Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl
- Carnegie Institution for Science, United States
Figures
Figure 1 with 1 supplement

ARF6 ChIP-Seq analyses.
(A) Distribution of ARF6 binding peaks relative to gene structure (−5000 base pairs from transcription start site to +1000 base pairs downstream of 3′ end). (B) Most of the early auxin-activated genes are ARF6 targets. Numbers above the columns indicate number of genes up- or down-regulated by 30 or 120 min of auxin treatments. (C) Venn diagram shows significant overlap among binding target genes of BZR1, PIF4 and ARF6. (D) ChIP-reChIP assay shows that BZR1 and ARF6 co-occupy shared target promoters. The enrichment of precipitated DNA was calculated as the ratio between transgenic plants and wild type control, normalized to that of the PP2A coding region as an internal control. Error bars indicate the SD of three biological repeats. (E) The G-box (CACGTG), HUD (CACATG), canonical AuxRE (TGTCTC) and TGTCGG are enriched in the ARF6 binding peaks associated with auxin-activated genes. GATCG (a random motif) is shown as a negative control. (F) Percentages of auxin-activated ARF6 binding peaks that have both E-box motif and core AuxRE (TGTC), only TGTC, or only E-box motifs. (G) Distribution of distance between E-box motifs and core AuxRE (TGTC) found in the ARF6 peaks associated with auxin-activated genes or total Arabidopsis genome. (H) ARF6 binding peaks having both E-box motifs and AuxRE have higher probability (%) of being associated with auxin-activated (30 or 120 min treatment) genes than the ARF6 binding peaks having only AuxRE. **p<0.01. (I) Venn diagram shows that genes activated by auxin, BR, or GA and genes repressed by light are enriched in the common binding targets of BZR1, PIF4 and ARF6. Numbers in the Venn diagram indicate percentage of corresponding genes (e.g., auxin-activated genes) in each section. Numbers in parentheses indicate percentage of genes in total Arabidopsis genome.
-
Figure 1—source data 1
(A) ARF6 target genes. ChIP-Seq analysis R (CSAR) software was used to identify binding peaks, with parameters (backg = 10, norm = −1, test = 'Ratio', times = 1e6, digits = 2) (Muino et al., 2011). Binding peaks with FDR <0.01 were finally defined as the ARF6 binding peak and genes having at least one ARF6 binding peak within its promoter (−3 kb) or coding region or 1 kb downstream from stop codon were considered direct target genes. max: maximum peak value; u3000, u2000, u1000: upstream 3000, 2000, or 1000 bp from transcription start site; d0: coding region; d1000: downstream 1000 bp from stop codon. (B) BZR1 target genes. ChIP-Seq experiment was performed using the BZR1p::BZR1-CFP transgenic seedlings grown in the dark for 5 days, and anti-YFP antibody. Data were analyzed as described in legend of Figure 1—source data 1A. (C) Previous PIF4 ChIP-seq result (Oh et al., 2012) was re-analyzed with same statistical method as described in Figure 1—source data 1A, to define PIF4 target genes.
- https://doi.org/10.7554/eLife.03031.004
-
Figure 1—source data 2
Auxin-activated genes previously identified in hypocotyls (Chapman et al., 2012) were compared with ARF6 target genes identified by ChIP-Seq to identify the auxin-activated ARF6 target genes in hypocotyls.
30 or 120 min: genes are activated after 30 or 120 min of auxin treatment.
- https://doi.org/10.7554/eLife.03031.005
Figure 1—figure supplement 1
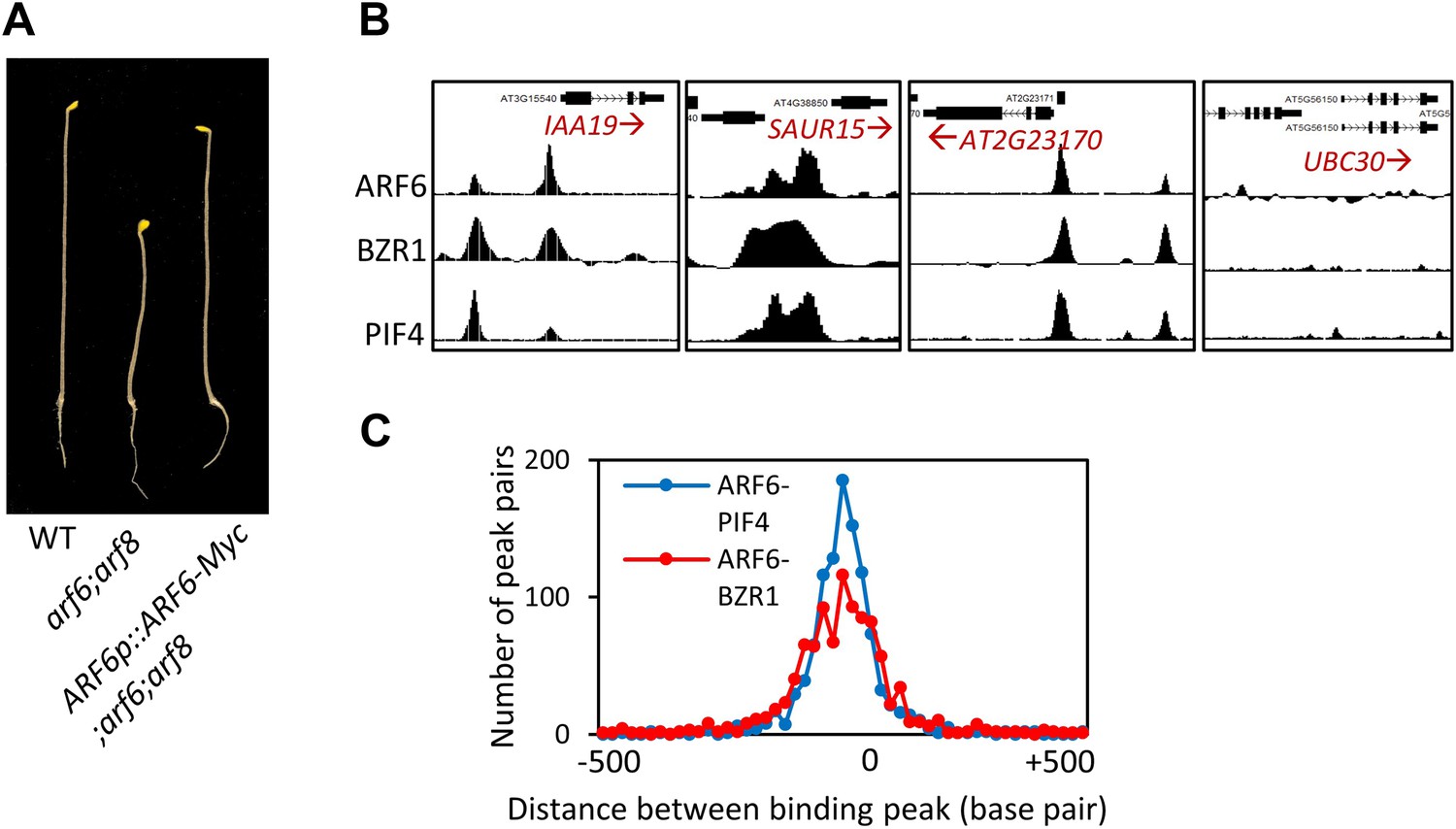
(A) ARF6-Myc regulated by ARF6 native promoter restored short hypocotyl of the arf6;arf8 double mutant. Seedlings were grown in the dark for 6 days. Representative seedlings are shown. (B) Representative ARF6, BZR1, and PIF4 binding peaks in the promoters of ARF6, BZR1 and PIF4 common target genes (IAA19, SAUR15 and AT2G23170) and UBC30 promoter as a negative control. (C) Distance distribution of ARF6 and PIF4 binding peaks or ARF6 and BZR1 binding peaks in the ARF6, BZR1 and PIF4 common target genes.
Figure 2 with 3 supplements
ARF6 interacts with BZR1 and PIF4.
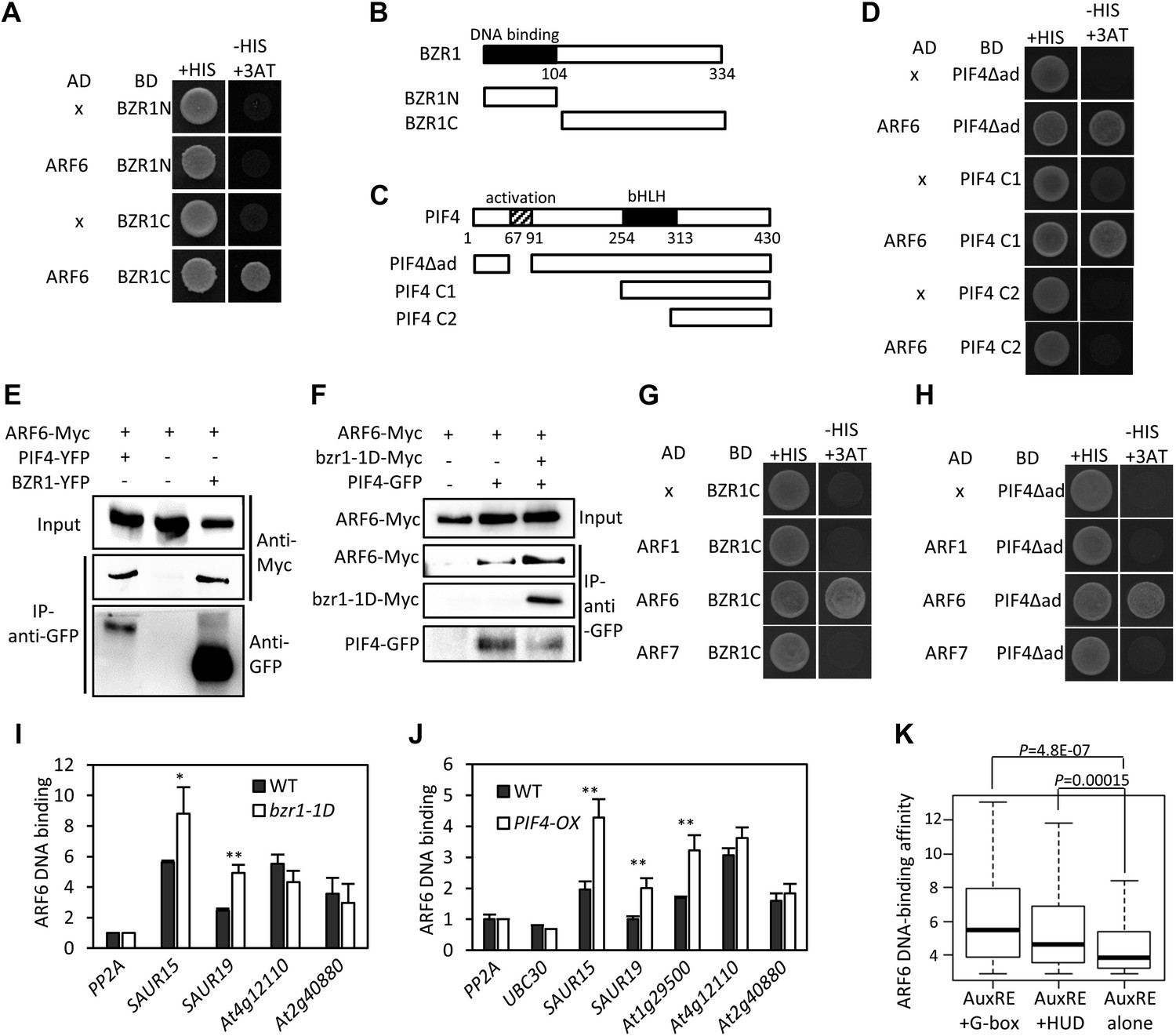
(A) ARF6 interacts with BZR1 in yeast two-hybrid assay. Yeast clones were grown on the synthetic dropout (+HIS) medium or synthetic dropout medium without histidine (−HIS) plus 1 mM 3AT. (B and C) Box diagram of various fragments of BZR and PIF4 used in (A, D, G, H). (D) ARF6 interacts with PIF4 in yeast two-hybrid assay. (E) ARF6 interacts with BZR1 and PIF4 in vivo. Transgenic plants expressing the indicated fusion proteins were used for immunoprecipitation using anti-GFP antibody, and the immunoblots were proved with anti-Myc antibody to detect interaction with the Myc-tagged ARF6 protein. (F) BZR1 enhances the ARF6–PIF4 interaction. Arabidopsis mesophyll protoplasts were transfected to express ARF6-Myc alone or together with PIF4-GFP and bzr1-1D-Myc as indicated, and the extracted proteins were immunoprecipitated by anti-GFP antibody. Gel blots were probed with anti-Myc or anti-GFP antibody. (G and H) BZR1 (G) and PIF4 (H) interact with ARF6, but not with ARF1 and ARF7 in yeast two-hybrid assays. (I) ARF6 DNA-binding is enhanced by bzr1-1D. Seedlings (35S::ARF6-Myc (WT) and 35S::ARF6-Myc;bzr1-1D (bzr1-1D)) grown on the 2 μM PPZ in the dark for 6 days were used for ChIP assays. Error bars in the (I and J) indicate the SD of three biological repeats. *p<0.05 and **p<0.01. (J) PIF4-OX enhances ARF6 DNA-binding. Seedlings (35S::ARF6-Myc (WT) and 35S::ARF6-Myc;PIF4-OX (PIF4-OX)) grown under light were used for ChIP assays of ARF6 binding to the indicated promoters. (K) Box plot shows that ARF6 binding peaks having both E-box motifs and AuxRE tend to have higher ARF6 DNA-binding affinity. ARF6 DNA-binding affinity was based on the peak score from the ARF6 ChIP-Seq analysis with CSAR.
Figure 2—figure supplement 1
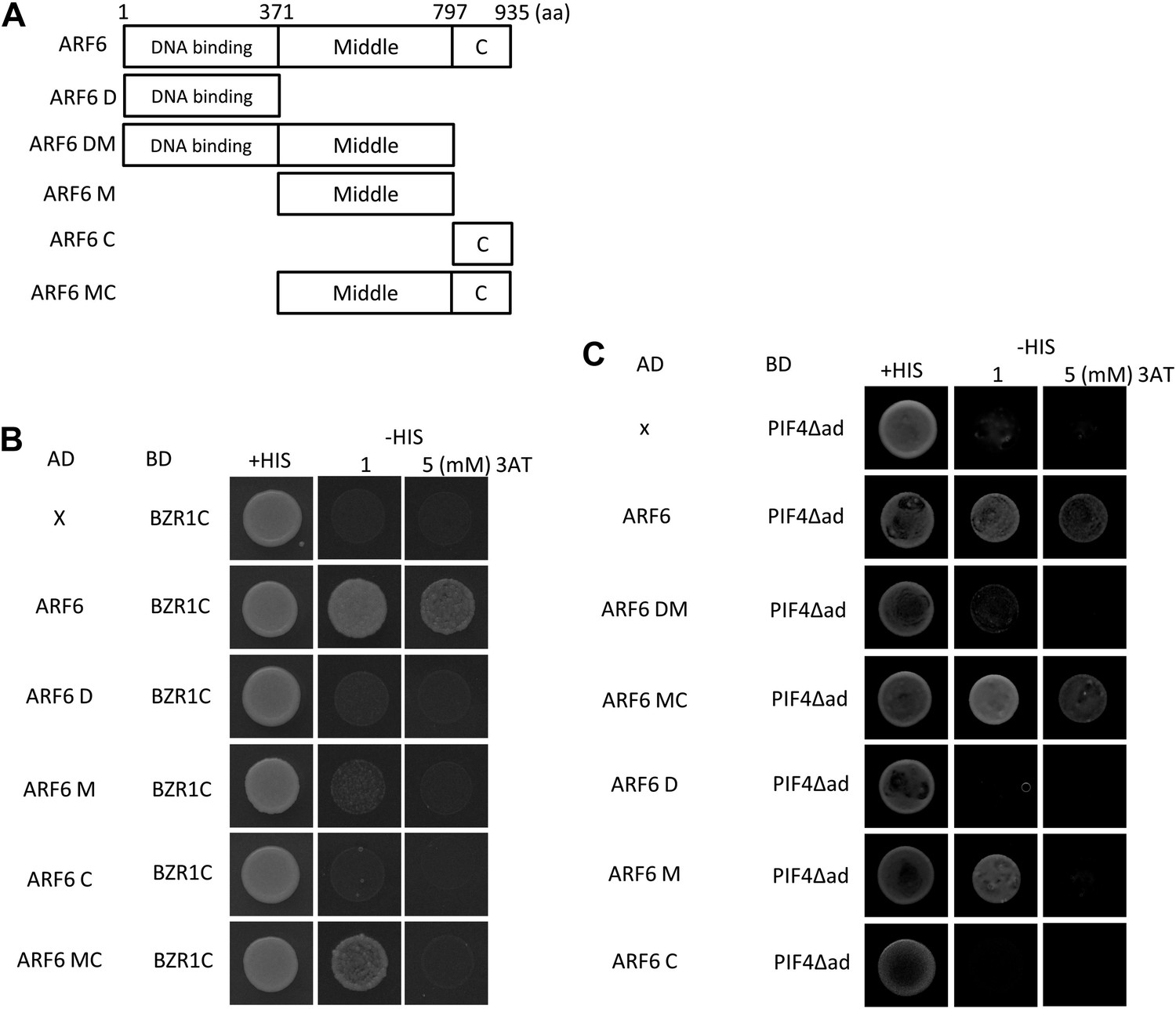
(A) Box diagram of various fragments of ARF6 used in the yeast-two hybrid assay. (B) ARF6 middle and C-terminal domains are required for the interaction with BZR1. Yeast clones were grown on the synthetic dropout (+HIS) or synthetic dropout without histidine (−HIS) plus 1 or 5 mM 3AT medium. AD: activation domain fusion vector, BD: DNA binding domain fusion vector, x: empty vector. (C) ARF6 middle and C-terminal domains are required for the interaction with PIF4.
Figure 2—figure supplement 2
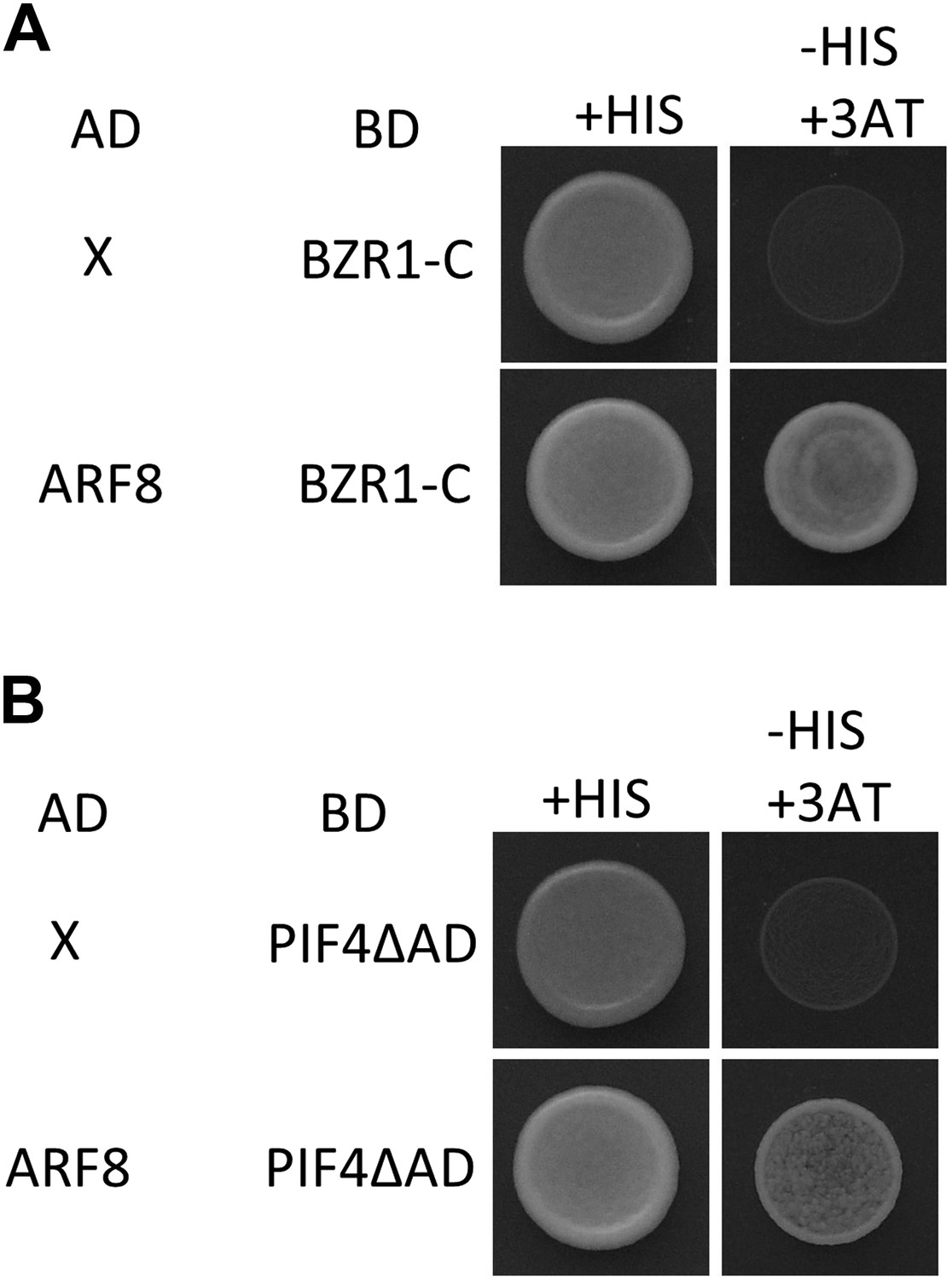
ARF8 interacts with both BZR1 and PIF4.
Yeast clones were grown on the synthetic dropout (+HIS) or synthetic dropout without histidine (−HIS) plus 1 mM 3AT medium. AD: activation domain fusion vector, BD: DNA binding domain fusion vector, x: empty vector.
Figure 2—figure supplement 3
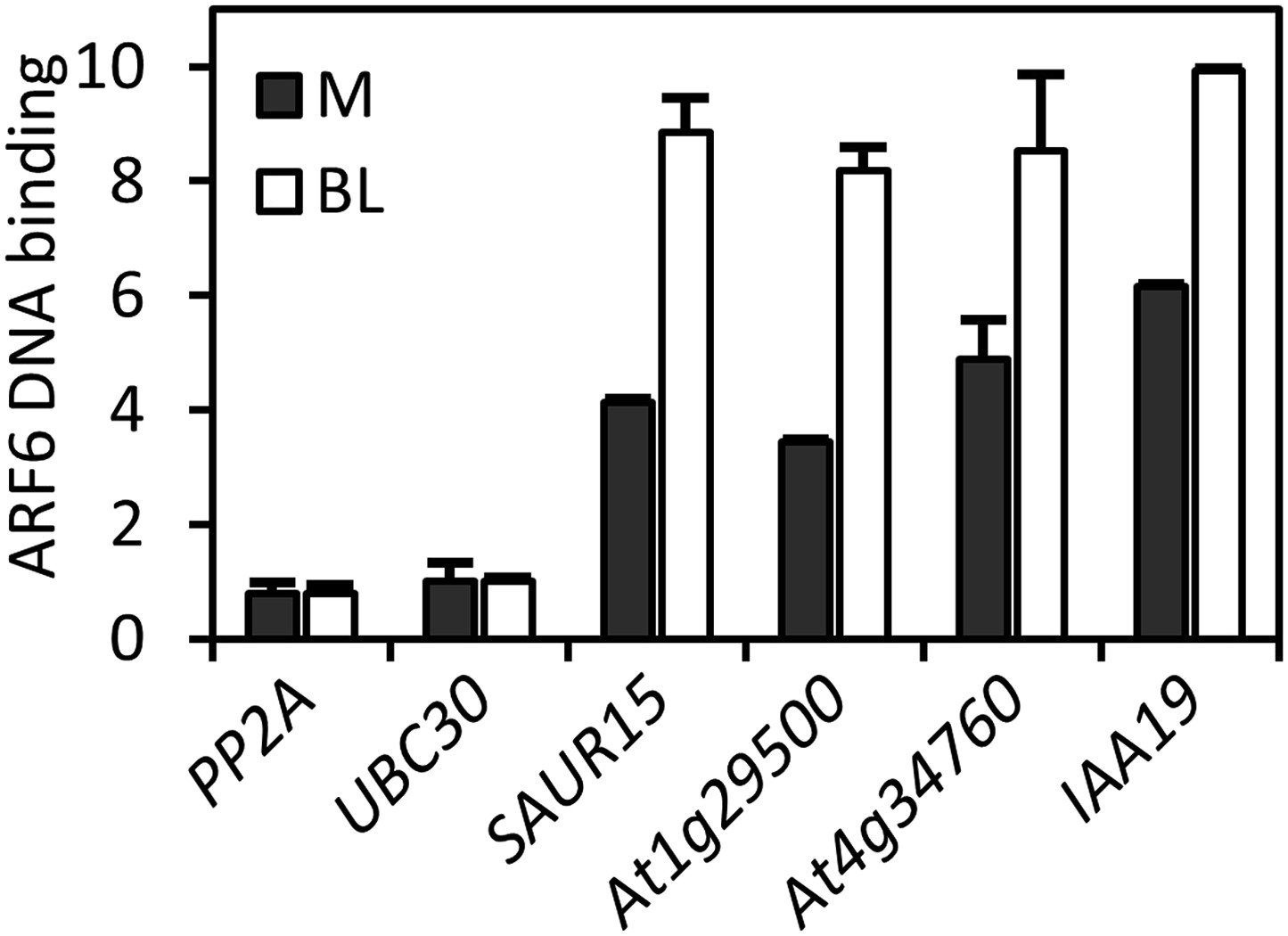
ARF6 DNA-binding on the common targets of ARF6 and BZR1 is enhanced by BR treatment.
Seedlings (35S::ARF6-Myc) grown on the medium containing 2 μM PPZ (M) or 2 μM PPZ + 100 nM brassinolide (BL) were used for the ChIP assay to determine ARF6 DNA-binding. Enrichment of DNA was calculated as the ratio between transgenic plants and wild type (Col-0), normalized to that of the PP2A coding region as an internal control.
Figure 3 with 2 supplements
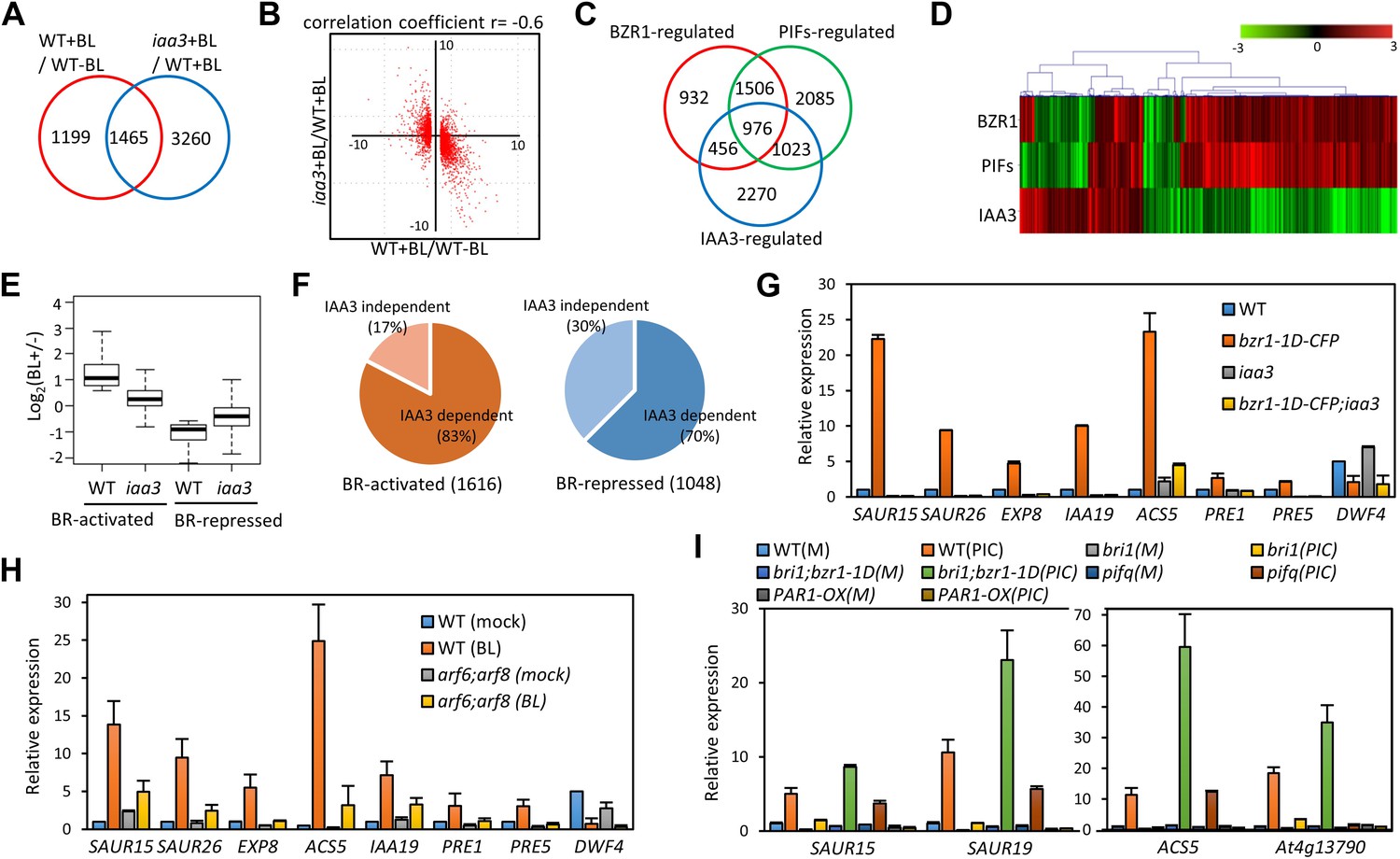
ARF6, BZR1, and PIF4 synergistically induce gene expression.
(A) Significant overlap between BR-regulated genes and IAA3-regulated genes. (B) Scatter plot of log2-fold change values in the 1465 overlapping set of IAA3- and BR-regulated genes. (C) Significant overlap among BZR1-, PIFs-, and IAA3-regulated genes. (D) Heat map of the 976 genes co-regulated by BZR1, PIFs, and IAA3. Scale bar indicates fold changes (log2 value). (E) Box plot representation of the 1616 BR-activated or the 1048 BR-repressed genes in the WT and iaa3/shy2-2. (F) Percentage of IAA3-dependent and IAA3-independent BR-regulated genes. Genes that were not significantly affected by BR treatment in iaa3/shy2-2 are defined as IAA3-dependent BR-regulated genes. (G) qRT-PCR analysis of BZR1-regulated genes in etiolated seedlings grown on 2 μM PPZ medium. Similar results are obtained from two independent biological repeats. Error bars indicate the SD of three technical repeats. (H) qRT-PCR analysis of BR-regulated genes in the seedlings treated with either mock or 100 nM BL for 4 hr. Error bars indicate the SD of three biological repeats. (I) qRT-PCR analysis of auxin responsive genes in the seedlings grown on medium containing no hormone (M) or 1 μM picloram, an artificial auxin. Error bars indicate the SD of three biological repeats.
-
Figure 3—source data 1
BR-regulated genes in wild type and their BR-responsive expression in the iaa3 mutant.
Seedlings were grown on 2 µM propiconazole medium for 5 days in the dark and treated with mock or 100 nM brassinolide (BL) for 4 hr. BR-regulated genes were defined by 1.5-fold difference between wild type (+BL) and wild type (−BL) with p-value<0.01.
- https://doi.org/10.7554/eLife.03031.012
-
Figure 3—source data 2
Genes whose expression levels are affected in the iaa3 mutant.
Seedlings of wild type and iaa3 were grown on 2 µM propiconazole medium for 5 days in the dark and treated with 100 nM brassinolide for 4 hr. The IAA3-regulated genes were defined by 1.5-fold difference between iaa3 and wild type with p<0.01.
- https://doi.org/10.7554/eLife.03031.013
Figure 3—figure supplement 1
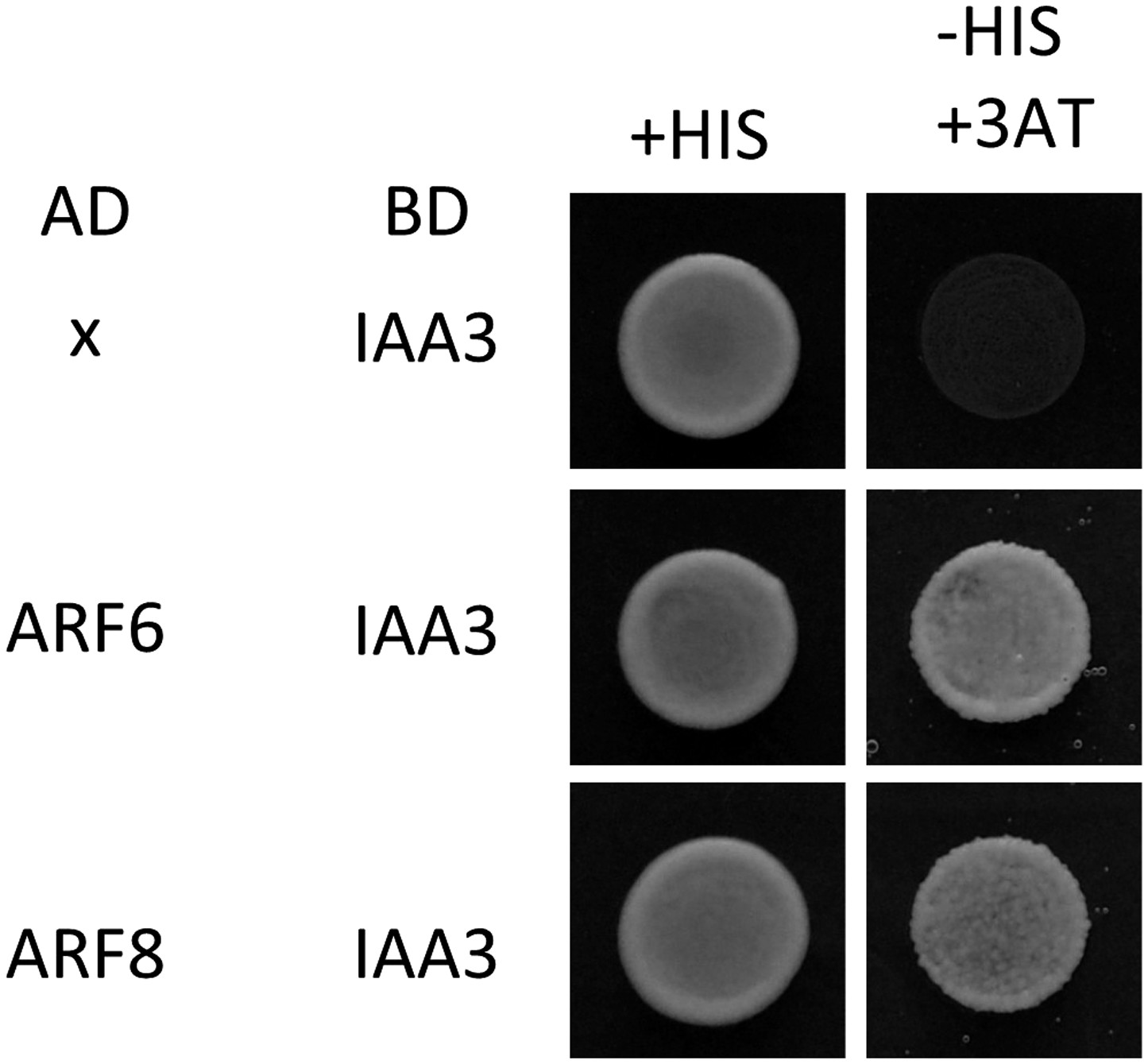
IAA3 interacts with both ARF6 and ARF8.
Yeast clones were grown on the synthetic dropout (+HIS) or synthetic dropout without histidine (−HIS) plus 1 mM 3AT medium. AD: activation domain fusion vector, BD: DNA binding domain fusion vector, x: empty vector.
Figure 3—figure supplement 2
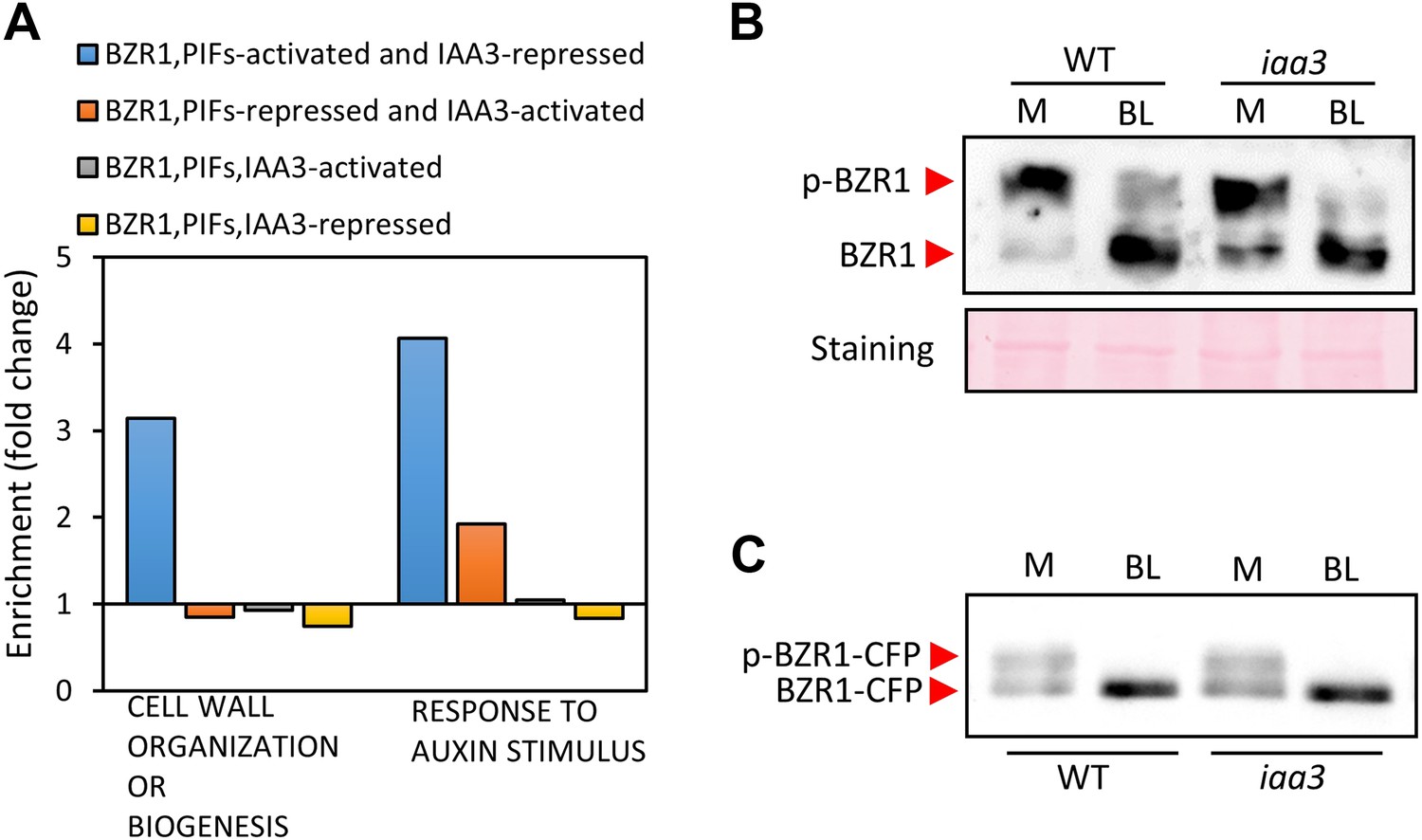
(A) Gene ontology analysis shows that the genes involved in cell wall organization or biogenesis, and the auxin responsive genes are enriched in the BZR1, PIFs-activated but IAA3-repressed genes. (B) Endogenous BZR1 phosphorylation status is not affected in the iaa3/shy2-2 mutant. Seedlings (Col-0) were grown on the medium containing 2 μM BRZ for 5 days in the dark and then treated with either mock (M) or 100 nM brassinolide (BL) for 30 min. Endogenous BZR1 was detected by anti-BZR1 antibody. p-BZR1: phosphorylated BZR1, BZR1: de-phosphorylated BZR1. (C) BZR1-CFP phosphorylation status is not affected in the iaa3 mutant. Transgenic plants expressing BZR1-CFP driven by native BZR1 promoter (BZR1p::BZR1-CFP) in the wild type or iaa3 were grown on the regular MS medium for 5 days under white light and then treated with either mock (M) or 100 nM brassinolide (BL) for 1 hr. BZR1-CFP was detected by anti-GFP antibody. p-BZR1-CFP: phosphorylated BZR1-CFP, BZR1-CFP: de-phosphorylated BZR1-CFP.
Figure 4
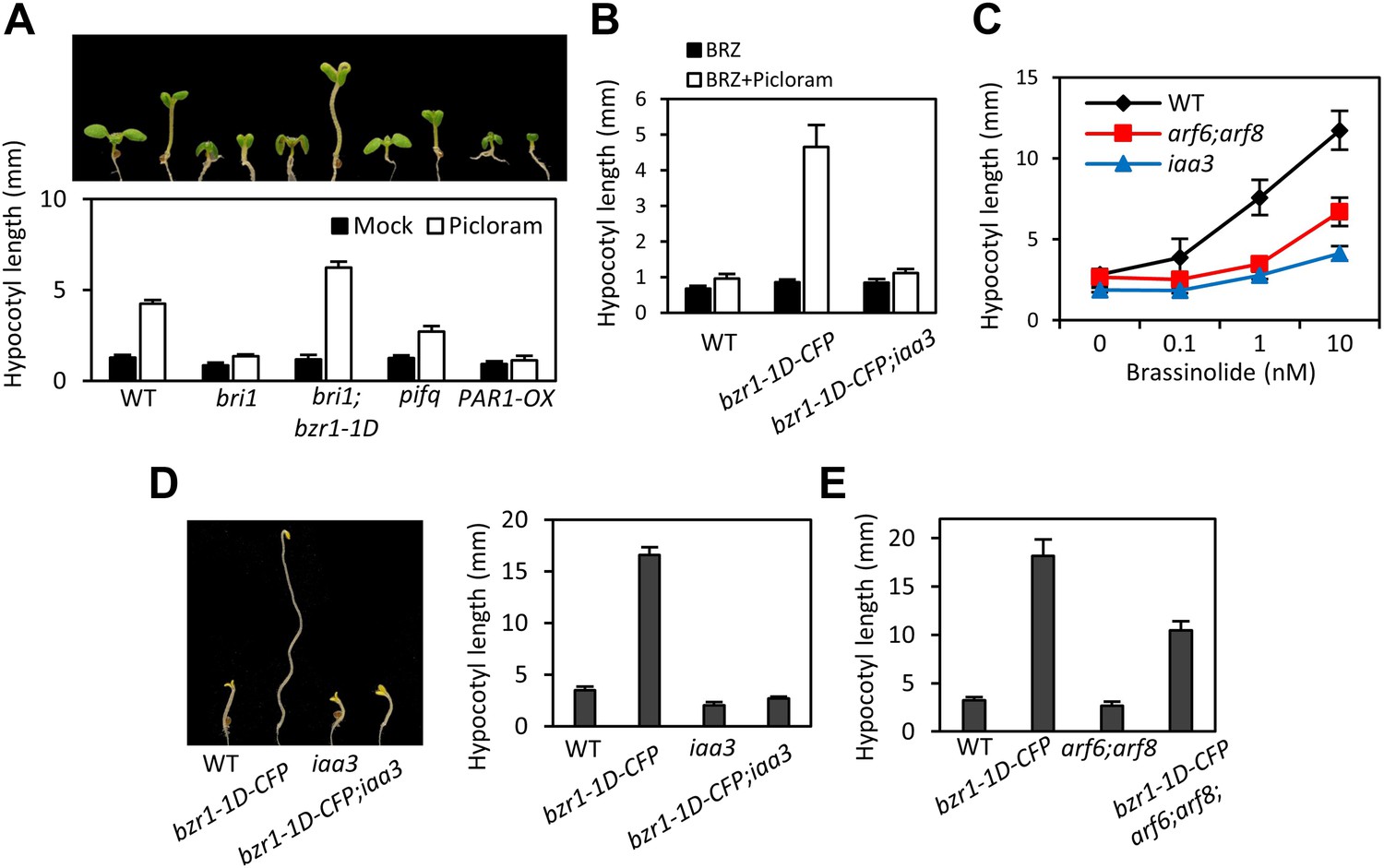
ARF6, BZR1, and PIF4 act interdependently in promoting hypocotyl elongation.
(A) BZR1 and PIFs are required for auxin promotion of hypocotyl elongation. Seedlings were grown on 5 μM artificial auxin picloram or mock medium. (B) Hypersensitivity of bzr1-1D to auxin is abolished by iaa3/shy2-2 mutation. Seedlings were grown on the medium containing 2 μM brassinazole (BRZ) with or without 5 μM artificial auxin picloram. (C) ARF6 and ARF8 are required for BR promotion of hypocotyl elongation. Seedlings were grown on medium containing 2 μM BRZ plus various concentration of BL in the dark. (D) The iaa3/shy2-2 mutation inhibits BZR1 promotion of hypocotyl elongation. Representative seedlings are shown in left panel and quantification of hypocotyl lengths are shown in right graph. Seedlings were grown on the 2 μM BRZ medium in the dark. (E) ARF6 and ARF8 are required for BZR1 promotion of hypocotyl elongation. Seedlings were grown on the 2 μM BRZ medium in the dark. All error bars in (A–E) indicate SD (n = 10 plants).
Figure 5 with 1 supplement

The HLH/bHLH module mediates developmental regulation of auxin sensitivity.
(A) PAR1 interacts with BEE2 and HBI1 in yeast two-hybrid assay. (B) ARF6 interacts with BEE2 and HBI1 in yeast two-hybrid assay. (C) The pre-amiR, IBH1-OX, and HBI1-SRDX plants are less sensitive to auxin. Seedlings were grown on hormone-free or 5 μM picloram medium for 7 days. Error bars indicate SD (n = 10 plants). (D) Auxin activation of gene expression is diminished in the pre-amiR and IBH1-OX plants. 7-day-old seedlings were treated with mock (M) or 1 μM IAA for 2 hr. (E) Young stems are more sensitive to auxin than mature stems. Young stems (2 cm stem from the top) and mature stems (2 cm stem from the bottom) were treated with mock (M) or 1 μM IAA for 2 hr. (F) Auxin sensitivity of mature stem is enhanced by PRE1-OX, HBI1-OX, and PIF4-OX. Numbers indicate ratios between IAA-treated and mock-treated. (G) The DWF4 expression is high in the young stems. (H) BZR1 is less phosphorylated in young stems than in mature stems. Proteins extracted from the young and mature stems of the BZR1p::BZR1-CFP transgenic plants were analyzed by anti-YFP immunoblotting. Ponceau S staining is shown for loading control. (I) Auxin sensitivity of mature stem is restored by bzr1-1D. Sections of mature and young stems from plants of same height were treated with IAA for 2 hr, and the expression levels of SAUR15 were analyzed by qRT-PCR. Numbers in (E, F, I) indicate ratios of the expression levels of IAA-treated to mock-treated. Error bars in (D–I) indicate the SD of three biological repeats.
Figure 5—figure supplement 1
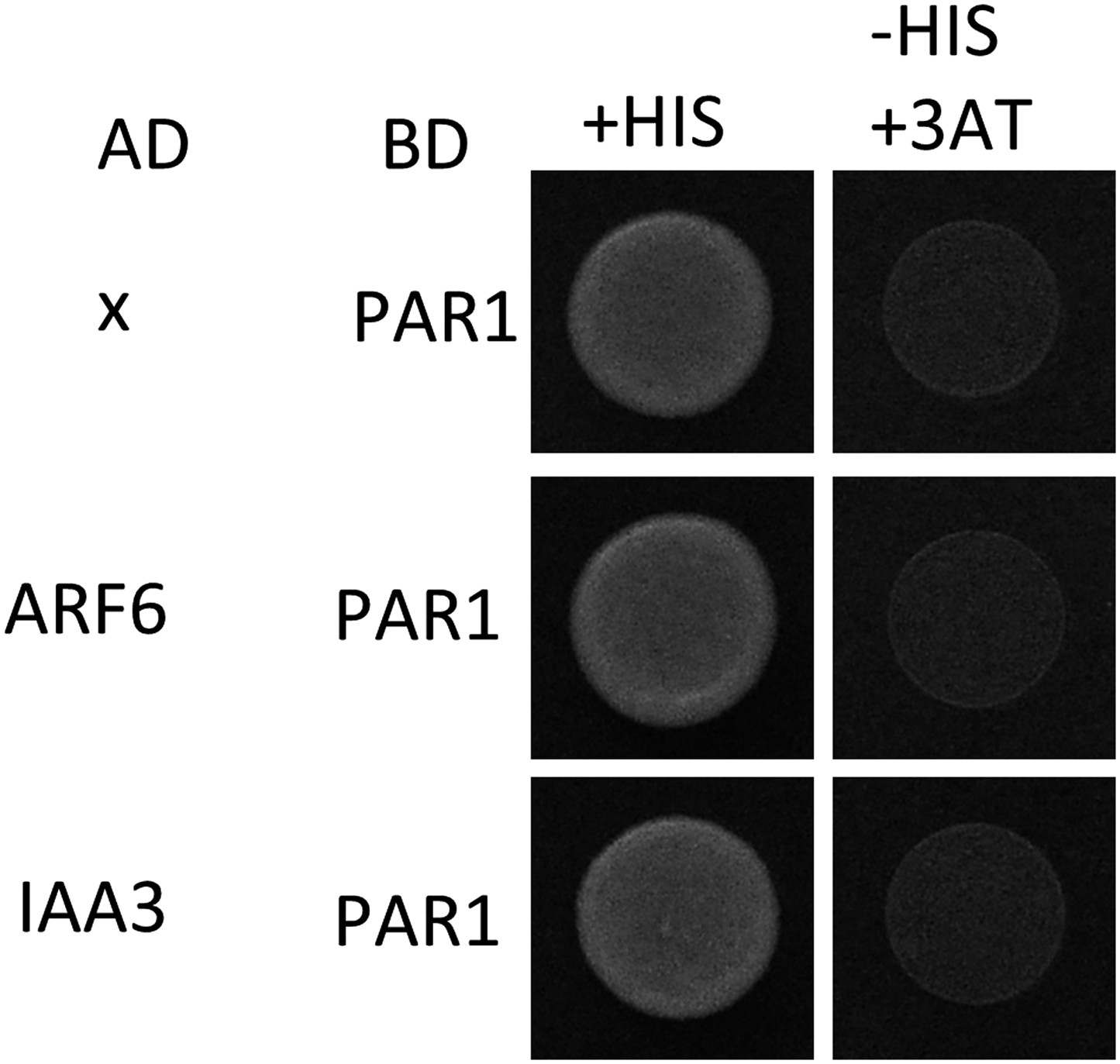
PAR1 does not interact with ARF6 and IAA3.
Yeast clones were grown on the synthetic dropout (+HIS) or synthetic dropout without histidine (−HIS) plus 1 mM 3AT medium. AD: activation domain fusion vector, BD: DNA binding domain fusion vector, x: empty vector.
Figure 6 with 2 supplements
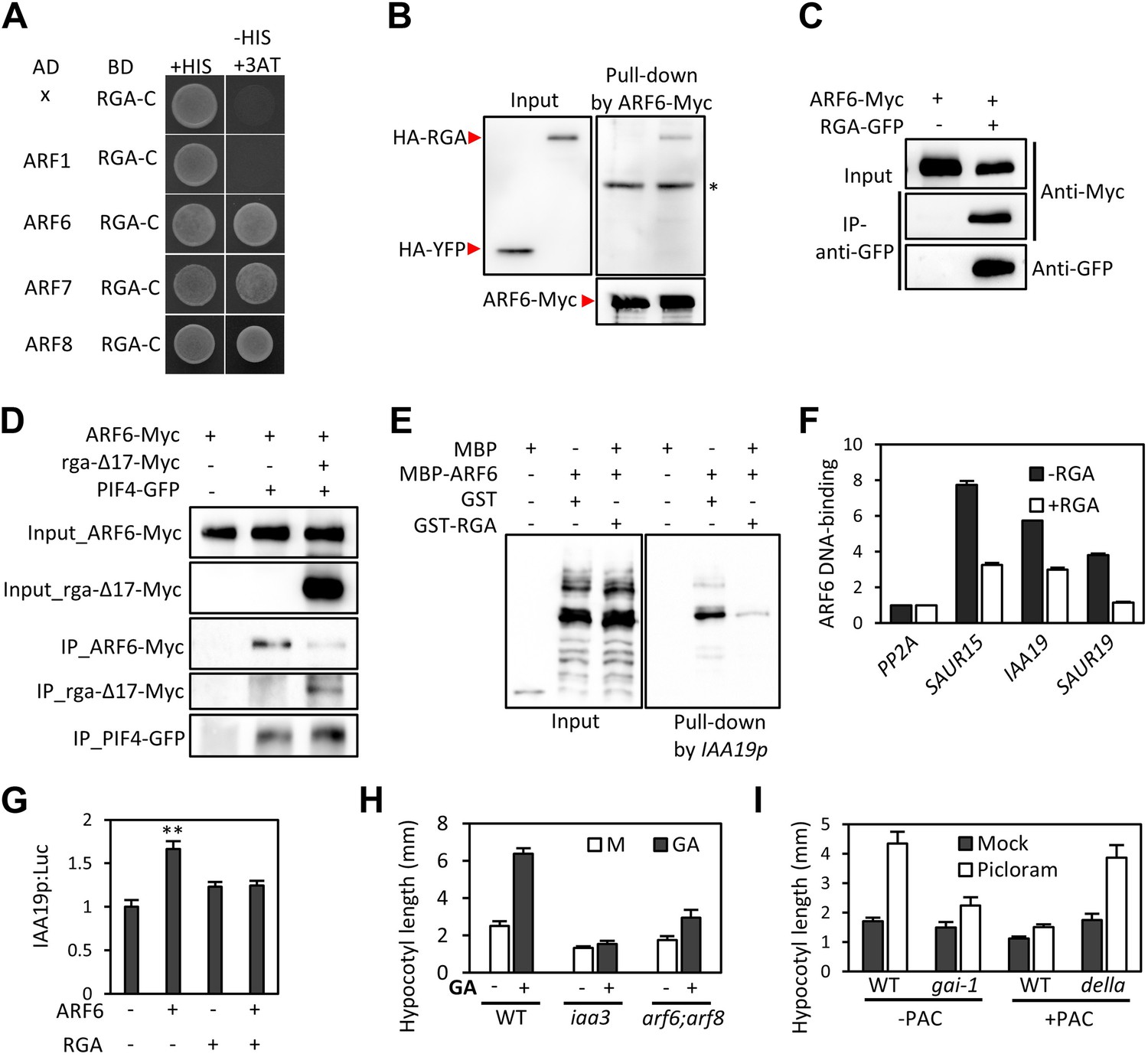
RGA interacts with ARF6 and blocks ARF6 binding to DNA.
(A) RGA interacts with ARF6, ARF7, and ARF8, but not ARF1 in the yeast two-hybrid assay. RGA with deletion of N-terminal 208 amino acids (RGA-C) was used for the assay. (B) RGA interacts with ARF6 in vitro. In vitro-translated HA-YFP and HA-RGA proteins were incubated with in vitro-translated ARF6-Myc protein bound to magnetic beads, and the pulled-down proteins were analyzed by immunoblot with anti-HA antibody. * indicates IgG band. (C) RGA interacts with ARF6 in vivo. Protein extracts from protoplasts transfected with ARF6-Myc or ARF6-Myc and RGA-GFP were immunoprecipitated with anti-GFP antibody, and analyzed by immunoblots with anti-GFP or anti-Myc antibody. (D) RGA disrupts the PIF4–ARF6 interaction. Arabidopsis mesophyll protoplasts were transfected to express ARF6-Myc alone or with PIF4-GFP and rga-Δ17-Myc as indicated, and the extracted proteins were immunoprecipitated by anti-GFP antibody. Gel blots were probed with anti-Myc or anti-GFP antibody. (E) RGA inhibits ARF6 binding to the IAA19 promoter in DNA pull-down assay. (F) RGA inhibits ARF6 DNA-binding ability in vivo. Protoplasts transfected with GFP-Myc (negative control) or ARF6-Myc with or without RGA-GFP were used for ChIP assay. Error bars indicate the s.d. of two technical repeats. Similar results were obtained in two independent experiments. (G) RGA inhibits ARF6 transcriptional activation activity. IAA19p::Luc was co-transfected with ARF6-GFP, RGA-GFP, or both, into Arabidopsis mesophyll protoplasts. The IAA19p::Luc activities were normalized by the 35S::renilla luciferase. Error bars indicate the s.e. of 10 biological repeats (n = 10) and **p<0.01. (H) Auxin signaling mutants are less sensitive to GA. Seedlings were grown on the 10 μM paclobutrazole with or without 1 μM GA in the dark. Error bars indicate SD (n = 10 plants). (I) DELLA inhibits the auxin promotion of hypocotyl elongation. Seedlings were grown on MS medium for 3 days and then transferred to the medium containing mock or 5 μM picloram, with or without 10 μM paclobutrazol (PAC), and incubated for 4 days. Error bars indicate SD (n = 10 plants).
Figure 6—figure supplement 1
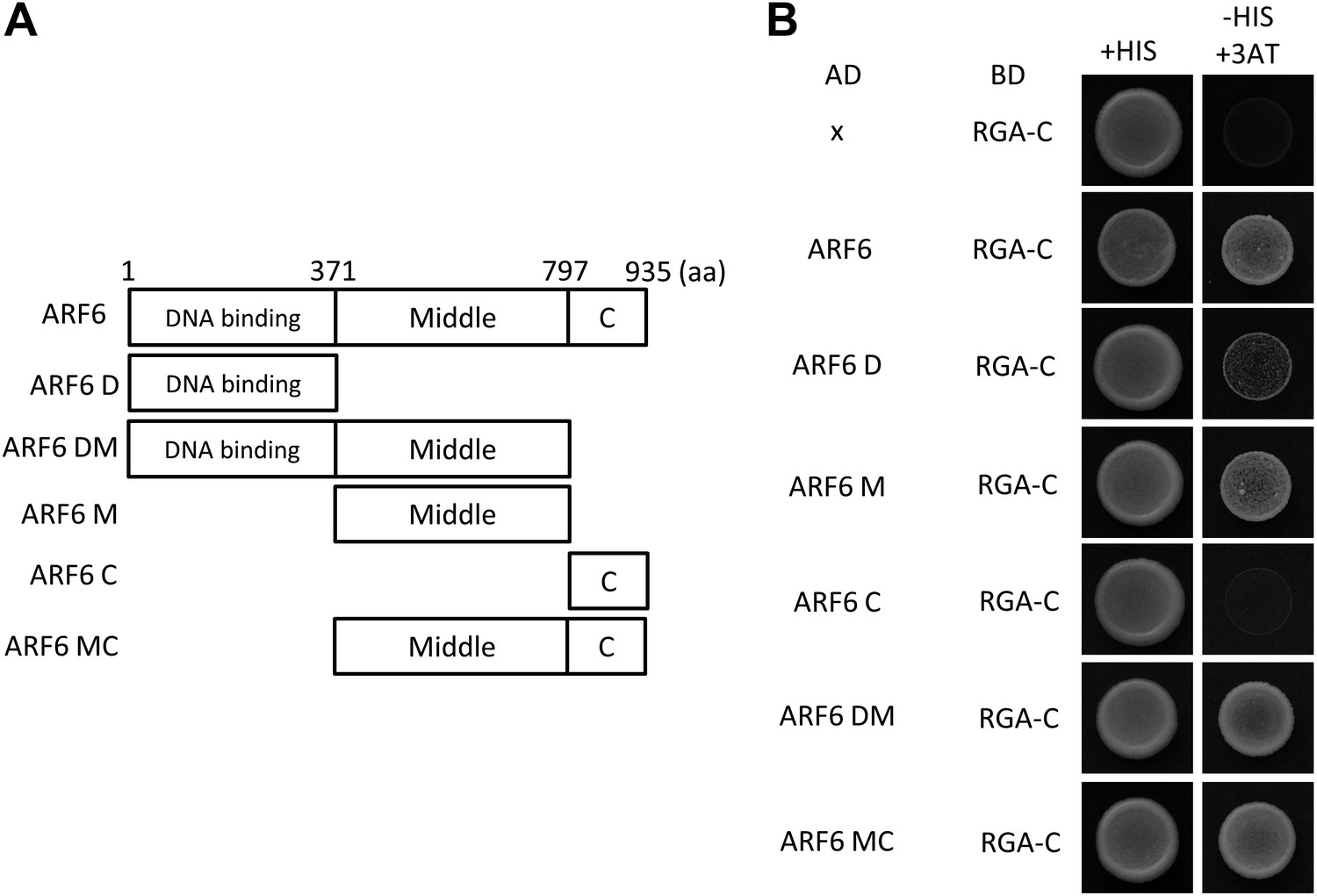
(A) Box diagram of various fragments of ARF6 used in the yeast-two hybrid assay. (B) RGA interacts strongly with a middle domain of ARF6 (ARF6 M) and interacts weakly with a DNA binding domain of ARF6 (ARF6 D). RGA with deletion of N-terminal 208 amino acids (RGA-C) was used for the assay. Yeast clones were grown on the synthetic dropout (+HIS) or synthetic dropout without histidine (−HIS) plus 1 mM 3AT medium. AD: activation domain fusion vector, BD: DNA binding domain fusion vector, x: empty vector.
Figure 6—figure supplement 2
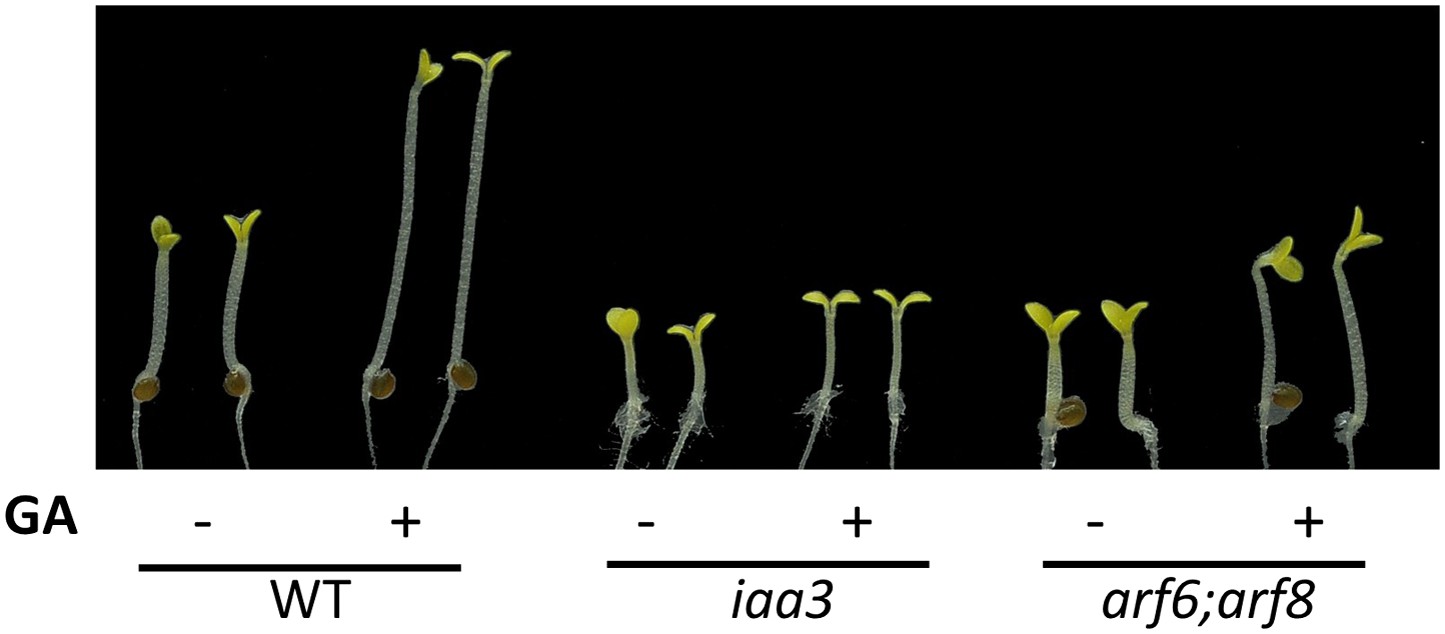
Auxin signaling mutants are less sensitive to GA.
Seedlings were grown on the 10 μM paclobutrazole with or without 1 μM GA in the dark. Representative seedlings are shown.
Figure 7
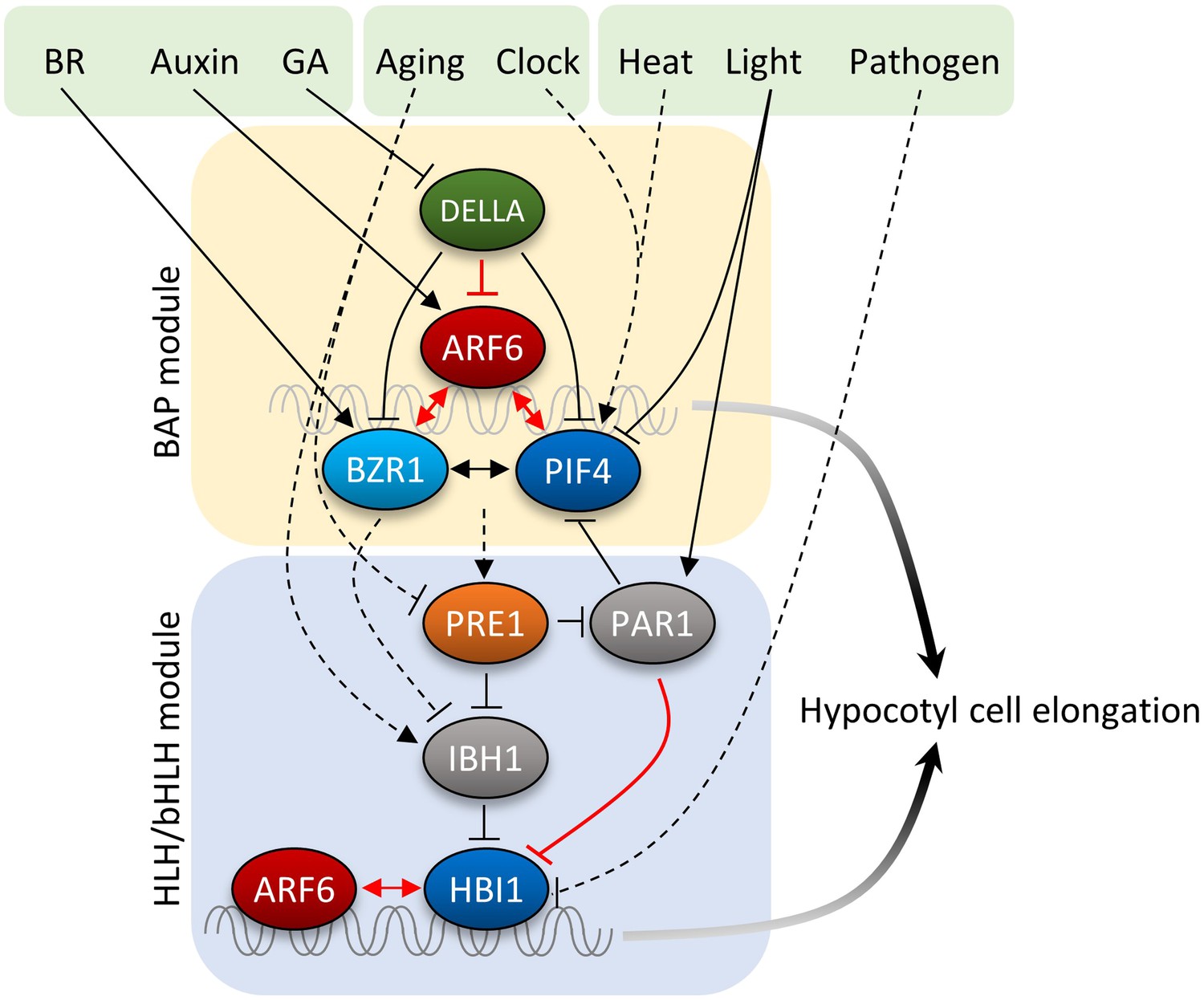
Diagram of the central growth regulation circuit.
In the diagram, solid lines indicate protein–protein interaction or post-translational modification, and dashed lines indicate transcriptional regulation. Red lines indicate new discoveries made in this study. In the BAP module, all three transcription factors, BR-regulated BZR1, auxin-regulated ARF6, and light/temperature-regulated PIF4, interact with each other and cooperatively regulate shared target genes and hypocotyl cell elongation. GA-regulated DELLA interacts with all BAP transcription factors and inhibits their DNA binding. Downstream of BAP module, the HLH/bHLH module, consisting of PRE1, IBH1/PAR1 and HBI1/PIF, modulates BAP activities through HLH–bHLH interactions. The BAP transcription factors positively regulate PRE1 in the HLH/bHLH module forming positive feedback loops. Development and pathogen signals are integrated into the central growth regulation network through PRE1/IBH1 and HBI1, respectively.
Additional files
-
Supplementary file 1
Primer list for qRT-PCR, ChIP-PCR and DNA pull-down assays.
- https://doi.org/10.7554/eLife.03031.023
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl
eLife 3:e03031.
https://doi.org/10.7554/eLife.03031




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}