Neurotrophin-3 regulates ribbon synapse density in the cochlea and induces synapse regeneration after acoustic trauma
- Boston Children's Hospital, United States
- Harvard Medical School, United States
- University of Michigan, United States
- Massachusetts Eye and Ear Infirmary, United States
Figures
Figure 1 with 1 supplement
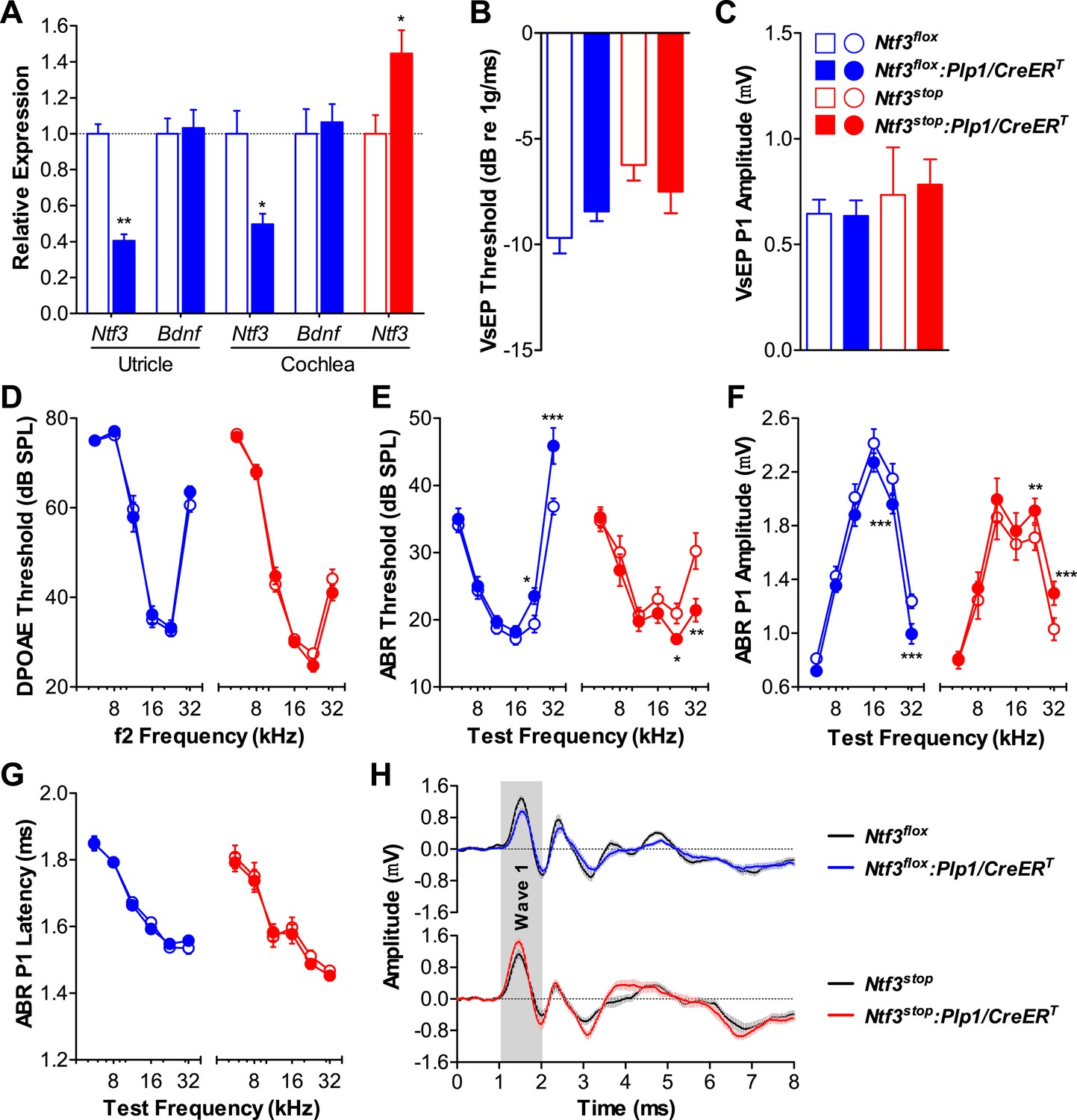
Ntf3 expression by postnatal supporting cells is required for cochlear, but not vestibular function.
(A) RT-qPCR shows that postnatal tamoxifen injection reduces or increases Ntf3 mRNA levels in adult Ntf3flox:Plp1/CreERT or Ntf3stop:Plp1/CreERT inner ears, respectively; n = 5–6. *p < 0.05, **p < 0.01 by two-tailed unpaired t test. (B and C) Postnatal Ntf3 knockout (blue) or overexpression (red) from supporting cells does not alter VsEP thresholds (B) or their peak 1 (P1) amplitudes at 0 dB (C); n = 4–8. (D–F) Postnatal knockout or overexpression of Ntf3 from supporting cells reduces or enhances cochlear function, respectively. Ntf3 knockout (blue) elevates ABR thresholds (E) and decreases ABR P1 amplitudes (F), without changing DPOAE thresholds (D); n = 16–17. Ntf3 overexpression (red) reduces ABR thresholds (E) and increases ABR P1 amplitudes (F), without changing DPOAE thresholds (D); n = 21. ABR P1 amplitudes were assessed at 70 dB SPL. *p < 0.05, **p < 0.01, ***p < 0.001 by two-way ANOVA. (G) ABR P1 latencies are not affected by either Ntf3 knockout (blue) or Ntf3 overexpression (red) at all frequencies examined. Key in C applies to A–G. (H) Mean ABR waveforms from responses to 32 kHz tone pips from Ntf3 knockouts and their controls (upper) and Ntf3 overexpressors and their controls (lower). Gray shading indicates ABR wave 1. Both ABR P1 latencies (G) and waveforms (H) results were assessed at 70 dB SPL; n = 13–17.
Figure 1—figure supplement 1
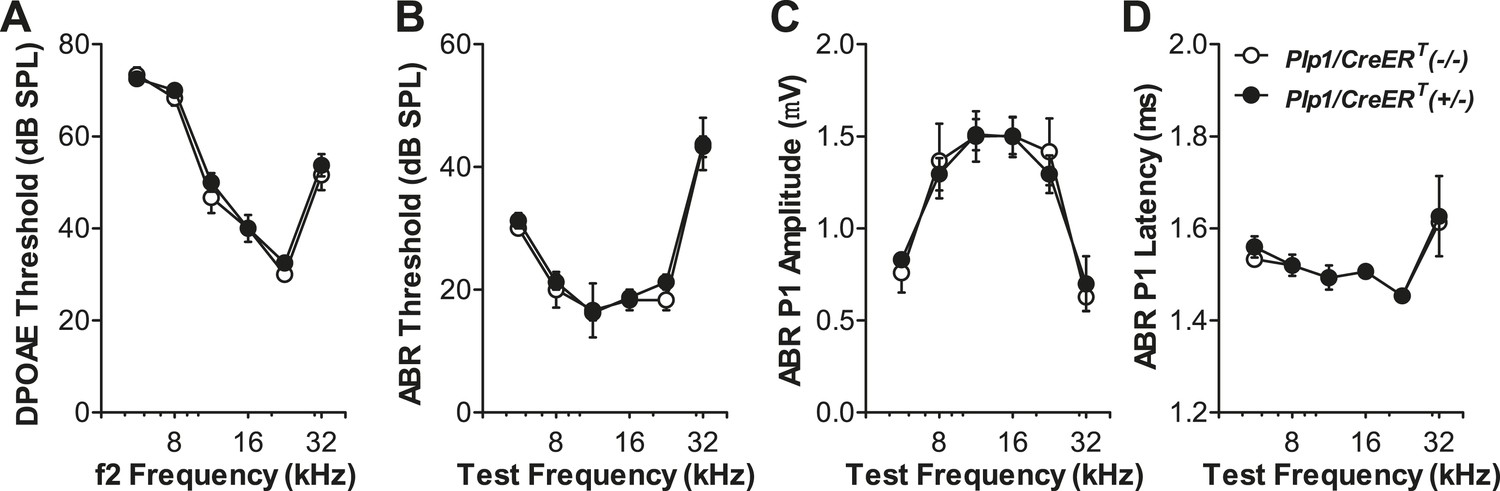
Expression of the Plp1/CreERT allele does not affect the cochlear function.
Plp1/CreERT(+/−) and Plp1/CreERT(−/−) littermates exhibit similar DPOAE thresholds (A), ABR thresholds (B), ABR P1 amplitudes (C), and ABR P1 latencies (D) at all frequencies tested. ABR P1 amplitudes and latencies were assessed at 70 dB SPL. Tamoxifen was injected at P0-1 and physiological tests were performed at P42; n = 4. Key in D applies to all panels.
Figure 2
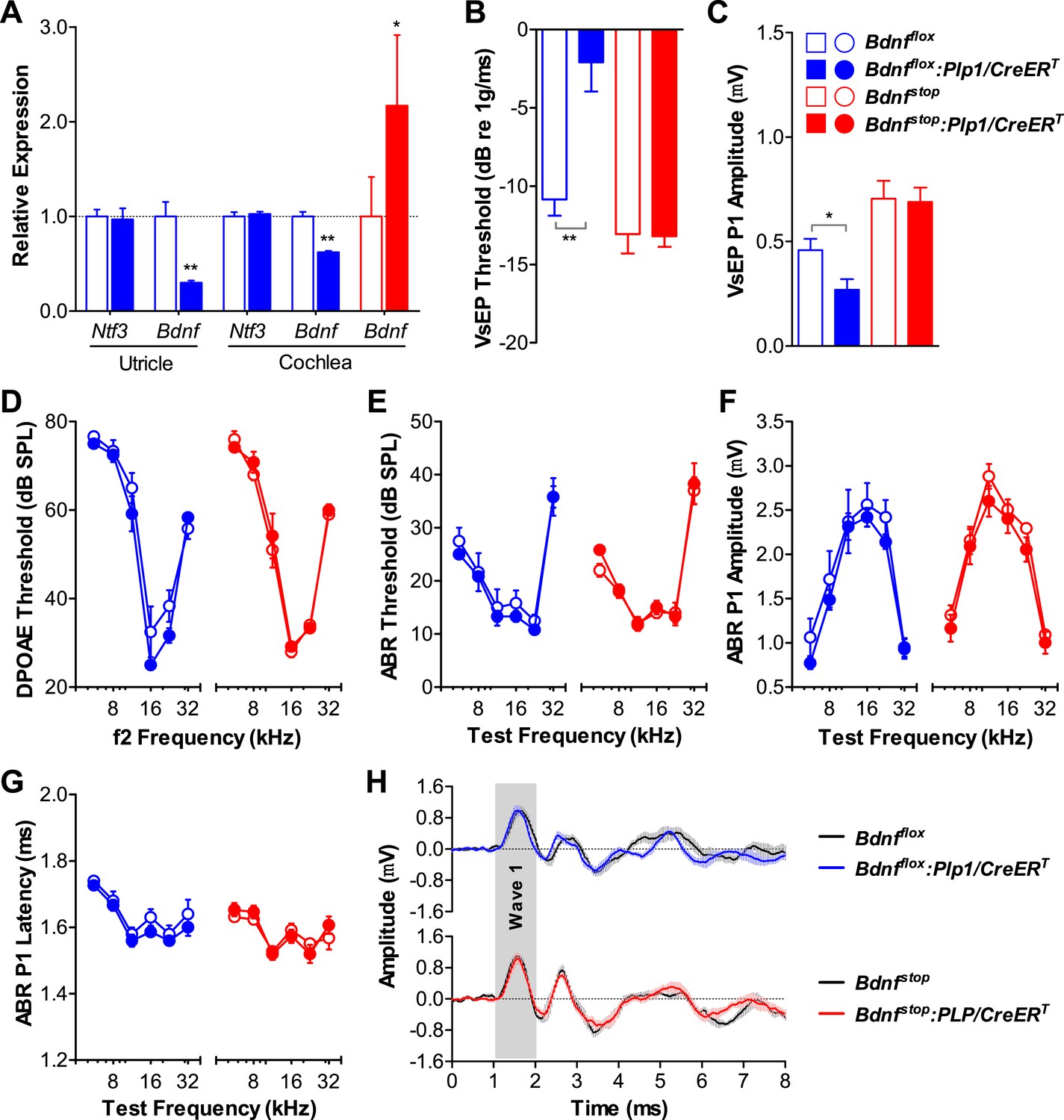
Bdnf expression by postnatal supporting cells is required for vestibular, but not cochlear function.
(A) RT-qPCR shows that postnatal tamoxifen injection results is reduced or increased Bdnf mRNA levels in adult Bdnfflox:Plp1/CreERT or Bdnfstop:Plp1/CreERT inner ears, respectively; n = 6. (B and C) Postnatal Bdnf knockout (blue) from supporting cells leads to higher VsEP thresholds (B) and lower VsEP P1 amplitudes (C) at the high stimulus level (0 dB re 1 g/ms). Postnatal Bdnf overexpression (red) does not affect the vestibular function (B and C); n = 6–11. *p < 0.05; **p < 0.01 by two-tailed unpaired t tests. (D–H) Neither Bdnf knockout (blue) nor overexpression (red) from postnatal supporting cells alters cochlear function, as shown by normal DPOAE thresholds (D), ABR thresholds (E), ABR P1 amplitudes (F), ABR P1 latencies (G), and ABR P1 waveforms (H) at 70 dB SPL compared to control littermates; n = 6. Key in C applies to A–G. Gray shading (H) indicates ABR wave 1.
Figure 3
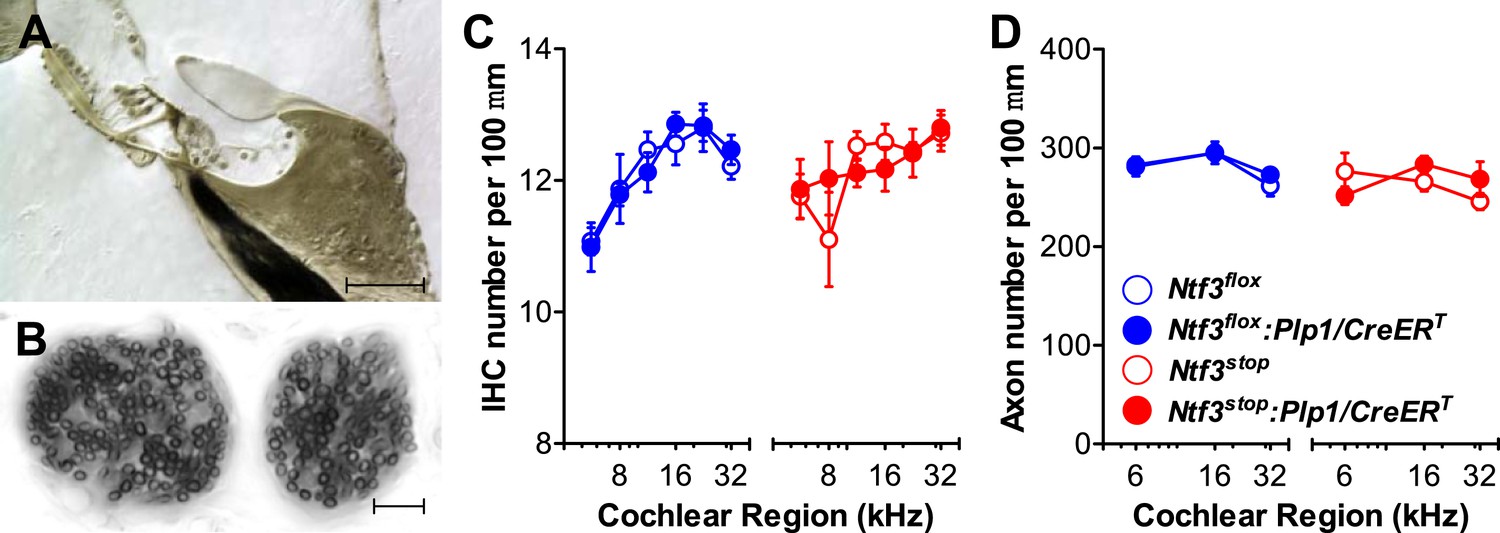
Ntf3 expression by postnatal supporting cells does not affect the organ of Corti morphology, hair cell or axon numbers.
(A and B) Photomicrographs of the basal turn of an Ntf3flox:Plp1/CreERT cochlea (∼32 kHz region) showing a cross-section of the organ of Corti (A; scale bar = 100 µm) and a tangential section through the osseous spiral lamina showing peripheral axons of cochlear nerve fibers (B; scale bar = 10 µm). (C) The number of IHCs per 100 µm of organ of Corti is not altered by either Ntf3 knockout (blue) or overexpression (red) at any cochlear regions; n = 5–6. (D) The number of cochlear-nerve peripheral axons per 100 µm of osseous spiral lamina (near the habenula perforata) is not affected by either Ntf3 knockout (blue) or overexpression (red); n = 6. Key in D applies to C and D.
Figure 4
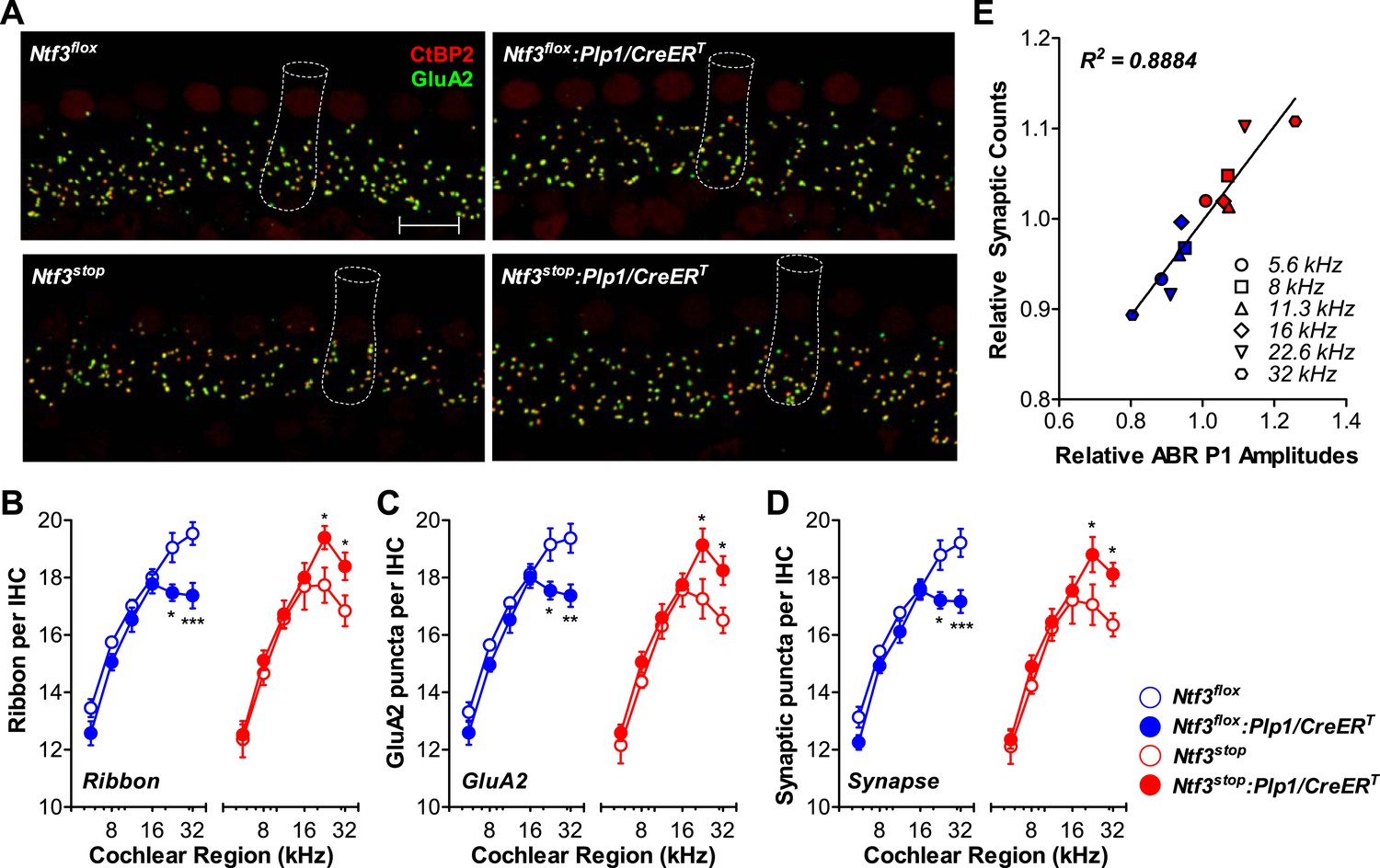
Ntf3 expression by postnatal supporting cells regulates hair cell ribbon synapse density at high frequencies.
(A) Representative confocal images (maximal projection from a focal series) of IHC synapses from 32 kHz region of Ntf3flox, Ntf3flox:Plp1/CreERT, Ntf3stop, and Ntf3stop:Plp1/CreERT cochleae immunolabeled for pre-synaptic ribbons (CtBP2-red) and post-synaptic receptor patches (GluA2-green) (scale bar = 10 µm). The dashed lines show the approximate outline of one IHC. CtBP2 antibody also weakly stains IHC nuclei. (B–D) Quantitative data shows that Ntf3 knockout reduces, and overexpression increases, the number of pre-synaptic ribbons (B), post-synaptic GluA2 receptor patches (C), and putative ribbon synapses, defined as juxtaposed CtBP2- and GluA2-positive puncta (D) at high frequency cochlear regions; n = 5–6. *p < 0.05, **p < 0.01, ***p < 0.001 by two-way ANOVA. (E) Relative synaptic counts vs relative ABR P1 amplitudes of Ntf3 knockouts (blue) or overexpressors (red) shows a linear correlation. Data points were obtained by normalizing synaptic counts and ABR P1 amplitudes of Ntf3 mutants to the values of their respective controls at each of the frequency regions analyzed. Key in D applies B–E.
Figure 5
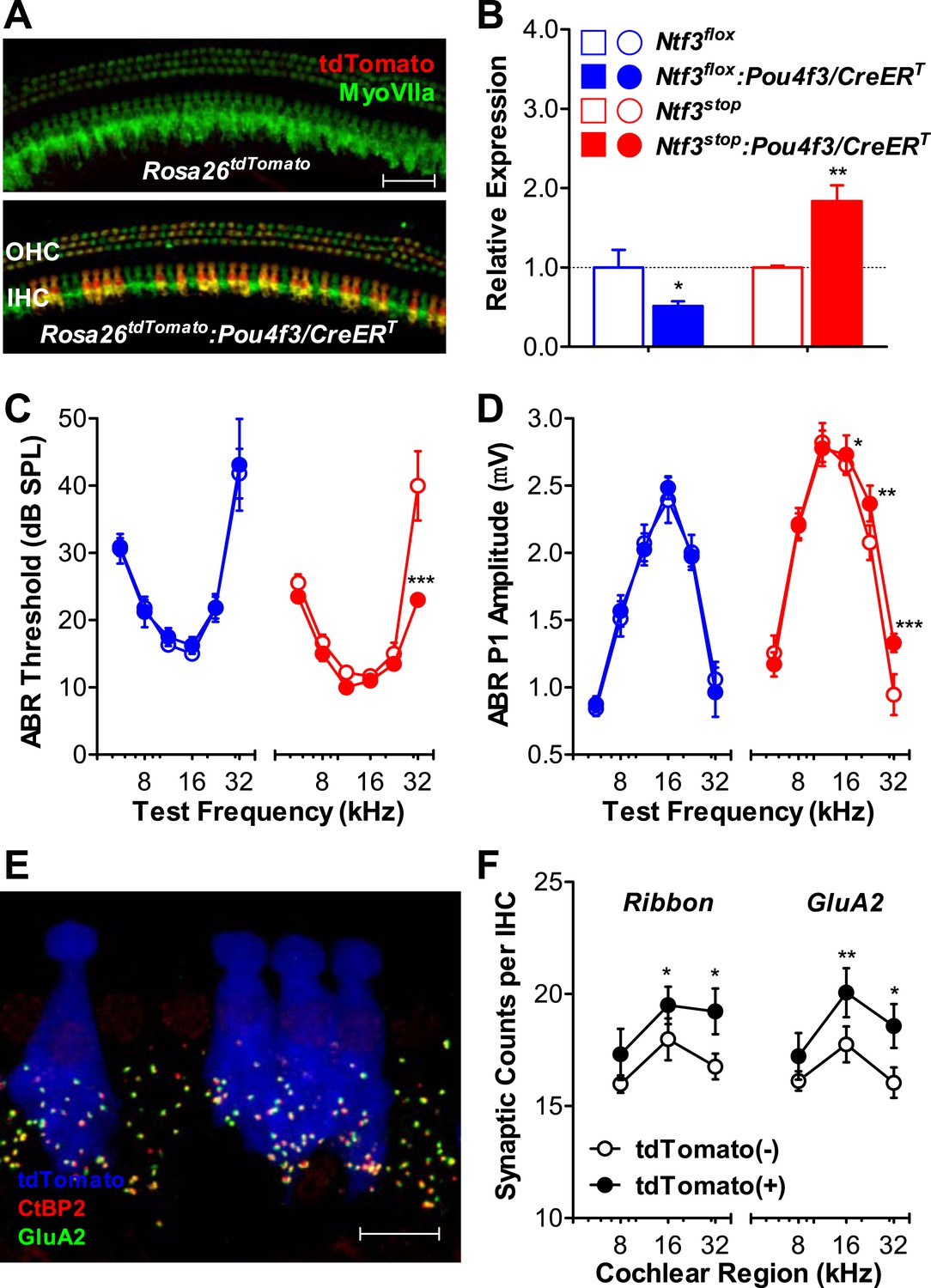
Ntf3 overexpression by postnatal hair cells increases cochlear sensitivity and synaptic densities at high frequencies.
(A) Pou4f3/CreERT allows for hair cells specific inducible gene recombination. Rosa26tdTomato:Pou4f3/CreERT mice and their littermate Rosa26tdTomato controls were injected with tamoxifen at P1-P3, and the cochleas were collected at P60. Inducible recombination is seen as tdTomato fluorescence co-localized with MyoVIIa immunostaining. Scale bar = 50 µm. (B) RT-qPCR shows that postnatal tamoxifen injection reduced Ntf3 mRNA in Ntf3flox:Pou4f3/CreERT cochlea and increased Ntf3 expression in Ntf3stop:Pou4f3/CreERT cochlea, compared to their respective controls; n = 4–5. *p < 0.05, **p < 0.01 by two-tailed unpaired t tests. (C and D) Postnatal overexpression of Ntf3 in hair cells (red) reduced ABR thresholds (C) and increased ABR P1 amplitudes (D) at high frequencies; n = 9–10. Postnatal knockout of Ntf3 from hair cells (blue) had no effect on these measures; n = 8–11. ABR P1 amplitudes were assessed at 70 dB SPL. *p < 0.05, **p < 0.01, ***p < 0.001 by two-way ANOVA. (E) Confocal maximal projection of 7 adjacent IHCs from the 32 kHz region of a hair cell-specific Ntf3 overexpressor, after immunostaining for synapses as in Figure 4A; tdTomato indicates recombined hair cells. Scale bar = 10 µm. (F) Synaptic counts are increased in recombined (tdTomato+, Ntf3 overexpressing) IHCs compared to neighboring unrecombined cells; n = 6 cochleae, with at least 100 hair cells in each group. *p < 0.05, **p < 0.01 by two-tailed paired t tests. Key in B applies to B–D.
Figure 6
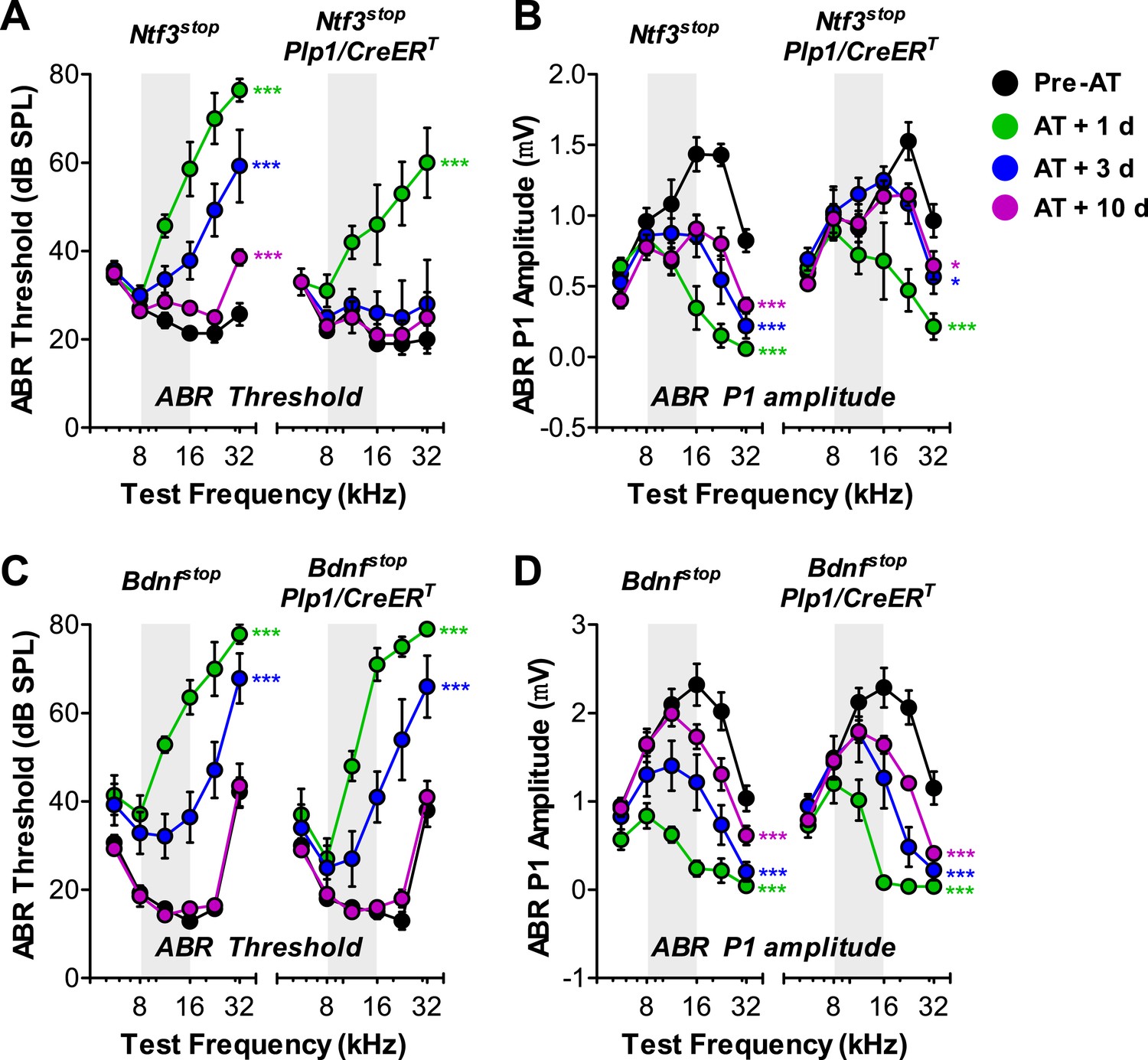
Overexpression of Ntf3, but not Bdnf, promotes recovery from noise-induced attenuation of cochlear responses.
(A and B) Ntf3 overexpression accelerates the recovery of ABR thresholds (A) and promotes recovery of ABR P1 amplitudes (B) after acoustic trauma (AT); n = 5–7. (C and D) Bdnf overexpression does not affect the recovery of ABR thresholds (A) and ABR P1 amplitudes (B) after acoustic trauma; n = 5–7. *p < 0.05, ***p < 0.001 by two-way ANOVA. Gray shading indicates the noise exposure spectrum. Key in B applies to all panels.
Figure 7
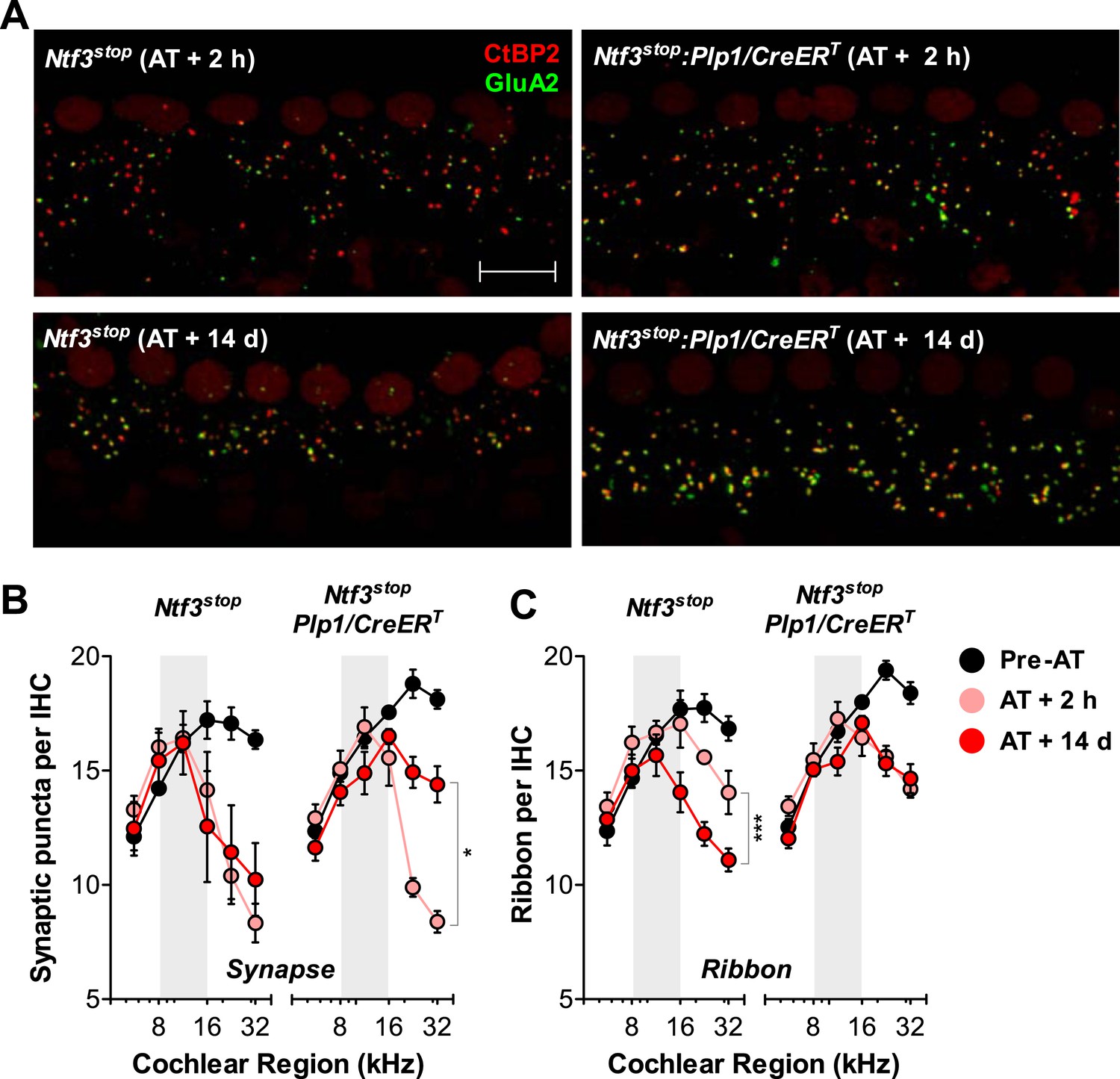
Ntf3 overexpression from postnatal supporting cells promotes recovery from noise-induced synaptic degeneration.
(A) Representative confocal images of IHC synapses from 32 kHz region of Ntf3stop and Ntf3stop:Plp1/CreERT cochleae immunolabeled for CtBP2 and GluA2 at 2 hr (upper panels) or 14 days after acoustic trauma (AT); scale bar = 10 µm. (B and C) Ntf3 overexpression promotes regeneration of IHC synapses (B) and prevents the progressive loss of IHC ribbons (C) between 2 hr (AT + 2 hr) and 14 days (AT + 14 days) after acoustic trauma. n = 3–8. *p < 0.05, ***p < 0.001 by two-way ANOVA. Gray shading indicates the noise exposure spectrum. Key in C applies to B–C.
Figure 8
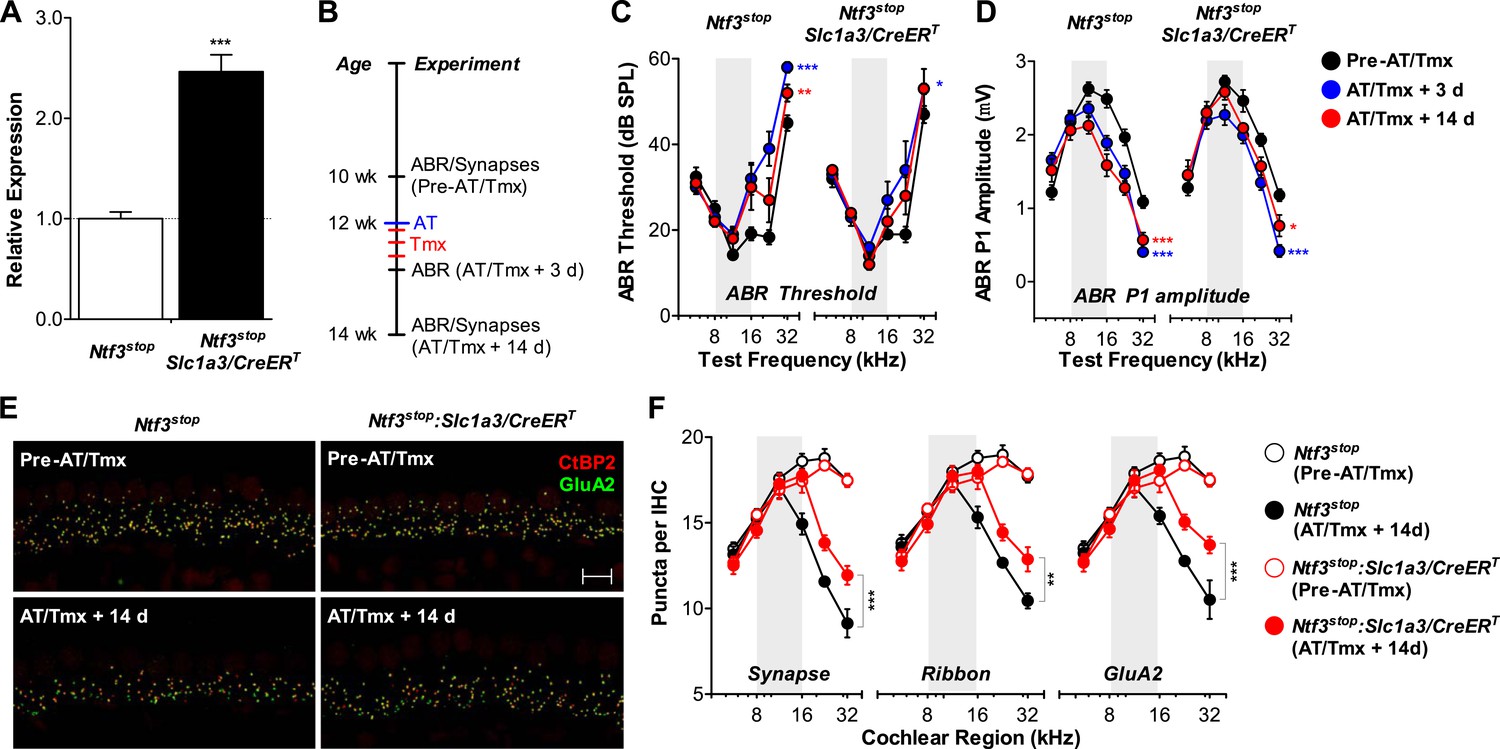
Ntf3 overexpression from adult supporting cells after acoustic trauma promotes auditory function recovery and synaptic regeneration.
(A) RT-qPCR shows that tamoxifen treatments of adult mice increased Ntf3 expression in Ntf3stop:Slc1a3/CreERT cochlea; n = 6. ***p < 0.001 by two-tailed unpaired t tests. (B) Time line of the experiment showing the ages of mice for ABR measurements, acoustic trauma (AT), tamoxifen inductions (Tmx), and sample collections for synaptic counts. (C–D) The effects of Ntf3 overexpression from adult supporting cells on ABR thresholds (C) and P1 amplitudes (D) after noise exposure; n = 5–9. *p < 0.05, **p < 0.01, ***p < 0.001 by two-way ANOVA. Key in D applies to C–D. (E) Representative confocal images of IHC synapses from 32 kHz region of Ntf3stop and Ntf3stop:Slc1a3/CreERT cochleae immunolabeled for CtBP2 and GluA2. The samples were collected from mice without AT/Tmx (Pre-AT/Tmx) or 14 days after AT/Tmx (AT/Tmx + 14 days). Scale bar = 10 µm. (F) Ntf3 overexpression after acoustic trauma promotes regeneration of IHC ribbon synapses; n = 5–6. **p < 0.01, ***p < 0.001 by two-way ANOVA. Gray shading indicates the noise exposure spectrum.
Author response image 1

Ribbon synaptic density of C57BL/6J and FVB/N mouse strains. (A) Representative confocal images (maximal projection from a focal series) of IHC synapses from 32 kHz region of 8 wk-old C57BL/6J and FVB/N cochleae immunolabeled for pre-synaptic ribbons (CtBP2-red) and post-synaptic receptor patches (GluA2-green) (scale bar = 10 µm). (B) Synaptic counts of C57BL/6J and FVB/N cochleae at 32 kHz; n = 4-6. *p<0.05 by unpaired student’s t-test.
Tables
Table 1
Statistical analysis (two-way ANOVA) of ABR threshold and P1 amplitude changes between control and neurotrophin overexpressing (Plp1/CreERT) mice after acoustic trauma
| Ntf3 overexpressor vs control | Bdnf overexpressor vs control | ||||
|---|---|---|---|---|---|
| p Value | F Statistic | p Value | F Statistic | ||
| ABR threshold | AT + 1 day | 0.0051 | 8.469 | 0.6801 | 0.1716 |
| AT + 3 days | <0.0001 | 19.00 | 0.6752 | 0.1774 | |
| AT + 10 days | <0.0001 | 17.34 | 0.9862 | 0.0003 | |
| ABR P1 amplitude | AT + 1 day | 0.0342 | 4.695 | 0.1899 | 1.7580 |
| AT + 3 days | <0.0001 | 18.08 | 0.5692 | 0.3276 | |
| AT + 10 days | <0.0001 | 18.13 | 0.0421 | 4.3160 | |
Table 2
Statistical analysis (two-way ANOVA) of synaptic density changes between control and Ntf3 overexpressing (Plp1/CreERT) mice after acoustic trauma
| p Value | F Statistic | ||
|---|---|---|---|
| Synapse | AT + 2 hr | 0.9732 | 0.001 |
| AT + 14 days | 0.0065 | 8.358 | |
| Ribbon | AT + 2 hr | 0.8134 | 0.056 |
| AT + 14 days | 0.0002 | 15.54 | |
Table 3
Statistical analysis (two-way ANOVA) of ABR threshold and P1 amplitude changes between control and Ntf3 overexpressing (Slc1a3/CreERT) mice before and after acoustic trauma and tamoxifen treatments
| p Value | F Statistic | ||
|---|---|---|---|
| ABR threshold | Pre-AT/Tmx | 0.7774 | 0.081 |
| AT/Tmx + 3 days | 0.1674 | 1.965 | |
| AT/Tmx + 14 days | 0.5301 | 0.400 | |
| ABR P1 amplitude | Pre-AT/Tmx | 0.2826 | 1.166 |
| AT/Tmx + 3 days | 0.3695 | 0.821 | |
| AT/Tmx + 14 days | 0.0008 | 12.71 | |
Additional files
-
Source code 1
Amira and Blob Projection software.
- https://doi.org/10.7554/eLife.03564.015
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Neurotrophin-3 regulates ribbon synapse density in the cochlea and induces synapse regeneration after acoustic trauma
eLife 3:e03564.
https://doi.org/10.7554/eLife.03564




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}