Touch-sensitive stamens enhance pollen dispersal by scaring away visitors
- Institute of Evolution and Ecology, School of Life Sciences, Central China Normal University, China
- Department of Biology, Washington University, United States
Figures
Figure 1 with 2 supplements
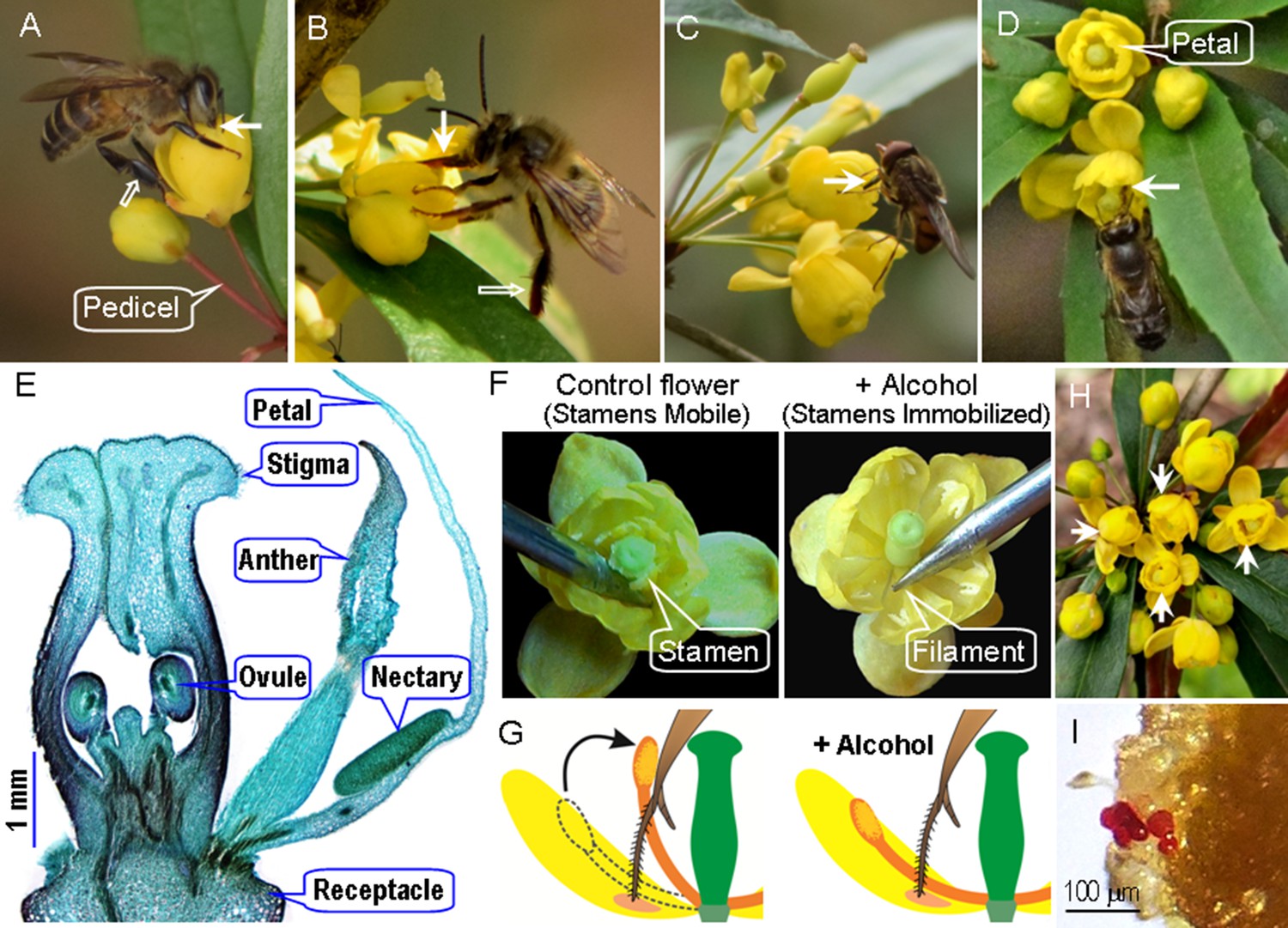
Flower traits, foraging behavior of visitors, and manipulations of stamen movements in Berberis julianae, which has stamens characterized by a touch-sensitive rapid movement toward the flower center.
The major pollinators, workers of Apis cerana (A), and a long-tongued bee, Habropoda sichuanensis (B), sucking nectar while their tongues (arrow) may contact filaments, anthers, and/or stigmas. These bees do not groom Berberis pollen into their corbiculae, and their legs are therefore without pollen loads (hollow arrows). (C) Rhingia campestris feeding on nectar and pollen. A bee visiting two flowers with experimentally immobilized and hence touch-insensitive stamens (D). (E) A cross section of a floral bud, showing the two anther valves and two nectaries at the base of each petal. (F) Natural flower with mobile stamens (left) bending inward when their filament bases are touched by a needle; stamen-immobilized (SI) flower (right) whose pedicel had been immersed in 75% alcohol for over 30 min. (G) Diagram of stamen-mobile and SI flowers, illustrating the stamen movement when a bee’s tongue touches the filament. (H) A floral array on an inflorescence in the field with four alcohol-treated SI flowers (arrows). (I) Stained pollen grains (red) deposited on a stigma under open pollination in the field.
Figure 1—figure supplement 1
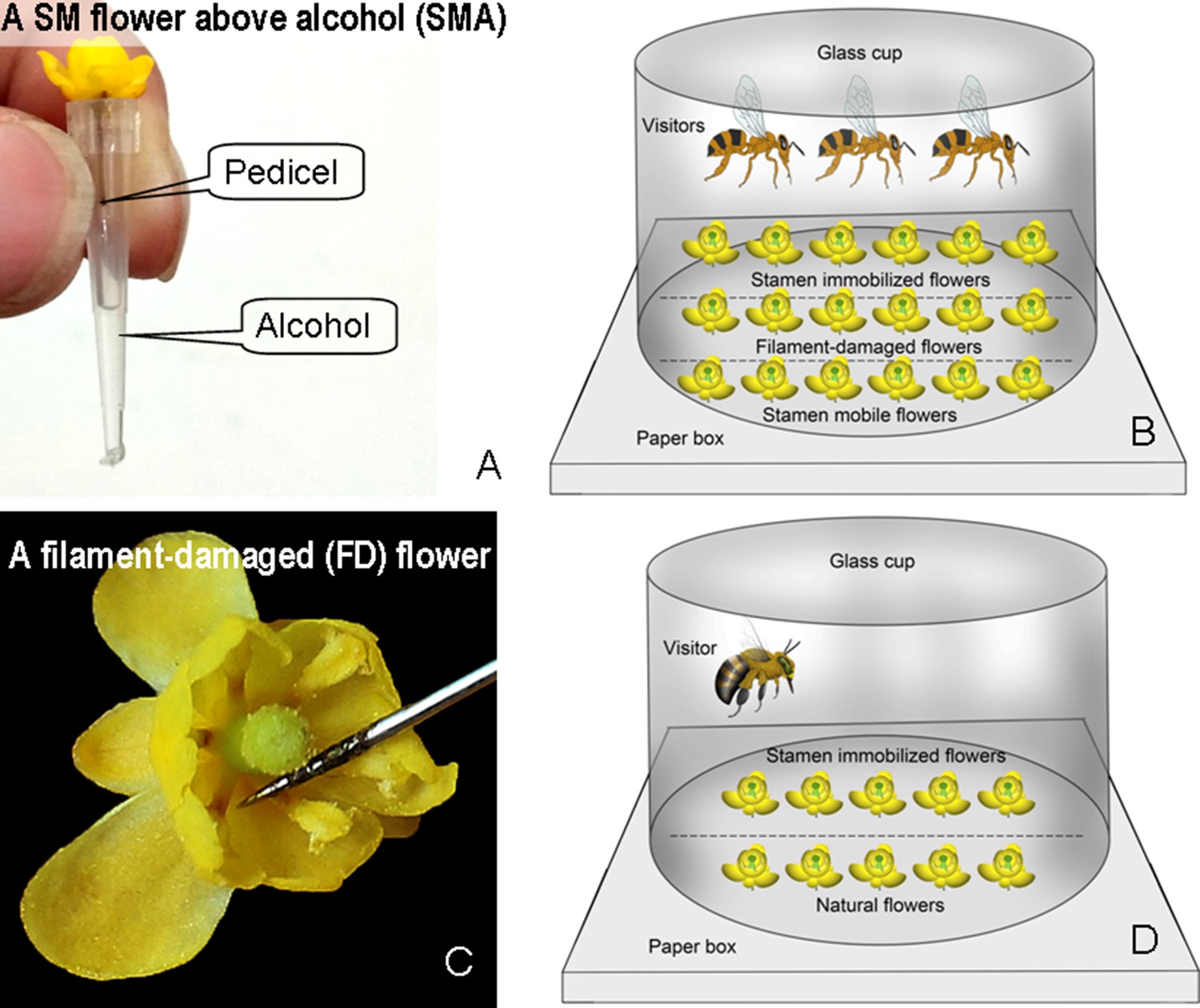
Flower manipulations and experimental floral arrays in Berberis julianae.
(A) An SMA flower with its pedicel fixed above 10 μL of alcohol. (B) An experimental array with stamens-mobile (SM), stamens-immobilized (SI), and filament-damaged (FD) flowers under enclosed conditions with three individuals of Apis cerana. (C) An FD flower in which all filaments have been damaged, but anthers and nectaries are intact. (D) An experimental array with SM and SI flowers enclosed together with one of the four visitor species caught while foraging on flowers nearby.
Figure 1—figure supplement 2

Habit, floral traits, developing berries, and feeding behavior of flower visitors in three Berberidaceae species whose stamens are touch-sensitive.
Bird’s eye view of a flowering individual in an open habitat (A) and an infructescence (B) of Berberis julianae in Sichuan. A hawkmoth (C) and a bumblebee worker (Bombus friseanus Skorikov, 1933) (D) drinking nectar from Berberis jamesiana in Yunnan. (E) A worker of Apis cerana drinking nectar from Mahonia bealei in Wuhan; its tongue has triggered the anthers moving inward (red arrow). (F) Pollen grains on the tongue of A. cerana visiting M. bealei. The bees’ legs are without visible pollen loads (pink circles), showing that these bees do not actively collect M. bealei pollen. (G) Cross section of an M. bealei flower showing the two anther valves with pollen grains attached.
Figure 2
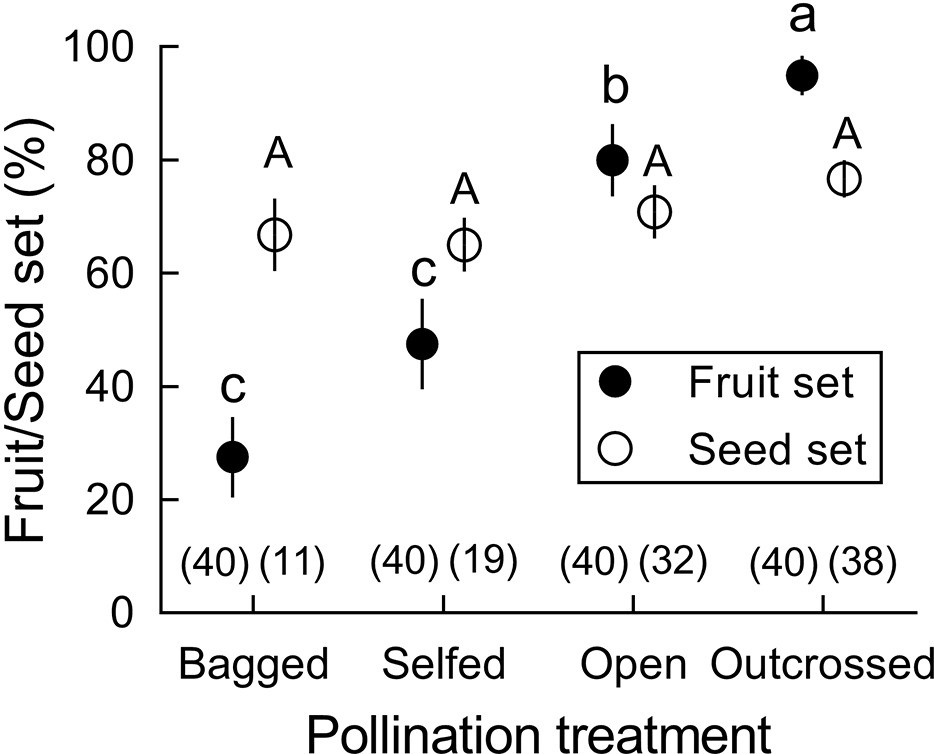
Fruit and seed set (mean ± Standard Error) after four pollination treatments in Berberis julianae.
Different letters beside mean values indicate significant differences among the four treatments under a generalized linear model (GLM). Fruit set differed significantly (Wald χ2 = 34.598, p<0.001, df = 3) but seed set per fruit did not when zero data were excluded (Wald χ2 = 1.973, p=0.578, df = 3). Sample size for each treatment is given in brackets above the X-axis.
Figure 3

Visitation rates (A, C) and handling times (B, D) of Apis cerana in four treatments of flowers of Berberis julianae.
Stamens mobile (SM, controls), stamens immobilized (SI), natural flowers in a fixed position above alcohol (SMA), and flowers with their filaments damaged (FD) so that the stamens became immobile but retained their pollen sacs and the nectar glands to the right and left of each filament. The box plots indicate the median (mid lines), the interquartile range (boxes), and 1.5× the interquartile range (whiskers). Different lowercase letters indicate significant differences, and the numbers in brackets indicate sample sizes. Sample sizes (n) are given in brackets.
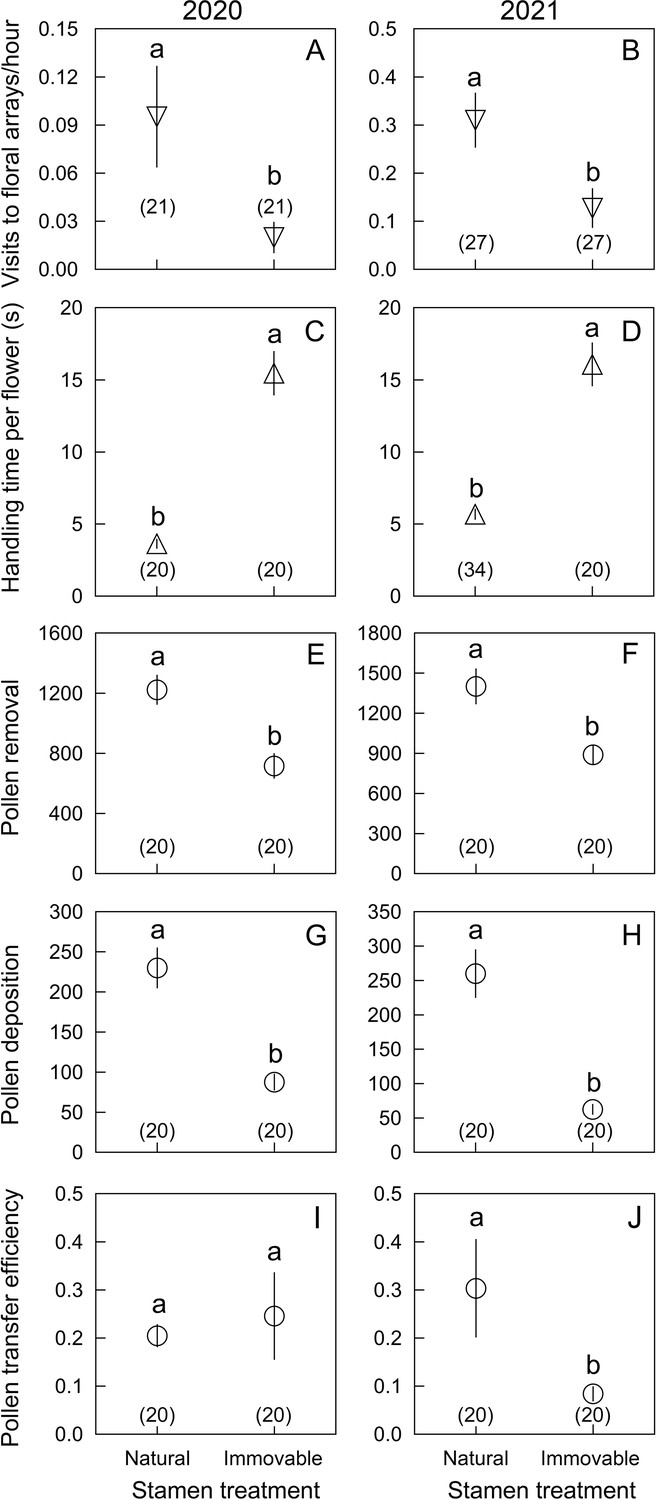
Figure 4
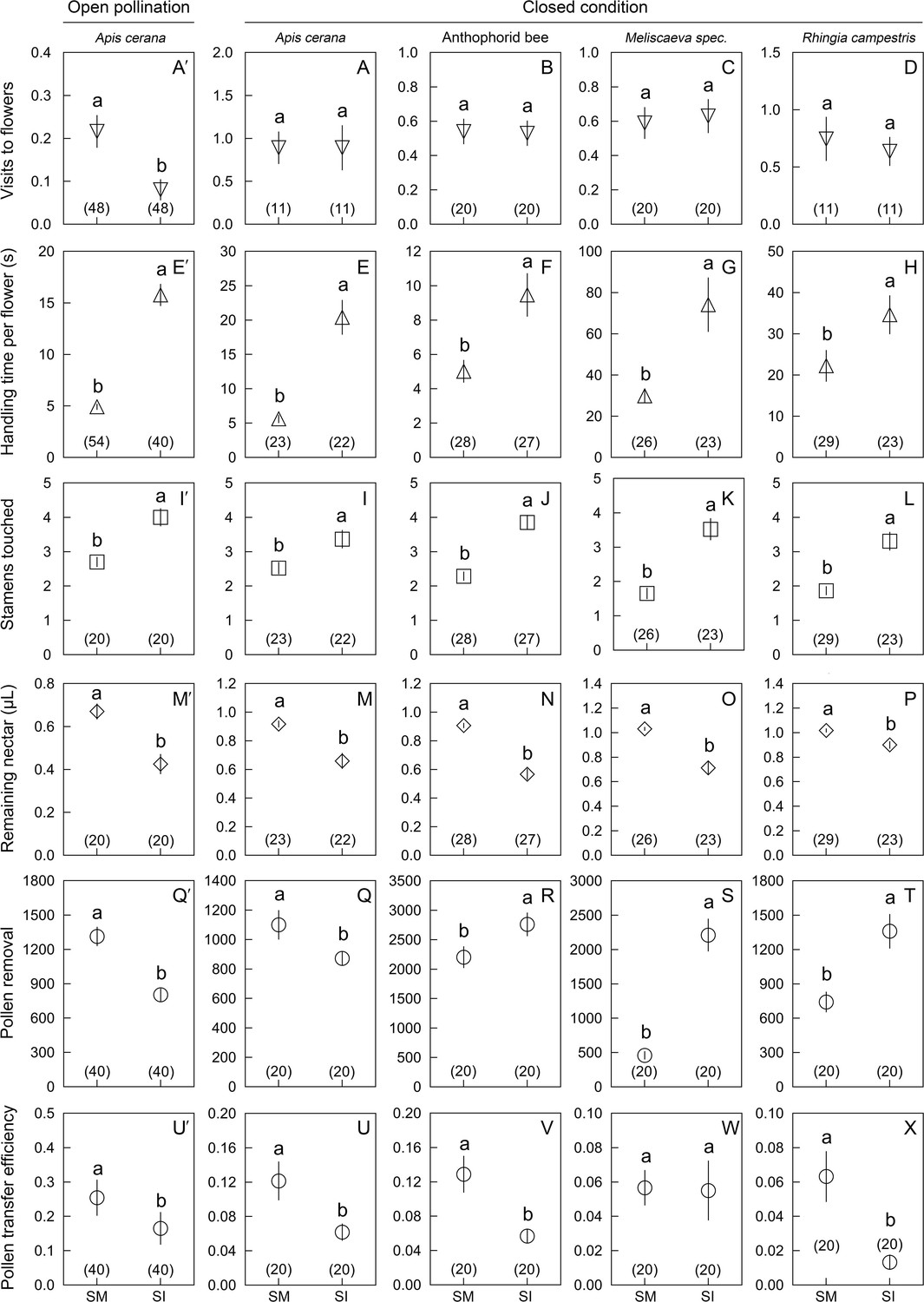
Comparisons of six parameters (mean ± Standard Error) in Berberis julianae to examine the effects of stamen movements on insect visitor foraging behavior and their roles in pollination.
The major pollinator, Apis cerana, was studied in 2020 and 2021 under open pollination (far left) and enclosed conditions (set-up shown in Figure 1—figure supplement 1 and Video 3), whereas the anthophorid bee pollinators and the two flies (Meliscaeva spec. and Rhingia campestris) were compared under enclosed conditions in 2021. Different lowercase letters above error bars indicate significant differences between control (stamen mobile [SM]) and alcohol-treated (stamen immobilized [SI]) flowers. (A′–D) Visitation rates of four visitor species, showing that A. cerana visited control flowers more frequently than SI flowers under open pollination (A′), but no visitor species discriminated between SI and SM flowers under enclosed conditions (A–D). All visitor species spent more time (E′–H) and touched more stamens (I′–L) in SI flowers than in control SM flowers. Visitors removed more nectar from SI flowers, resulting in less nectar remaining per flower (M′–P). Pollen removal by A. cerana was lower from SI than from SM flowers (Q′, Q), but higher in the other three visitor species (R–T). Compared to SM flowers, pollen transfer efficiency was significantly decreased in SI flowers (U′, U, V, X), although it did not differ in Meliscaeva spec. (W). Sample sizes (n) are given in brackets.
Figure 5 with 1 supplement
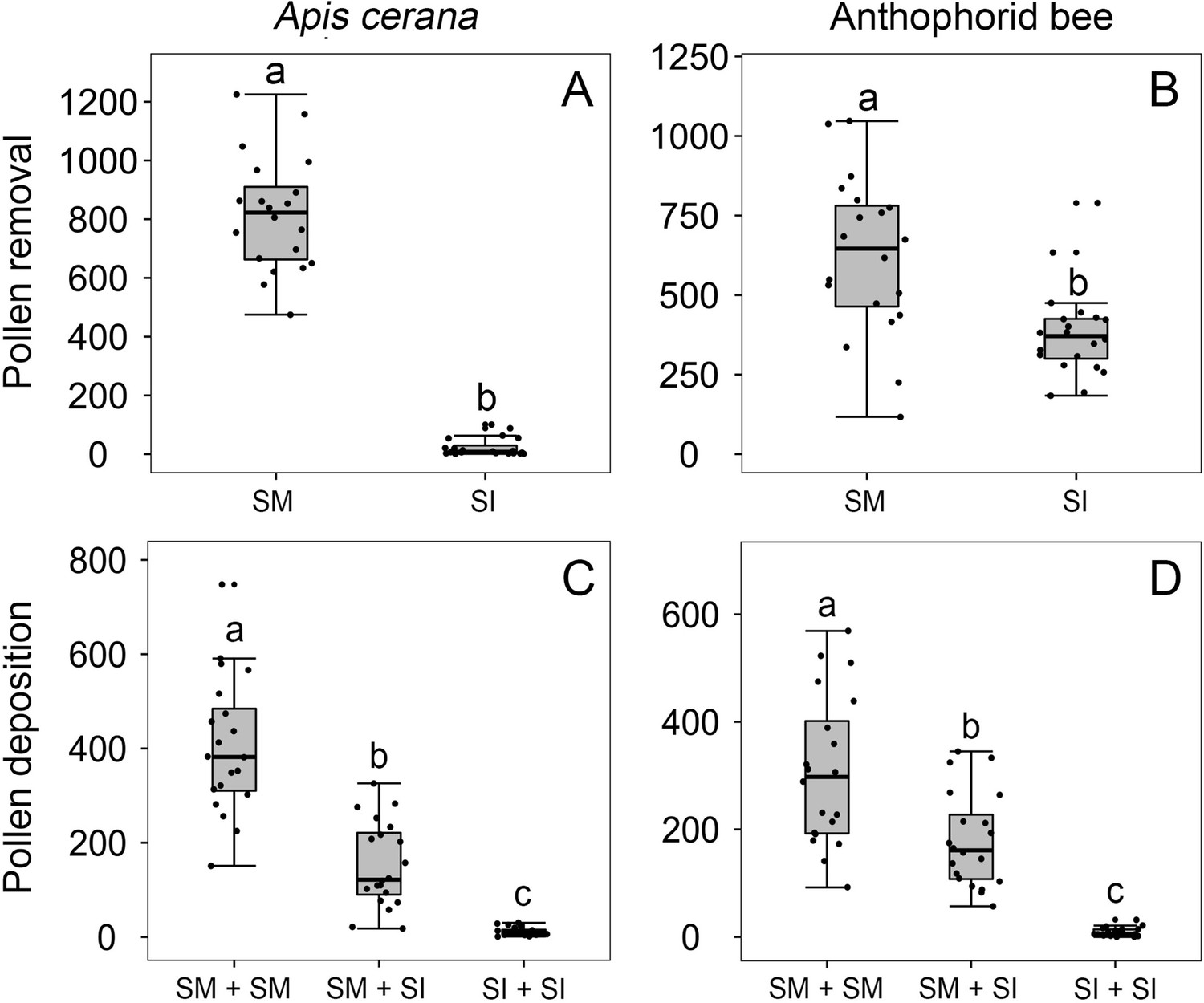
Effects of stamen forward-snapping on pollen placement on the pollinator body and pollen deposition on stigmas after single visits by Apis cerana (A, C) and the anthophorid bees (B, D).
Numbers of pollen grains placed on bees’ tongues during a single visit were significantly higher when stamens were mobile (SM) than when stamens were experimentally immobilized (SI) (A, B). Numbers of pollen grains deposited on the stigma of the second-visited flower (pollen recipient) during single visits by A. cerana (C) and the anthophorids (D) in three trials with the sequence being SM + SM flowers; SM + SI flowers; and SI + SI flowers. The box plots indicate the median (mid lines), the interquartile range (boxes), and 1.5× the interquartile range (whiskers). Different lowercase letters indicate significant differences among three trials.
Figure 5—figure supplement 1
Foraging behavior of Apis cerana and its effect on pollination in Berberis julianae flowers with experimentally immobilized stamens and controls in 2020 and 2021 including visits to floral arrays per flower (A, B), insect handling time per flower (C, D), and pollen removal (E, F), pollen deposition (G, H) and pollen transfer efficiency (I, J) by single visits.
Mean and SEs (standard-error bars) are presented, with different lowercase letters indicating significant differences between control (SM) and stamen-immobilized (SI) flowers.
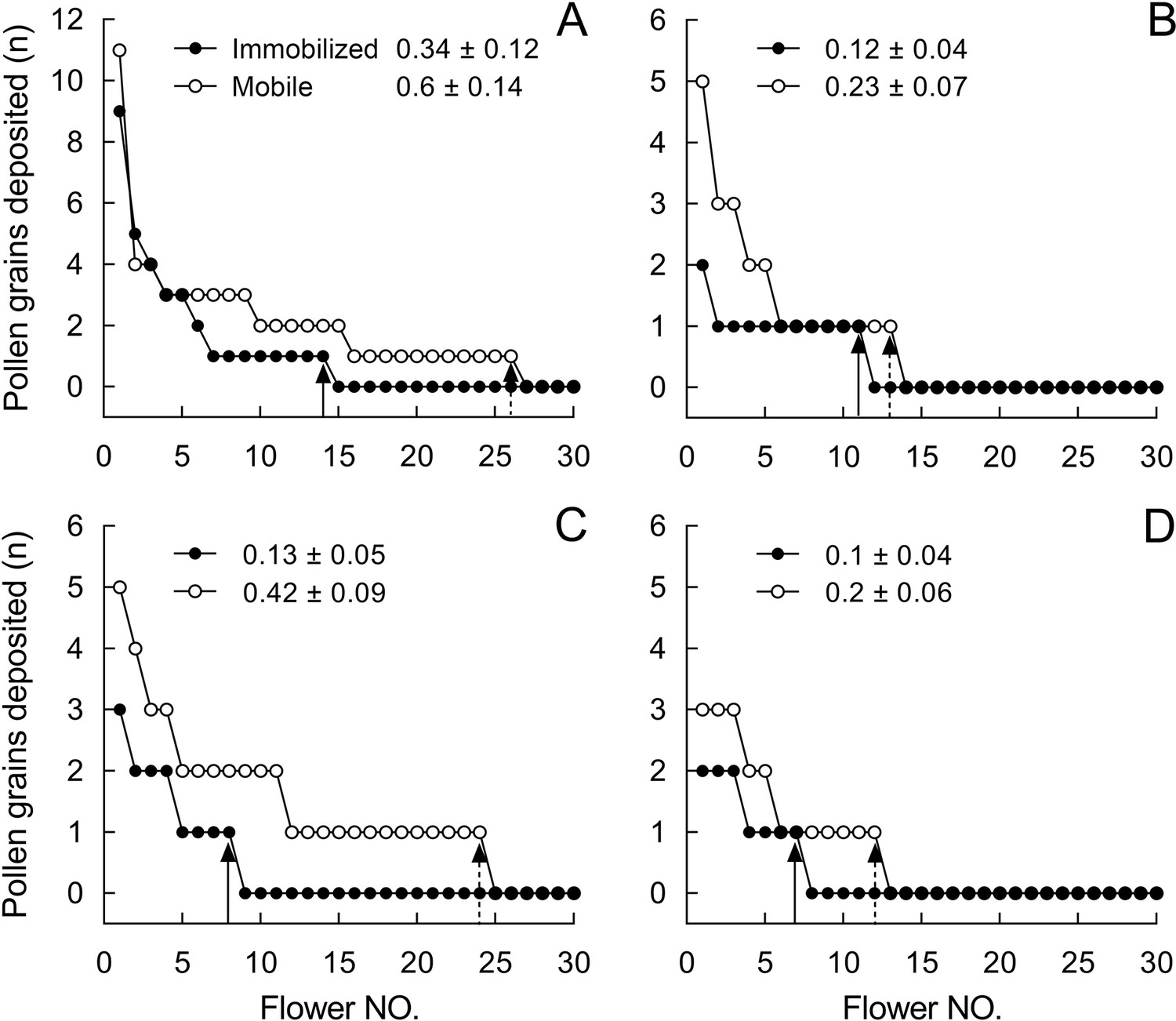
Figure 6 with 1 supplement

Number of stained pollen grains deposited on the stigmas from control flowers with mobile stamens (open circles) and from flowers with experimentally immobilized stamens (closed circles) in Berberis julianae.
Mean pollen number and SEs (numbers of flowers with stained pollen deposition/total number of sampled flowers of pollen recipients) are given for treated and control flowers in each of four trials (A–D). Note that only 15 flowers are shown, although each pollen-tracking test sampled over 100 flowers to examine the effect of stamen movements on pollen dispersal; for example, of 260 pollen-recipient flowers, only 4 flowers received pollen grains from stained stamens-immobilized (SI) flowers, that is, (4/260) in (A).
Figure 6—figure supplement 1
Number of stained pollen grains deposited on the stigmas from control flowers with mobile stamen (open circles) and from flowers with experimentally immobilized stamens (closed circles) in Berberis jamesiana.
Mean pollen number (n = 100 flowers) and SEs are given in each of four trials (A–D). Each pollen-tracking test involved 100 pollen-recipient flowers, but the 70 additional flowers not receiving any pollen are not shown.
Videos
Video 1
Stamens of Berberis julianae become touch-insensitive after the flower pedicels had been immersed in 75% alcohol for 35 min.
Video 2
Stamens of Mahonia bealei become touch-insensitive after the flower pedicels had been immersed in 75% alcohol for 30 min.
Video 3
Under enclosed conditions (Figure 1—figure supplement 1D), individual syrphid flies (Meliscaeva spec.) took up nectar from a Berberis julianae flower with immobilized stamens for much longer, giving them time to touch four stamens, while leaving more quickly and touching only two stamens in a control flower with mobile stamens.
Additional files
-
Supplementary file 1
Floral traits (mean ± SE) and duration of stamen movements in Berberis jamesiana, B. julianae, B. forrestii, and Mahonia bealei.
- https://cdn.elifesciences.org/articles/81449/elife-81449-supp1-v1.docx
-
Supplementary file 2
Foraging behaviors of two types of major insect visitors to Berberis julianae.
Behaviors of the bees and flies visiting Berberis julianae, focusing on visitation rates (visits per flower per hour, mean ± SE), visits per flower, handling time, number of stamens touched, nectar volume remaining per flower, pollen grains removed, pollen grains deposited, and pollen transfer efficiency.
- https://cdn.elifesciences.org/articles/81449/elife-81449-supp2-v1.docx
-
Supplementary file 3
G-test of independence confirming that Berberis julianae flowers with mobile stamens (SM flowers) donated pollen to more recipient flowers at three distance classes than did flowers with experimentally immobilized stamens (SI flowers).
- https://cdn.elifesciences.org/articles/81449/elife-81449-supp3-v1.docx
-
Supplementary file 4
G-test of independence confirming that Berberis jamesiana flowers with mobile stamens (SM flowers) donated pollen to more recipient flowers at two distance classes than did flowers with experimentally immobilized stamens (SI flowers).
- https://cdn.elifesciences.org/articles/81449/elife-81449-supp4-v1.docx
-
Supplementary file 5
Concentrations of berberine (mean ± SE) in different tissues of Berberis julianae under generalized linear models.
- https://cdn.elifesciences.org/articles/81449/elife-81449-supp5-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/81449/elife-81449-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Touch-sensitive stamens enhance pollen dispersal by scaring away visitors
eLife 11:e81449.
https://doi.org/10.7554/eLife.81449




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}