A dual nature of γδ T cell immune memory responses
- Immunology and Metabolism Unit, Life and Medical Sciences (LIMES) Institute, University of Bonn, Germany
- Institut of Immunology, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, Germany
- Department of Internal Medicine and Radboud Center for Infectious Diseases, Radboud University Medical Center, Netherlands
Figures
Figure 1
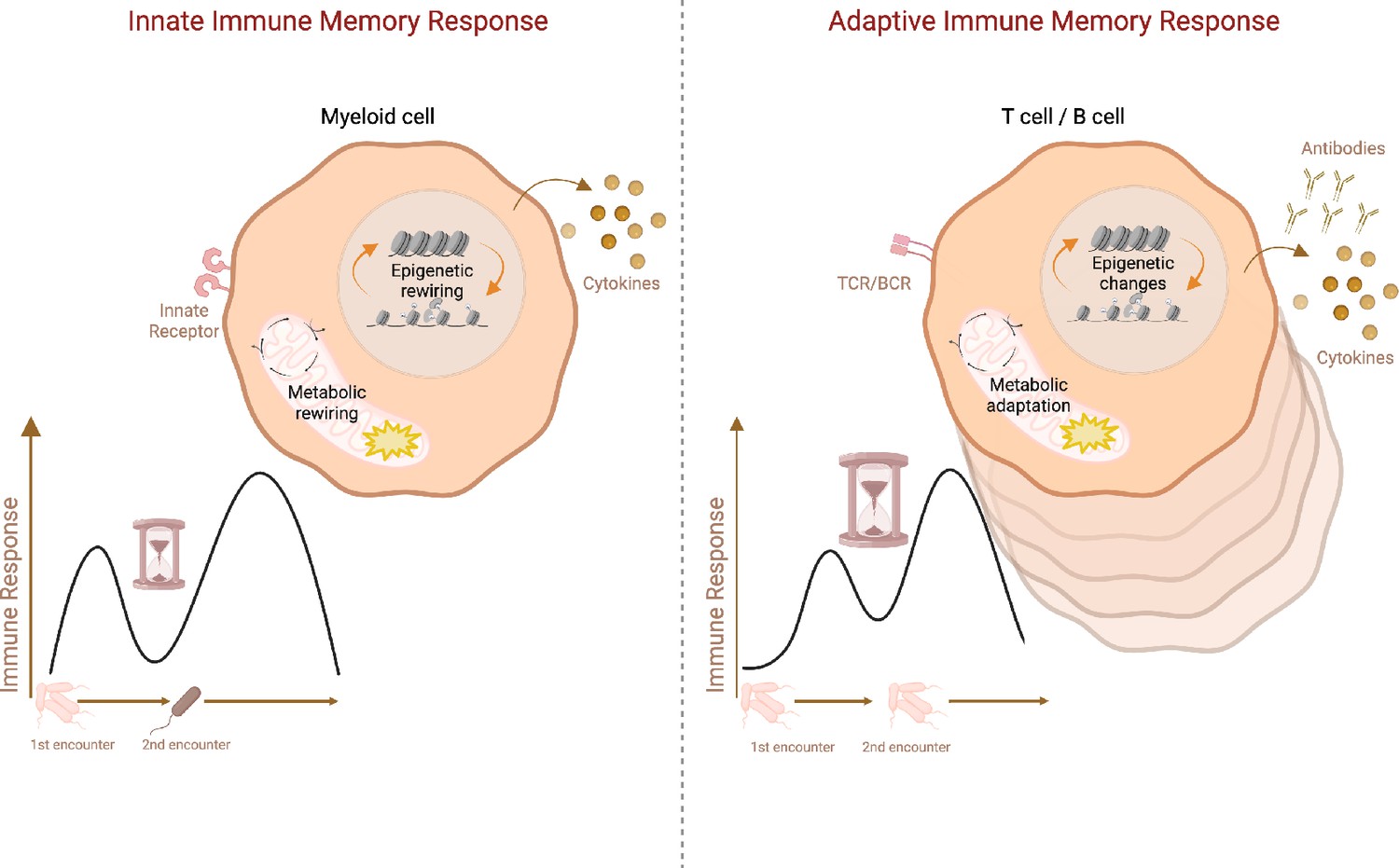
Schematic representation of adaptive vs. innate immune memory characteristics.
Myeloid cells and lymphocytes mount immune memory responses characterized by the enhanced effector function upon secondary exposure. While innate immune cells produce more inflammatory cytokines upon secondary challenge with heterologous stimuli, adaptive memory immune cells rapidly proliferate and generate copious amounts of antibodies and cytokines upon rechallenge with the same antigen. Antibodies, as well as memory B cells and T cells, persist in the host while cytokines produced by innate immune cells return to the baseline after the resolution of infection. Innate immune memory lasts relatively shorter than adaptive immune memory. Both adaptive and innate immune memory formation is accompanied by epigenetics and metabolic rewiring, facilitating transcriptional responses and allowing more robust immune reactions upon secondary challenge. TCR: T-cell receptor, BCR: B-cell receptor. Created with BioRender.com.
Tables
Table 1
Immune memory responses of gamma delta (γδ) T cells.
BCG: Bacille Calmette-Guerin; CMV: Cytomegalovirus; HSV: herpes simplex virus; IMQ: imiquimod; MMR: measles-mumps-rubella; MPV: Mpox virus.
| Species | T cell subset | Experimental setting | Location | Adaptive immune memory responses | Innate immune memory responses | Ref | ||
|---|---|---|---|---|---|---|---|---|
| Inducing agent | Immune memory response characteristics | Inducing agent | Immune memory response characteristics | |||||
| Human and non-human primates | Vδ2 | In vitro | Peripheral blood | BCG | Enhanced proliferation upon M. tuberculosis stimulation | Hoft et al., 1998; Kabelitz et al., 1991 | ||
| Vδ2 | In vivo | Peripheral blood | BCG | Enhanced proliferation and IFN-γ production upon restimulation | Kabelitz et al., 1991 | |||
| γδ | In vivo | Pulmonary and peripheral blood | BCG | Enhanced proliferation upon reinfection | Shen et al., 2002; Lai et al., 2003 | |||
| Vδ2 | In vivo | Peripheral blood | Listeria monocytogenes | Enhanced proliferation and effector function upon reinfection | Ryan-Payseur et al., 2012 | |||
| γδ | In vivo | Peripheral blood | Plasmodium falciparum | Enhanced proliferation and IFN-γ production upon restimulation | Teirlinck et al., 2011 | |||
| Vδ1 | In vivo | Peripheral blood | Plasmodium falciparum | Clonal expansion, recurrent parasite-exposure driven expansion and differentiation | von Borstel et al., 2021; Rutishauser et al., 2020 | |||
| Vδ2 | In vivo | Peripheral blood | SARS-CoV-2 mRNA vaccine | Enhanced proliferation and IFN-γ production upon revaccination | Terzoli et al., 2024 | |||
| Vδ2 | In vivo | Peripheral blood | MPV | Enhanced proliferation and IFN-γ production upon rechallenge | Shao et al., 2009 | |||
| Vδ1 | In vivo | Peripheral blood | CMV | Rapid proliferation and infection resolution after reinfection | Pitard et al., 2008 | |||
| Vδ2 | In vivo | Pulmonary compartment | Listeria monocytogenes | Enhanced IFN-γ and perforin production; lower pulmonary pathology and less weight loss upon M. tuberculosis infection | Shen et al., 2019 | |||
| γδ | In vitro | Peripheral blood | BCG | Enhanced TNF and IFN-γ production upon C. albican challenge; transcriptional rewiring | Suen et al., 2024 | |||
| Vδ2 | In vitro | Peripheral blood | HSV | Enhanced lysing ability of infected cells upon PHA or mycobacteria stimulation | Bukowski et al., 1994 | |||
| γδ | In vitro | Peripheral blood | MMR | Enhanced TNF and IFN-γ production upon CD3 stimulation; transcriptional and metabolic rewiring | Röring et al., 2024 | |||
| Mouse | Vγ4Vδ1 | In vivo | Intestinal mucosa | Listeria monocytogenes | Enhanced proliferation and infection clearance upon rechallenge | Sheridan et al., 2013 | ||
| Vγ4 | In vivo | Intestinal epithelium | Listeria monocytogenes | Enhanced IL-17 production and clustering with L monocytogenes replication foci upon secondary infection | Romagnoli et al., 2016 | |||
| Vγ6 | In vivo | Peritoneum, draining mediastinal lymph nodes | Staphylococcus aureus | Enhanced IL-17 production and infection clearance after reinfection | Murphy et al., 2014 | |||
| Vγ6 | In vivo | Kidney | Staphylococcus aureus | Reduced renal bacterial load upon reinfection | Bertram et al., 2023 | |||
| Vγ1 | In vivo | Liver, lung, spleen | MCMV | Enhanced proliferation and survival rate upon rechallenge | Khairallah et al., 2015 | |||
| γδ | In vitro | Liver, spleen | Plasmodium chabaudi | Enhanced CD107a expression and IFN-γ production upon rechallenge; transcriptional reprogramming | Kumarasingha et al., 2020 | |||
| Vγ4Vδ4 | In vivo | Skin | IMQ | Enhanced proliferation and IL-17 production upon IMQ rechallenge | Ramírez-Valle et al., 2015 | |||
| Vγ4Vδ4 | In vivo | Skin; ear | IMQ | Enhanced proliferation and IL-17 production and neutrophil recruitment upon IMQ rechallenge | Hartwig et al., 2015 | |||
| Vγ4Vδ1 | In vitro | Gut; bulk mesenteric lymph nodes | Listeria monocytogenes | Enhanced proliferation and IFN-γ and IL-17A production upon S. enterica serovar Typhimurium and C. rodentium challenge | Khairallah et al., 2022 | |||
| Cow | γδ | In vitro | Airway and peripheral blood | BCG | Increased IFN-γ producing γδ T cells | Guerra-Maupome and McGill, 2019 | ||
| γδ | In vitro | Peripheral blood | BCG | Enhanced IL-6 and TNF production upon Escherichia coli, LPS and Pam3CSK4 stimulation; epigenetic rewiring | Samuel et al., 2024 | |||
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A dual nature of γδ T cell immune memory responses
eLife 14:e104887.
https://doi.org/10.7554/eLife.104887




{kind=link}