Temporal transcriptional response to ethylene gas drives growth hormone cross-regulation in Arabidopsis
- The Salk Institute for Biological Studies, United States
- Carnegie Mellon University, United States
- University of Toronto, Canada
- University of California, San Diego, United States
- Howard Hughes Medical Institute, The Salk Institute for Biological Studies, United States
- University of California, San Diego School of Medicine, United States
- School of Computer Science, Carnegie Mellon University, United States
Figures
Figure 1 with 5 supplements
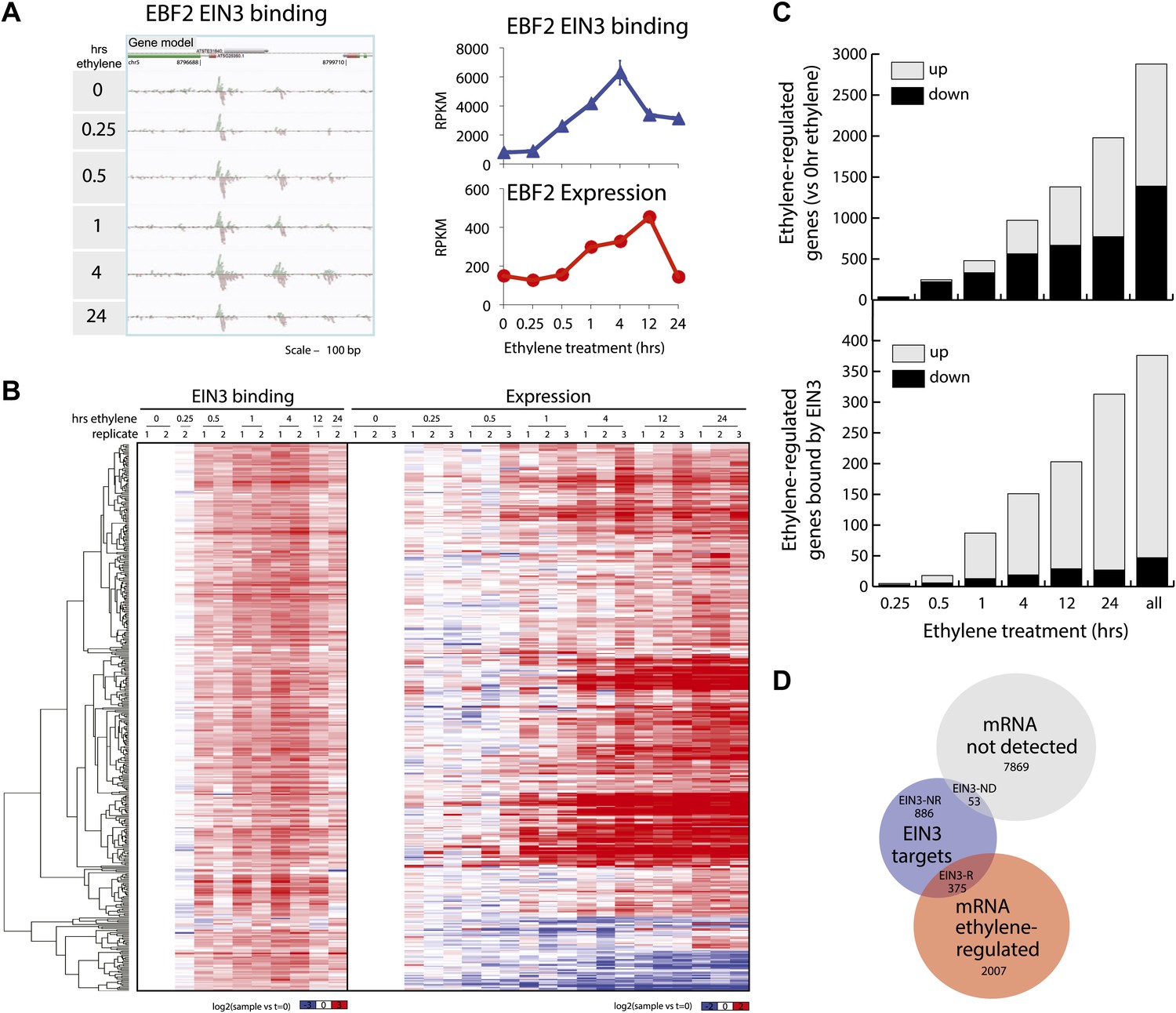
Dynamics of ethylene-induced EIN3 binding and transcription supports the role of EIN3 as an activator of the ethylene response.
(A) Ethylene treatment results in an increase of EIN3 binding in three regions of the EBF2 promoter, corresponding to an increase in steady-state mRNA levels. Binding and transcription levels are indicated by reads per kilobase per million reads in sample (RPKM). Gene model: green (exon), red (UTR), grey (intron/transposon). (B) Patterns of EIN3 binding and expression of ethylene-regulated targets are strikingly evident over a timecourse of ethylene gas treatment. EIN3 binding increases with ethylene treatment to a maximum at 4 hr of ethylene treatment for all candidate targets. Each line in the heatmap represents the RPKM value for the representative EIN3 binding site (left panel) and transcript (right panel). (C) (Upper panel) Equivalent numbers of genes are up- and down-regulated upon ethylene treatment. (Lower panel) Majority of EIN3 targets differentially expressed upon ethylene treatment are up-regulated. (D) A subset of EIN3 targets is transcriptionally regulated by ethylene (EIN3-R).
Figure 1—figure supplement 1
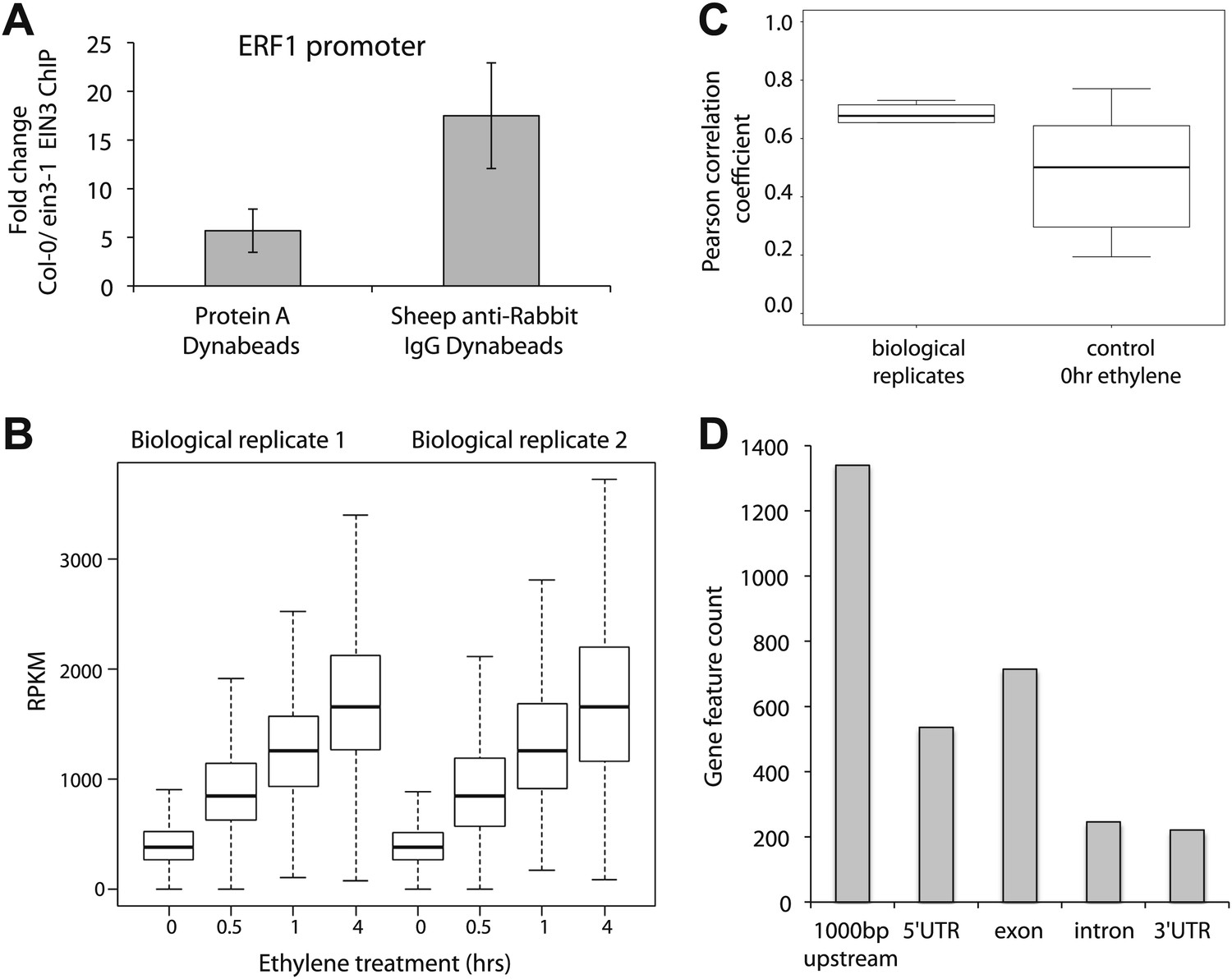
EIN3 antibody reproducibly enriches DNA in chromatin immunoprecipitation.
(A) Enrichment of the known target of EIN3, the promoter of ERF1, using Dynabeads Protein A and Dynabeads Sheep anti-Rabbit IgG to collect protein-DNA complexes. The average fold change for two technical ChIP replicates with three QPCR technical replicates each is shown. (B) Reproducibility in the two biological replicates for EIN3 ChIP performed upon treatment of ethylene gas for 0, 0.5, 1, and 4 hr. (C) Average RPKM of EIN3 binding sites 0, 0.5, 1, and 4 hr of ethylene gas treatment. (D) EIN3 binding preferentially occurs in the promoter regions of genes (1000 bp upstream of the TSS).
Figure 1—figure supplement 2
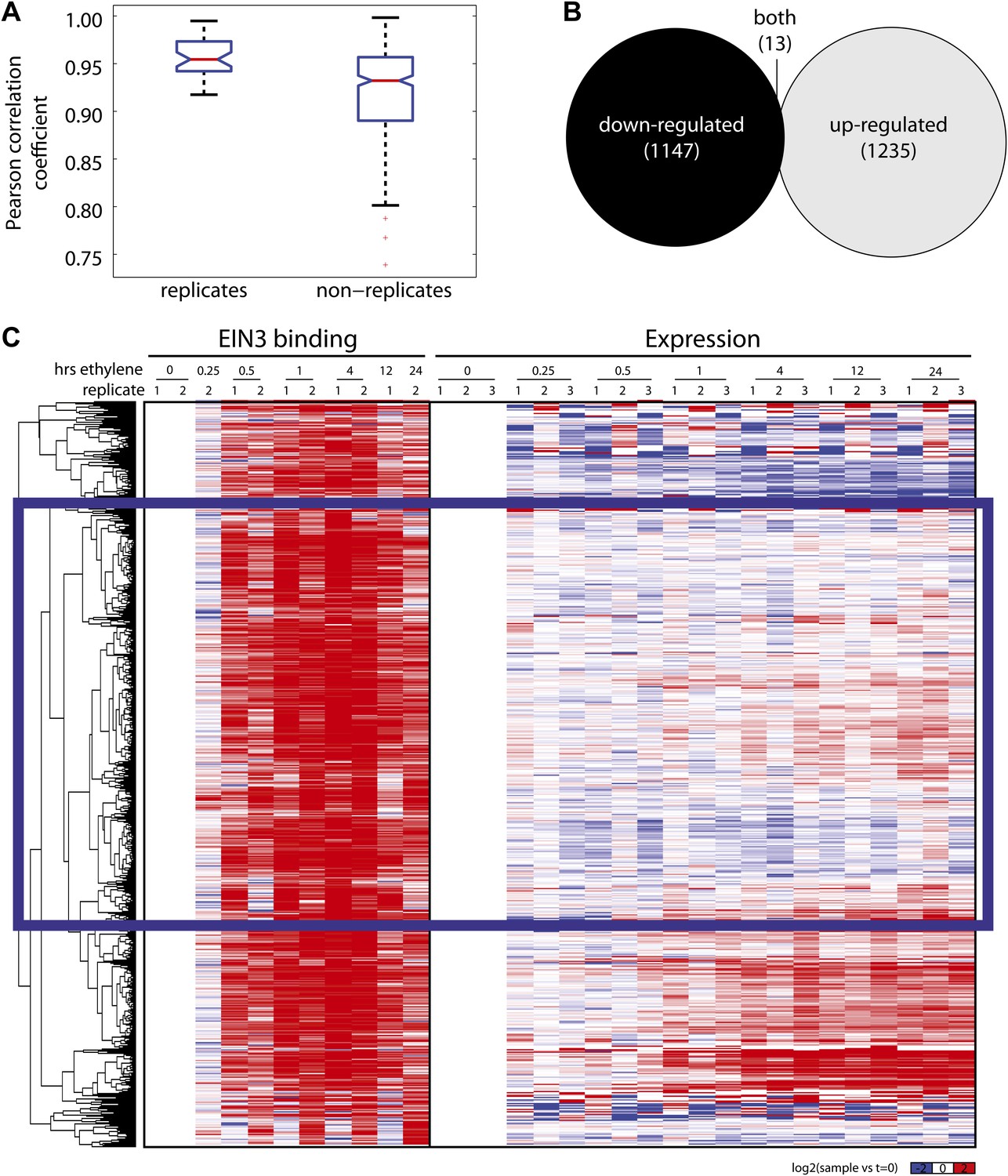
Ethylene-regulated genes are induced and repressed.
(A) Reproducibility of RNA-Seq experiments. Ethylene-regulated have a higher reproducibility amongst replicates than non-replicates. (B) Genes that are both up- and down-regulated occur at different timepoints of ethylene treatment. (C) The majority of EIN3 targets exhibit increased binding upon ethylene treatment, however, changes in steady-state levels of mRNA do not occur for the majority of these targets. EIN3-NR genes are indicated by the blue rectangle.
Figure 1—figure supplement 3
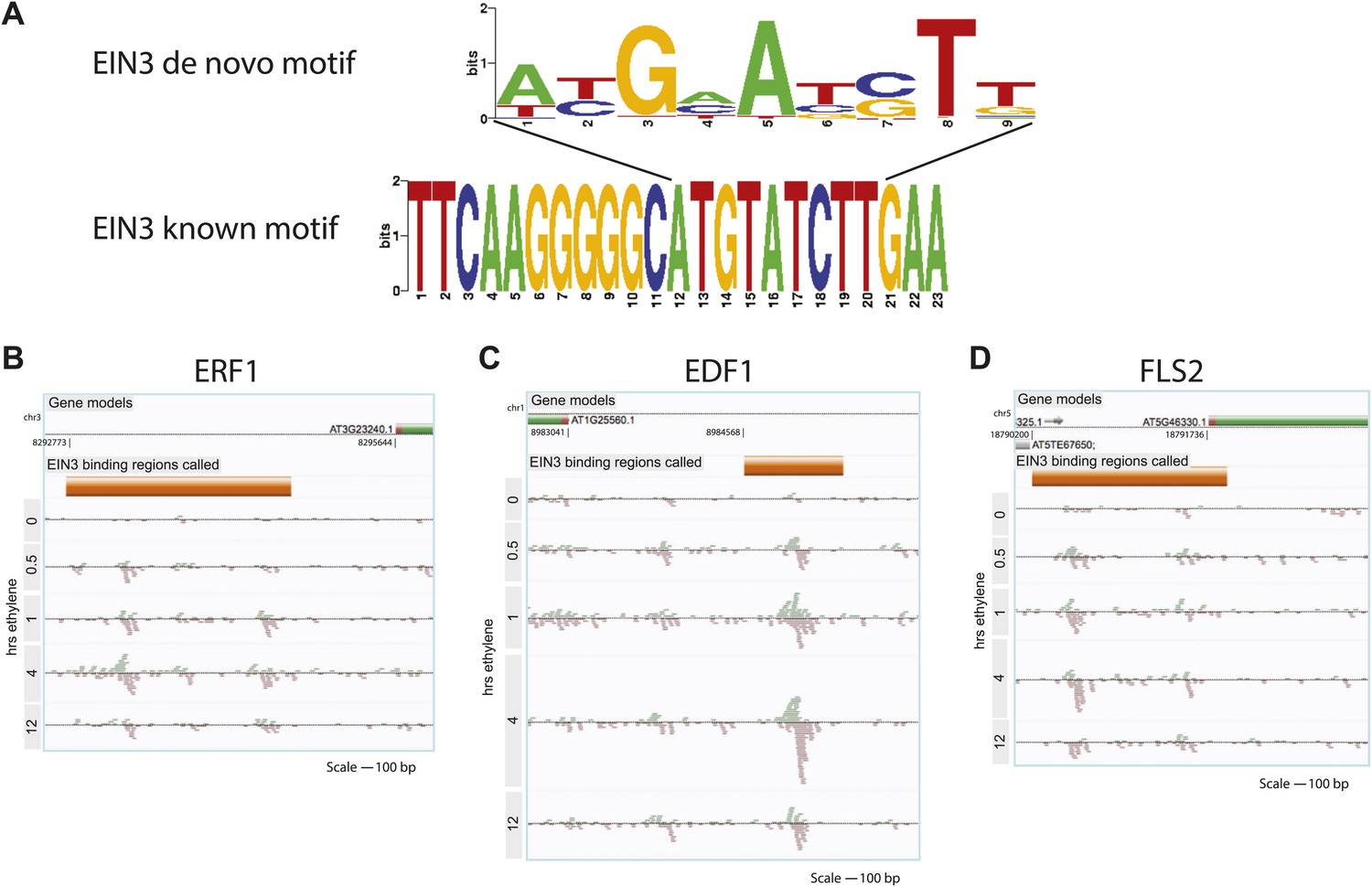
Binding of EIN3 to previously known targets.
(A) De novo motif from the top 50 EIN3 binding sites with the best match to the known EIN3 motif (E-value = 1.12 × 10−5). EIN3 binding of the promoters of (B) ERF1, (C) EDF1, and (D) FLS2.
Figure 1—figure supplement 4
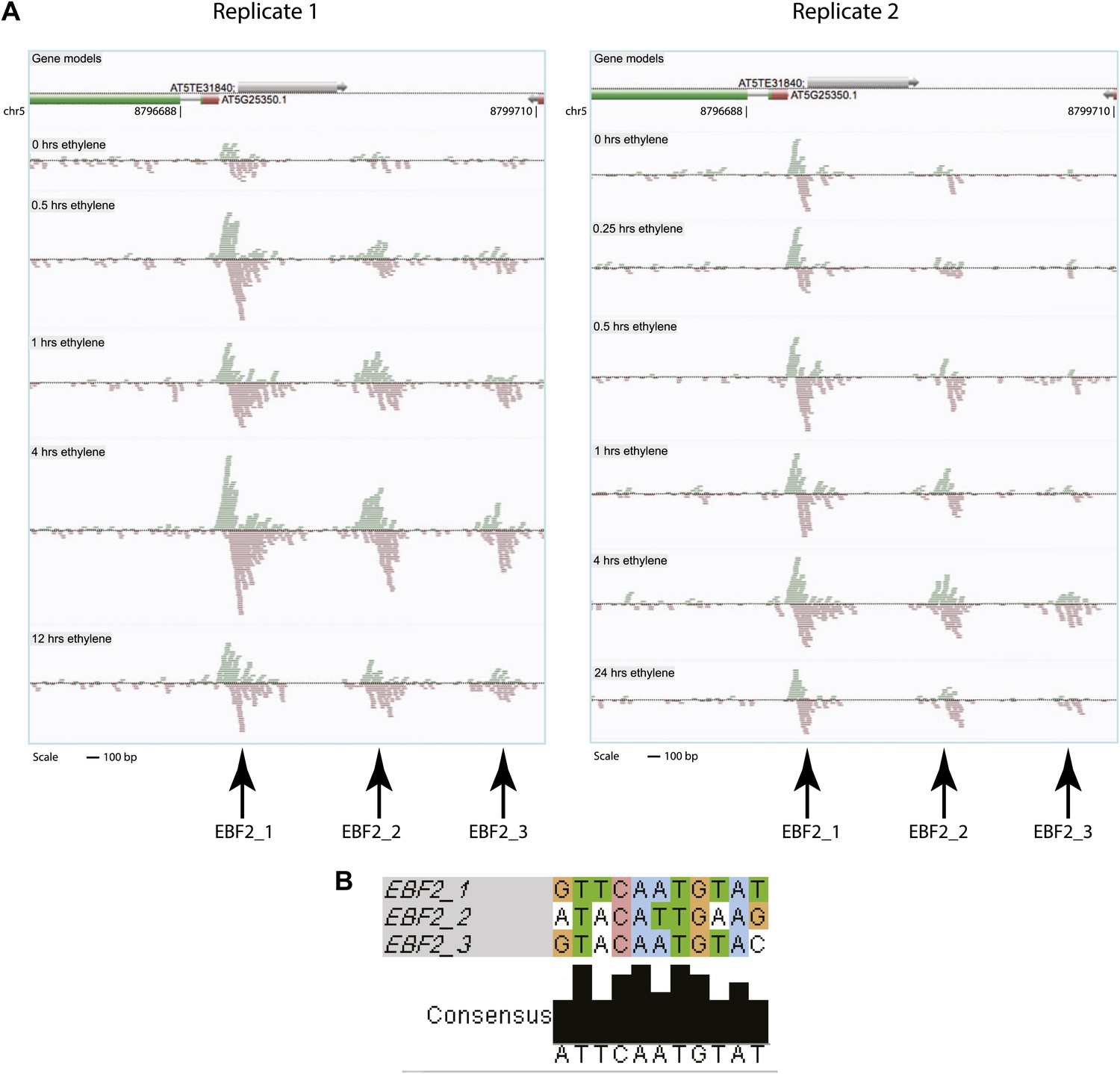
EIN3 ChIP-Seq identified an additional binding in the EBF2 promoter.
(A) Binding of EIN3 to the EBF2 promoter increases upon ethylene gas treatment. EIN3 binding is strongest in the most proximal site to the TSS, and weakest in the most distal site to the TSS, which is not called by our stringent analysis. The most distal EIN3 binding site was not detected by our analysis but displays the characteristic ChIP-Seq binding pattern. (B) Alignment of motifs of the three binding sites in the EBF2 promoter.
Figure 1—figure supplement 5
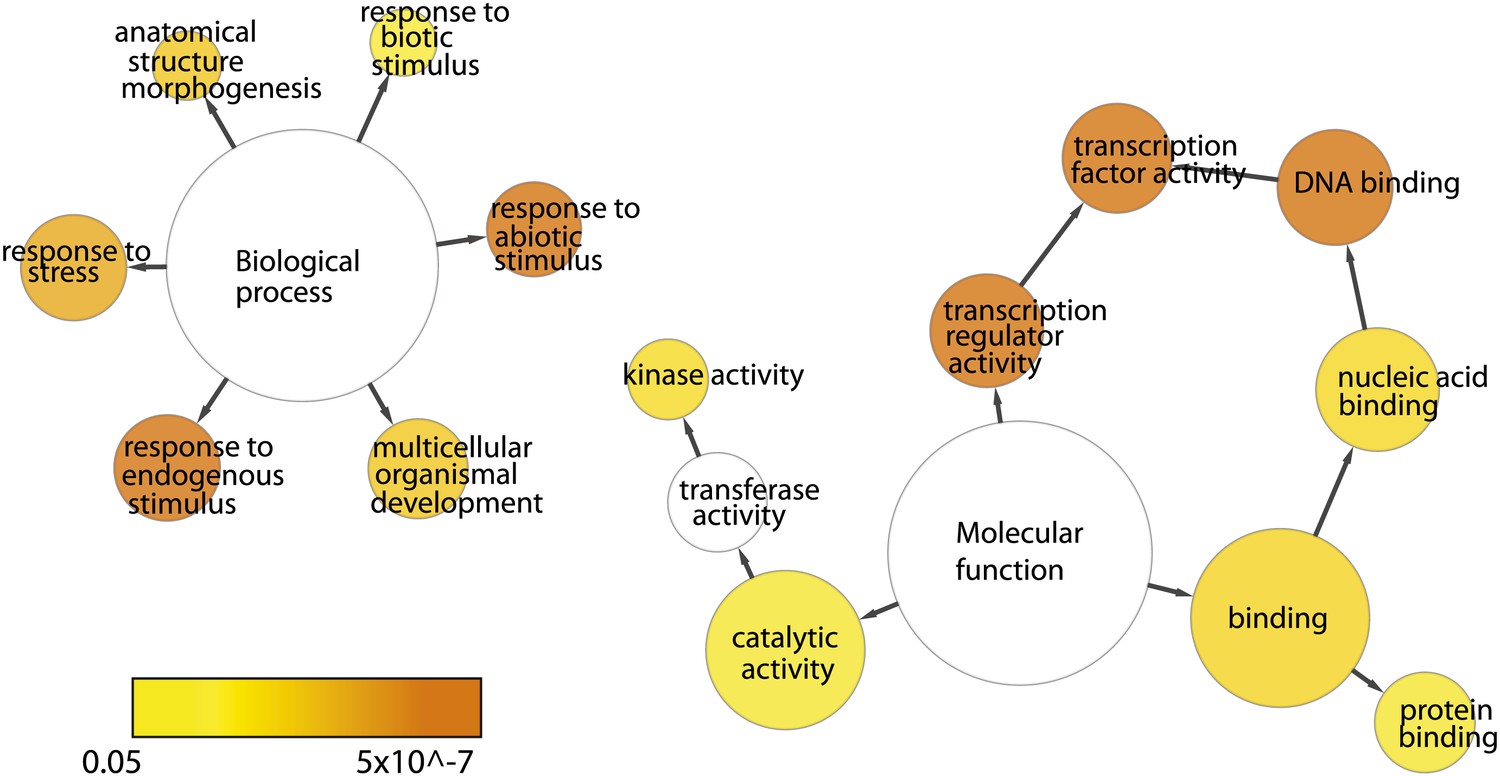
Functional categories are over-represented for EIN3 targets that are ethylene-regulated (EIN3-R).
Network was generated using BiNGO (v.2.44) using the GOSlim_Plants ontology, Benjamini and Hochberg p-value legend is indicated below.
Figure 2 with 3 supplements

The ethylene transcriptional response occurs in four distinct waves of transcriptional induction.
(A) Ethylene-regulated EIN3 targets (EIN3-R) exhibit increased binding at transcription start sites (TSS) upon ethylene treatment (black arrows) in comparison to those not transcriptionally regulated by ethylene (EIN3-NR and EIN3-ND). Each boxplot represents the distribution of EIN3 ChIP-Seq RPKMs near the TSS. (B) Distribution of gene families among EIN3-R targets reveals over-representation of gene families related to hormone responses function. (Inset) Percentage of hormone-related genes in EIN3 binding and transcription categories. (C) DREM paths representing waves of induction of steady-state levels of transcription by ethylene for genes that are regulated by EIN3, implicating different modes of transcriptional regulation in the ethylene response. Right panels contain all genes for each wave.
Figure 2—figure supplement 1
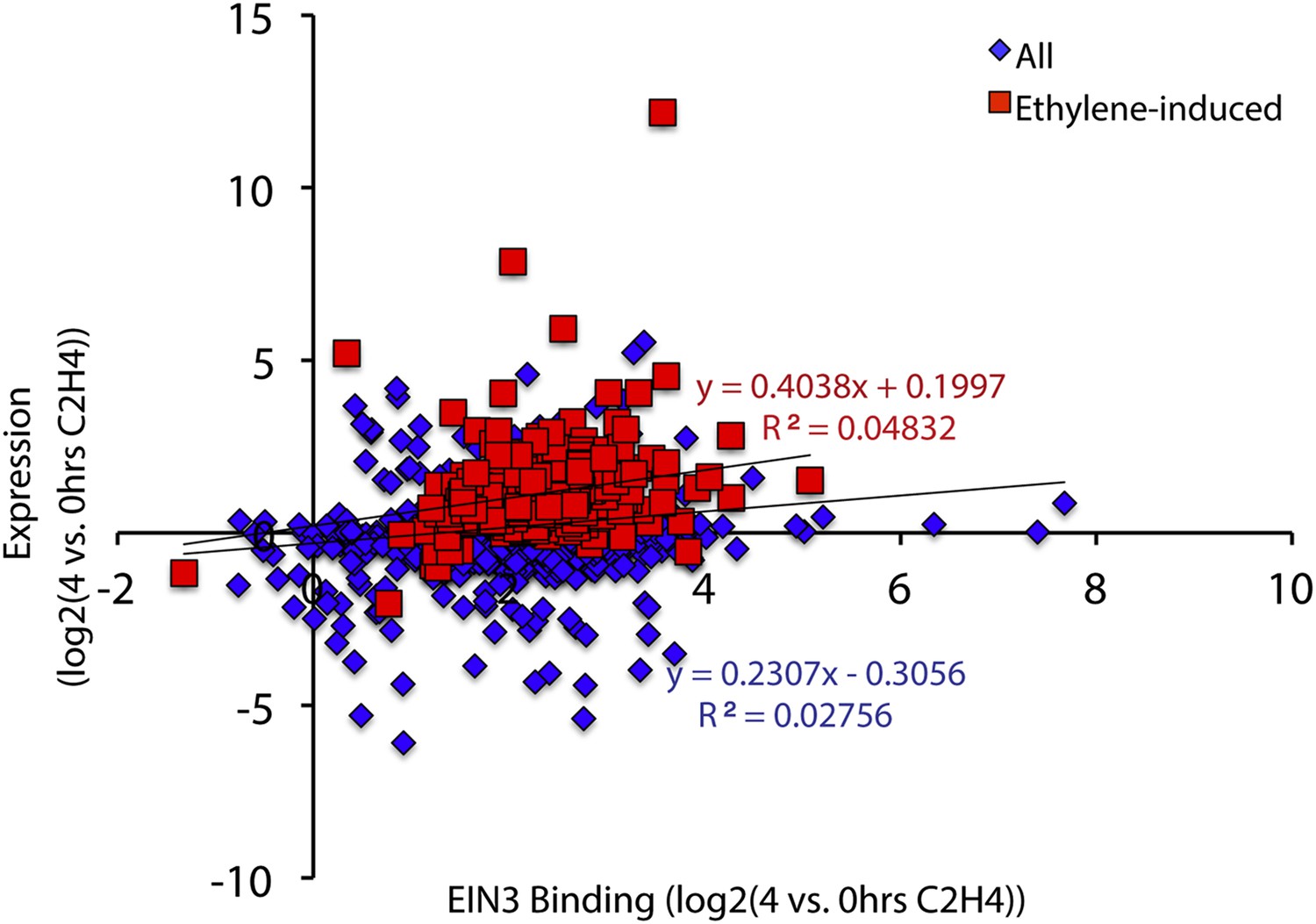
Quantitative correlation between EIN3 binding and ethylene-regulated expression.
Linear regression revealed that there is no quantitative correlation of changes in EIN3 binding and ethylene-regulated steady-state levels of transcription.
Figure 2—figure supplement 2
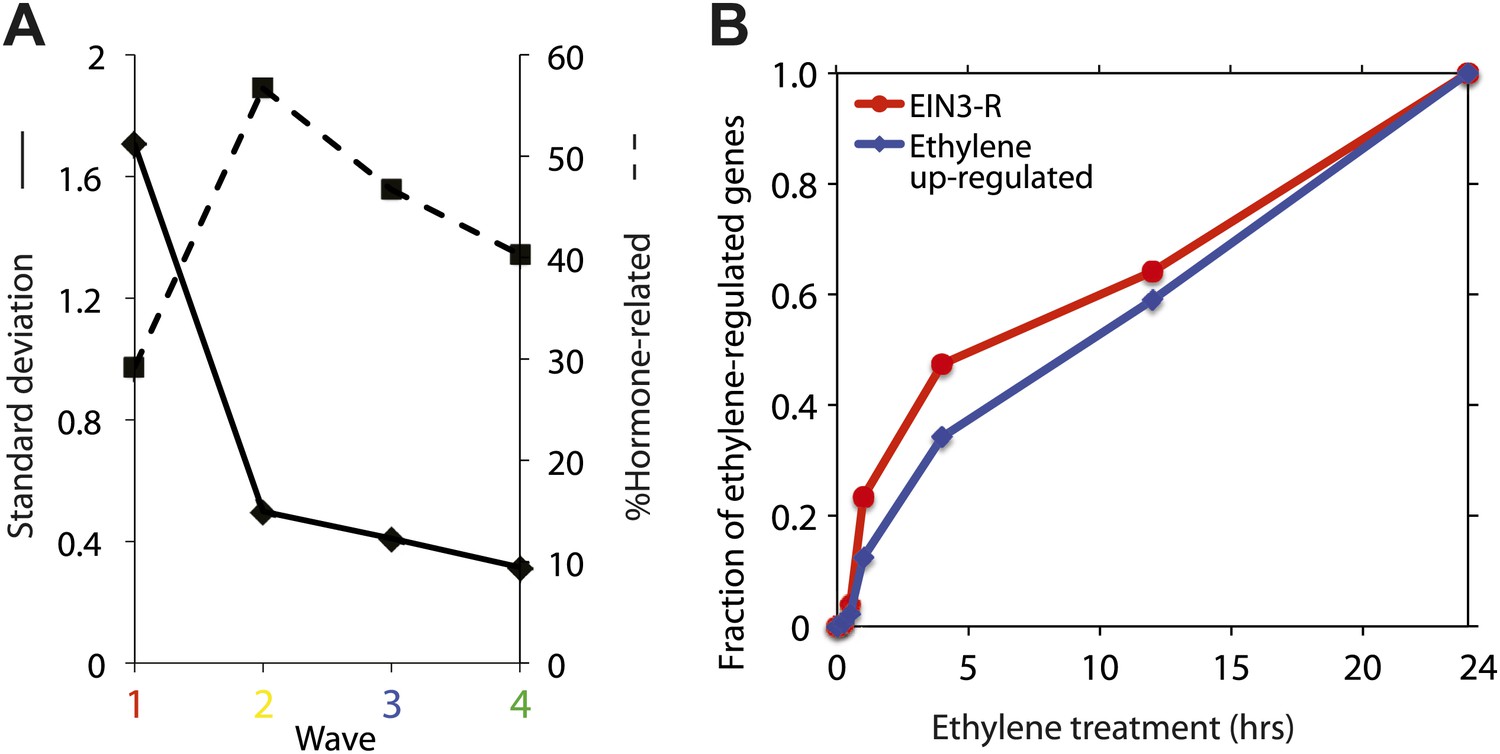
Temporal characterization of the ethylene transcriptional response.
(A) The EIN3-modulated ethylene transcriptional response occurs in four waves with various levels of noise. A decrease in standard deviation correlates to an increase of hormone-related genes. (B) The kinetics of the ethylene transcriptional response on a linear scale. Approximately 35–50% of the ethylene-regulated genes are transcriptionally affected by the first 4 hr of ethylene treatment, for all ethylene transcriptional response genes and EIN3-modulated ethylene transcriptional response genes, respectively.
Figure 2—figure supplement 3
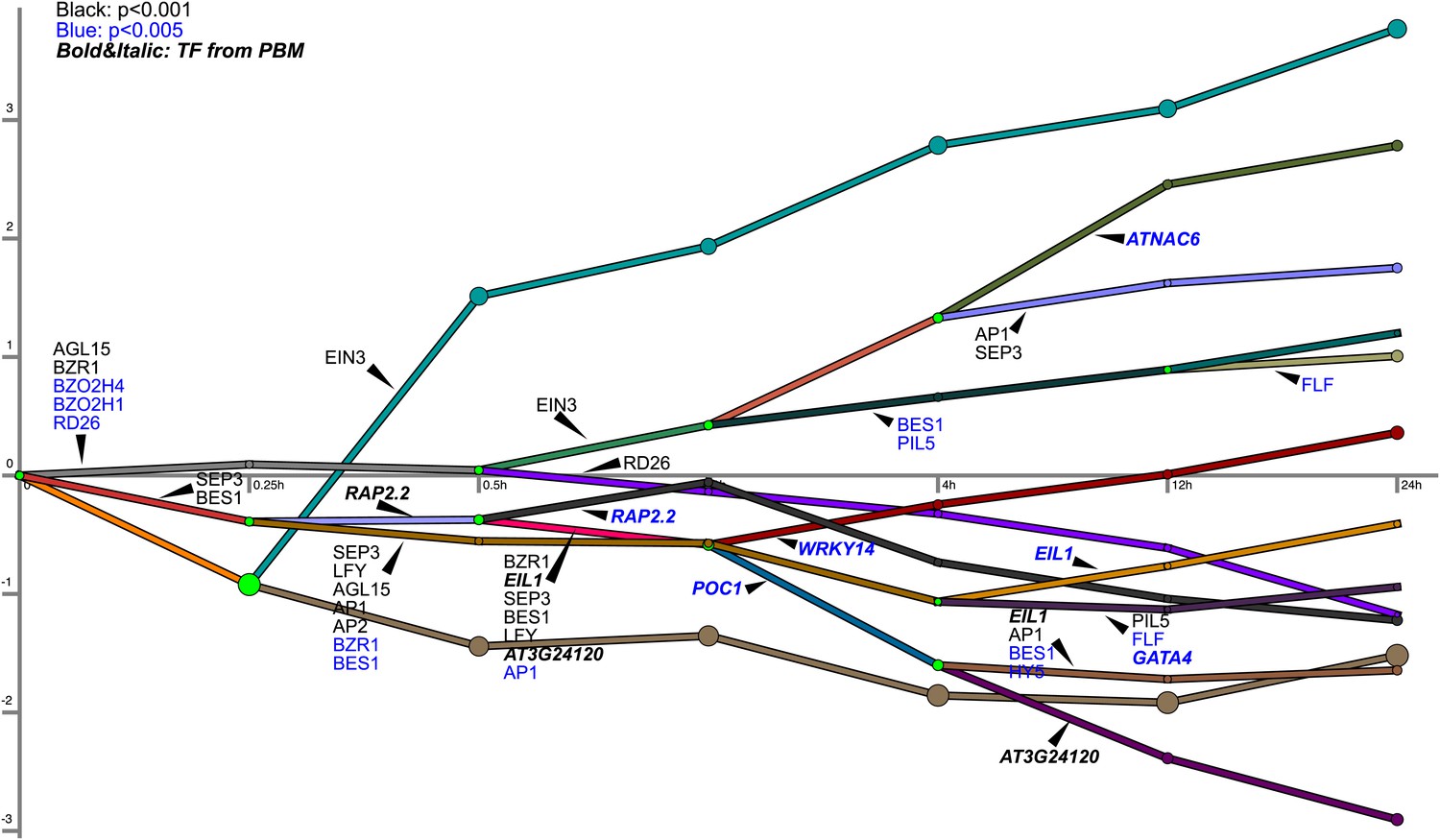
Ethylene transcription and associated transcription factor regulation kinetics from DREM analysis.
This dynamic regulatory map contains major bifurcation or convergence events in temporal expression data and determines the transcription factors potentially responsible for these events. EIN3-modulation is significant (10−10) in the expression trajectories labeled EIN3 (teal, dark brown, purple, green-blue; first four endpoint trajectories from the top of the figure). The EIN3-associated trajectories represent the four waves of ethylene-regulated expression regulated by EIN3, as further discussed in the main text. The TF associations are based on motifs from AGRIS and the PBM data in this paper. There are two caveats based on the TF association annotation: (1) It is possible the TF controlling these RNA is not represented in the database, and (2) Genes may not be regulated by the specific TF indicated, but instead by a homolog or other TF that has a similar target sequence not in the database.
Figure 3 with 1 supplement

Functional classification of EIN3 candidate targets reveals genes involved in hormone responses.
(A) Feedback (ethylene signaling components, above) of the ethylene response and feedforward (downstream effectors, below). Downstream effectors in green are transcriptionally induced by ethylene. Known EIN3 targets are noted by asterisks; all other EIN3 candidate targets were discovered by this study. (B and C) EIN3 candidate targets are involved in hormone co-regulation. Node color represents hormone annotation, as indicated in B; large nodes are EIN3 candidate targets. Dark grey edges represent protein-protein interactions (PPI) and light grey edges are protein–DNA interactions (PDI). Hormone annotation legend: abscisic acid (ABA), brassinosteroid (BR), cytokinin (CK), ethylene (ETH), gibberellin (GA), auxin (IAA), methyl jasmonate (MJ), salicylic acid (SA), >1, more than one hormone. (D) EIN3-mediated ethylene co-regulation occurs at many different levels. PPIs are from the Arabidopsis Interactome Mapping Consortium, and EIN3 PDIs are from this study.
Figure 3—figure supplement 1

Motifs of EIN3 targets that are transcriptionally regulated by ethylene were determined in vitro using protein binding microarrays.
https://doi.org/10.7554/eLife.00675.014
Figure 4 with 4 supplements
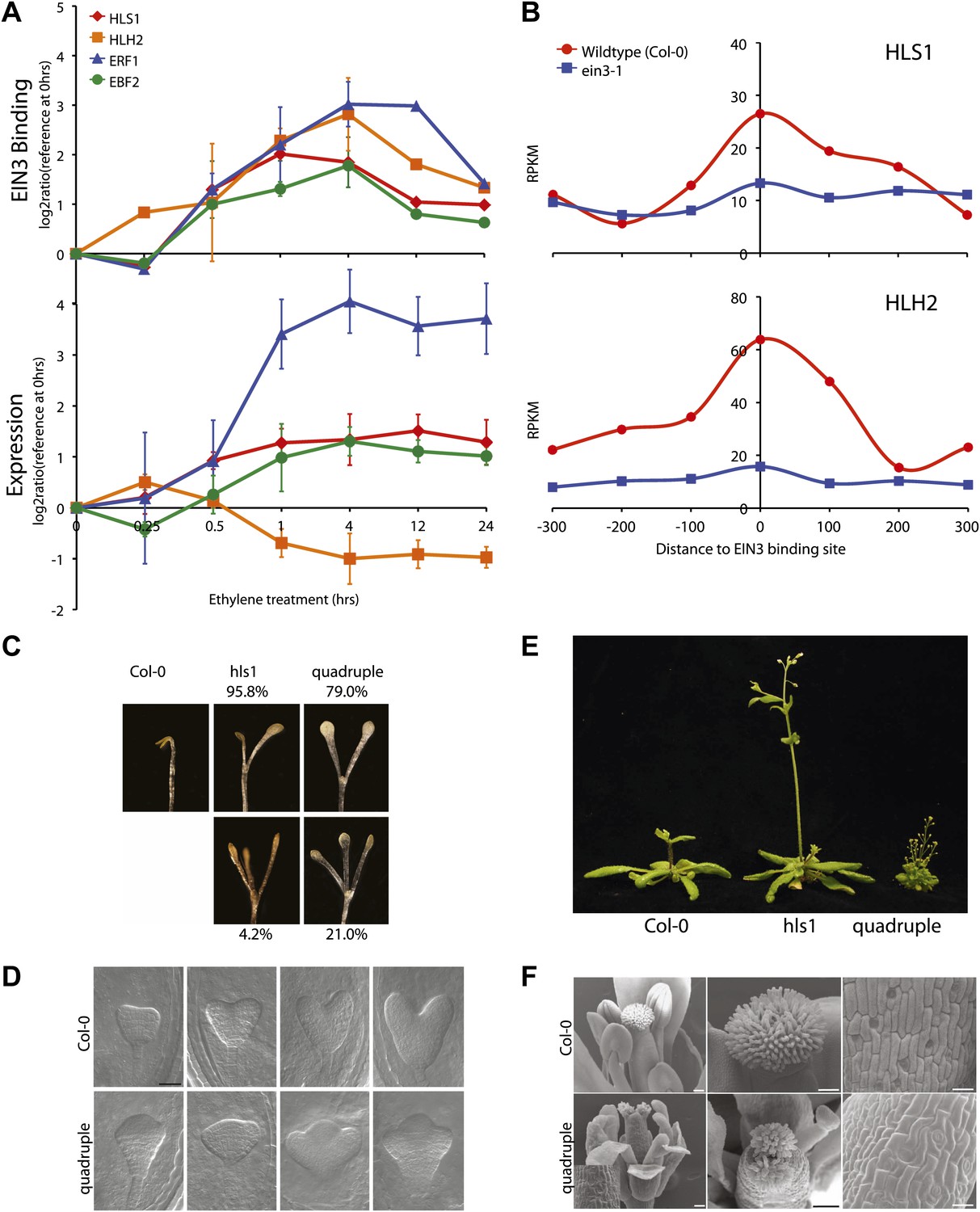
EIN3 binding facilitates HLS1 ethylene-auxin hormone co-regulation.
(A) (Top panel) EIN3 targets HLS1 and HLH2. Temporal EIN3 binding and expression patterns are shown with known EIN3 targets as a control. HLH1 and HLH3 are not expressed in etiolated seedlings. (B) Binding of EIN3 to HLS1/HLH2 promoters is dependent on presence of EIN3. (C)–(F) Mutations in HLS1 and its homologs reveal severe growth and developmental defects. (C) Tri-cotyledon phenotypes in apical hook of quadruple mutants. Images were taken at the same magnification. (D) HLS1 gene family has a role in embryo patterning. SEM image scale bar, 50 μm. (E) Adult three-week-old plants displayed dwarfed phenotypes similar to axr1. (F) Quadruple mutants display floral defects similar to arf3/ettin. Inset and panels on the right show abnormal guard cell patterning. SEM scale bars, 100 μm.
Figure 4—figure supplement 1
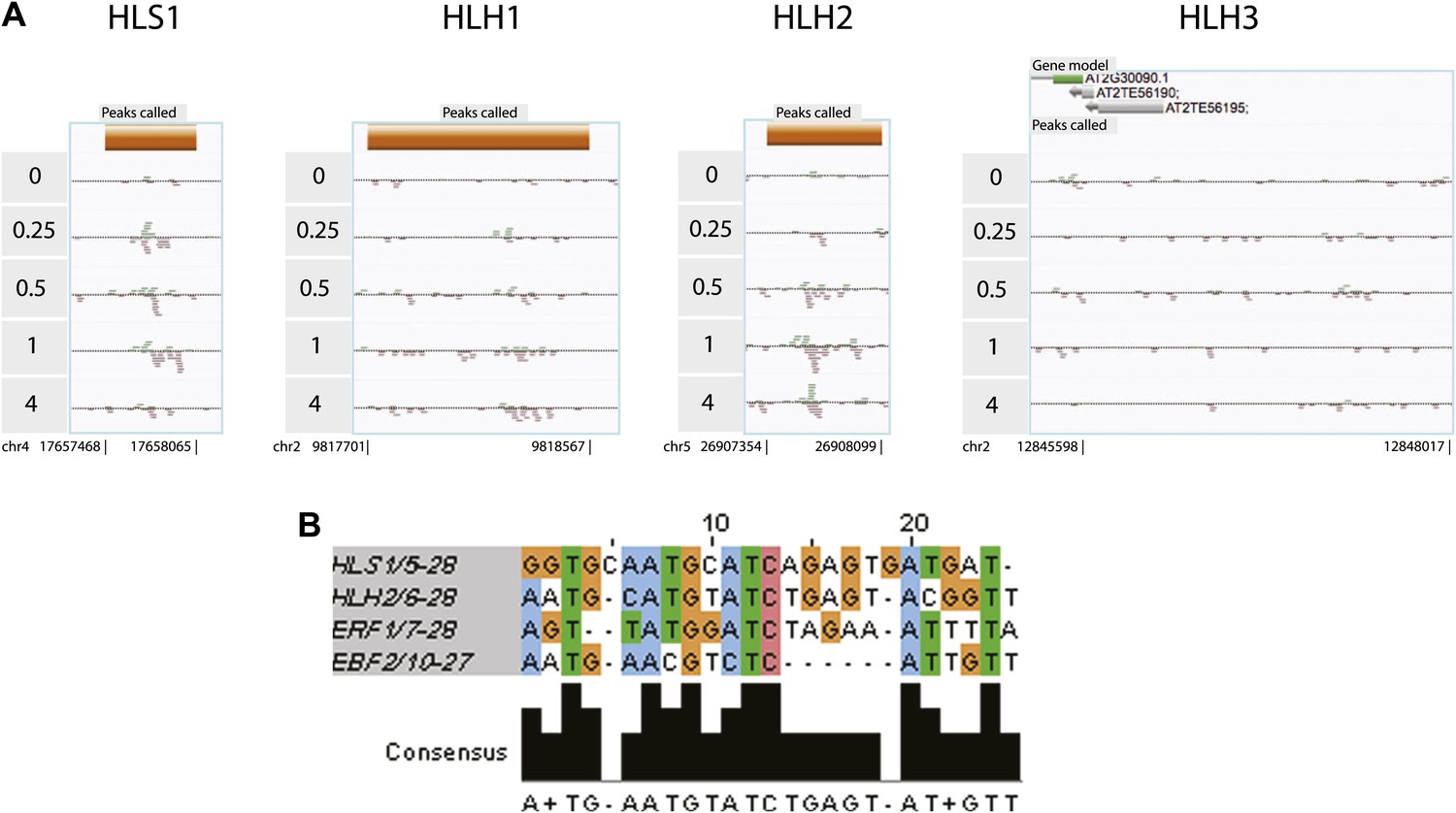
HLS1, HLH1, and HLH2 are targets of EIN3.
(A) EIN3 binding in HLS1, HLH1, HLH2 promoters. (B) EIN3 binding motifs in HLS1, HLH2 reveal a consensus. ERF1 and EBF2 motifs are shown as a reference.
Figure 4—figure supplement 2
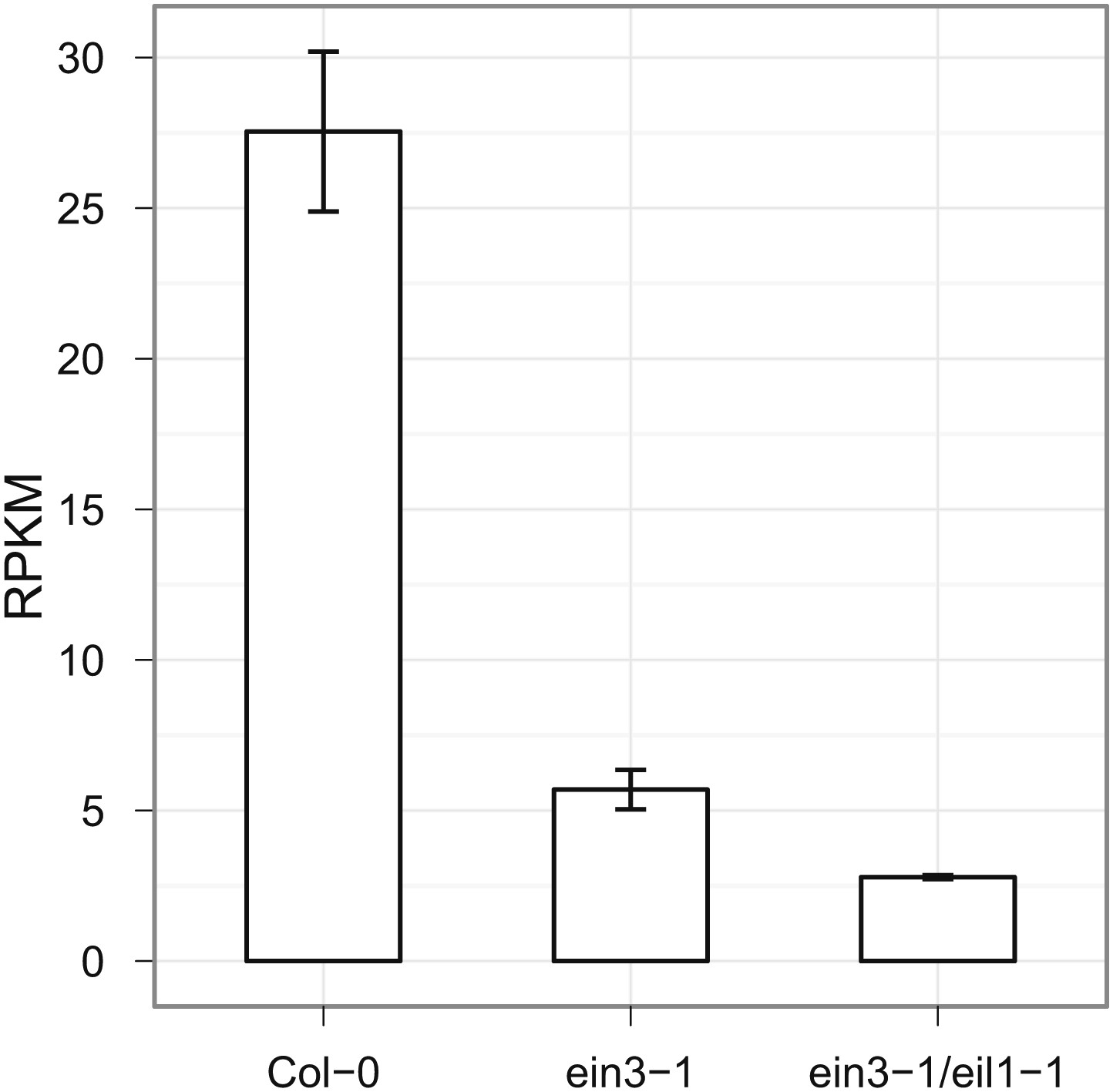
HLS1 expression is decreased in ein3-1, and ein3-1/eil1-1 mutants.
https://doi.org/10.7554/eLife.00675.017
Figure 4—figure supplement 3

HLS1-like homologs (HLHs) are similar to HLS1 in protein sequence and domain structure.
(A) Conservation of HLS1 and HLHs proteins. Amino acid sequence alignment of HLS1 and its three homologs are shown. Gaps are represented as ‘.’. Shading indicates identical sequences (black), conserved changes (gray), similar residues (light gray). (B) Phylogeny of HLS1 and HLHs and proteins from other organisms containing acetyltransferase domains. Amino acid sequences were aligned using Clustal, then a bootstrap 50% majority-rule consensus tree was constructed using PAUP. Abbreviations for species are as follows: Hs, Homo sapiens; Sc, Saccharomyces ceravisiae; Mm, Mus musculus; At, Arabidopsis thaliana; Bn, Brassica napus; Zm, Zea mays; Os, Oryza sativa; Ps, Pisum sativum; Ec, E. coli. (C) Location of T-DNA insertions in HLH genes. Boxes represent the exons of each HLH gene. Triangles represent the T-DNA alleles that are characterized in detail. Not all T-DNA insertion alleles in the HLH genes are shown.
Figure 4—figure supplement 4
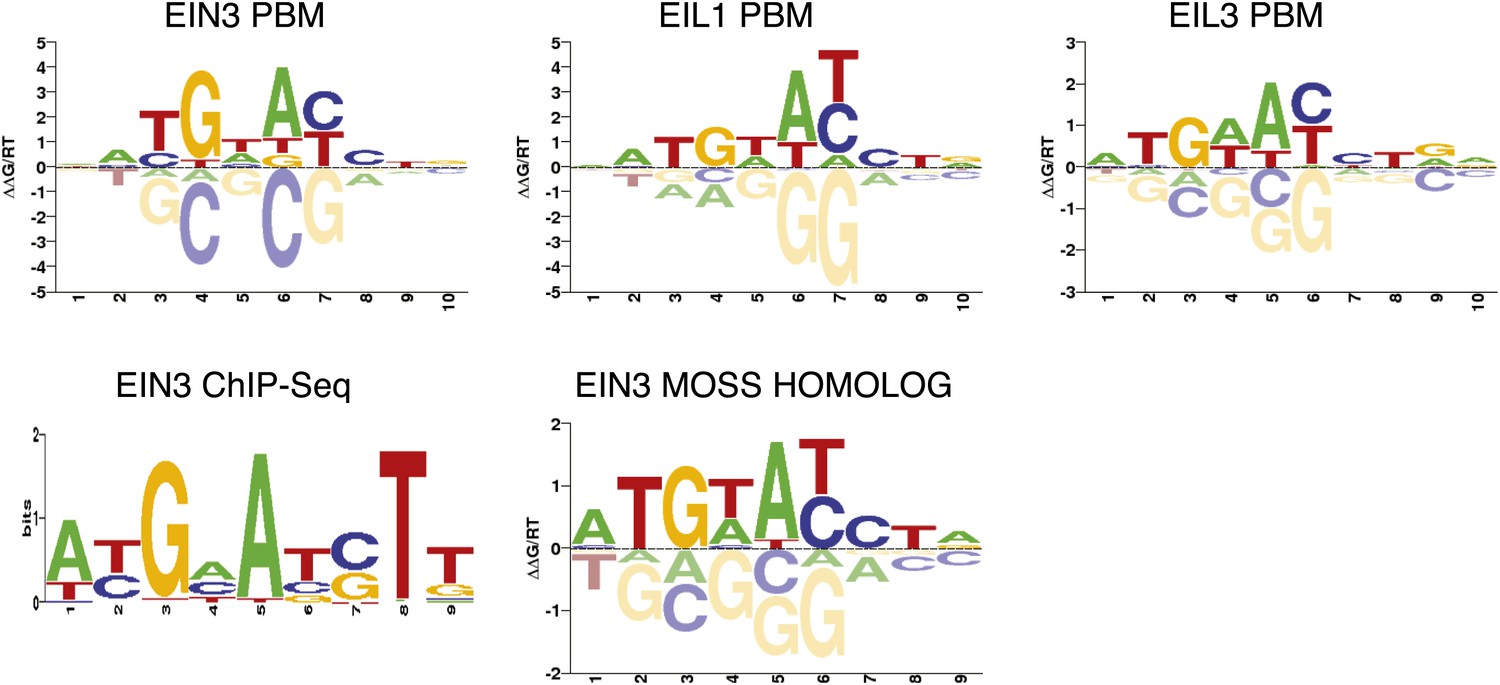
Arabidopsis thaliana EIN3, EIL1, EIL3, and Physcomitrella patens EIN3 DNA-binding motifs from protein binding microarray experiments.
EIN3 ChIP-Seq DNA binding motif is shown for comparison. EIN3 ChIPSeq and protein binding motif alignment was performed with STAMP (E-value = 1.59 × 10−6).
Additional files
-
Supplementary file 1
(A) Summary of sequencing reads from EIN3 ChIP-Seq and mRNA-Seq experiments. (B) EIN3 candidate targets. (C) EIN3 candidate target gene distribution of gene ontology terms. (D) Protein binding microarray transcription factor information. (E) Motifs of EIN3 targets are over-represented in the promoters of ethylene transcriptional response genes.
- https://doi.org/10.7554/eLife.00675.020
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Temporal transcriptional response to ethylene gas drives growth hormone cross-regulation in Arabidopsis
eLife 2:e00675.
https://doi.org/10.7554/eLife.00675




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}