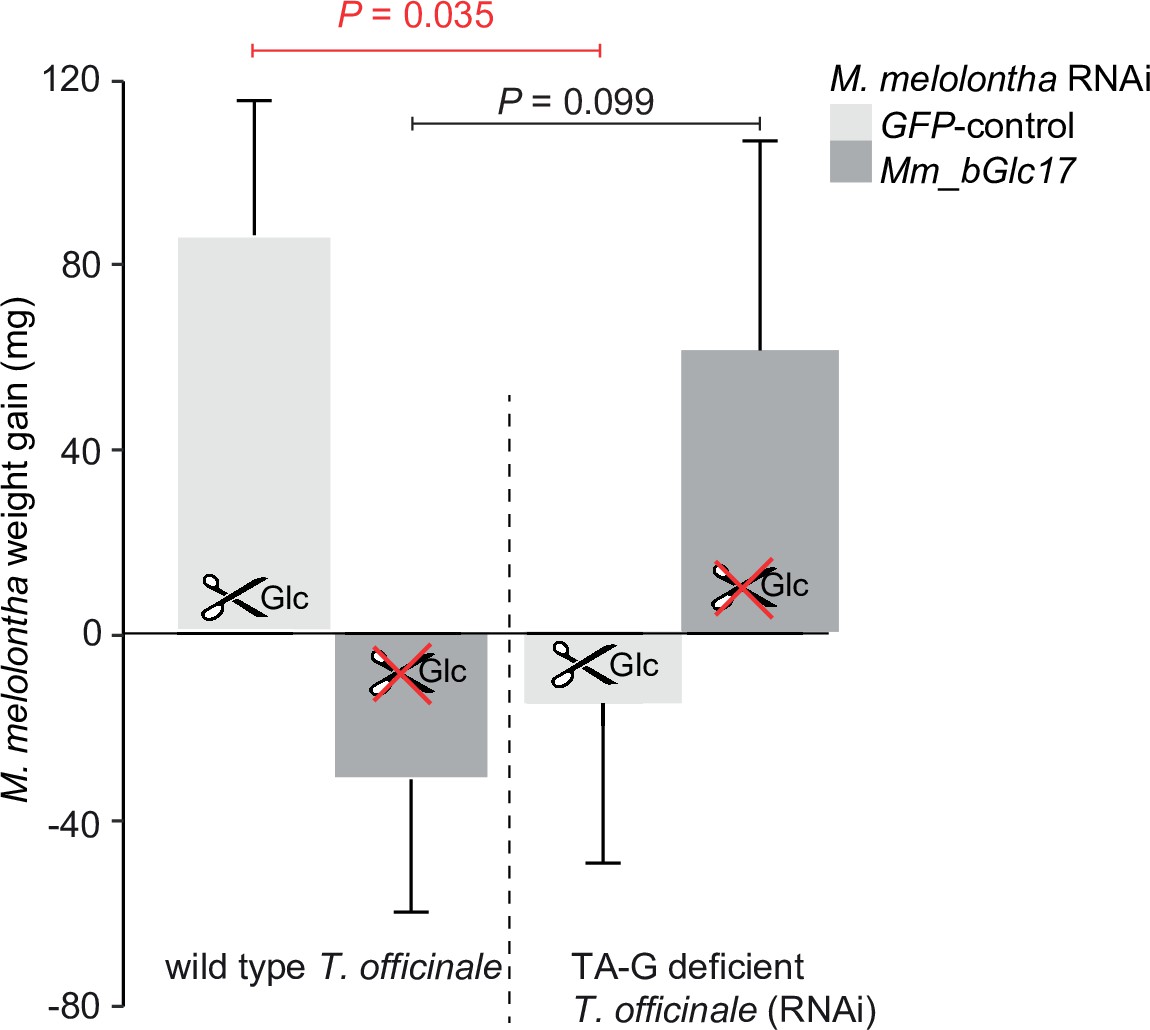
(A) Taraxinic acid β-D-glucopyranosyl ester (TA-G) deglucosylation activity (TA/(TA + TA G)) of gut extracts from Melolontha melolontha larvae in which different β-glucosidases were silenced through RNA interference (RNAi), resulting in stable and specific silencing of the individual glucosidases, Figure 4—figure supplements 2–3. Silencing of Mm_bGlc17 significantly reduced hydrolysis of TA-G by gut extracts. A green fluorescent protein-derived double-stranded RNA (GFP dsRNA) construct was used as a negative control. Mm_bGlc16&17-treated larvae received a 50:50 (v/v) mixture of both dsRNA species. Deglucosylation activity was normalized to that of boiled control samples to correct for the background of non-enzymatic hydrolysis. N = 9–10. p-value of a one-way analysis of variance (ANOVA) is shown. Different letters indicate a significant difference according to Tukey’s honest significance test. Error bars = SEM. (B) Weight gain of Mm_bGlc17-silenced and GFP-control M. melolontha larvae growing on transgenic TA-G-deficient or control Taraxacum officinale lines. N = 11–15. p-values refer to a two-way ANOVA and Student’s t-tests. Error bars = SEM. For comparing growth of GFP- and Mm_bGlc17-silenced larvae between TA-G-deficient and control lines, refer to Figure 4—figure supplement 4. The experiment was repeated once with similar results; Figure 4—figure supplement 5. (C) Gene expression (relative to actin) of Mm_bGlc17-silenced and GFP-control M. melolontha larvae feeding on transgenic TA-G-deficient or control T. officinale lines. N = 12–14. p-values refer to a two-way ANOVA (log-transformed data) and Kruskal-Wallis rank sum tests (non-transformed values). (D) Choice of Mm_bGlc17-silenced and GFP-control larvae between transgenic TA-G-deficient and control T. officinale lines. Silencing of Mm_bGlc17 abolished the choice of control larvae for TA-G-deficient lines. p-values refer to binomial tests. Choice was stable over time; see Figure 4—figure supplement 6. Raw data are available in Figure 4—source data 1.
-
Figure 4—source data 1
Source data of main and supplementary figures of Figure 4.
- https://cdn.elifesciences.org/articles/68642/elife-68642-fig4-data1-v1.xlsx
Figure 4—figure supplement 1
Injection of dsRNA by a sterile syringe between the second and third segment of Melolontha melolontha.
Figure 4—figure supplement 2
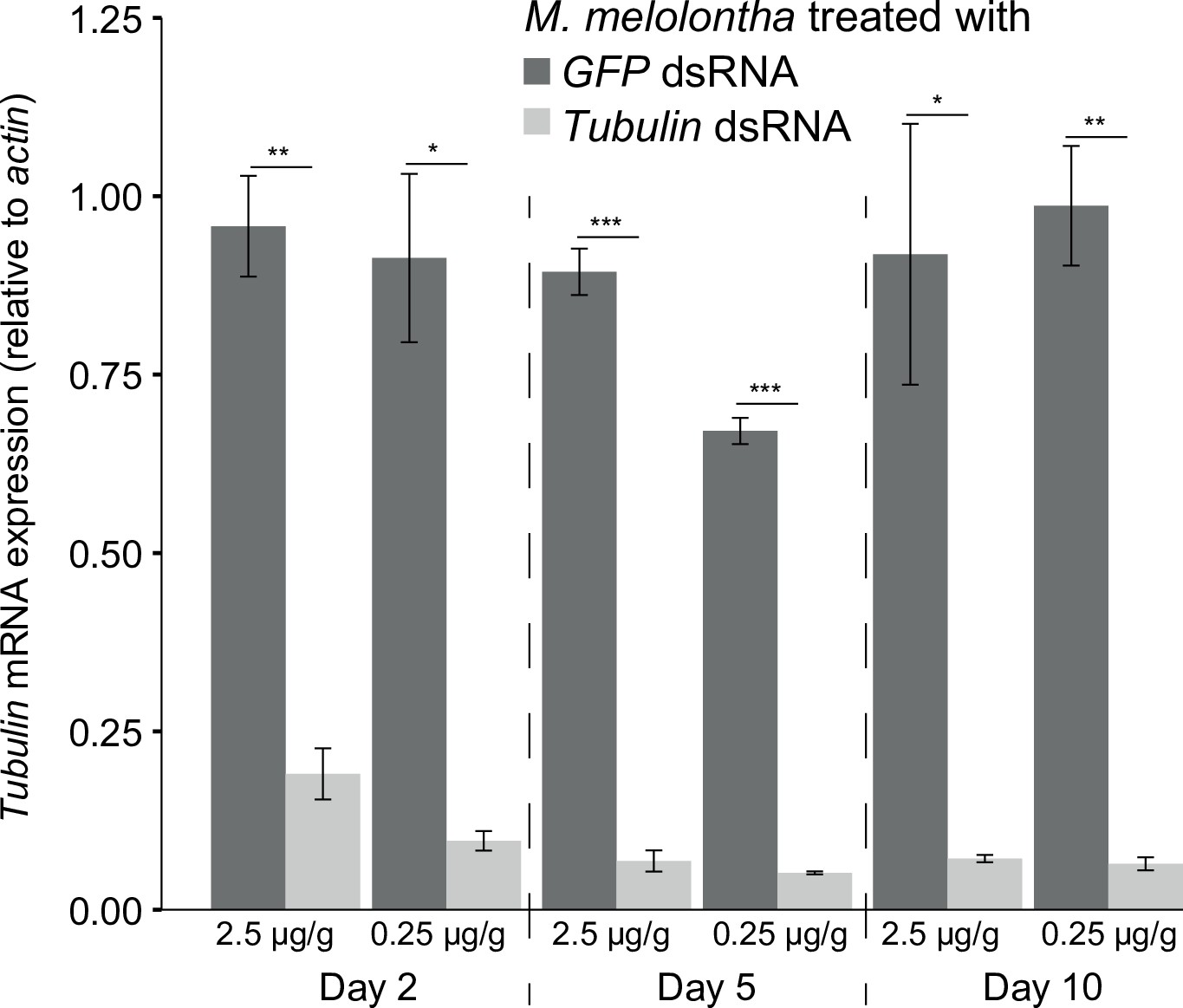
Tubulin mRNA expression in Melolontha melolontha larvae treated with 2.5 µg or 0.25 µg GFP or tubulin dsRNA per g larval mass 2, 5, and 10 days after injection (N = 3).
Error bars denote SEM. Asterisks indicate significant differences in relative tubulin expression between green fluorescent protein (GFP) and tubulin double-stranded RNA (dsRNA)-treated larvae according to the two-tailed Student’s t-test (*p<0.5; **p<0.01; ***p<0.001).
Figure 4—figure supplement 3
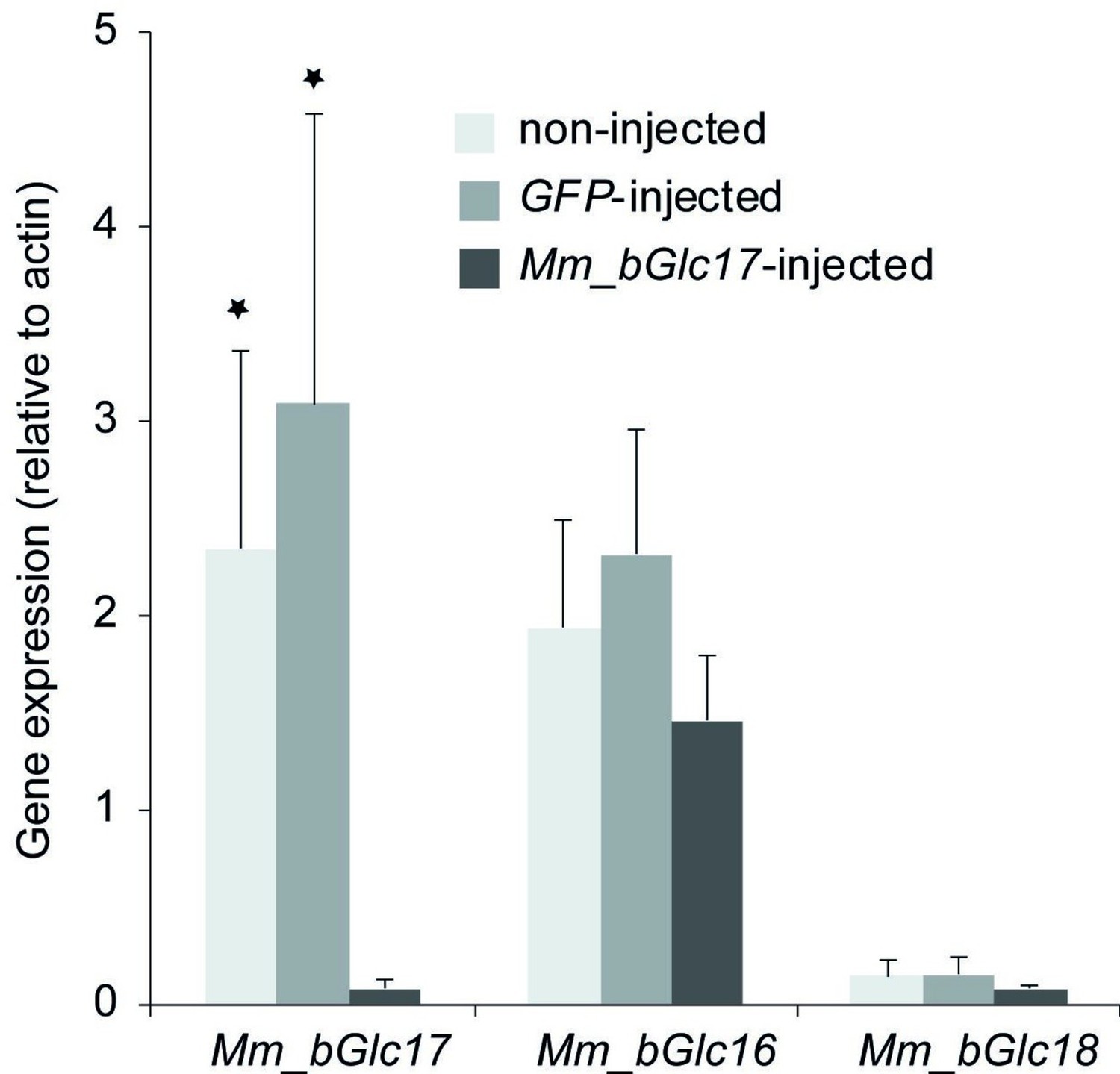
Gene expression (relative to actin) of Mm_bGlc16, Mm_bGlc17, and Mm_bGlc18 in Mm_bGlc17 dsRNA-injected larvae 2 days after dsRNA application.
Asterisks indicate significant differences in the relative gene expression between non-injected and Mm_bGlc17-injected, as well as between green fluorescent protein (GFP)-injected and Mm_bGlc17-injected larvae according to two-tailed Student’s t-tests (*p<0.5). Error bars denote SEM. N = 6–8.
Figure 4—figure supplement 4
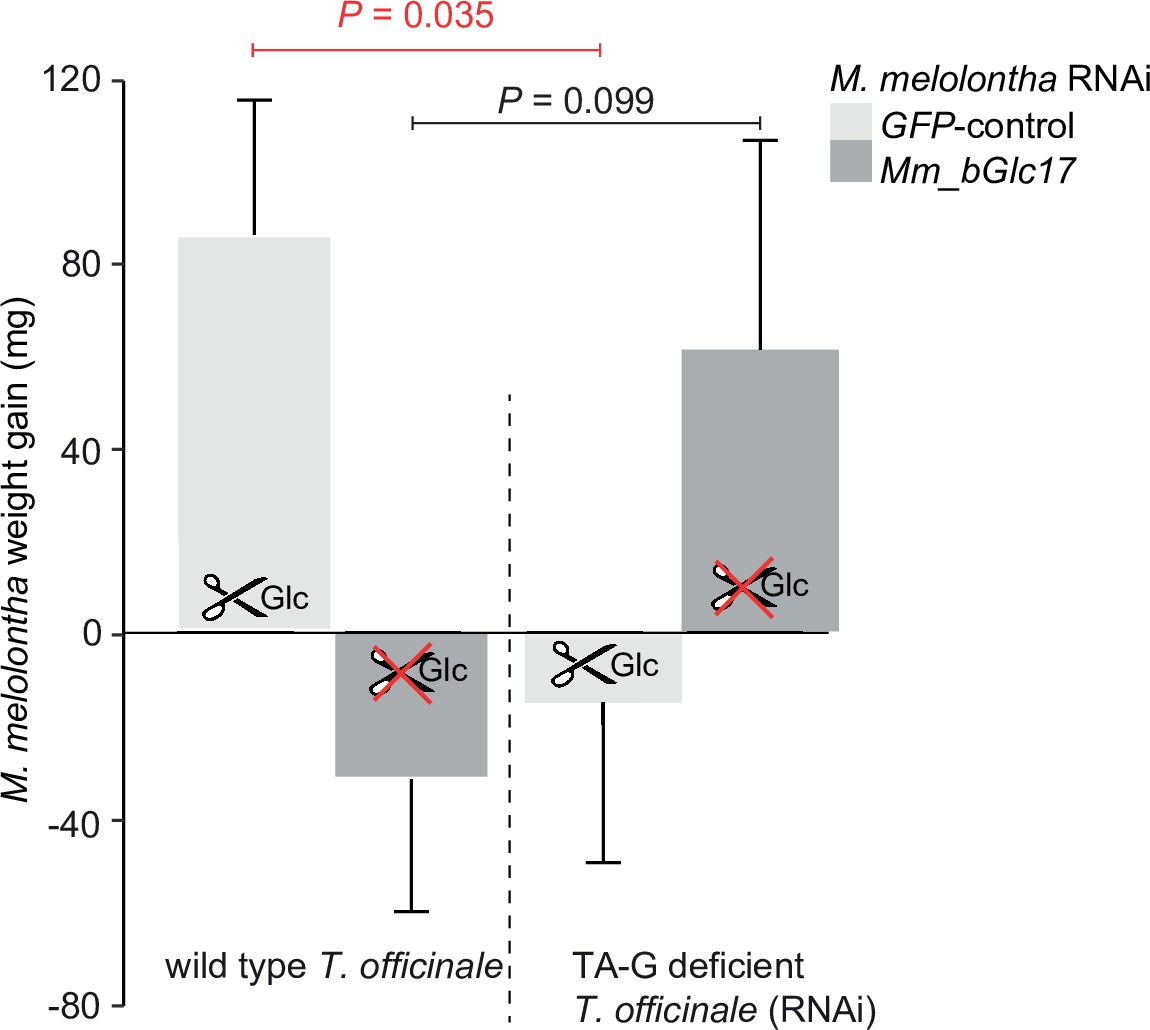
Weight gain of Mm_bGlc17-silenced and GFP-control Melolontha melolontha larvae growing on transgenic TA-G-deficient or control Taraxacum officinale lines (N = 11–15).
p-values refer to Student’s t-tests. Error bars = SEM.
Figure 4—figure supplement 5
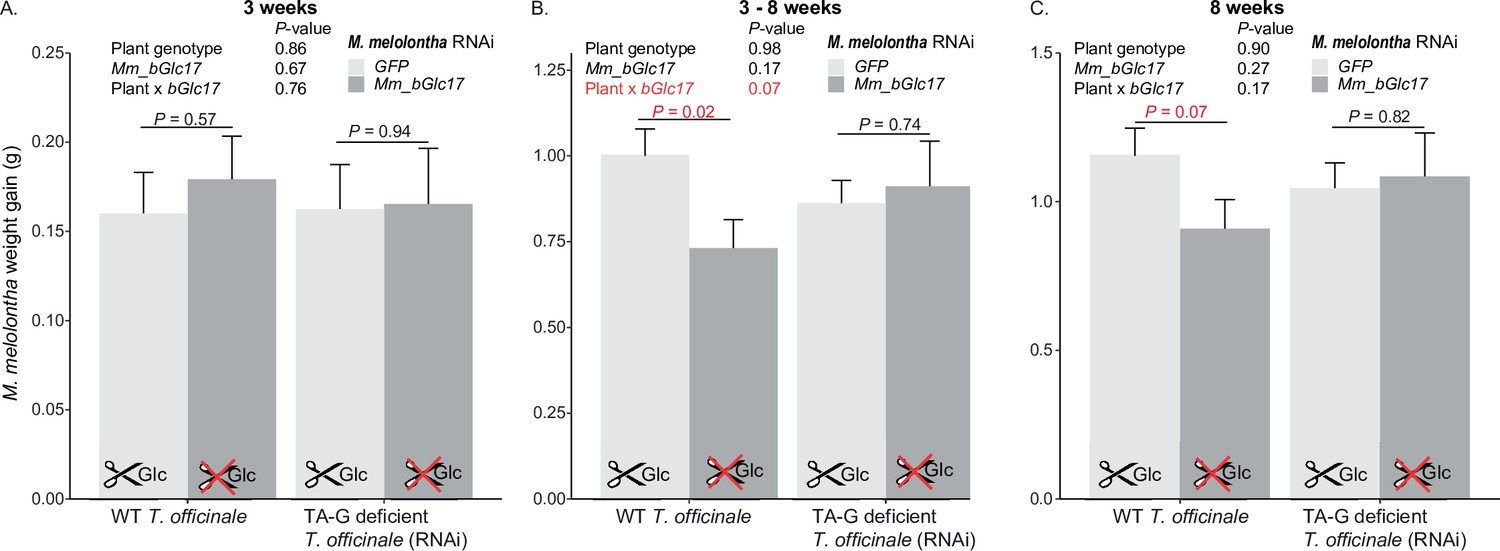
Repetition of the experiment on weight gain of Mm_bGlc17-silenced and GFP-control Melolontha melolontha larvae growing on transgenic TA-G-deficient or control Taraxacum officinale lines.
(A) within the first 3 weeks, (B) 2-8 weeks, (C) 8 weeks (N=12-20) p-values refer to two-way analyses of variance (ANOVAs) and Student’s t-tests. Error bars = SEM.
Figure 4—figure supplement 6

Choice of Melolontha melolontha larvae that were treated with either GFP (A) or Mm_bGlc17 (B) dsRNA between TA-G-deficient transgenic and wild-type Taraxacum officinale plants 1–4 hr after the start of the experiment.
p-values refer to binomial tests. Larvae that were not chosen were excluded from the analysis. GFP = green fluorescent protein; TA-G = taraxinic acid β-D-glucopyranosyl ester; dsRNA = double-stranded RNA.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Melolontha melolontha) | Mm_bGlc17 | This paper | See Supplementary file 1 | |
| Genetic reagent (Taraxacum officinale) | TA-G-deficient plants | Doi:10.1371/journal.pbio.1002332 | A34-RNAi-1 | |
| Genetic reagent (M. melolontha) | Mm_bGlc17 silenced | This paper | See ‘Materials and methods’ (‘TA-G deglucosylation activity in RNAi-silenced M. melolontha larvae’) | |
| Cell line (Trichoplusia ni) | High Five Cells | Life Technologies, Carlsbad, CA, USA | ||
| Transfected construct (T. ni) | pIB/V5-His TOPO | Life Technologies, Carlsbad, CA, USA | ||
| Biological sample (M. melolontha) | M. melolontha | This paper | Collection of different natural populations | |
| Biological sample (T. officianle) | T. officinale | Doi: | Different natural genotypes | |
| Recombinant DNA reagent | Plasmid pGJ 2648 | Other | Supplied by Dr. Christian Schulze-Gronover, Fraunhofer Institute for Molecular Biology and Applied Ecology | |
| Recombinant DNA reagent | pCR2.1-TOPO plasmids | Life Technologies | ||
| Recombinant DNA reagent | cDNAs | This paper | See Supplementary file 3 | |
| Sequence-based reagent | Primers | This paper | See Supplementary file 2 | |
| Sequence-based reagent | KAPA SYBR FAST qPCR Master Mix | Kapa Biosystems | ||
| Commercial assay or kit | innuPREP RNA Mini Kit | Analytik Jena, Jena, Germany | ||
| Commercial assay or kit | RNeasy Plant Mini Kit; RNeasy Lipid Tissue Mini Kit | Qiagen | ||
| Commercial assay or kit | SMARTer RACE cDNA Amplification Kit | Clontech, Mountain View, CA, USA | ||
| Commercial assay or kit | FuGeneHD-Kit | Promega, Madison, WI, USA | ||
| Commercial assay or kit | GeneJET Gel Extraction Kit; DreamTaq DNA Polymerase; MEGAscript RNAi Kit | Thermo Fisher Scientific, Waltham, MA, USA | ||
| Commercial assay or kit | KAPA SYBR FAST qPCR Kit Optimized for LightCycler 480 | Kapa Biosystems, Wilmington, MA, USA | ||
| Chemical compound, drug | Bis(p-nitrophenyl)phosphate; castanospermine; acarbose | Sigma Aldrich | ||
| Chemical compound, drug | Blasticidin; Express Five culture medium | Life Technologies | ||
| Chemical compound, drug | x Protease Inhibitor HP Mix | SERVA Electrophoresis, Heidelberg, Germany | ||
| Chemical compound, drug | TA-G; TA; BXDs; 4-MSOB | This paper | See ‘Materials and methods’ (‘Enzymatic assays of recombinant proteins’ and ‘Synthesis of TA-G metabolite standards’) | |
| Chemical compound, drug | Salicin | Alfa Aeser | ||
| Chemical compound, drug | Cellobiose | Fluka | ||
| Chemical compound, drug | Glc-MU | Sigma Aldrich |
Additional files
-
Supplementary file 1
Nucleotide sequences of the heterologously expressed M. melolontha β-glucosidases.
- https://cdn.elifesciences.org/articles/68642/elife-68642-supp1-v1.docx
-
Supplementary file 2
Primer information.
- https://cdn.elifesciences.org/articles/68642/elife-68642-supp2-v1.docx
-
Supplementary file 3
Fragment nucleotide sequence of tubulin, GFP, Mm_bGlc16, Mm_bGlc17, and Mm_bGlc18 in pCR2.1-TOPO and pIB/V5-His-TOPO vectors.
- https://cdn.elifesciences.org/articles/68642/elife-68642-supp3-v1.docx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/68642/elife-68642-transrepform1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A beta-glucosidase of an insect herbivore determines both toxicity and deterrence of a dandelion defense metabolite
eLife 10:e68642.
https://doi.org/10.7554/eLife.68642




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}