Habitat fragmentation mediates the mechanisms underlying long-term climate-driven thermophilization in birds
- MOE Key Laboratory of Biosystems Homeostasis & Protection, College of Life Sciences, Zhejiang University, China
- Department of Ecology and Evolutionary Biology, University of California, Los Angeles, United States
- Center for Global Change and Ecological Forecasting, Zhejiang Zhoushan Island Ecosystem Observation and Research Station, Institute of Eco Chongming, Zhejiang Tiantong Forest Ecosystem National Observation and Research Station, School of Ecological and Environmental Sciences, China
Figures
Figure 1

The general process of thermophilization and the hypothesized framework of thermophilization in fragmented habitats.
Two general processes of thermophilization are shown on the top: increasing colonization rate of warm-adapted species, and increasing extinction rate of cold-adapted species over time (a). Compared to continuous habitats, a hotter microclimate caused by lower buffering ability on fragmented patches may attract more warm-adapted species to colonize while causing cold-adapted species to extirpate or emigrate faster (b). Loss of cold-adapted species can also be exacerbated when highly fragmented patches harbor lower habitat heterogeneity (e.g. resource, microrefugia), which will also reduce colonization of warm-adapted species (c). Isolated patches due to habitat fragmentation will block warm-adapted species’ colonization and cold-adapted species’ emigration under warming (d). The three distinct patches signify a fragmented landscape and the community in the middle of the three patches was selected to exemplify colonization-extinction dynamics in fragmented habitats. Relative species richness is shown by the number of bird silhouettes in the community . Note that extinction here may include both the emigration of species and then the local extinction of species and that CTI can also change simply due to differential colonization-extinction rates by thermal affinity if the system is not at equilibrium prior to the study. In our study system, we have no way of knowing whether our island system was at equilibrium at onset of the study, thus, focusing on changing rates of colonization-extinction over time presents a much stronger tests of thermophilization.
Figure 2 with 1 supplement

The location of Thousand Island Lake and the experimental islands.
The map was created using ESRI (Environmental Systems Resource Institute) ArcMap software (version 10.3). The base map sources include Esri, Maxar, Earthstar Geographics, and the GIS User Community.
Figure 2—figure supplement 1
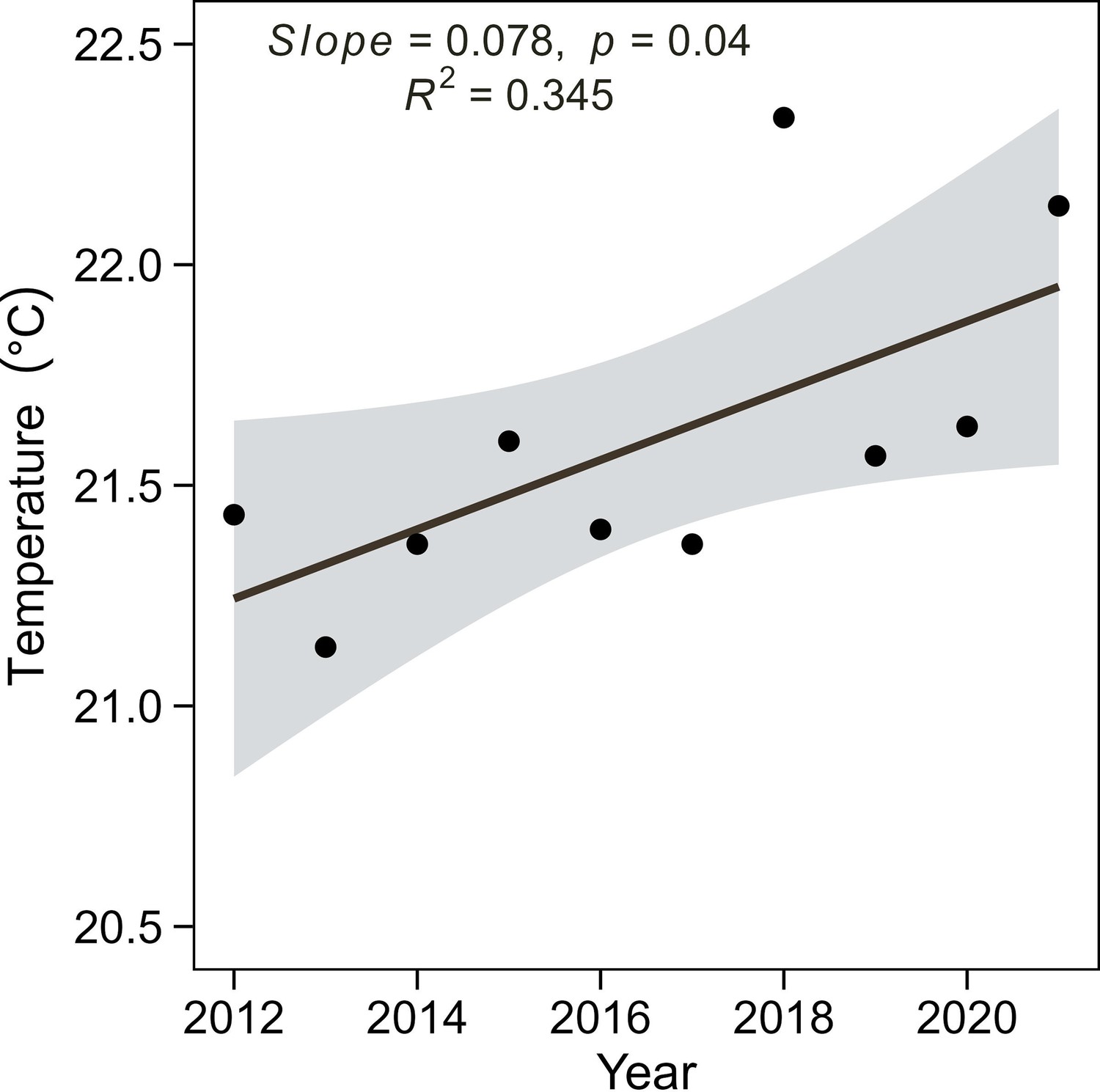
Breeding season temperature from the year 2012–2021 in the Thousand Island Lake, China.
Breeding season temperature was calculated as the average of the mean monthly temperature for breeding seasons (April – June). Black lines and grey shade indicate the regression line and 95% confidence interval of the linear regression model (slope = 0.078, r2=34.53%, p=0.04).
Figure 3 with 2 supplements

Temporal trend in CTIoccur and CTIabun in the TIL.
Vertical lines in (a) are posterior estimated CTI (mean ± SD); solid black line and shaded area are predictions and 95% credible intervals extracted from JAGS modeling posterior mean CTIoccur (n = 530) as a function of year, and using island identity as a random effect while accounting for the variation in posterior CTIoccur. Points and error bars in (b) are observed CTIabun (mean ± 2 SD); solid black line and shaded area are predicted values and 95% confidence intervals estimated from LMM modeling observed CTIabun (n = 530) as a function of year, and using island identity as a random effect.
Figure 3—figure supplement 1
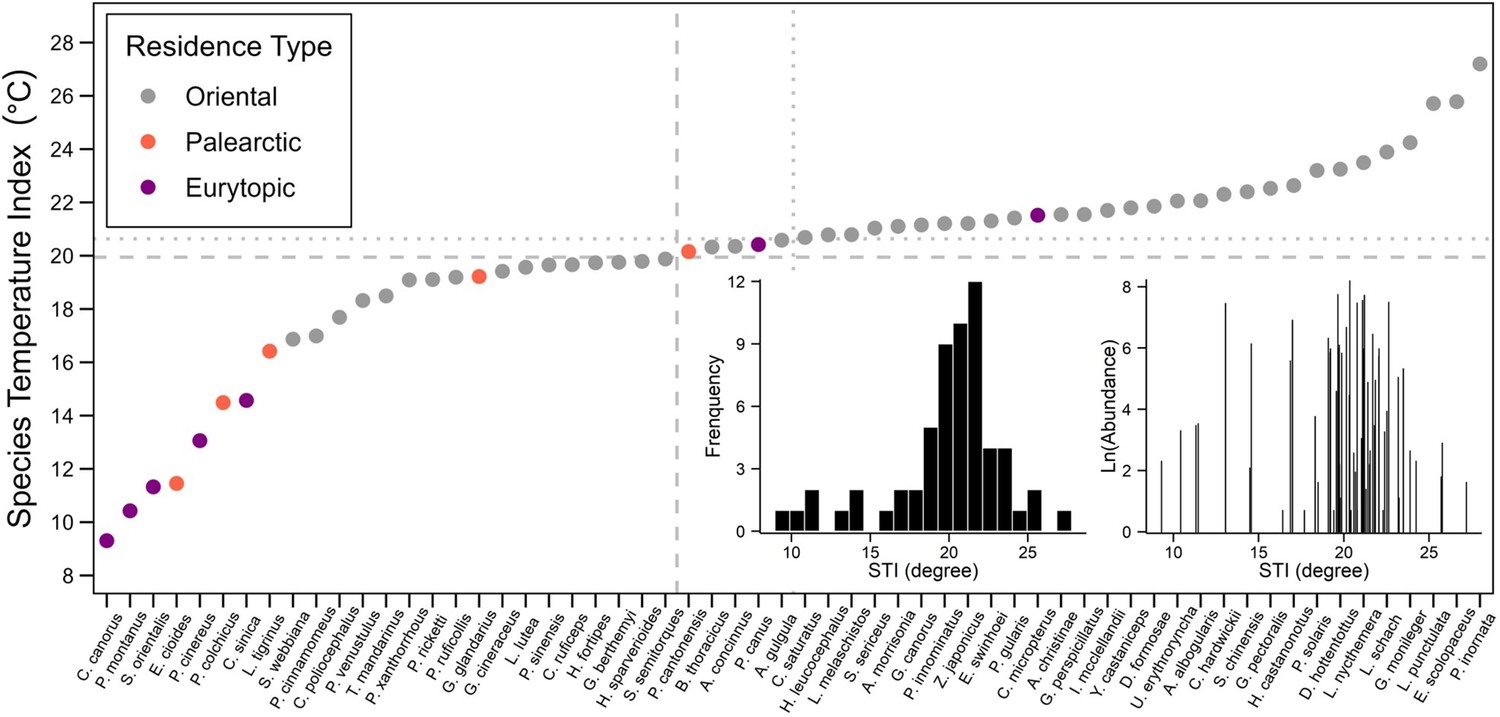
Species temperature index of 60 bird species in the Thousand Islands Lake, China.
The dotted horizontal line indicates the median of STI values; the dashed horizontal line indicates the mean of STI values. Grey points are Oriental species, red points are Palearctic species and purple points are Eurytopic species. The bottom right presents the frequency of STI values (right) and the overview relationship between total abundance (natural logarithm transversion of total abundance over 10 years and on all islands) and STI.
Figure 3—figure supplement 2
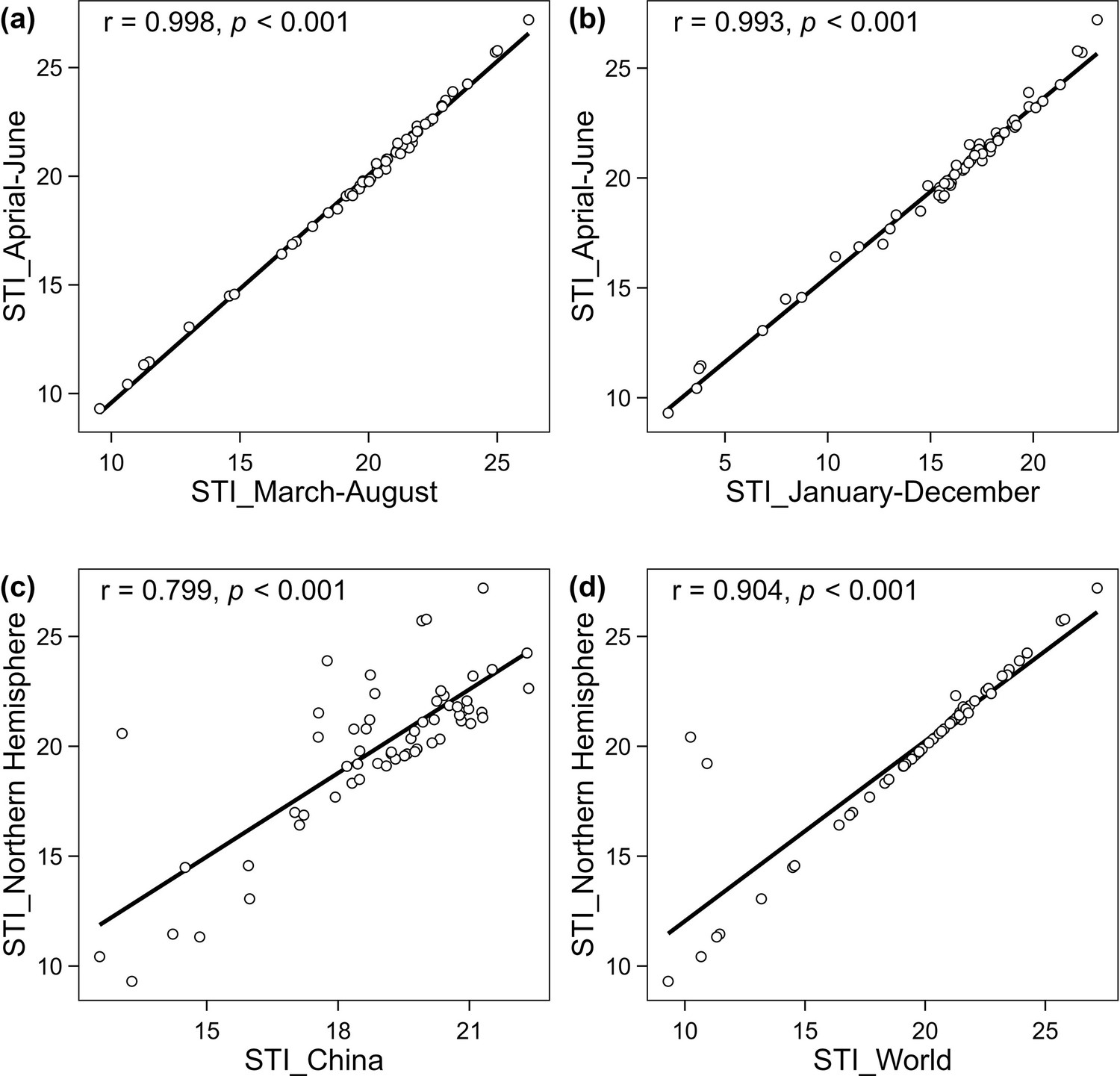
Robustness of 60 species’ STI to changes in data resources.
The units on the horizontal and vertical axes are degrees Celsius. Pearson correlation coefficients between STI used in our study and STI calculated with different time-windows: March to August (a) and January to December (b), were significantly high. Pearson correlation coefficients between STI used in our study and STI calculated with different distributional ranges: the distributional range in China (c) and distributional range across the world (d), were significantly high.
Figure 4 with 2 supplements

Relationship between STI and occupancy trend, abundance trend and the relationship among species thermal preference and trend in occupancy, colonization, and extinction rates.
Each point and error bar in (a) represents the temporal trend of occupancy rate (posterior mean and 95% credible interval of year effect on occupancy rate) for each species, with the filled dots indicating a significant year effect while the hollow dots indicating nonsignificant year effect. The dotted vertical line indicates the median of STI values. Cold-adapted species are plotted in blue and warm-adapted species are plotted in orange. The number of species with significant occupancy trends in each quadrant was added to the plot. The black dashed line and shaded area are the predicted values and 95% confidence intervals of the weighted linear regression model (n = 60). Each point in (b) represents the posterior mean estimate of year in colonization, extinction or occupancy rate for each of 60 species. The color of the point indicates the temporal trend in occupancy. Similar to (a), each point and error bar in (c) represents the temporal trend of abundance (year effect and 95% confidence interval of year effect on abundance) for each species. The black dashed line and shaded area are the predicted values and 95% confidence intervals of the weighted linear regression model (n = 60).
Figure 4—figure supplement 1
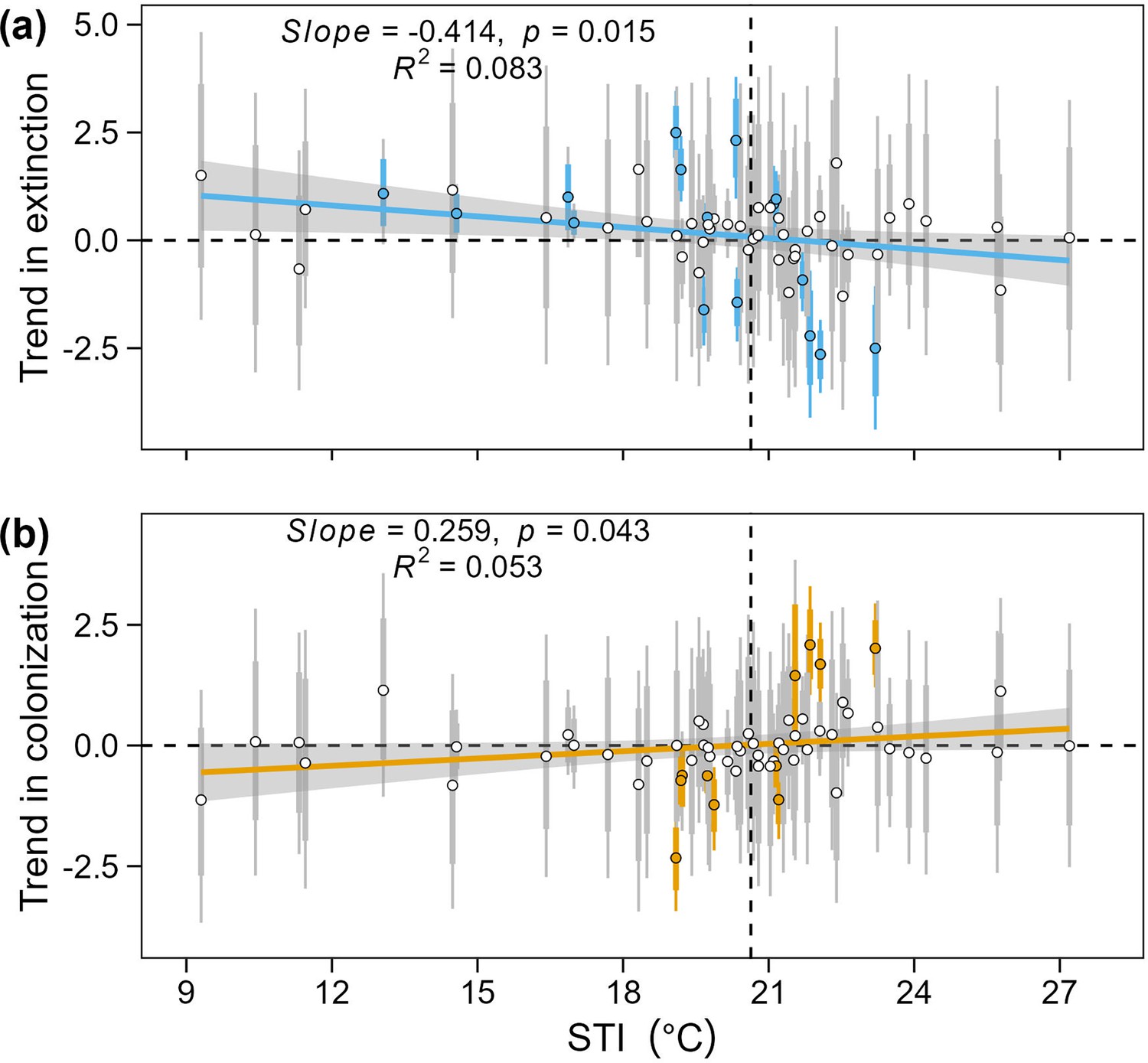
Relationship between STI and extinction trend or colonization trend.
Relationship between STI and mean estimates and 95%, and 80% credible intervals of posterior distribution for temporal trends of species extinction rate (a) and species colonization rate (b). Points and error bars in blue or orange indicate significant posterior estimates and error bars in grey indicate non-significant effects. Results of linear regression models modeling posterior mean of temporal trends in colonization (n = 60) or extinction (n = 60) weighted by standard deviations were added to plots.
Figure 4—figure supplement 2
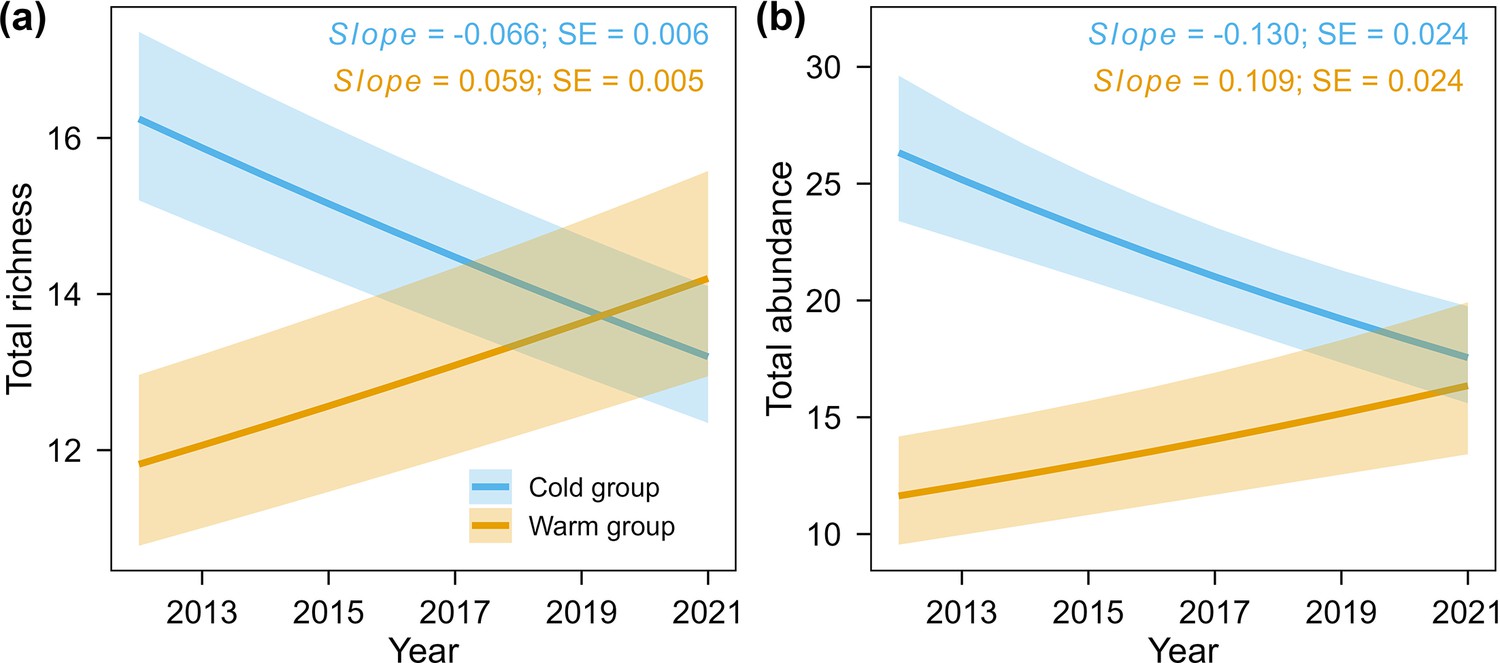
Temporal trend in total richness and abundance for warm-adapted species and cold-adapted species groups, seperately.
Partial regression effects of year effect from LMM on total species richness (a) and total abundance (b) for warm versus cold-adapted species groups. The solid lines indicate significant effects. The shaded bands are 95% confidence intervals of fitted values with other variables at their mean values. Total richness (n = 530) for each of the 53 transects in each year was calculated as the mean of the posterior estimate of species absence/presence, while total abundance (n = 530) for each of the 53 transects in each year was calculated from raw maximum abundance data.
Figure 5 with 1 supplement

Posterior estimates of logit-scale parameters related to cold-adapted species’ extinction rates and warm-adapted species’ colonization rates.
Points are species-specific posterior means on the logit-scale extracted from MSOM, where parameters >0 indicate positive effects (on extinction [a] or colonization [b]) and parameters <0 indicate negative effects. Only cold-adapted species (23 selected species) with positive trends in extinction (a) and warm-adapted species (15 selected species) with positive trends in colonization (b) are shown, which are two main processes contributing positively to thermophilization. Points in blue/orange indicate significant effects (95% or 80% credible intervals) and points in white indicate non-significant effects. All points were jittered slightly by 0.22 in width.
Figure 5—figure supplement 1
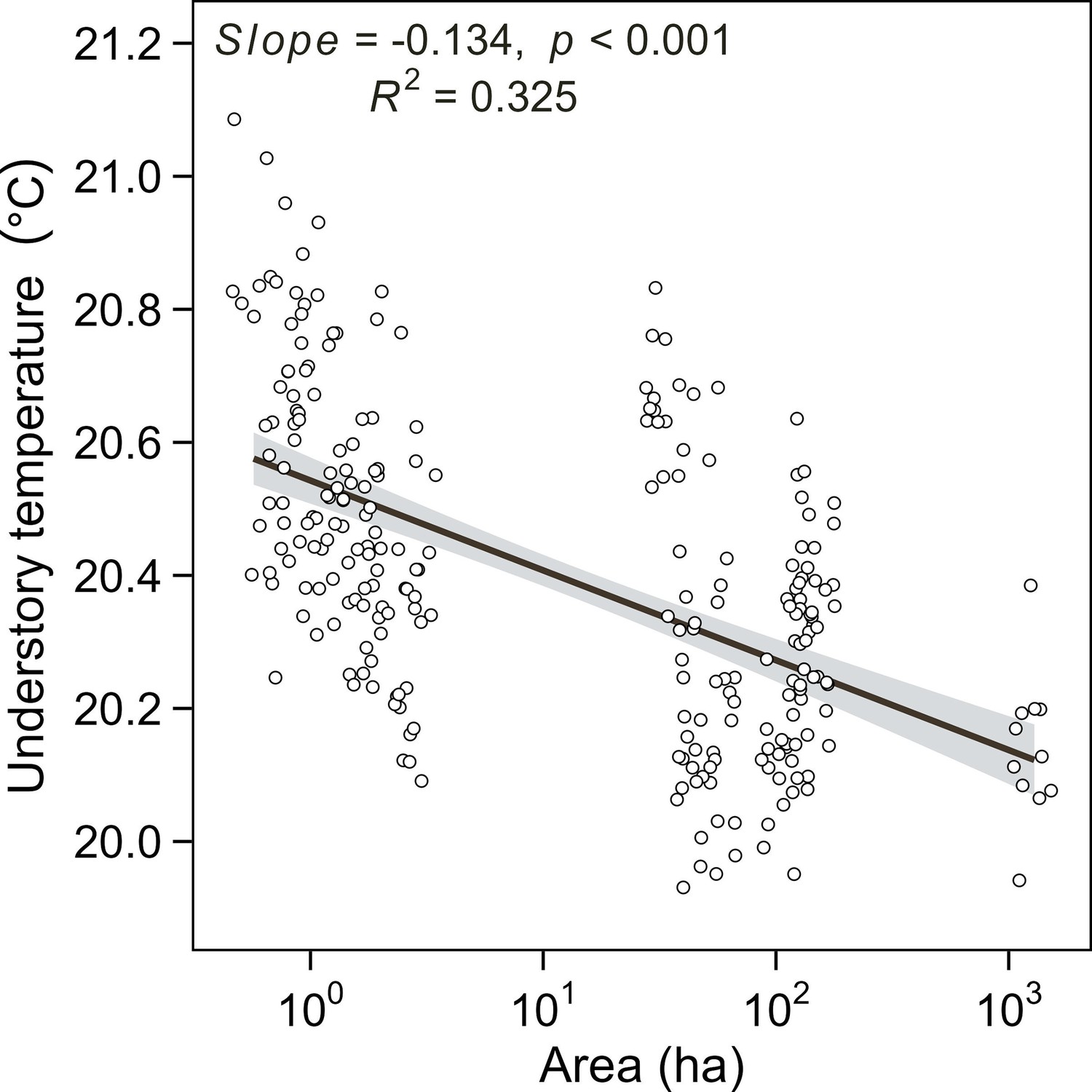
Relationship between island area and understory breeding season air temperature of 20 islands monitored in the year 2022.
The understory air temperature was monitored in 20 selected study islands differing in island area. Each plot represents the average air temperature (April – June) of a site within the islands. Lines and shaded areas are predicted values and 95% confidence intervals from linear regression models.
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Habitat fragmentation mediates the mechanisms underlying long-term climate-driven thermophilization in birds
eLife 13:RP98056.
https://doi.org/10.7554/eLife.98056.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}