Subdued, a TMEM16 family Ca2+-activated Cl− channel in Drosophila melanogaster with an unexpected role in host defense
- University of California, San Francisco, United States
- Howard Hughes Medical Institute, University of California, San Francisco, United States
Figures
Figure 1
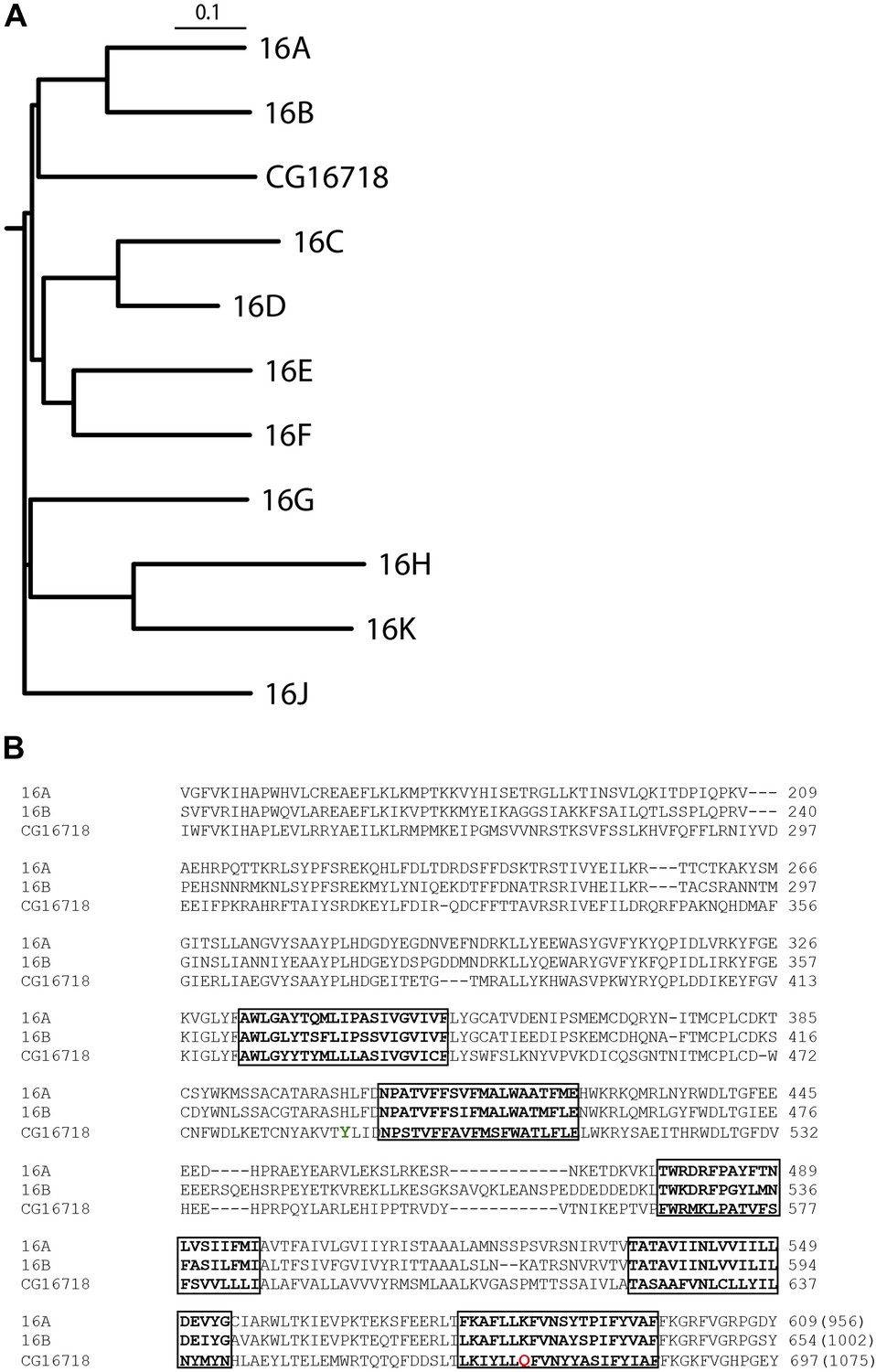
CG16718 (Subdued) aligns closely with mammalian CaCCs TMEM16A and B (Mus musculus).
(A) Multiple sequence alignment of protein sequences was done with ClustalW2 and phylogenetic tree construction was done in PHYLIP 3.67 (Drawgram). (B) Putative transmembrane segments are highlighted with boxes, and mutated residues (Figure 4) featured in this report are marked in color on the primary sequence alignment Y489 is shown in green and Q672 is shown in red. Bracketed values at the end of the alignment indicate the number of residues in the whole channel.
Figure 2
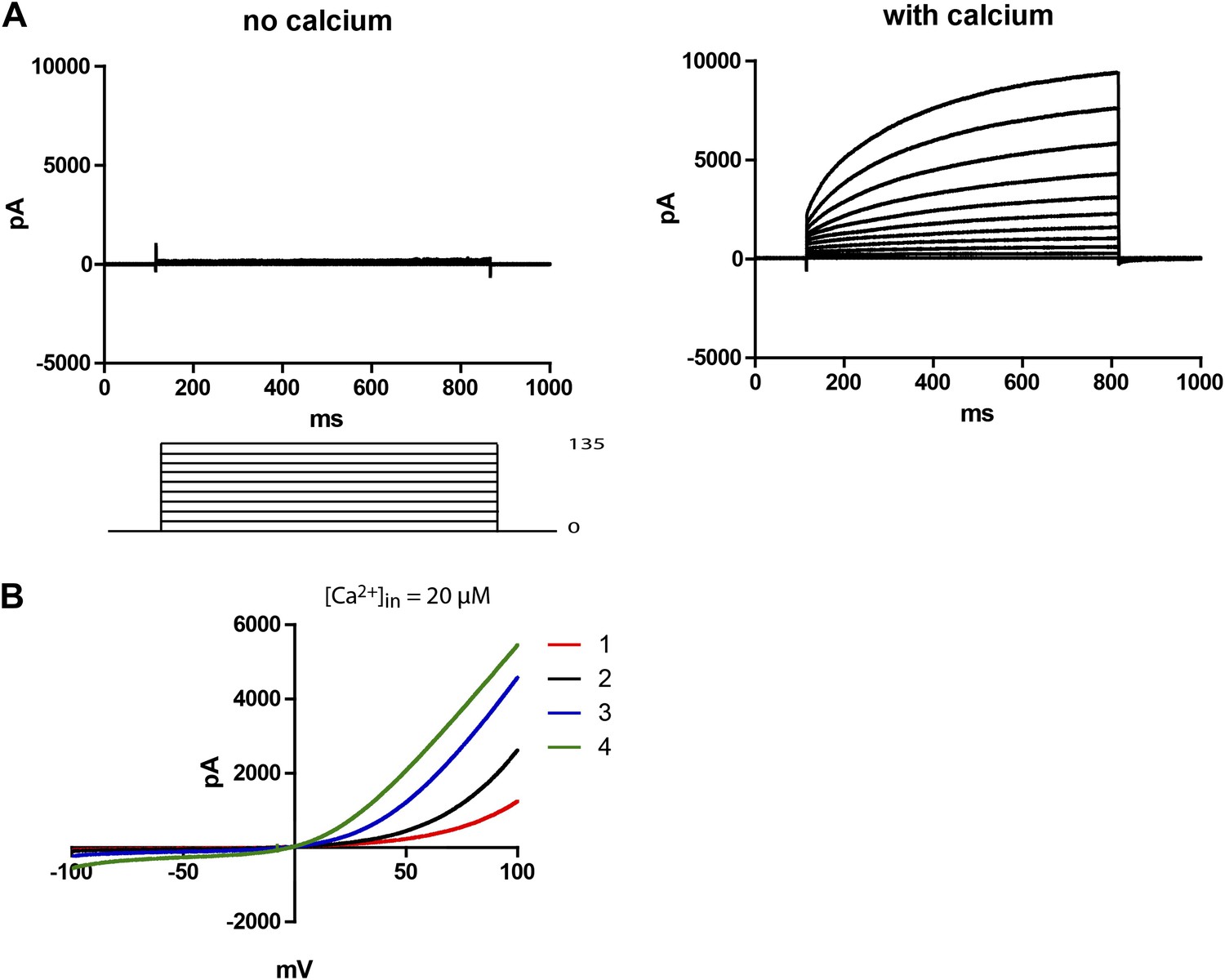
Subdued is a calcium and voltage-dependent channel.
(A) Subdued-transfected HEK 293T cells were used for recording in whole cell patch clamp experiments. No current was observed without calcium in the pipette (left), but large time dependent currents were observed (right) when calcium was added to the pipette (200 µM free calcium). Unless otherwise mentioned, all recordings in this study were done with 200 µM free calcium in the pipette and symmetrical NaCl in the pipette and bath solution. (B) Representative traces showing outward rectification of Subdued current in voltage ramps from −100 to +100 mV. Traces 1–4 were taken sequentially and show an increase in current and decrease in rectification over time (20 µM free calcium in pipette). The ramps were done at a rate (dV/dT) of 0.067 V/s.
Figure 3
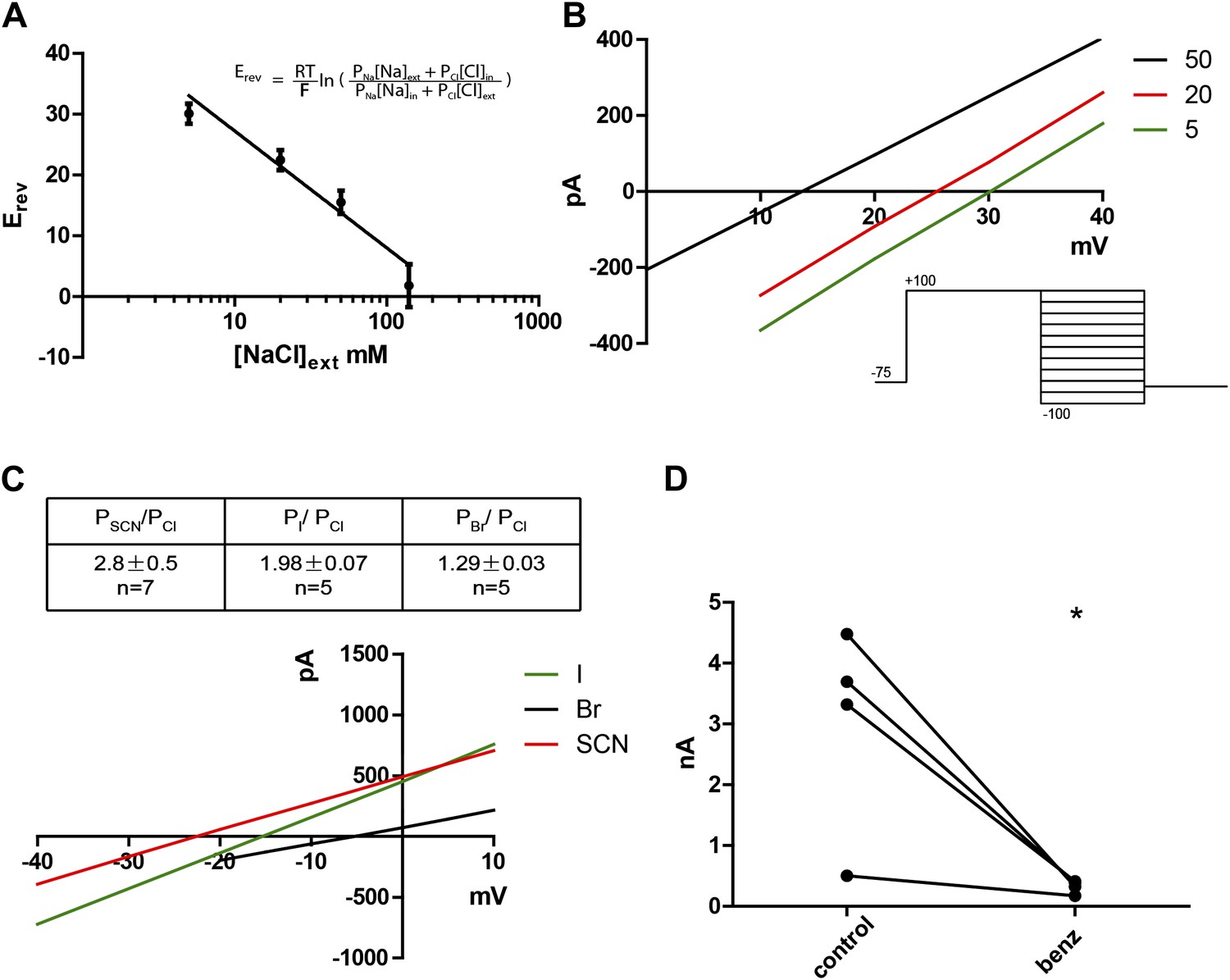
Subdued displays hallmark ionic selectivity of classic CaCCs and is blocked by a known CaCC inhibitor.
(A) NaCl gradients were introduced across the membrane by varying NaCl concentrations of external solutions. The reversal potential (Erev) at each concentration was obtained and fitted to the Goldman-Hodgkin-Katz (GHK) equation from which the PNa/PCl was determined to be 0.16. (B) Representative I/V plots obtained by varying external NaCl (in mM). A diagram of the voltage protocol used to measure Erev is shown below. After a 750 ms activating pre-pulse to +100 mV, instantaneous tail currents were measured from test voltages −100 to +100 mV in 20 mV steps. After an initial estimate of Erev using this protocol, test potentials and voltage increments were refined, while pre-pulse conditions and the length of voltage steps remained constant. For each NaCl concentration, n = 4 to n = 9. (C) Subdued preferentially permeates larger anions with the selectivity sequence: SCN > I > Br > Cl. Bi-ionic conditions were introduced by varying the external solution. Erev and permeability ratios were obtained by the same methodology as described for (B). Representative I/V plots obtained by varying the anion in external solutions using the same voltage protocols as described in (B) are shown. (D) Subdued is significantly and reversibly blocked by 20 µM benzbromarone (n = 4, p<0.05, Student’s t-test).
Figure 4
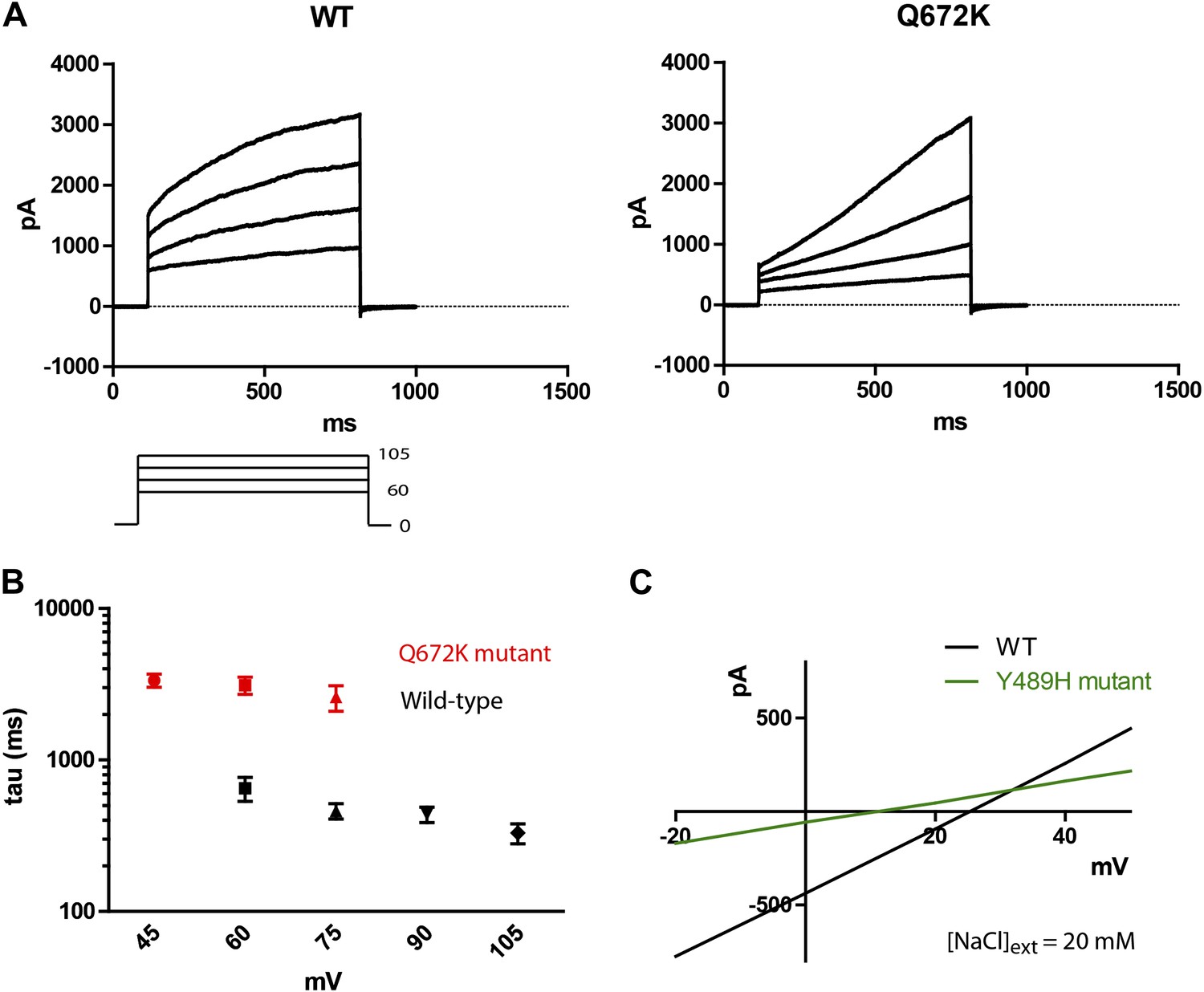
Mutations of subdued change properties of observed currents.
(A) Different kinetic properties in the wild-type (WT) and Q672K mutant channel revealed by a voltage step protocol (750 ms in 15 mV increments). (B) Semi-log plots of mean activation time constants (τ) as a function of voltage. τ was derived from the single exponential fitting of the current traces obtained from 750 ms (WT, n = 6) and 5 s (Q672K, n = 4) voltage steps. Time constants at 60 and 75 mV were significantly different for the two channels (p<0.001, Student’s t-test). (C) A Y489H mutation decreases chloride selectivity compared to the WT channel. A representative I/V plot showing the shift in Erev in a 20 mM external NaCl solution. WT Erev was determined to be (25 ± 4) mV, n=5; the Y489H mutant Erev was significantly different at (11 ± 2) mV, (n = 4, p<0.05, Student’s t-test). Data were obtained using methodology described in Figure 2B.
Figure 5 with 3 supplements

Subdued plays a role in host defense in Drosophila melanogaster.
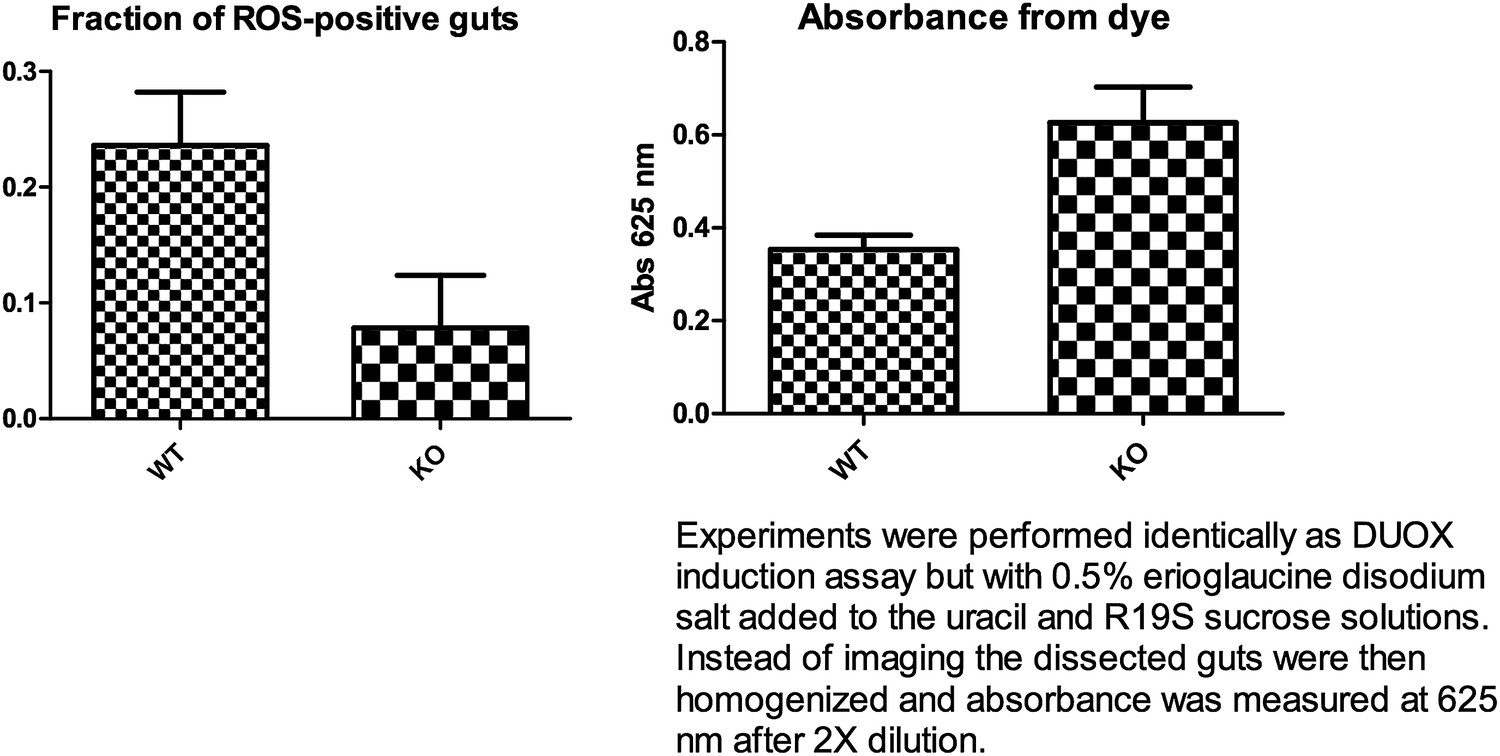
(A) Subdued knockout flies display susceptibility to Serratia infection. Wild-type (WT) and knockout (KO) flies were fed on a Db11/sucrose solution and their survival monitored for 192 hr post-infection. Two independently generated KO strains, KO2 and KO11, were used. WT flies lived significantly longer compared to KO2 and KO11 (n = 4, p<0.001, two-way ANOVA). (B) KO flies accumulate higher titers of bacteria in the whole animal. 20 whole flies were homogenized 48 hr post-infection. Serially diluted homogenates were plated on agar and inspected for Db11 colony forming units (CFU). Significantly more bacteria were recovered from the KO flies (n = 7, Student’s t-test, p<0.01). (C) KO flies do not consume more food than WT flies. Four fly guts were dissected and homogenized from vials of 20 flies fed with Db11/sucrose solution containing 0.5% wt/vol erioglaucine disodium salt (FDC Blue #1) 72 hr post-infection. The amount of food eaten by the flies was estimated by measuring absorbance of the dye. KO flies tended to consume significantly less food than WT flies (n = 5, p<0.01, repeated measures one-way ANOVA and Tukey’s multiple comparison test). No significant difference was observed between the KO strains. (D) The homogenate obtained from experiments described in (C) was inspected for Db11 colony forming units (CFU). Significantly higher amounts of bacteria were recovered from KO compared to WT fly guts (n = 6, p<0.05, repeated measures one-way ANOVA and Tukey’s multiple comparison test). No significant difference was observed between KO strains.
Figure 5—figure supplement 1
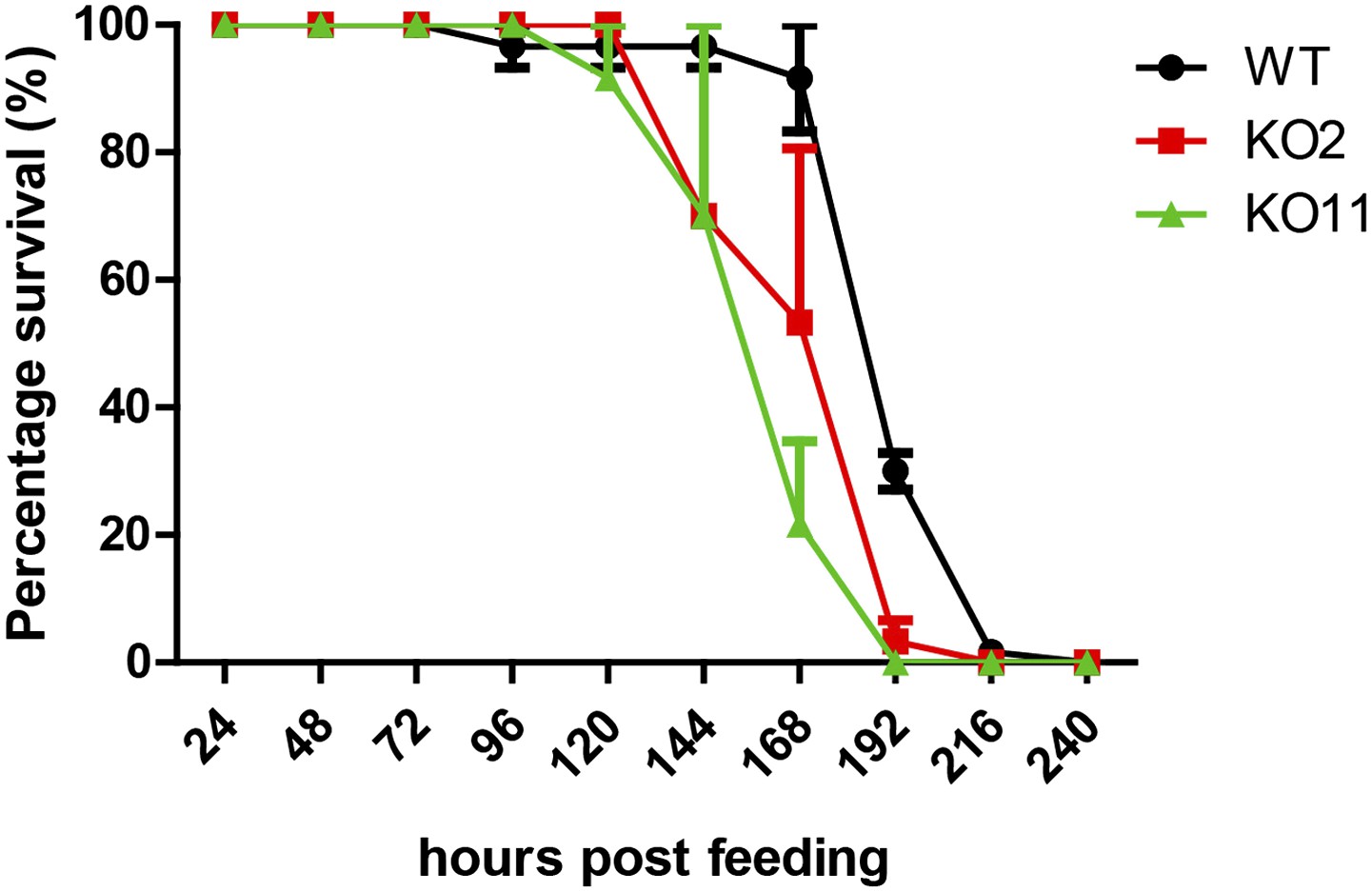
Knockout (KO) flies do not display significant lethality relative to wild-type (WT) flies upon ingestion of UV-killed Db11.
Db11/sucrose solution was irradiated with UV for 15 min prior to administration to the flies. While non-irradiated Db11 caused high mortality in KO strains 96 hr post-infection, UV-irradiated Db11 did not bring about noticeable lethality in all three strains at this time point. Increased lethality at later time points could arise from general reduced fitness of the KO strains. UV-irradiated bacterial feeding was done in a separate Db11-free incubator (n = 3).
Figure 5—figure supplement 2
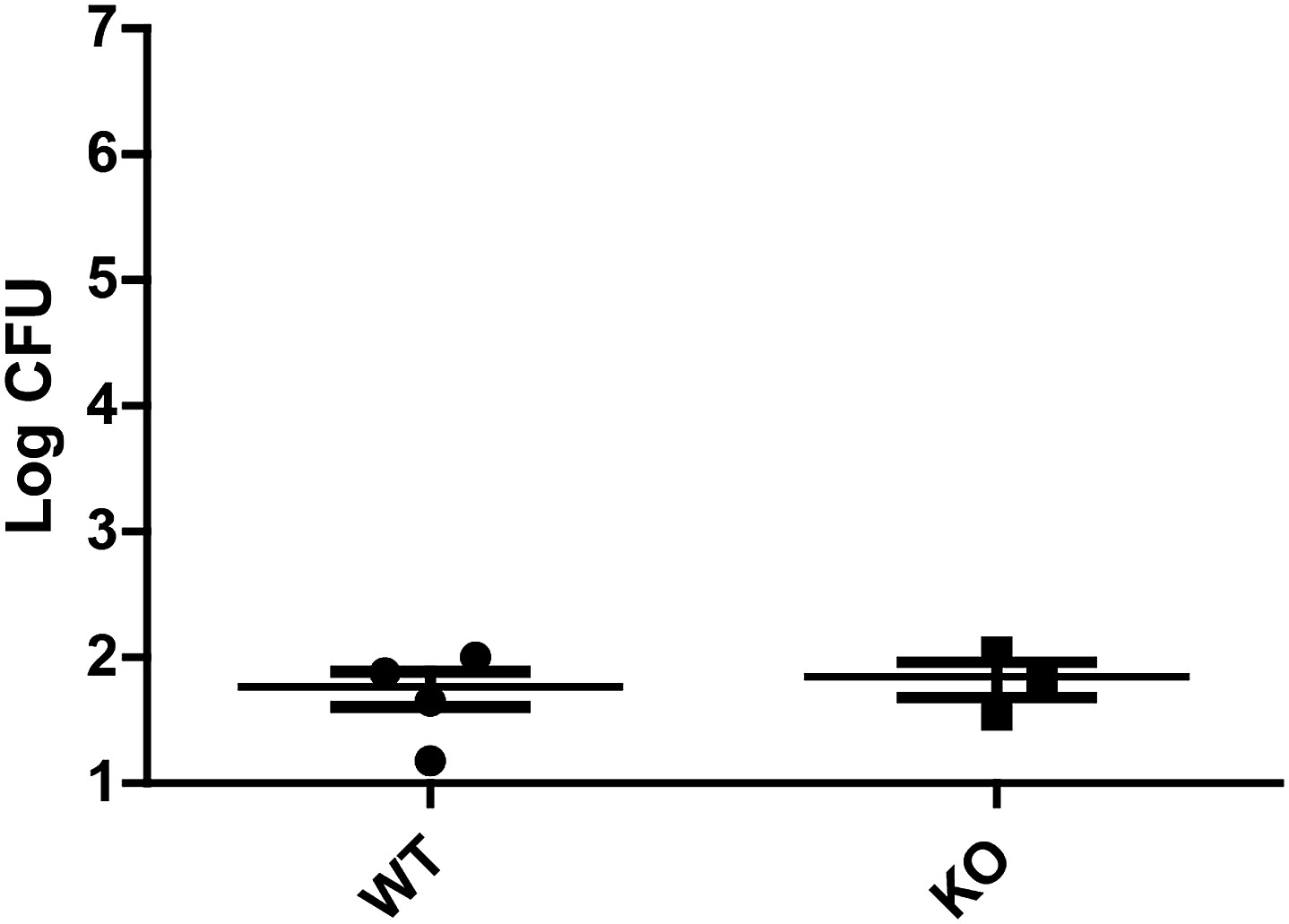
Feeding flies gentamicin greatly reduces Db11 counts from gut dissections.
Nehme et al. observed that after 24 hr of Db11 feeding, a significant amount of bacteria had entered the hemocoel and adhered to dissected guts in hemocyte-impaired flies. To explore the possibility that KO flies accumulated bacteria in the hemocoel which then adhered to the guts, flies were fed with Db11/sucrose solution for 24 hr and then switched to vials containing 500 µg/ml gentamicin with Db11/sucrose solution. After another 48 hr, four fly guts were dissected and the bacteria recovered from the guts were plated and counted. The low Db11 CFU counts from flies fed with gentamicin show that hemocoel-resident Db11 that adhere to the gut do not contribute greatly to the population of bacteria recovered from gut dissections. The data also suggest that it is unlikely that KO flies have severe impairment of hemocyte and hemocoel defenses.
Figure 5—figure supplement 3
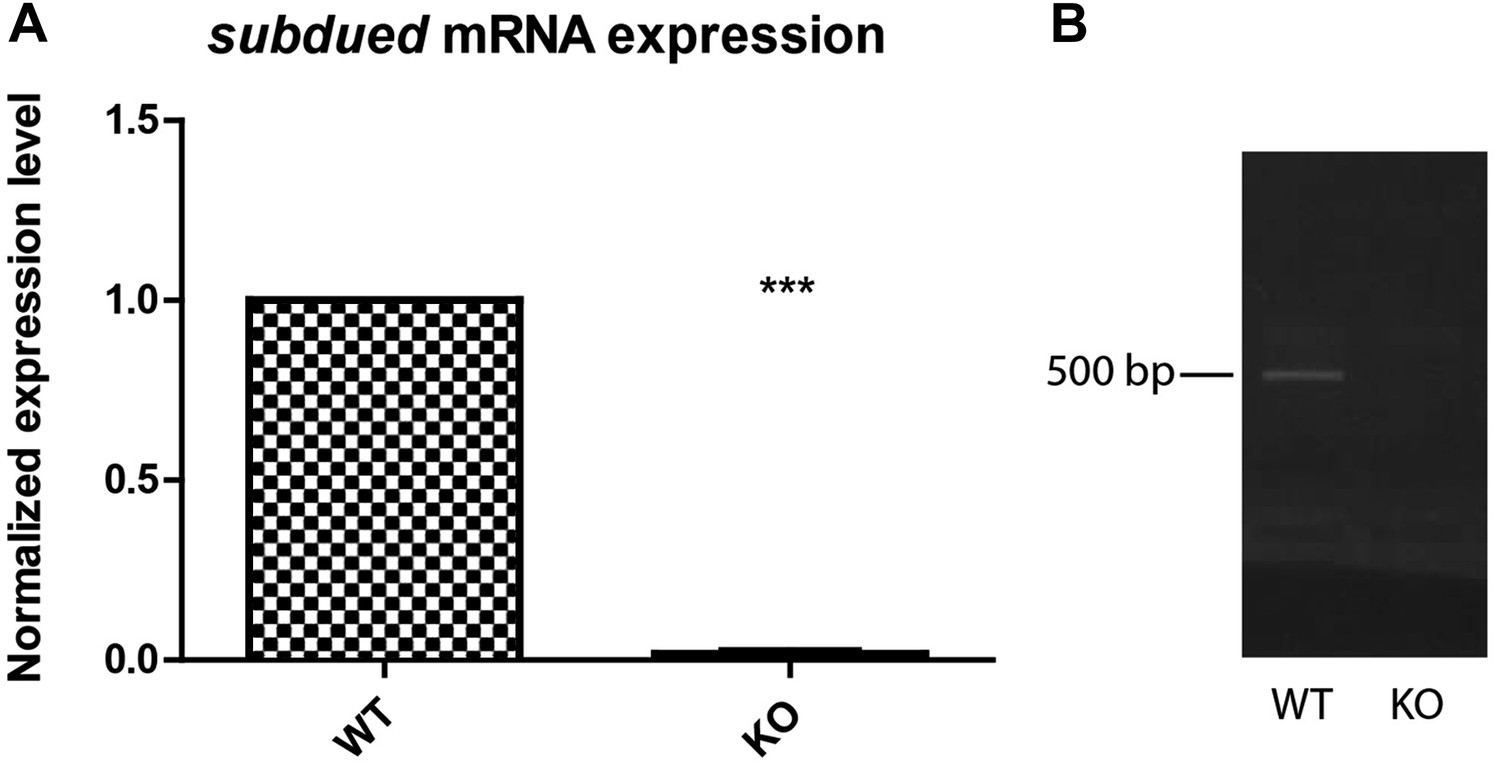
Drosophila guts express the subdued gene.
RNA extraction was performed on four whole dissected guts and subdued expression was determined by performing reverse transcription and quantitative PCR (A, n = 3, p<0.001), using KO guts as a control. For further verification, a different primer set was used to perform regular PCR using the same cDNA as a template (B).
Author response image 1
Author response image 2
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Subdued, a TMEM16 family Ca2+-activated Cl− channel in Drosophila melanogaster with an unexpected role in host defense
eLife 2:e00862.
https://doi.org/10.7554/eLife.00862




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}