Tree crickets optimize the acoustics of baffles to exaggerate their mate-attraction signal
- University of Bristol, United Kingdom
- Indian Institute of Science, India
Figures
Figure 1 with 2 supplements
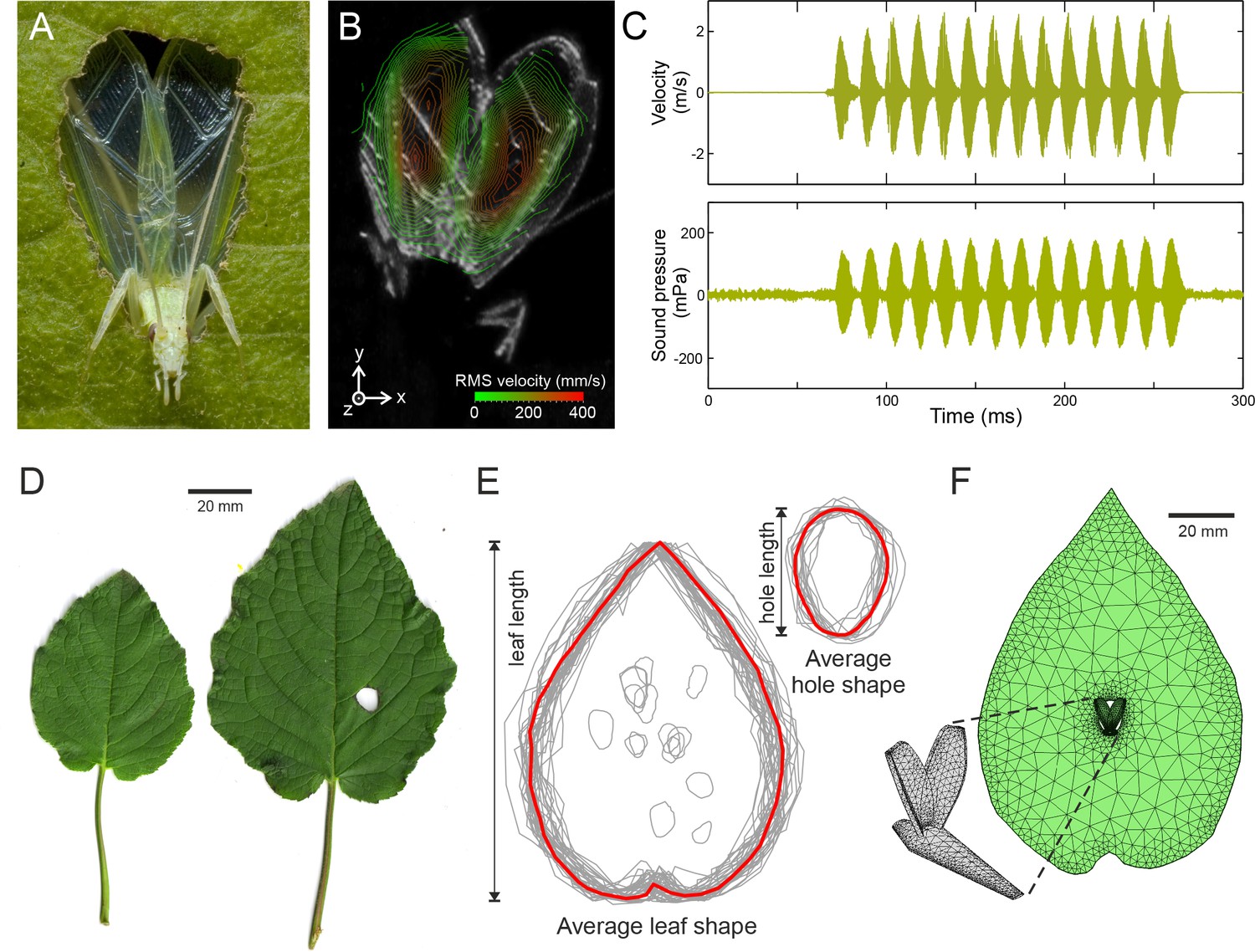
Tree cricket calling and baffle-use behaviour.
(A) A wild male Oecanthus henryi calling from a baffle (photograph reproduced from [Mhatre et al., 2012]). (B) RMS wing velocity (in the z-direction) of a male calling from a baffle hole (see Figure 1—figure supplement 1 for details). (C) Time-resolved wing velocity measured at the location of highest velocity (upper trace), and sound pressure level 200 mm from the cricket (lower trace). Peak velocity averaged 1.8 ± 0.6 m·s−1, peak displacement reached 95.67 ± 29.54 µm, and peak accelerations of 4629 ± 1733 g were measured. This gave rise to peak SPLs of 108.8 ± 48.5 mPa (74.0 ± 3.9 dB). (D) Examples of host plant leaves (Hyptis suaveolens) offered to a male in choice experiments. The larger leaf shows a cricket-made baffle hole. (E) Outlines of all leaves and baffle holes in the choice experiment normalized for size and overlaid to calculate the average leaf and baffle hole shapes (red outline). (F) Finite element model of an average shaped baffle with detail showing cricket model. Model baffle hole is shown at the leaf centre, but can be arbitrarily positioned across the leaf surface.
Figure 1—figure supplement 1
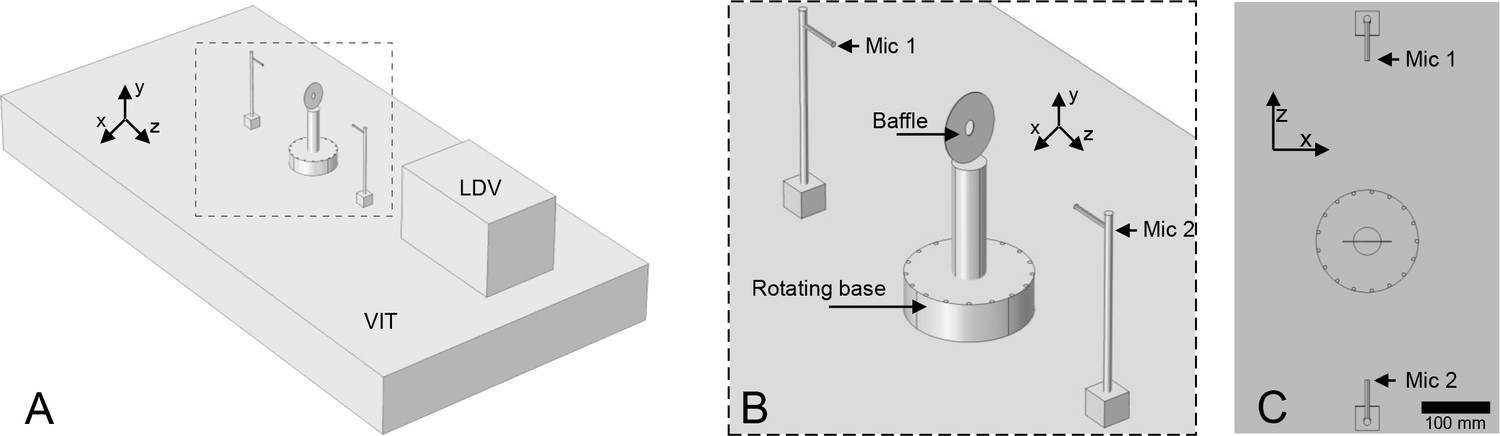
Experimental set-up for vibrometry of calling tree crickets.
The insects were released onto the baffle assembly and would preferentially call from the baffle. The base was rotated so the insect’s wings were oriented normal to the laser. Once the animal began calling, measurements from the two microphones were taken simultaneously with the vibrometry data. A: Diagrammatic representation of complete set-up. B: Close up view of baffle assembly. C: Plan view of baffle assembly and microphone positions. LDV: Laser Doppler vibrometer, VIT: Vibration isolation table, Mic 1: front microphone, Mic 2: rear microphone. Figure to scale.
Figure 1—figure supplement 2
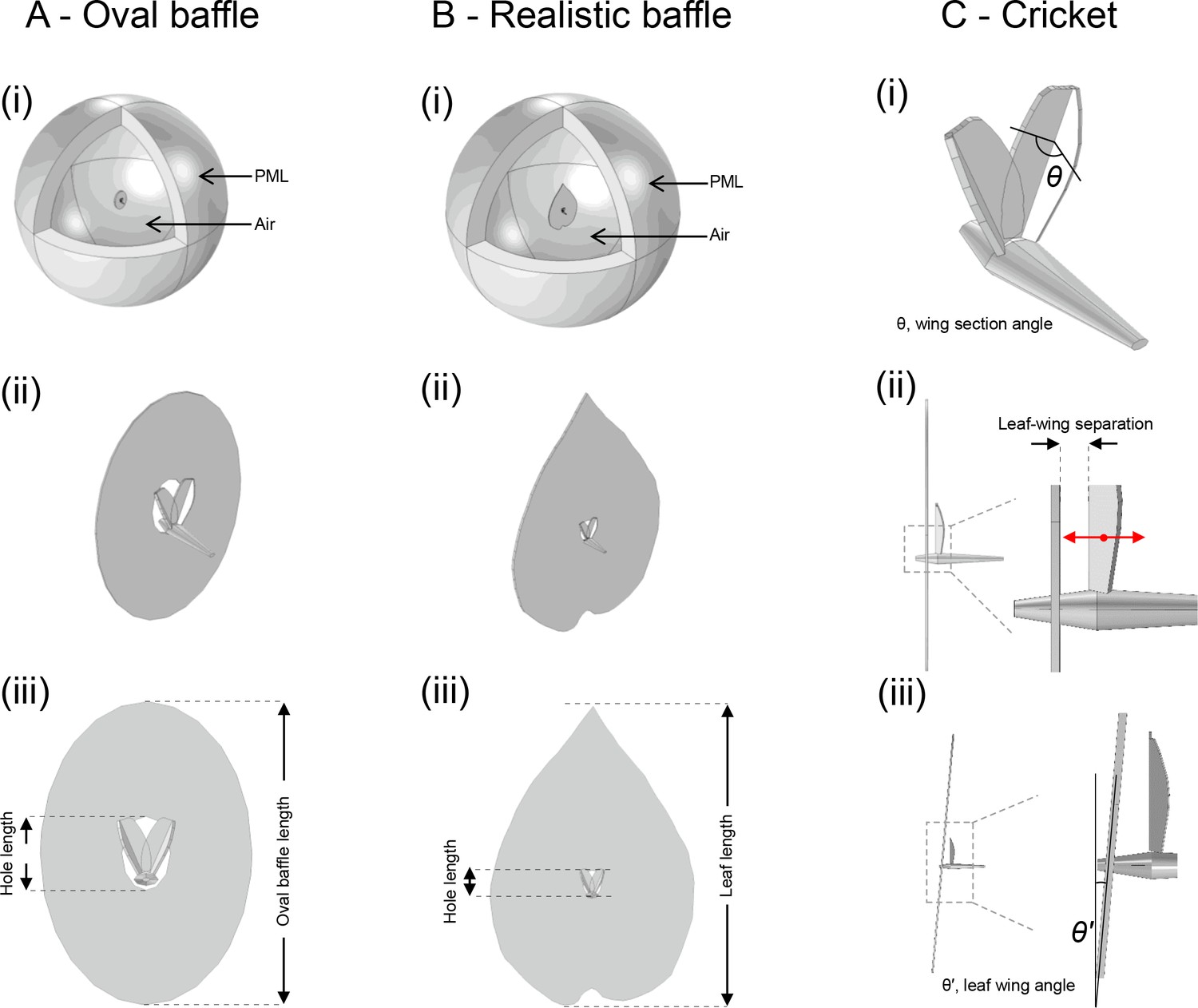
Finite element model of cricket calling from idealized and realistic baffles.
(A) (i) Cricket and oval baffle in air surrounded by perfectly matched layer (PML). (ii) Relative position of cricket in oval baffle. (iii) Definitions of oval baffle dimensions as used in manuscript. (B) (i) Cricket and realistic leaf in air surrounded by perfectly matched layer (PML). (ii) Relative position of cricket in realistic leaf baffle. (iii) Definitions of realistic leaf baffle dimensions as used in manuscript. (C) (i) Geometry of cricket body and forewings showing wing section angle (costal angle), real cricket value was approximately 137°. (ii) Wing to leaf separation distance, real cricket values was approximately 1 mm. Red arrow shows direction of prescribed velocity condition. (iii) Leaf to wing tilt angle, shown here as −5° tilt for illustration (the tilt was maintained at 0° in the models).
Figure 2 with 1 supplement

The sound field predicted by finite element modeling from an unbaffled and baffled cricket.
A y-z plane cut through the 3D sound field radiated by the virtual FEA cricket (A) calling in free-space and (B) through an oval baffle (baffle length x width: 69 × 41 mm; hole length x width: 17 × 11 mm). The cricket’s body aligns with the z-axis and the wing-plane is in the x-y plane as depicted in the inset. The pressure field is represented by iso-amplitude surfaces that join all points in space with identical dB SPL. Baffling reduces acoustic short-circuiting and enhances SPL around the hole, resulting in higher far-field radiation.
Figure 2—figure supplement 1
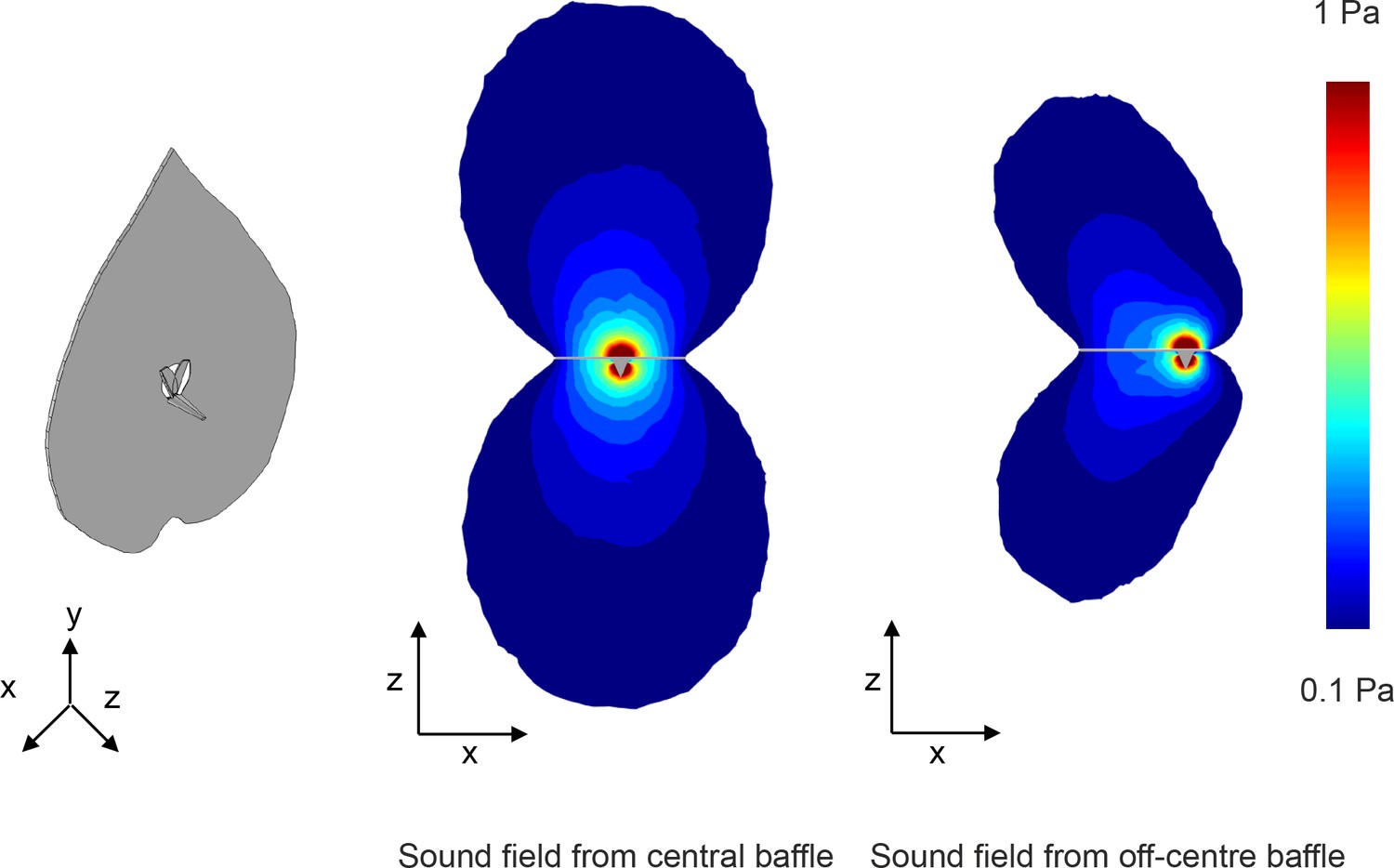
Effect of baffle hole position on the shape of the resulting sound field.
We show absolute pressure plots in the z-x plane, with the plane passing through the center of the baffle hole. In the off-center scenario the magnitude of the absolute pressure field is reduced due to the acoustic short circuiting effect becoming more significant at the closer edge.
Figure 3 with 1 supplement
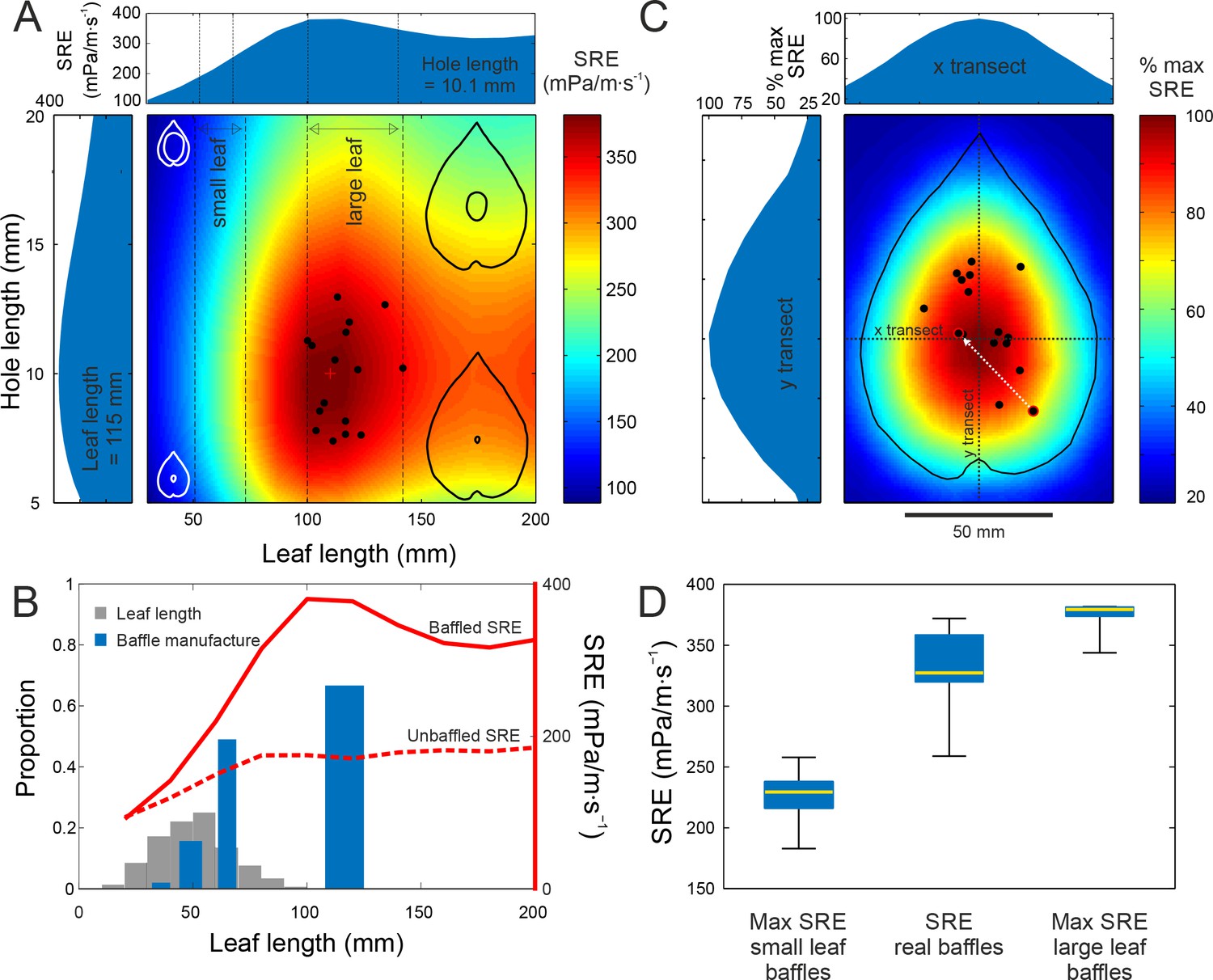
Baffle design efficiency and optimization behaviour.
(A) FEA predicted SRE representing the effect of different leaf and hole size combinations (cartoons not to scale), overlaid with data from cricket-made baffles (black dots). Leaf and hole lengths are used as a proxy for size. In nature, as in our models, leaves retain their aspect ratio at different sizes, and length is an adequate measure. Vertical stippled lines mark the ranges of small and large leaf sizes offered to the crickets during choice experiments. Side graphs show SRE at average leaf size (115 mm) and average hole size, which is also the optimal size (10.1 mm). The red cross marks the optimal baffle. (B) Blue bars depict the proportion of males that made baffles on different leaf sizes in a no-choice experiment. The width of the bars depict the size range of the leaves in that size class. Grey bars depict the distribution of natural leaf sizes by depicting the proportion of leaves that fall into different size classes (N = 570 leaves). The two red lines depict the SRE associated with baffled calling (solid line), and with unbaffled calling from the leaf edge (wings parallel to the leaf surface, stippled line) at different leaf sizes. (C) Baffles created on real leaves were normalized to the average large leaf (length = 115 mm) and overlaid onto a map showing the percent SRE at different hole positions in relation to the maximum. Percent maximum SRE along horizontal and vertical transects are shown on side graphs. One male made two holes, first an eccentric, suboptimal one, followed by another at a more efficient location (white arrow). (D) Distributions of maximum SRE achievable on small and large leaves offered to the males, with SRE distributions of real baffles made by males (n = 15). Box and whisker plot depicts median (red line), the 25th and 75th percentile (blue box) and the range (whiskers).
Figure 3—figure supplement 1
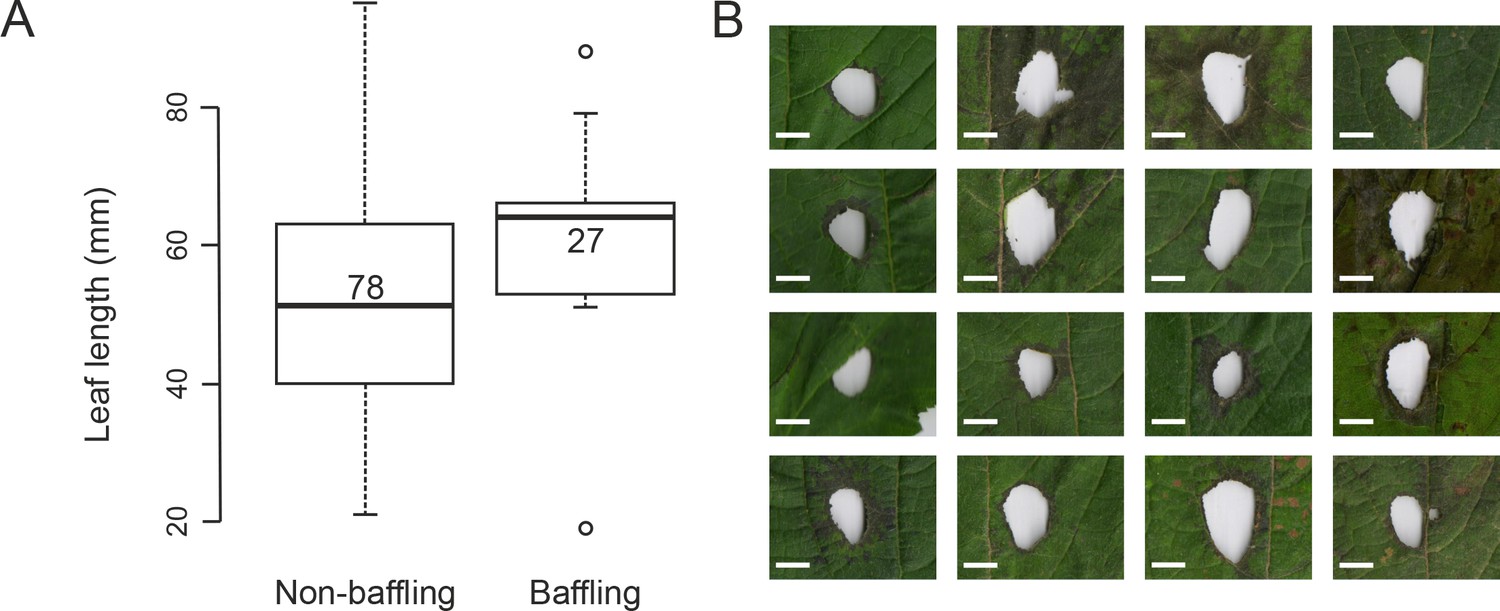
Baffle making behaviour and its natural context (A) Box-plot showing the distributions of leaf size used by wild non-bafflers and by baffling males in the field.
The leaves used by both sets of males for calling were larger than the most commonly found leaves in the wild. The leaves used by the baffling males were, however, bigger than those used by non-baffling males, suggesting that males actively select larger leaves for making baffles (Mann-Whitney U test; Length: W = 668, p<0.005, numbers in boxes are the sample sizes). (B) All the baffle holes made on large H. suaveolens leaves by the male tree crickets (scale bars: 10 mm). Note that the holes are always placed between large leaf veins.
Figure 4
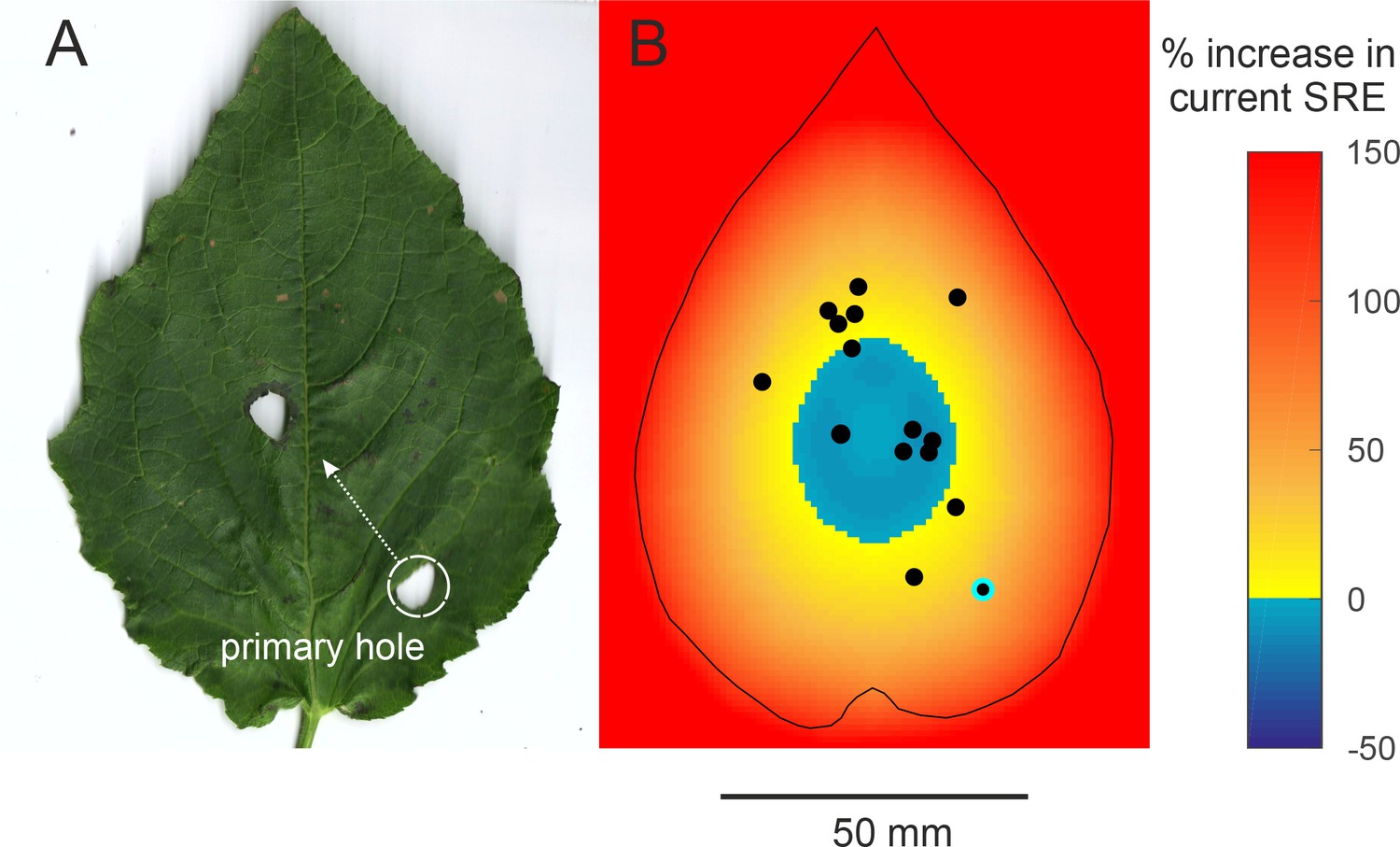
Efficiency of two-holed baffles (A) The baffle of a male who made a very eccentric primary hole and then cut another hole at a more central location.
The SRE of a two-holed baffle such as this is difficult to predict a priori. A central baffle hole is more efficient, but two holes allow more acoustic short-circuiting. (B) An SRE map of a two-holed baffle was made to address this question. In this scenario, the primary hole can be positioned anywhere on the leaf but the second hole is always at the optimal central position. The colour on the map depicts the percent increase in SRE that a male at that position can accrue by making a second optimally positioned hole. SRE increases only if the primary hole was eccentric; if the primary hole was close to the optimal position, however, making a second hole in effect reduces the baffle’s SRE. The dots depict the positions of the holes of real males. The primary hole of the only male with the two-holed baffle is highlighted.
Videos
Video 1
Finite element analysis simulation of instantaneous sound pressure radiated by unbaffled and baffled calling cricket.
https://doi.org/10.7554/eLife.32763.003
Video 2
Video of male O. henryi making a baffle and calling from it.
https://doi.org/10.7554/eLife.32763.007Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.32763.013
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Tree crickets optimize the acoustics of baffles to exaggerate their mate-attraction signal
eLife 6:e32763.
https://doi.org/10.7554/eLife.32763




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}