The evolutionary modifications of a GoLoco motif in the AGS protein facilitate micromere formation in the sea urchin embryo
- Department of Molecular Biology, Cellular Biology, Biochemistry, Brown University, United States
Figures
Figure 1
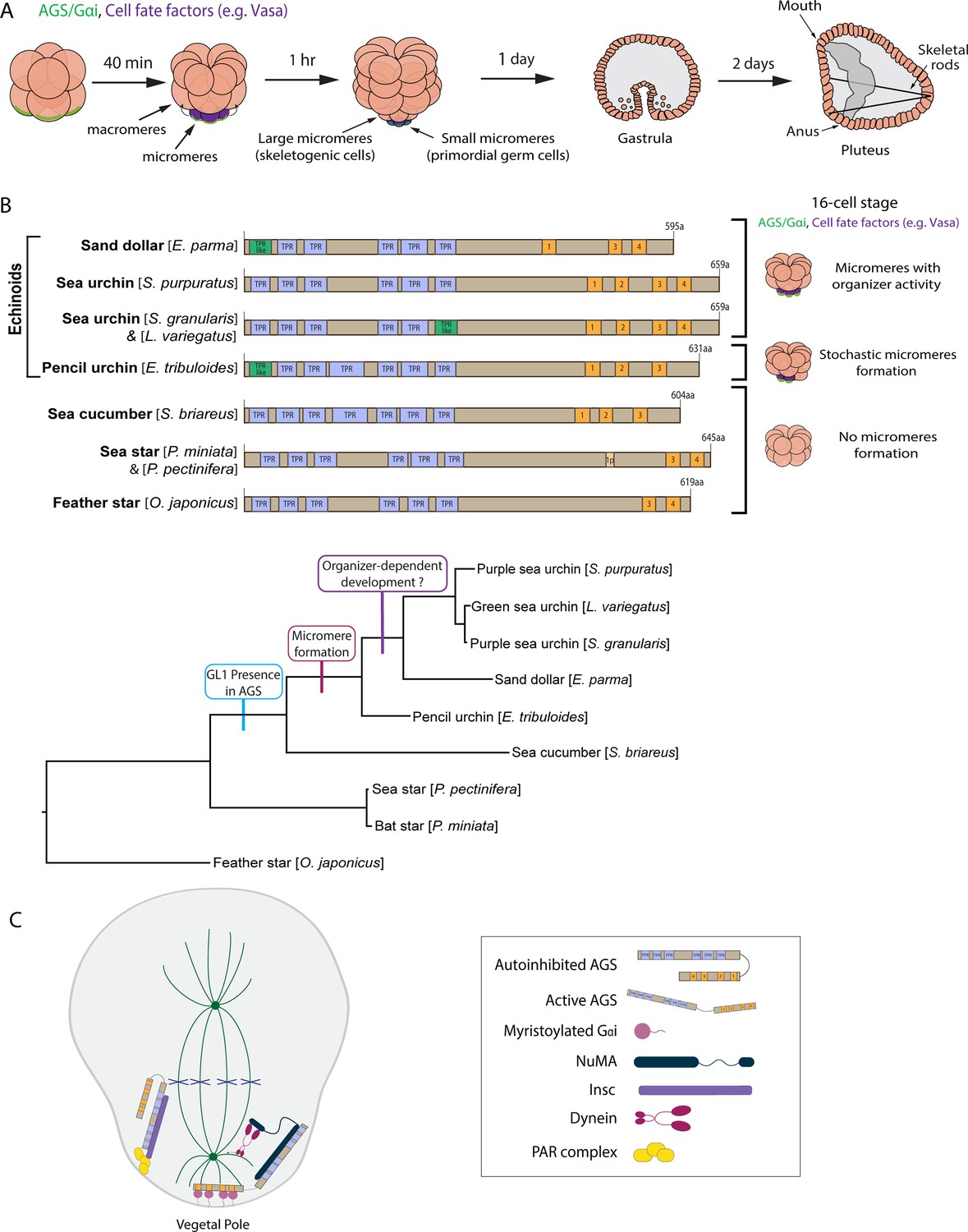
The evolutionary modification of the SpAGS protein corresponds to the introduction of micromeres and inductive signaling during echinoid diversification.
(A) Schema depicting sea urchin embryonic development from eight-cell stage to pluteus. Green represents the colocalization of AGS and Gαi at the vegetal cortex, and purple represents the early segregation of fate factors such as Vasa. (B) Comparative diagrams of predicted motifs of each echinoderm AGS protein, based on NCBI blast search results for AGS sequences. Conserved TPR motifs are indicated in blue, and GL motifs in orange. Green shows TPR-like motifs, which contain several amino acid changes. Lighter colors represent partial GL motifs. See Figure 2—figure supplement 2 for each echinoderm AGS sequence. The tree depicts SpAGS evolution among echinoderms with the introduction of the GL1 motif and micromeres. (C) Working model of AGS mechanism in asymmetric cell division (ACD) based on fly and mammalian models.
Figure 2 with 2 supplements
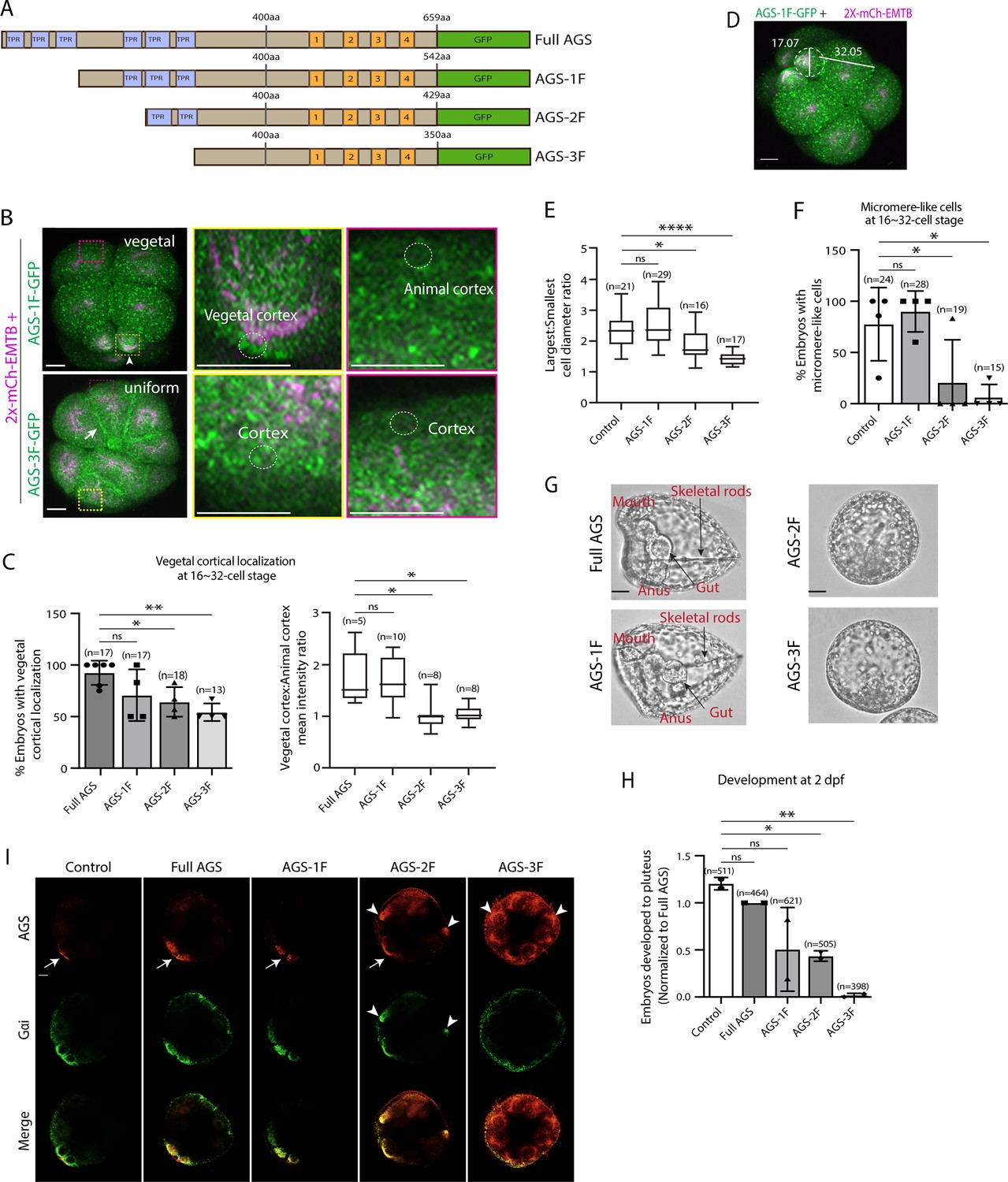
The N-terminal TPR domain restricts SpAGS localization and function at the vegetal cortex.
(A) Design of SpAGS-GFP N-terminal deletion constructs used in this study. TPR motifs are marked in blue, and GL motifs are in orange. (B) Representative 2D-projection images of the embryo injected with AGS-1F-GFP or AGS-3F-GFP and 2x-mCherry-EMTB, exhibiting vegetal (upper panel, arrowhead) and uniform (lower panel, arrow) cortical localization. The magnified images next to each panel demonstrate how we measured the mean intensities of the vegetal cortex (yellow squared) and animal cortex (magenta squared) using ImageJ. The results of the analysis are summarized in the corresponding graph (C). Embryos were injected with 0.15–0.3 μg/μl stock of SpAGS-GFP mRNA and 0.5 μg/μl stock of 2x-mCherry-EMTB mRNA. Z-stack images were taken at 1 μm intervals to cover a layer of the embryo. (C) Percentage of the embryos with vegetal cortical localization of SpAGS (left) and the ratio of the vegetal cortex-to-animal cortex mean intensity (right) at 16–32-cell embryos. Statistical analysis was performed against Full AGS by one-way ANOVA. (D) Representative 2D-projection confocal image of a 16-cell stage embryo injected with AGS-1F-GFP. The largest cell (macromere) and the smallest cell (micromere) diameters were measured using ImageJ. Z-stack images were taken at 1 μm intervals to cover a layer of the embryo. (E) The diameter ratio of the smallest cell (micromere-like cell) over the largest cell (macromere-like cell) was quantified for the embryos injected with the SpAGS mutants or EMTB-only (control). (F) Percentage of the embryos forming micromere-like cells was scored for each SpAGS mutant and EMTB-only (control). ‘Micromere formation’ is defined as the formation of a group of four cells that are smaller in size and made through a vertical cell division at the vegetal pole at the 16-cell stage. Since none of the AGS-3F-injected embryos formed normal micromeres, ‘micromere-like cells’ were counted based on their vertical cell division, not relative to their size. Statistical analysis was performed against control by one-way ANOVA. (G, H) Brightfield images show the representative phenotypes scored in the corresponding graph (H) at 2 dpf. We categorized embryos into three groups, namely, ‘full development’, with embryos reaching the pluteus stage with complete gut formation and skeleton; ‘delayed development’, with some gastrulation but no proper skeleton; and ‘failed gastrulation’. As many of the abnormal-looking embryos fell into the median of the latter two categories, we scored only the embryos reaching full development in the graph. Control represents embryos injected with a RITC dye only. Statistical analysis was performed against control by one-way ANOVA. (I) Single Z-slice confocal imaging was used to focus on the vegetal cortex. Embryos were stained with AGS (orange) and Gɑi (green) antibodies. White arrows and arrowheads indicate the signals at the vegetal cortex and ectopic cortical signals, respectively. Images represent over 80% of the embryos observed (n = 30 or larger) per group. n indicates the total number of embryos scored. *p<0.05, **p<0.01, and ****p<0.0001. Each experiment was performed at least three independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 2—source data 1
Numerical data for Figure 2C.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig2-data1-v1.xlsx
-
Figure 2—source data 2
Numerical data for Figure 2E.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig2-data2-v1.xlsx
-
Figure 2—source data 3
Numerical data for Figure 2F.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig2-data3-v1.xlsx
-
Figure 2—source data 4
Numerical data for Figure 2H.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig2-data4-v1.xlsx
Figure 2—figure supplement 1
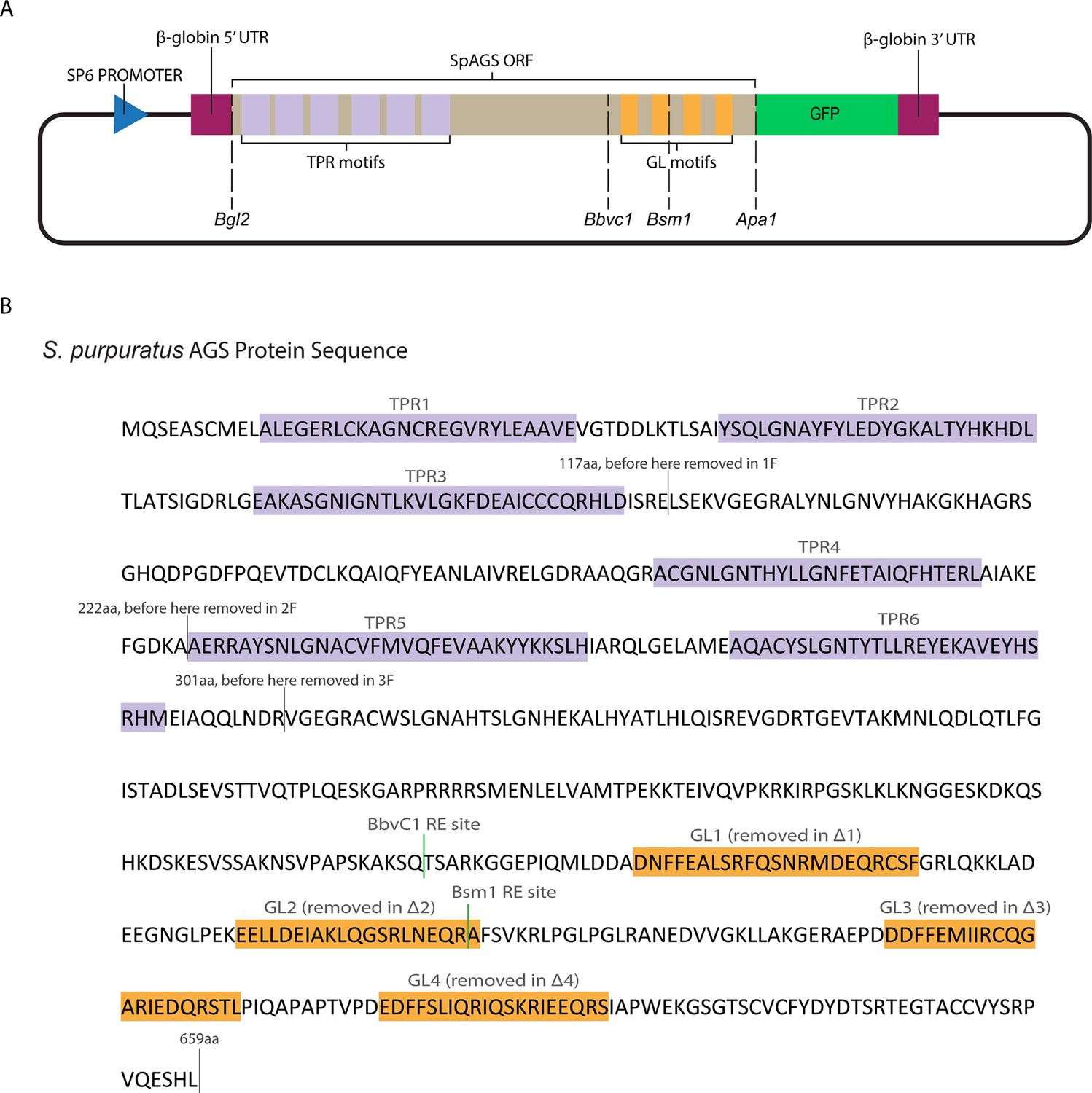
Dissection of SpAGS motifs.
(A) The construct design of SpAGS-GFP, including the restriction enzyme sites used to prepare SpAGS mutants. (B) The protein sequence of SpAGS. Predicted domains are labeled based on NCBI blast results, and the sequence portions deleted for each N-terminal construct are marked. The sequences for each GL motif used for deletion or swapping are indicated in orange. The internal restriction enzyme sites for BbvCI and BsmI are shown in green.
Figure 2—figure supplement 2

Echinoderm AGS sequence alignment.
All AGS are similar in the N-terminus with the predicted conserved TPR motifs (blue) but are highly variable in the C-terminus with the predicted GL motifs (yellow).
Figure 3
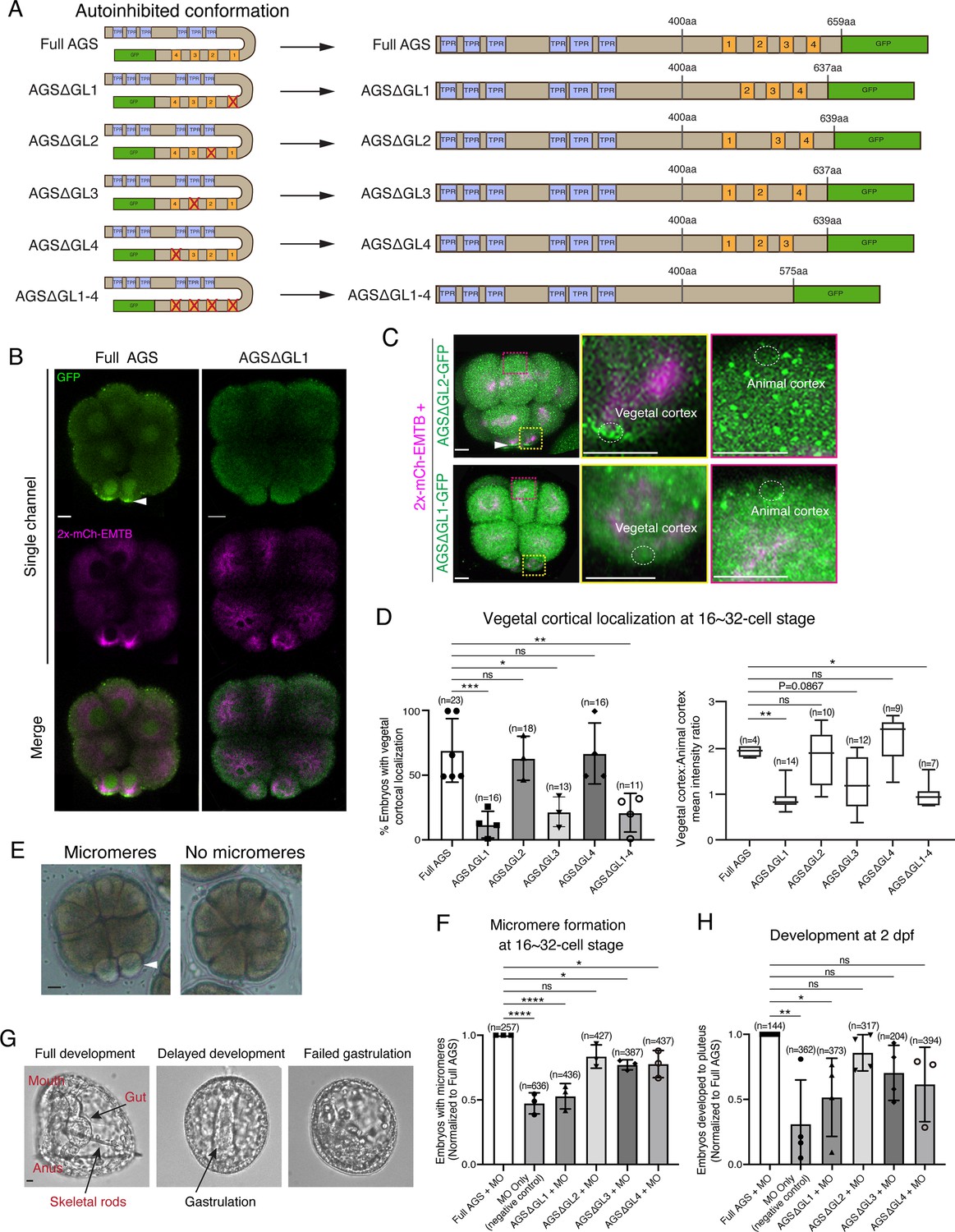
GL1 is essential for vegetal cortical recruitment of SpAGS at the 8–16-cell stage of the sea urchin embryo.
(A) Design of SpAGS-GFP C-terminal deletion mRNAs tested in this study. TPR motifs are marked in blue, and GL motifs are in orange. See Figure 2—figure supplement 2 for protein sequence. (B) Single Z-slice confocal imaging was used to focus on the vegetal cortex. Representative embryos injected with SpAGS-GFP or SpAGSΔGL1-GFP are shown. Embryos were injected with 0.3 μg/μl stock of SpAGS-mutant-GFP mRNA (green) and 0.5 μg/μl stock of 2x-mCherry-EMTB mRNA (magenta). The white arrowhead indicates vegetal cortical localization of AGS-GFP. (C, D) Representative 2D-projection images of the embryo injected with SpAGS-GFP mRNA and 2x-mCherry-EMTB mRNA, exhibiting vegetal cortical (upper panel, AGSΔGL2, arrowhead) and uniform cytoplasmic (lower panel, AGSΔGL1) localization. The magnified images next to each panel demonstrate how we measured the mean intensities of the vegetal cortex (yellow squared) and animal cortex (magenta squared) using ImageJ. The results of the analysis are summarized in the corresponding graph (D). Z-stack images were taken at 1 μm intervals to cover a layer of the embryo. Percentage of the embryos that had the GFP signal at the vegetal cortex (left) and the ratio of the vegetal cortex-to-animal cortex mean intensity (right) during the 16–32-cell stage were scored in the graphs. Statistical analysis was performed against Full AGS by one-way ANOVA. (E, F) Brightfield images show the representative phenotypes scored in the corresponding graph (F) at the 16-cell stage. White arrowhead shows micromeres. Embryos were injected with 0.15 μg/μl stock of SpAGS-GFP mRNAs and 0.75 mM SpAGS MO. The number of embryos forming micromeres was scored and normalized to that of Full AGS in the graph. Statistical analysis was performed by one-way ANOVA. (G, H) Brightfield images show the representative phenotypes scored in the corresponding graph (H) at 2 dpf. Embryos were injected with 0.15 μg/μl stock of SpAGS-GFP mRNAs and 0.75 mM SpAGS MO. The number of embryos developing to the pluteus stage was scored and normalized to that of Full AGS in the graph. Statistical analysis was performed by one-way ANOVA. n indicates the total number of embryos scored. *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Each experiment was performed at least three independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 3—source data 1
Numerical data for Figure 3D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig3-data1-v1.xlsx
-
Figure 3—source data 2
Numerical data for Figure 3F.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig3-data2-v1.xlsx
-
Figure 3—source data 3
Numerical data for Figure 3H.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig3-data3-v1.xlsx
Figure 4
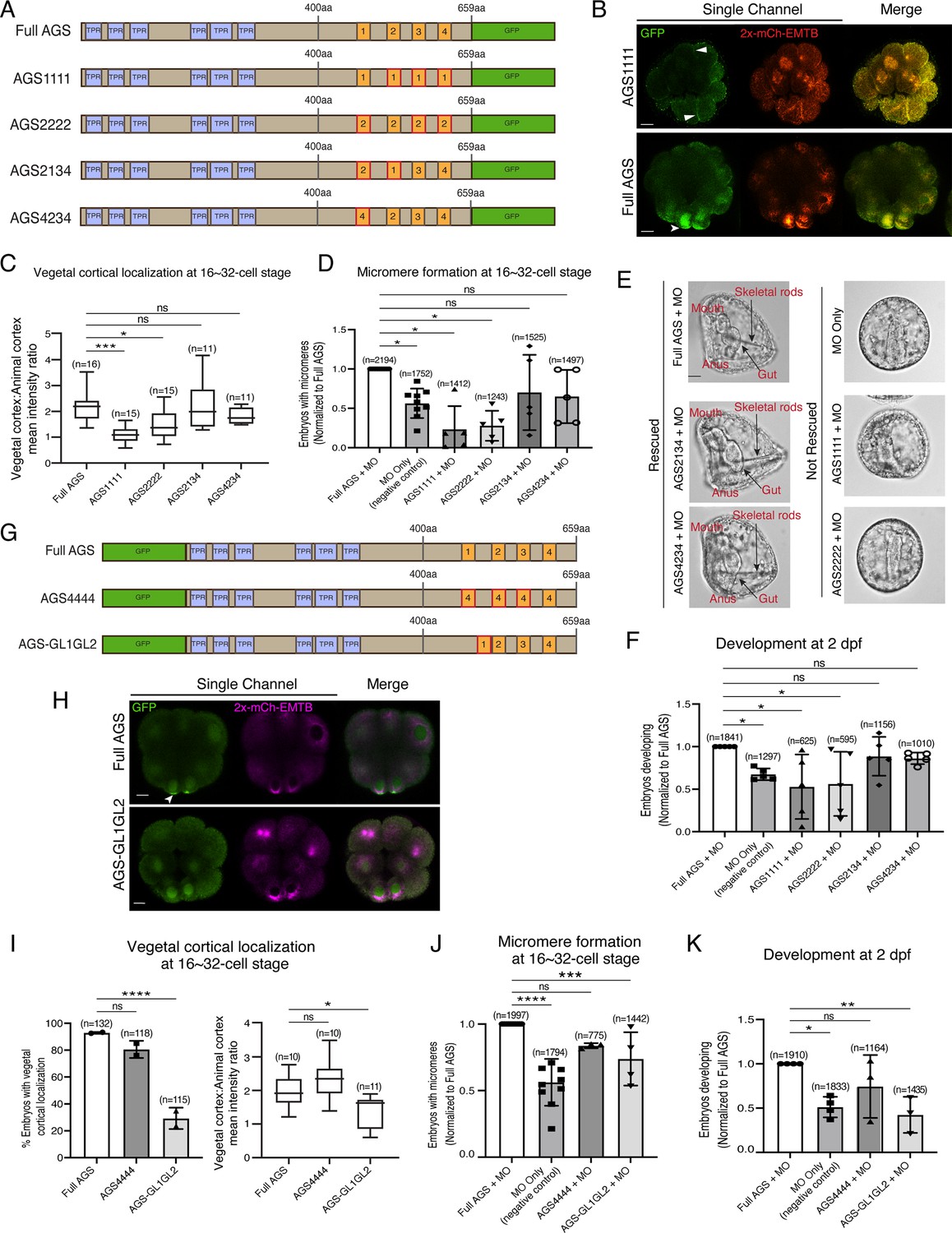
The position of GL1 and the sequences of GL3 and GL4 are important for SpAGS localization and function.
(A) Design of SpAGS-GFP C-terminal mutant constructs tested in this study. TPR motifs are marked in blue, and GL motifs are in orange. Red boxes show interchanged GL motifs. (B) Single Z-slice confocal images of sea urchin embryos at the 8–16-cell stage showing localization of SpAGS1111-GFP mutant. Embryos were injected with 0.3 μg/μl stock of SpAGS-mutant GFP mRNAs and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. White arrowheads indicate vegetal cortical localization of AGS-GFP proteins and ectopic localization of AGS1111 mutant. (C) The ratio of the vegetal cortex-to-animal cortex mean intensity in 16–32-cell embryos. Statistical analysis was performed against Full AGS by one-way ANOVA. (D–F) Embryos were injected with 0.15 μg/μl stock of SpAGS-GFP mRNAs and 0.75 mM SpAGS MO. The number of embryos making micromeres (D) and developing to gastrula or pluteus stage (F) was scored and normalized to that of the Full AGS control group. Brightfield images (E) show the representative phenotypes scored in the corresponding graph (F) at 2 dpf. Of note, AGS1111 and AGS2222 mutants caused substantial toxicity, degrading many embryos by 2 dpf and resulting in inconsistent scoring. Thus, we scored embryos reaching the pluteus stage, which revealed delayed development in this analysis. Statistical analysis was performed by one-way ANOVA. (G) Design of GFP-SpAGS C-terminal mutant constructs tested in this study. In AGS4444, we replaced all GL motifs with GL4. In AGS-GL1GL2, GL1 is shifted adjacent to GL2. TPR motifs are marked in blue, and GL motifs are in orange. Red boxes show modified GL motifs. (H) Single Z-slice confocal images of sea urchin embryos at the 8–16-cell stage showing localization of GFP-SpAGS-GL1GL2 mutant. Embryos were injected with 0.3 μg/μl stock of GFP-SpAGS mRNA and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. The white arrowhead indicates the vegetal cortical localization of GFP-AGS. (I) Percentage of the embryos with vegetal cortical localization of SpAGS mutants (left) and the ratio of the vegetal cortex-to-animal cortex mean intensity (right) in 16–32-cell embryos. Statistical analysis was performed against Full AGS by one-way ANOVA. (J, K) Embryos were injected with 0.15 μg/μl stock of GFP-SpAGS mRNAs and 0.75 mM SpAGS MO. The number of embryos making micromeres (J) and developing to gastrula or pluteus stage (K) was scored and normalized to that of the Full AGS. Statistical analysis was performed by one-way ANOVA. n indicates the total number of embryos scored. *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 4—source data 1
Numerical data for Figure 4C.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig4-data1-v1.xlsx
-
Figure 4—source data 2
Numerical data for Figure 4D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig4-data2-v1.xlsx
-
Figure 4—source data 3
Numerical data for Figure 4F.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig4-data3-v1.xlsx
-
Figure 4—source data 4
Numerical data for Figure 4I.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig4-data4-v1.xlsx
-
Figure 4—source data 5
Numerical data for Figure 4J.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig4-data5-v1.xlsx
-
Figure 4—source data 6
Numerical data for Figure 4K.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig4-data6-v1.xlsx
Figure 5 with 3 supplements
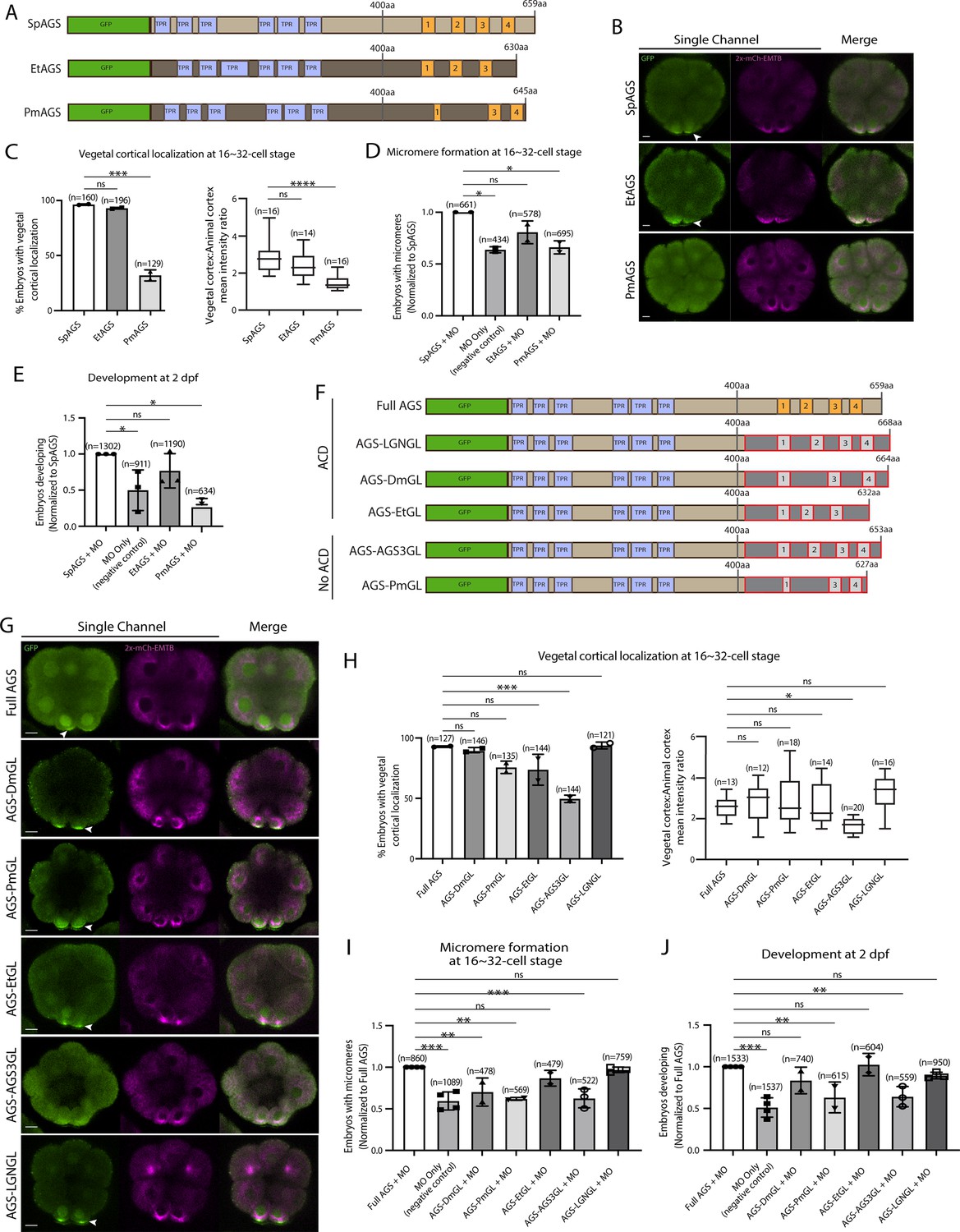
Molecular evolution of AGS C-terminus facilitates micromere formation.
(A) Design of GFP-AGS constructs from three different species tested in this study, namely, S. purpuratus (Sp), E. tribuloides (Et), and P. miniata (Pm). TPR motifs are marked in blue, and GL motifs are in orange. (B) Single Z-slice confocal images of sea urchin embryos at 16-cell stage showing localization of each GFP-AGS. Embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNAs and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. The white arrowhead indicates vegetal cortical localization of GFP-AGS. (C) Percentage of embryos with vegetal cortical localization of GFP-AGS (left) and the ratio of the vegetal cortex-to-animal cortex mean intensity (right) in 16–32-cell embryos. Statistical analysis was performed against SpAGS by one-way ANOVA. (D, E) Embryos were injected with 0.15 μg/μl stock of GFP-AGS mRNAs and 0.75 mM SpAGS MO. The number of embryos making micromeres (D) and developing to pluteus stage (E) was scored and normalized to that of the Full AGS. Statistical analysis was performed by one-way ANOVA. (F) Design of GFP-AGS C-terminal chimeric mutant constructs tested in this study. TPR motifs are marked in blue, and GL motifs are in orange. The brown section shows the SpAGS portion, and the red and dark gray boxes show the non-sea urchin (non-SpAGS) C-terminal sequence introduced. Protein sequences used include Drosophila Pins (Dm), P. miniata AGS (Pm), E. tribuloides AGS (Et), H. sapiens AGS3 (AGS3), and H. sapiens LGN (LGN). (G) Single Z-slice confocal images of sea urchin embryos at the 8–16-cell stage showing localization of each GFP-AGS. Embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNA and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. The white arrowheads indicate vegetal cortical localization of GFP-AGS. (H) Percentage of the embryos with vegetal cortical localization of GFP-AGS chimeric mutants (left) and the ratio of the vegetal cortex-to-animal cortex mean intensity (right) in 16–32-cell embryos. Statistical analysis was performed against Full AGS by one-way ANOVA. (I, J) Embryos were injected with 0.15 μg/μl stock of GFP-AGS mRNAs and 0.75 mM SpAGS MO. The number of embryos making micromeres (I) and developing to gastrula or pluteus stage (J) was scored and normalized to that of the Full AGS. Statistical analysis was performed by one-way ANOVA. n indicates the total number of embryos scored. *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 5—source data 1
Numerical data for Figure 5C.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-data1-v1.xlsx
-
Figure 5—source data 2
Numerical data for Figure 5D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-data2-v1.xlsx
-
Figure 5—source data 3
Numerical data for Figure 5E.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-data3-v1.xlsx
-
Figure 5—source data 4
Numerical data for Figure 5H.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-data4-v1.xlsx
-
Figure 5—source data 5
Numerical data for Figure 5I.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-data5-v1.xlsx
-
Figure 5—source data 6
Numerical data for Figure 4J.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-data6-v1.xlsx
Figure 5—figure supplement 1

SbAGS does not fully localize at the vegetal cortex.
(A) Design of 2x-GFP-AGS constructs that contain AGS orthologs from two different species tested in this study, namely, S. purpuratus (Sp; sea urchin), and S. briareus (Sb; sea cucumber). TPR motifs are marked in blue, and GL motifs are in orange. (B) Alignment of GL1 motif sequences among echinoderms. (C) Single Z-slice confocal images of sea urchin (Sp) or sea cucumber (Sb) embryos at the 16-cell stage showing localization of 2x-GFP-AGS. Embryos were injected with 0.2–0.3 μg/μl stock of GFP-AGS mRNA and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. The white arrowhead indicates vegetal cortical localization of GFP-AGS. (D) Left graph: the number of embryos with vegetal cortical localization of 2x-GFP-AGS in 16–32-cell embryos was scored and normalized to that of the control group (SpAGS). Right graph: the ratio of the vegetal cortex-to-animal cortex mean intensity. Statistical analysis was performed against the control (SpAGS) by t-test. n indicates the total number of embryos scored. *p<0.05. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 5—figure supplement 1—source data 1
Numerical data for Figure 5—figure supplement 1D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp1-data1-v1.xlsx
Figure 5—figure supplement 2
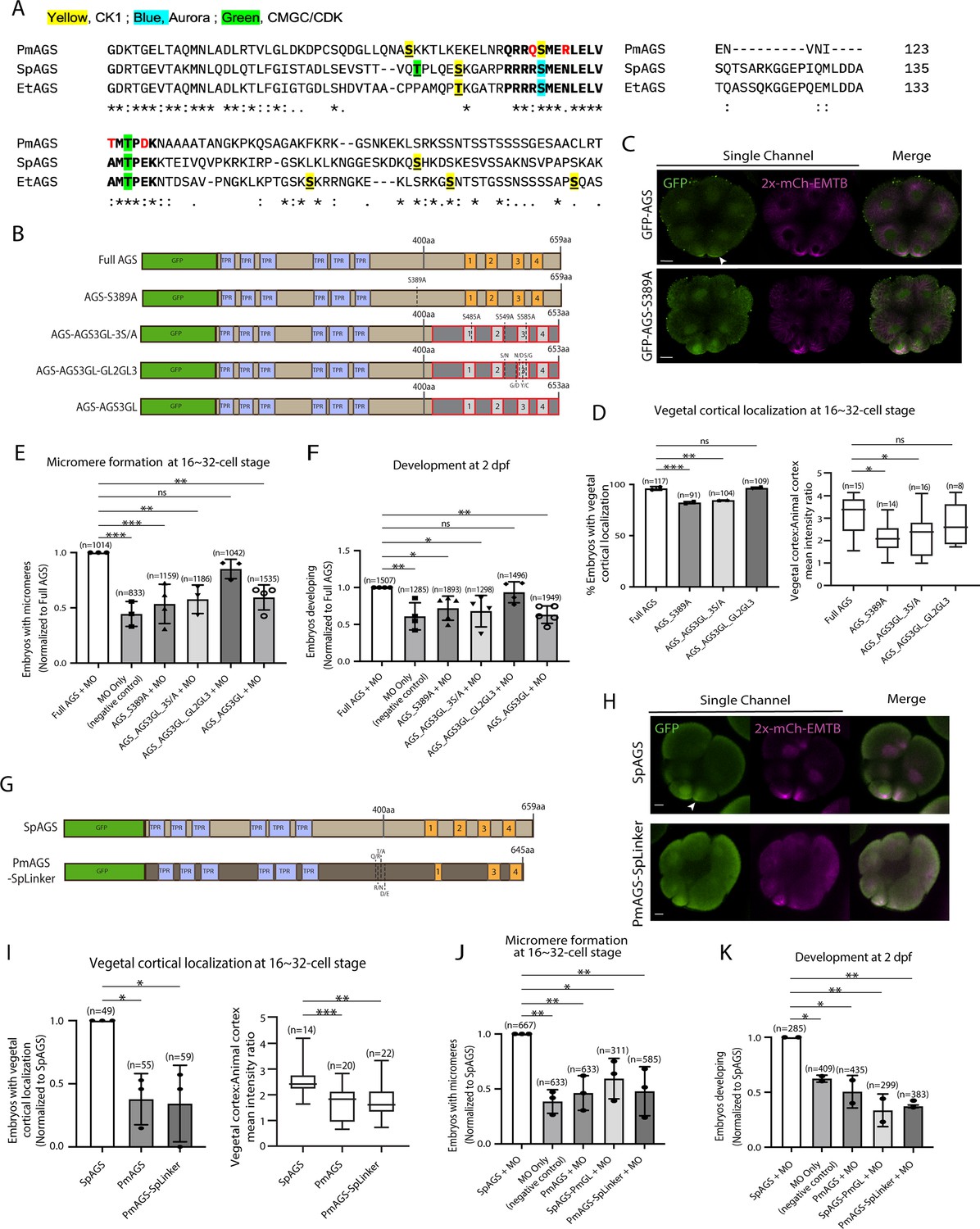
The linker domain and GL2-GL3 regions are important for AGS localization and function.
(A) Alignment of linker domain between echinoderms including sea urchin (SpAGS), pencil urchin (EtAGS), and sea star (PmAGS). Bold letters represent the conserved core linker domain. The yellow, blue, or green highlights indicate the CK1, Aurora, or CMGC/CDK phosphorylation sites predicted by GPS 6.0. The red letters indicate PmAGS amino acids mutated to those of SpAGS to construct the PmAGS-SpLinker mutant. (B) Design of GFP-AGS mutant constructs tested in this study. TPR motifs are marked in blue, and GL motifs are in orange. The brown section indicates the SpAGS sequence, and the red and gray boxes show the non-sea urchin (non-SpAGS) introduced at the C-terminus. The dotted lines represent single amino acid mutations. (C) Single z-slice confocal images of sea urchin embryos at the 8–16-cell stage showing localization of GFP-AGS-S389A mutant. Embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNA and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. The white arrowhead indicates vegetal cortical localization of GFP-AGS. (D) Percentage of the embryos with vegetal cortical localization of GFP-AGS mutants (left) and the ratio of the vegetal cortex-to-animal cortex mean intensity (right) in 16–32-cell embryos. Statistical analysis was performed against Full AGS by one-way ANOVA. (E, F) Embryos were injected with 0.15 μg/μl stock of GFP-AGS mRNAs and 0.75 mM SpAGS MO. The number of embryos forming micromeres (E) and developing to gastrula or pluteus stage (F) was scored, each of which was then normalized to that of the Full AGS. Statistical analysis was performed by one-way ANOVA. (G) Design of GFP-AGS constructs tested in this study from S. purpuratus (Sp) and P. miniata (Pm). TPR motifs are marked in blue, and GL motifs are in orange. The dotted lines represent single amino acid mutations. (H) Single Z-slice confocal images of sea urchin embryos at the 8–16-cell stage showing localization of GFP-AGS. Embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNAs and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. The white arrowhead indicates vegetal cortical localization of GFP-AGS. (I) The number of embryos with vegetal cortical localization of GFP-AGS mutants in 16–32-cell embryos was scored and normalized to that of the SpAGS (left graph). Right graph shows the ratio of the vegetal cortex-to-animal cortex mean intensity. Statistical analysis was performed against SpAGS by one-way ANOVA. (J, K) Embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNAs and 0.75 mM SpAGS MO. The number of embryos making micromeres (J) and developing to gastrula or pluteus stage (K) was scored and normalized to that of the SpAGS. Statistical analysis was performed by one-way ANOVA. n indicates the total number of embryos scored. *p<0.05, **p<0.01, and ***p<0.001. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 5—figure supplement 2—source data 1
Numerical data for Figure 5—figure supplement 2D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp2-data1-v1.xlsx
-
Figure 5—figure supplement 2—source data 2
Numerical data for Figure 5—figure supplement 2E.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp2-data2-v1.xlsx
-
Figure 5—figure supplement 2—source data 3
Numerical data for Figure 5—figure supplement 2F.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp2-data3-v1.xlsx
-
Figure 5—figure supplement 2—source data 4
Numerical data for Figure 5—figure supplement 2I.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp2-data4-v1.xlsx
-
Figure 5—figure supplement 2—source data 5
Numerical data for Figure 5—figure supplement 2J.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp2-data5-v1.xlsx
-
Figure 5—figure supplement 2—source data 6
Numerical data for Figure 5—figure supplement 2K.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig5-figsupp2-data6-v1.xlsx
Figure 5—figure supplement 3

Alignment of C-terminus GoLoco domain sequences used for chimeric mutants.
Sea urchin S. purpuratus (SpAGS_GL), Drosophila (DmPins_GL), sea star P. miniata (PmAGS_GL), pencil urchin E. tribuloides (EtAGS_GL), human H. sapiens LGN (HsLGN_GL), and human H. sapiens AGS3 (HsAGS3_GL). Bold letters indicate GoLoco motif sequences. The green highlight indicates additional serine amino acid present uniquely in HsAGS3_GL and mutated to alanine in AGS_AGS3GL_3S/A mutant. The highlighted amino acids between GL2 and GL3 and within GL3 are those mutated to match HsLGN_GL in AGS_AGS3GL_GL2GL3 mutant.
Figure 6
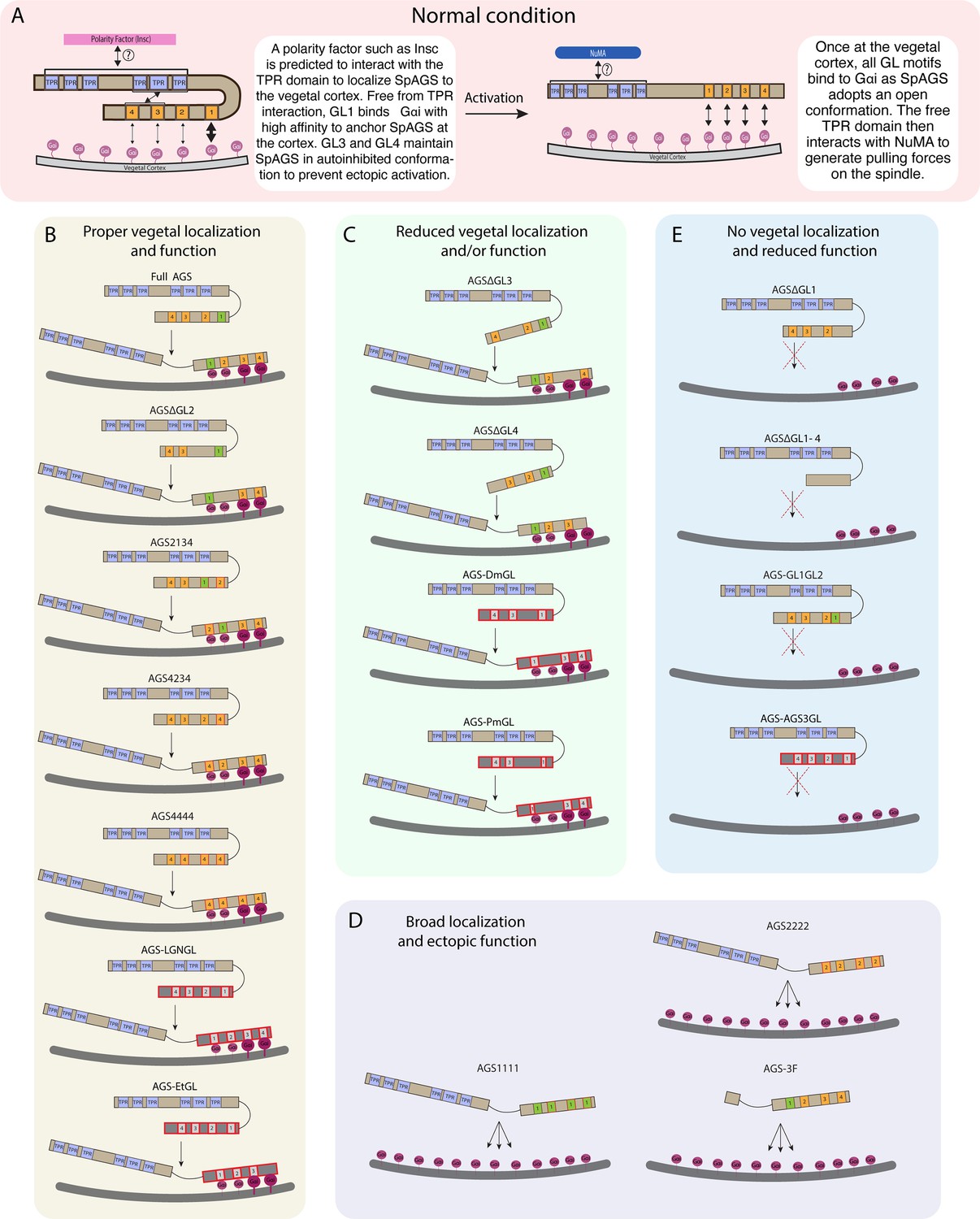
Summary diagrams of SpAGS dissection experiments.
(A) A model for the mechanism of SpAGS localization and function at the vegetal cortex. In a closed conformation, GL1 is critical for SpAGS recruitment and anchoring at the cortex through Gαi binding, while GL3 and GL4 maintain the autoinhibition. The TPR domain is hypothesized to interact with a polarity factor such as Insc to restrict SpAGS localization to the vegetal cortex. Upon Gαi binding, SpAGS adopts an open conformation, allowing all four GLs to bind to Gαi and the TPR domain to interact with NuMA for force generation on the astral microtubules. (B) A series of mutants that showed normal vegetal localization and functions. The position of GL1 is a more determining factor since mutants with GL1 replaced with other GL sequences localized and functioned properly. (C) A series of mutants that showed a reduced vegetal localization and/or function. The GL3 and GL4 are necessary to regulate AGS localization and function, likely by mediating its autoinhibitory mechanism through their binding to TPRs. Furthermore, AGS-DmGL and -PmGL were categorized in this group due to the reduced number of GL motifs. (D) A series of mutants that showed broad AGS localization and ectopic function. The TPR domain is critical for restricting AGS localization at the vegetal cortex since its removal spreads the AGS signal around all cortices. The sequences of GL3 and GL4 are also crucial for the SpAGS function. (E) A series of mutants that showed neither vegetal localization nor function. Removing or displacing GL1 led to significant disturbances in AGS localization and function, suggesting that having a GL motif at this specific position is critical for AGS interaction with Gαi and its anchoring to the cortex.
Figure 7 with 2 supplements
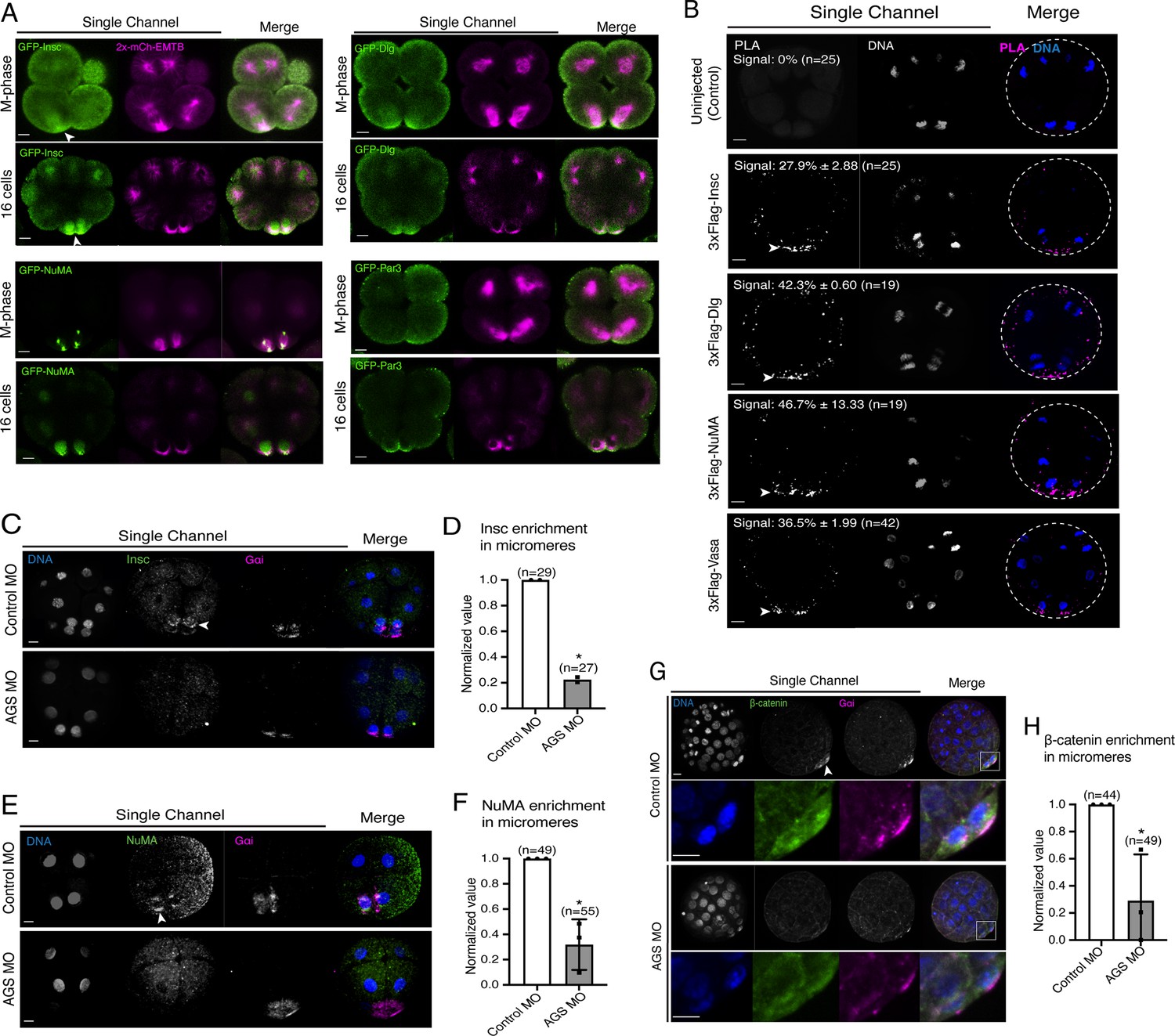
SpAGS is critical for the proper localization of asymmetric cell division (ACD) factors and fate determinants.
(A) Single Z-slice confocal imaging was used to focus on the vegetal cortex. Representative images of embryos during the metaphase and at the 16-cell stage show localization of each GFP-ACD factor, SpInsc, SpDlg, SpNuMA, and SpPar3. Embryos were injected with 0.5 μg/μl stock of GFP-ACD factor mRNAs and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. White arrowheads indicate vegetal cortical localization of GFP constructs. Images represent over 80% of the embryos observed (n = 30 or larger) per group across at least two independent cycles of experiments. (B) Single Z-slice confocal images of sea urchin embryos at the 8–16-cell stage showing the signals at the vegetal cortex by proximity ligation assay (PLA) assay with Flag and AGS antibodies. Embryos were injected with 0.3–1 μg/μl stock of 3xFlag-ACD factor mRNA. White arrowheads indicate the colocalization of AGS and another ACD factor at the vegetal cortex. The average % of the 8-cell and 8–16-cell embryos with the PLA signal across two independent cycles of experiments is indicated in each image. All embryos were scored independently of the angle since it was hard to identify the angle at the 8-cell stage. (C–F) Representative 2D-projection images of the embryo stained with Insc (C), NuMA (E), and β-catenin (G) antibodies (green) by immunofluorescence. Embryos were stained with Gɑi antibody (magenta) and Hoechst dye (blue) as well. Z-stack images were taken at 1 μm intervals to cover a layer of the embryo. White arrowheads indicate the signal in micromeres. Embryos were injected with 0.75 mM control MO or 0.75 mM SpAGS MO. The number of embryos showing the localization of Insc (D), NuMA (F), and β-catenin (H) in micromeres was scored and normalized to that of the control MO. Statistical analysis was performed by t-test. n indicates the total number of embryos scored. *p<0.05. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 7—source data 1
Numerical data for Figure 7D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-data1-v1.xlsx
-
Figure 7—source data 2
Numerical data for Figure 7F.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-data2-v1.xlsx
-
Figure 7—source data 3
Numerical data for Figure 7H.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-data3-v1.xlsx
Figure 7—figure supplement 1
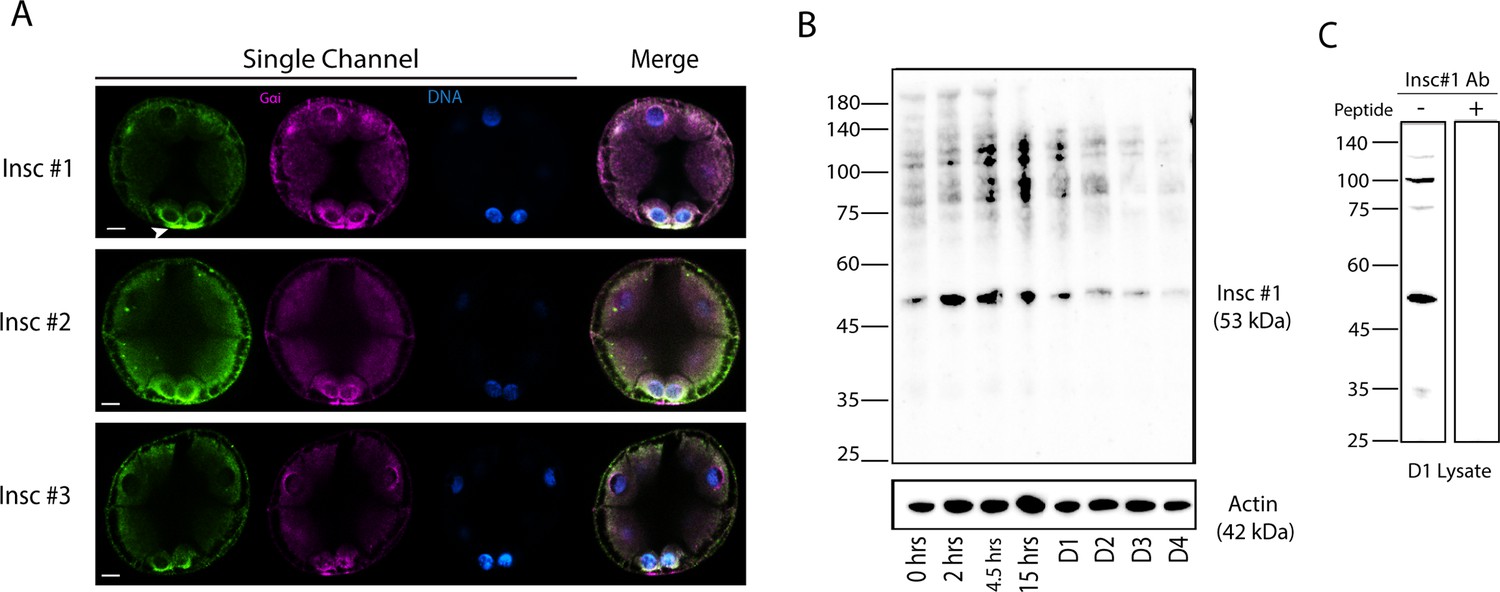
Insc protein expression during embryonic development.
(A) Endogenous Insc protein localization by immunofluorescence. Embryos were stained with three Insc antibodies (green) designed for different Insc amino acid sequence sections. Embryos were stained with Gɑi antibody (magenta) and Hoechst dye (blue). During the 16-cell stage, all antibodies show signal enriched at the vegetal pole. With #2 and #3 antibodies, some nonspecific cortex signals were also observed around the entire embryo. (B) Insc immunoblot analysis. Embryos were collected at 0, 2, 4.5, 15, 24, 48, 72, and 96 hr post fertilization and subjected to immunoblot with Insc #1 antibody. Actin (42 kDa) was used as a loading control. The expected size of Insc is 53 kDa. (C) Peptide competition assay with Insc #1 antibody. The 24 hr lysate was used. Each experiment was performed at least two independent times. Scale bars = 10 μm.
-
Figure 7—figure supplement 1—source data 1
Original blots with labels for Figure 7—figure supplement 1B.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-figsupp1-data1-v1.zip
-
Figure 7—figure supplement 1—source data 2
Original blot images for Figure 7–Supplement 1C.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-figsupp1-data2-v1.zip
-
Figure 7—figure supplement 1—source data 3
The PDF file of original blots with labels for Figure 7—figure supplement 1C.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-figsupp1-data3-v1.pdf
-
Figure 7—figure supplement 1—source data 4
Original blot images for Figure 7—figure supplement 1C.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig7-figsupp1-data4-v1.zip
Figure 7—figure supplement 2
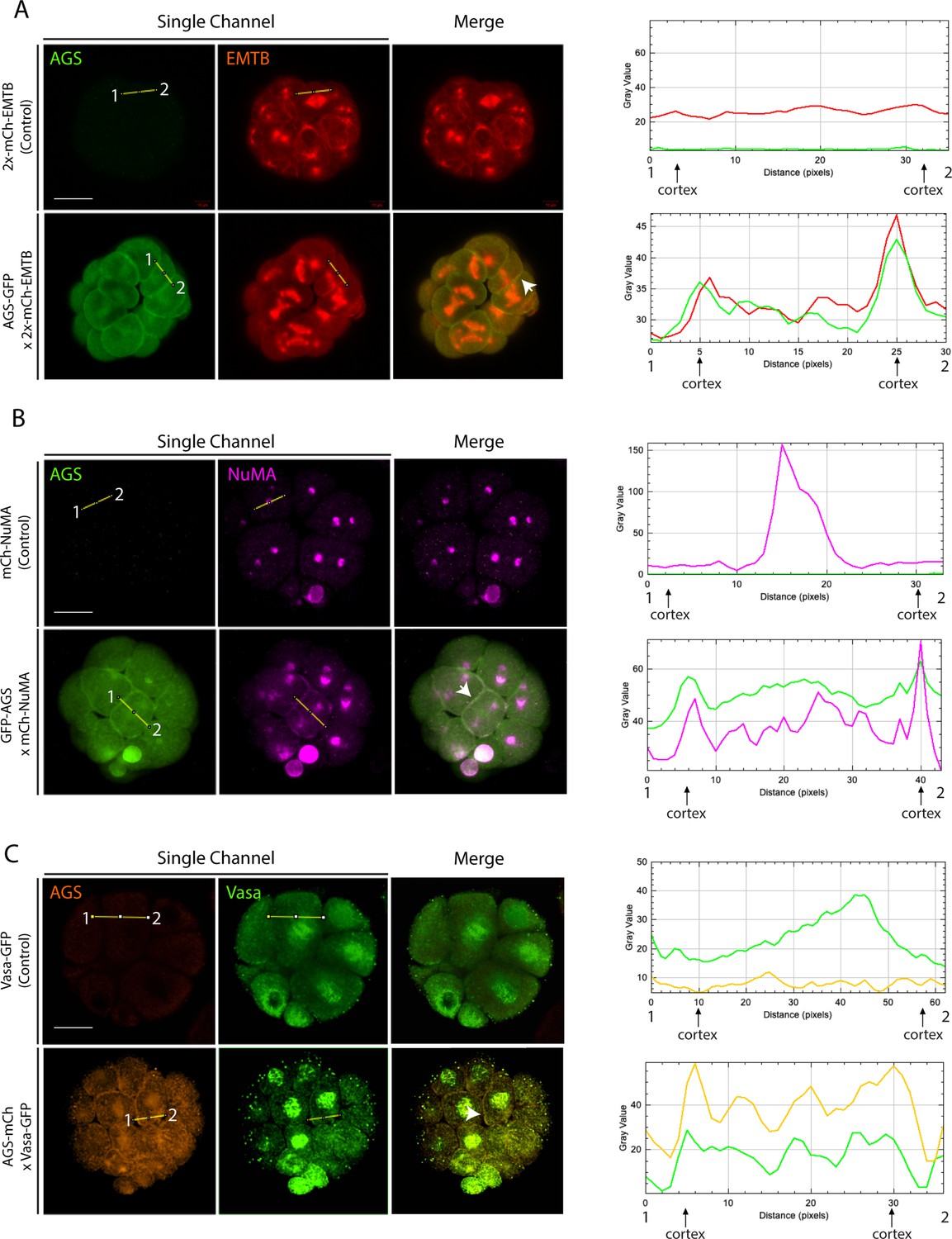
SpAGS colocalizes with micromere-specific fate determinants.
(A) Embryos were co-injected with 2x-mCherry-EMTB (0.5 μg/μl stock) mRNA with or without GFP-AGS (0.5 μg/μl stock) mRNA. (B) Embryos were co-injected with mCherry-NuMA (0.15 μg/ul) mRNA with or without GFP-AGS (0.5 μg/μl stock). (C) Embryos were co-injected with Vasa-GFP (1 μg/μl stock) mRNA with or without AGS-mCherry (0.5 μg/μl stock) mRNA. The intensity of each signal, from one cortex to the other, was measured and plotted from point 1 to 2 on the corresponding graph (right) using ImageJ. White arrows indicate the cortical colocalization of each construct. All images represent over 50% of the embryos observed (n = 30 or larger) per group. Scale bars = 20 μm.
Figure 8 with 2 supplements
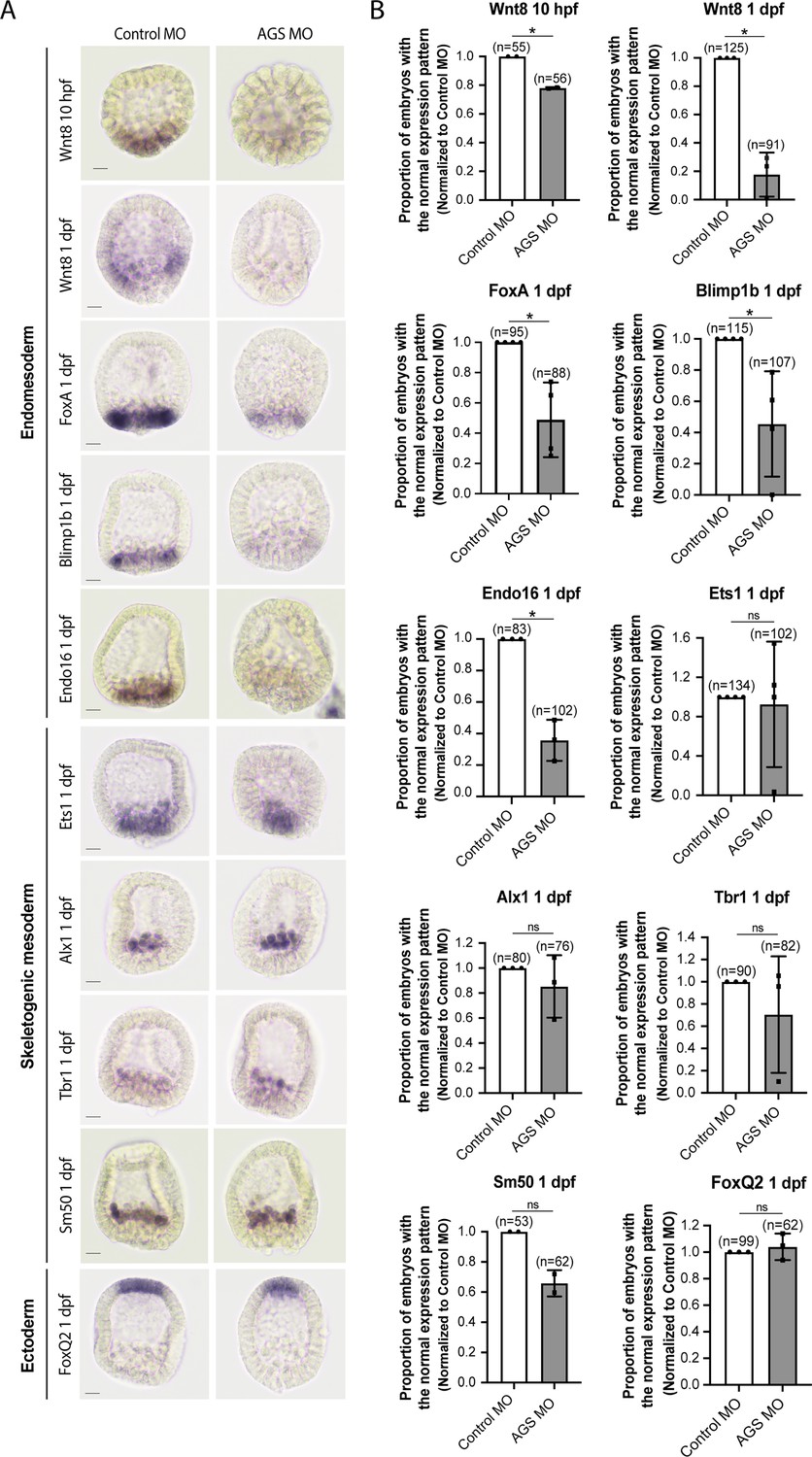
SpAGS is critical for the downstream gene expressions regulated by micromere signaling.
Embryos were injected with 0.75 mM control MO or 0.75 mM SpAGS MO. Brightfield images (A) show the representative in situ hybridization (ISH) staining for each cell lineage marker scored in the corresponding graph (B). The number of embryos showing the normal signal patterns of each marker gene was scored and normalized to that of the control MO. Statistical analysis was performed by t-test. n indicates the total number of embryos scored. *p<0.05. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 20 μm.
-
Figure 8—source data 1
Numerical data for Figure 8B.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig8-data1-v1.xlsx
Figure 8—figure supplement 1

Sea urchin (SpDlg) and sea star (PmDlg) sequence alignment.
Blue, yellow, and green highlights indicate the PDZ, SH3, and GUK domains, respectively.
Figure 8—figure supplement 2

Sea urchin (SpDlg) and sea star (PmDlg) sequence alignment.
Blue, yellow, and green highlights indicate the PDZ, SH3, and GUK domains, respectively.
Figure 9
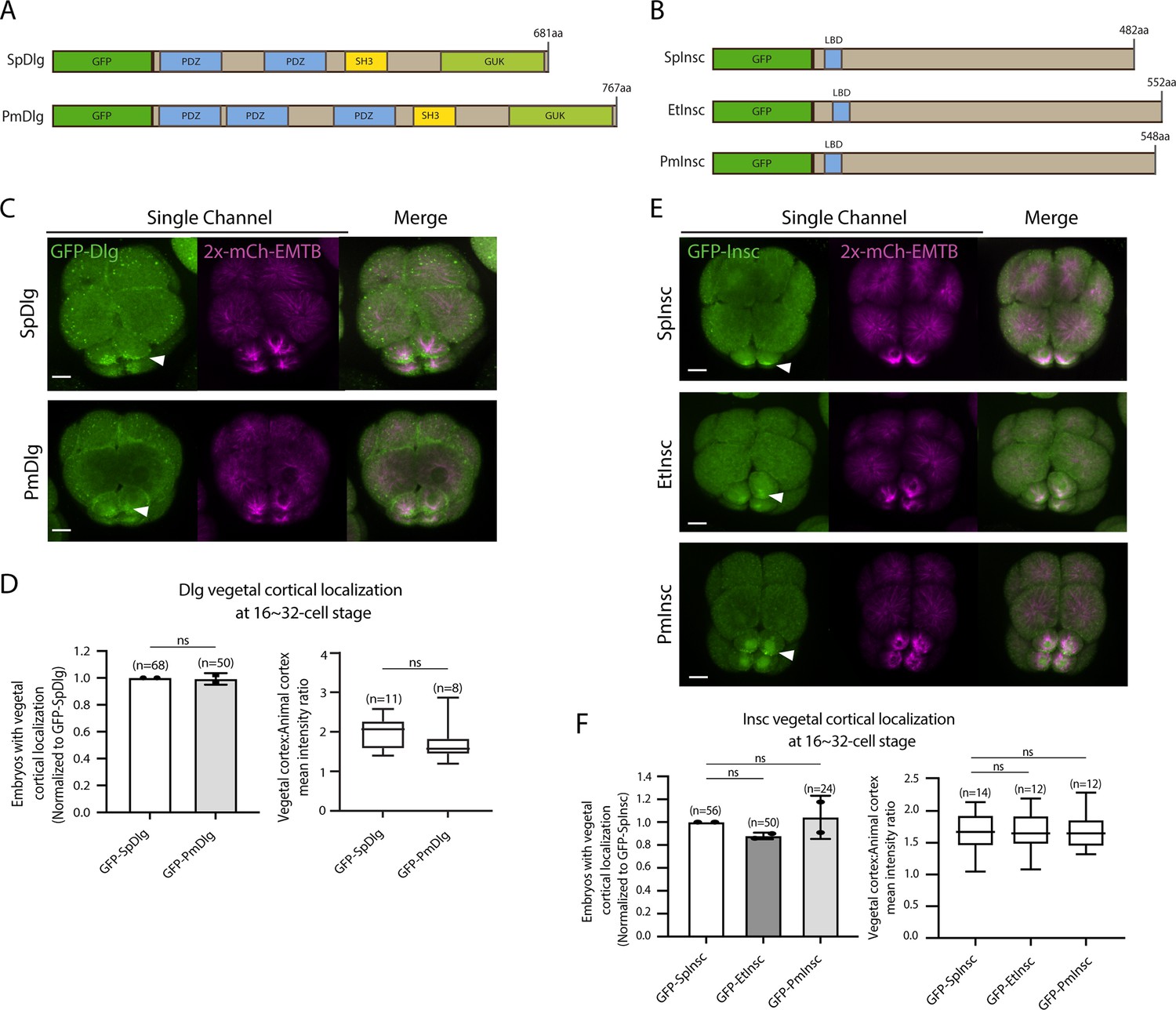
Dlg and Insc are not the variable factors of the asymmetric cell division (ACD) machinery in evolution.
(A, B) Design of GFP-Dlg (A) and GFP-Insc (B) constructs that were tested in this study. Of note, EtDlg was unavailable in the database due to the limited genomic information available for this species. (C–F) Representative 2D-projection images of sea urchin embryos at the 8–16-cell stage showing localization of each echinoderm GFP-Dlg (C) and GFP-Insc (E). Z-stack images were taken at 1 μm intervals. Embryos were injected with 0.5 μg/μl stock of GFP-Dlg or GFP-Insc mRNAs and 0.25 μg/μl stock of 2x-mCherry-EMTB mRNA. White arrowheads indicate vegetal cortical localization of GFP constructs. The number of embryos with vegetal cortical localization of GFP-Dlg (D) and GFP-Insc (F) in 16–32-cell embryos was scored and normalized to that of the GFP-SpDlg or GFP-SpInsc (left graph). Right graph shows the ratio of the vegetal cortex-to-animal cortex mean intensity. Statistical analysis was performed by t-test or one-way ANOVA. n indicates the total number of embryos scored. Each experiment was performed at least two independent times. Error bars represent standard error. Scale bars = 10 μm.
-
Figure 9—source data 1
Numerical data for Figure 9D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig9-data1-v1.xlsx
-
Figure 9—source data 2
Numerical data for Figure 9F.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig9-data2-v1.xlsx
Figure 10
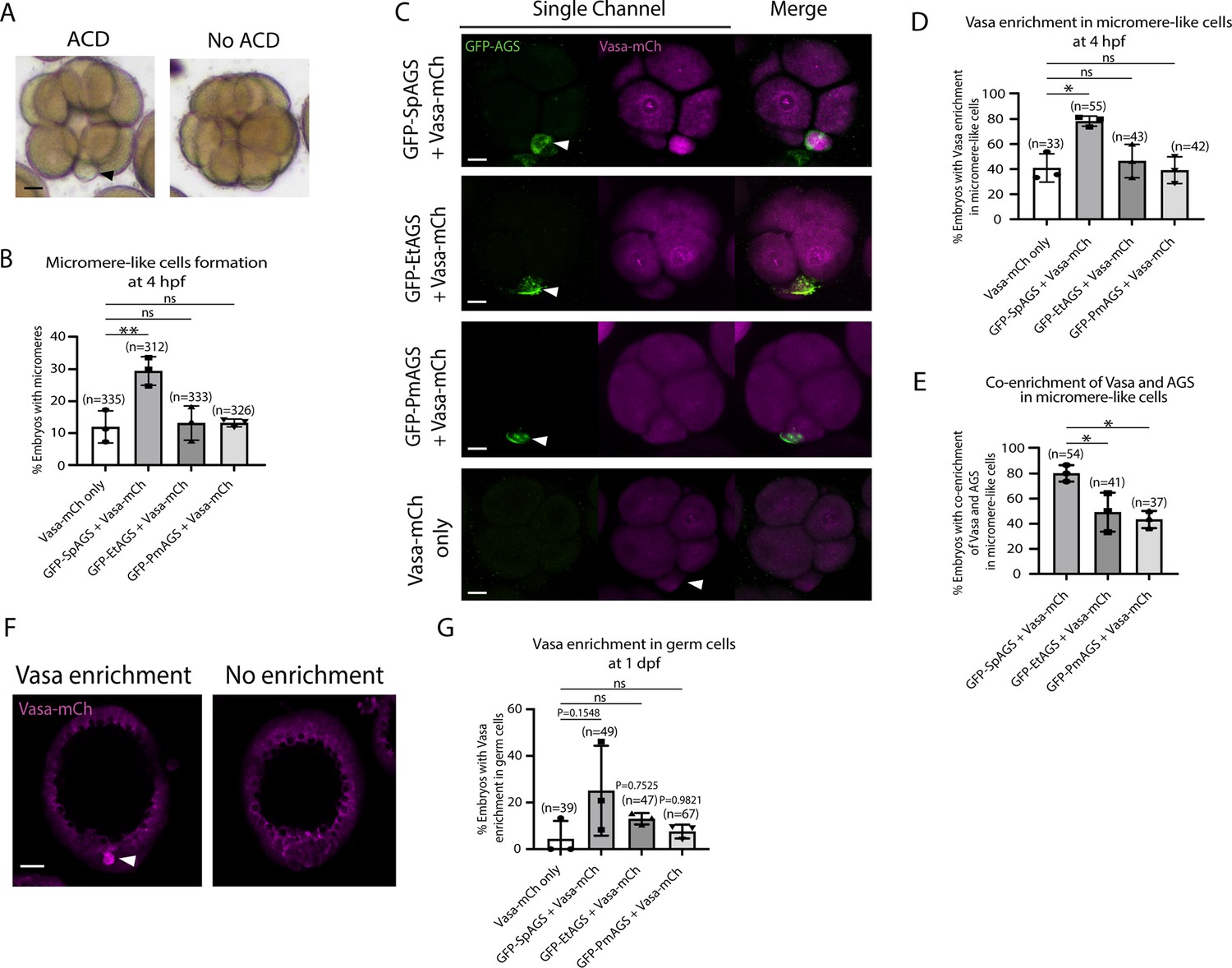
SpAGS but not EtAGS and PmAGS can induce a functional asymmetric cell division (ACD) in pencil urchin (Et) embryos.
(A) Representative brightfield images of pencil urchin (Et) embryos with or without micromere-like cells. Black arrowhead indicates micromere-like cells. (B) Et embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNAs and 1 μg/μl stock of Vasa-mCherry mRNA. The number of embryos making micromere-like cells was scored and normalized to that of the Vasa-mCherry only. Statistical analysis was performed against Vasa-mCherry only by one-way ANOVA. (C) Representative 2D-projection images of the injected Et embryos. Z-stack images were taken at 1 μm intervals to cover a layer of the embryo. White arrowheads indicate micromere-like cells. Scale bars = 10 μm. (D) The number of embryos showing Vasa enrichment in the micromere-like cells was scored and shown in percentage (%). Only the embryos that formed micromere-like cells were scored. Statistical analysis was performed against Vasa-mCherry only by one-way ANOVA. (E) Percentage of total embryos showing co-enrichment of Vasa and AGS in the micromere-like cells. Statistical analysis was performed against GFP-SpAGS by one-way ANOVA. (F, G) Representative 2D-projection images of Et embryos at 1 dpf. White arrowhead indicates Vasa enrichment in germ cells. Z-stack images were taken at 1 μm intervals. Percentage of total embryos showing Vasa enrichment in germ cells at 1 dpf was scored. Et embryos were injected with 0.3 μg/μl stock of GFP-AGS mRNAs. Statistical analysis was performed against Vasa-mCherry only by one-way ANOVA. n indicates the total number of embryos scored. *p<0.05 **p<0.01. Each experiment was performed at least three independent times. Error bars represent standard error. Scale bars = 20 μm.
-
Figure 10—source data 1
Numerical data for Figure 10B.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig10-data1-v1.xlsx
-
Figure 10—source data 2
Numerical data for Figure 10D.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig10-data2-v1.xlsx
-
Figure 10—source data 3
Numerical data for Figure 10E.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig10-data3-v1.xlsx
-
Figure 10—source data 4
Numerical data for Figure 10G.
- https://cdn.elifesciences.org/articles/100086/elife-100086-fig10-data4-v1.xlsx
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Antibody | Anti-SpAGS (rabbit serum) | Voronina and Wessel, 2006; doi:10.1111/j.1440-169X.2006.00895.x | N/A | IF (1:300) PLA (1:300) |
| Antibody | Anti-Gαi (mouse monoclonal) | Santa Cruz Biotech | sc-56536 | IF (1:30) |
| Antibody | Anti-β-catenin (rabbit polyclonal) | Yazaki et al., 2015; doi:10.1017/S0967199414000033 | N/A | IF (1:300) |
| Antibody | Anti-SpInsc (rabbit polyclonal) | This article | N/A | See ‘Insc antibody production and validation’ WB (1:2000) IF (1:200) |
| Antibody | Anti-SpNuMA (rabbit polyclonal) | Poon et al., 2019; doi:10.1038/s41467-019-11560-8 | N/A | IF (1:500) |
| Antibody | Anti-β-actin (8H10D10) (mouse monoclonal) | Cell Signaling Technology | 3700S | WB (1:5000) |
| Antibody | Anti-Flag (mouse monoclonal) | MilliporeSigma | F1804 | PLA (1:100) |
| Antibody | Alexa 488-conjugated goat anti-rabbit IgG (goat polyclonal) | Cell Signaling Technology | 4412 | IF (1:300) |
| Antibody | Alexa 555-conjugated goat anti-mouse IgG (goat polyclonal) | Cell Signaling Technology | 4409 | IF (1:300) |
| Antibody | HRP-conjugated anti-Protein A antibody (goat polyclonal) | Abcam | ab7245 | WB (1:2000) |
| Antibody | HRP-conjugated goat anti-mouse IgG (horse polyclonal) | Cell Signaling Technology | 7076 | WB (1:2000) |
| Antibody | Anti-Digoxigenin-AP, Fab fragments (sheep polyclonal) | Roche | 11093274910 | ISH (0.1–0.5 ng/μl) |
| Chemical compound, drug | Hoechst 33342 | Thermo Fisher Scientific | 62249 | IF (1:1000) |
| Chemical compound, drug | Tris buffered saline, with tween (TBST) | MilliporeSigma | T9039 | |
| Chemical compound, drug | Tris-MOPS-SDS Running Buffer | GenScript | M00138 | |
| Chemical compound, drug | Transfer buffer powder | GenScript | M00139 | |
| Chemical compound, drug | DIG RNA labeling mix | Roche | 11277073910 | |
| Commercial assay or kit | mMESSAGE mMACHINE SP6 Transcription Kit | Thermo Fisher Scientific | AM1340 | |
| Commercial assay or kit | MEGAscript SP6 Transcription kit | Thermo Fisher Scientific | AM1330 | |
| Commercial assay or kit | MEGAscript T7 Transcription kit | Thermo Fisher Scientific | AM1333 | |
| Commercial assay or kit | In-Fusion HD Cloning | Clontech | 639648 | |
| Commercial assay or kit | Duolink In Situ Red Starter Kit Mouse/Rabbit | MilliporeSigma | DUO92101 | |
| Recombinant DNA reagent | Plasmid: SpAGS-GFP | Poon et al., 2019; doi:10.1038/s41467-019-11560-8 | N/A | |
| Recombinant DNA reagent | Plasmids: SpAGS-dC-term-GFP, SpAGS-dGL1/2/3/4-GFP, SpAGS1111/2222/2134/4234-GFP | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmids: SpAGS-dN-term-GFP | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: SpAGS-mCherry | Poon et al., 2019; doi:10.1038/s41467-019-11560-8 | N/A | |
| Recombinant DNA reagent | Plasmid: GFP-SpAGS, GFP-EtAGS, GFP-PmAGS, 2x-GFP-SpAGS, 2x-GFP-SbAGS | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: GFP-SpAGS4444/GL1GL2/LGNGL/DmGL/EtGL/AGS3GL/PmGL/S389A/AGS3GL-3S/A/AGS3GL-GL2GL3 | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: GFP-PmAGS-SpLinker | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: GFP-SpDlg/PmDlg | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: GFP-SpInsc/EtInsc/PmInsc | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: GFP-NuMA | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: mCherry-NuMA | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: GFP-Par3 | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: Vasa-GFP | Yajima and Wessel, 2011; doi:10.1242/dev.054940 | N/A | |
| Recombinant DNA reagent | Plasmid: Vasa-mCherry | Uchida and Yajima, 2018; doi:10.1016/j.ydbio.2018.06.015 | N/A | |
| Recombinant DNA reagent | Plasmid: 3xFlag-GFP-SpAGS/SpDlg/SpNuMA, 3xFlag-Vasa-GFP | This article | N/A | See ‘Plasmid construction’ |
| Recombinant DNA reagent | Plasmid: 2x-mCherry-EMTB | Addgene | 26742 | |
| Software, algorithm | EchinoBase | http://www.echinobase.org/Echinobase/ | N/A | Echinoderm protein sequences |
| Software, algorithm | NCBI blast | https://blast.ncbi.nlm.nih.gov/Blast.cgi | N/A | Protein motif search |
| Software, algorithm | Clustal Omega | https://www.ebi.ac.uk/Tools/msa/clustalo/ | N/A | Protein sequence alignment |
| Software, algorithm | ImageJ | https://imagej.nih.gov/ij/ | N/A | Quantitative analysis |
| Software, algorithm | GraphPad PRISM 8 | https://www.graphpad.com/scientific-software/prism/ | N/A | Statistical analysis |
| Sequence-based reagent | SpAGS-MO | NM_001040405.1 | Morpholino antisense oligos | GGCCCGTTTCACAAAGCCTTTGTTT |
| Sequence-based reagent | SpAlx1 | XM_011663478.2 | ISH probe primers | F: GGATATTTTCTCGACCGGGATC R: CGAGTAACCGTTCATCATCCCC |
| Sequence-based reagent | SpBlimp1b | NM_214574.3 | ISH probe primers | F: ATGGGGTGCAACGACAACGCCGTG R: CTATGATTTGTTCGTACGATTGAG |
| Sequence-based reagent | SpEndo16 | NM_214519.1 | ISH probe primers | F: GCAGAGTTCAACAGAATCGAC R: GCCAGTAGACGTAGCAGAAG |
| Sequence-based reagent | SpEts1 | XM_030976919.1 | ISH probe primers | F: TCAATCATGGCGTCTATGCACTG R: ACAGCTGCAGGGATAACAGG |
| Sequence-based reagent | SpFoxA | NM_001079542.1 | ISH probe primers | F: ATGGCCAATAGTGCCATGATCTCG R: TCACATTGCATGGTTTGCTTG |
| Sequence-based reagent | SpFoxQ2 | XM_003731512.3 | ISH probe primers | F: ATGACTTTATTCAGCATTGACAAC R: TAGCAGGATCCTACAGAAGACCAG |
| Sequence-based reagent | SpSm50 | NM_214610.3 | ISH probe primers | F: ATGAAGGGAGTTTTGTTTATTGTGG CTAGTC R: GTTATGCCAACGCGTCTGCCTCTTG AAGC |
| Sequence-based reagent | SpTbr1 | XM_786173.5 | ISH probe primers | F: CCACCGCTGCACCAGACGAC R: CTGCCGGCTGGCGCCAATTGCG |
| Sequence-based reagent | SpWnt8 | NM_214667.1 | ISH probe primers | F: ATGGATGTTTTTACGGAATTTGTTCG R: CTACAGCCTCGATCCAACGGGCTG |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The evolutionary modifications of a GoLoco motif in the AGS protein facilitate micromere formation in the sea urchin embryo
eLife 13:RP100086.
https://doi.org/10.7554/eLife.100086.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}