Shaping of inner ear sensory organs through antagonistic interactions between Notch signalling and Lmx1a
- University College London, United Kingdom
Figures
Figure 1 with 1 supplement
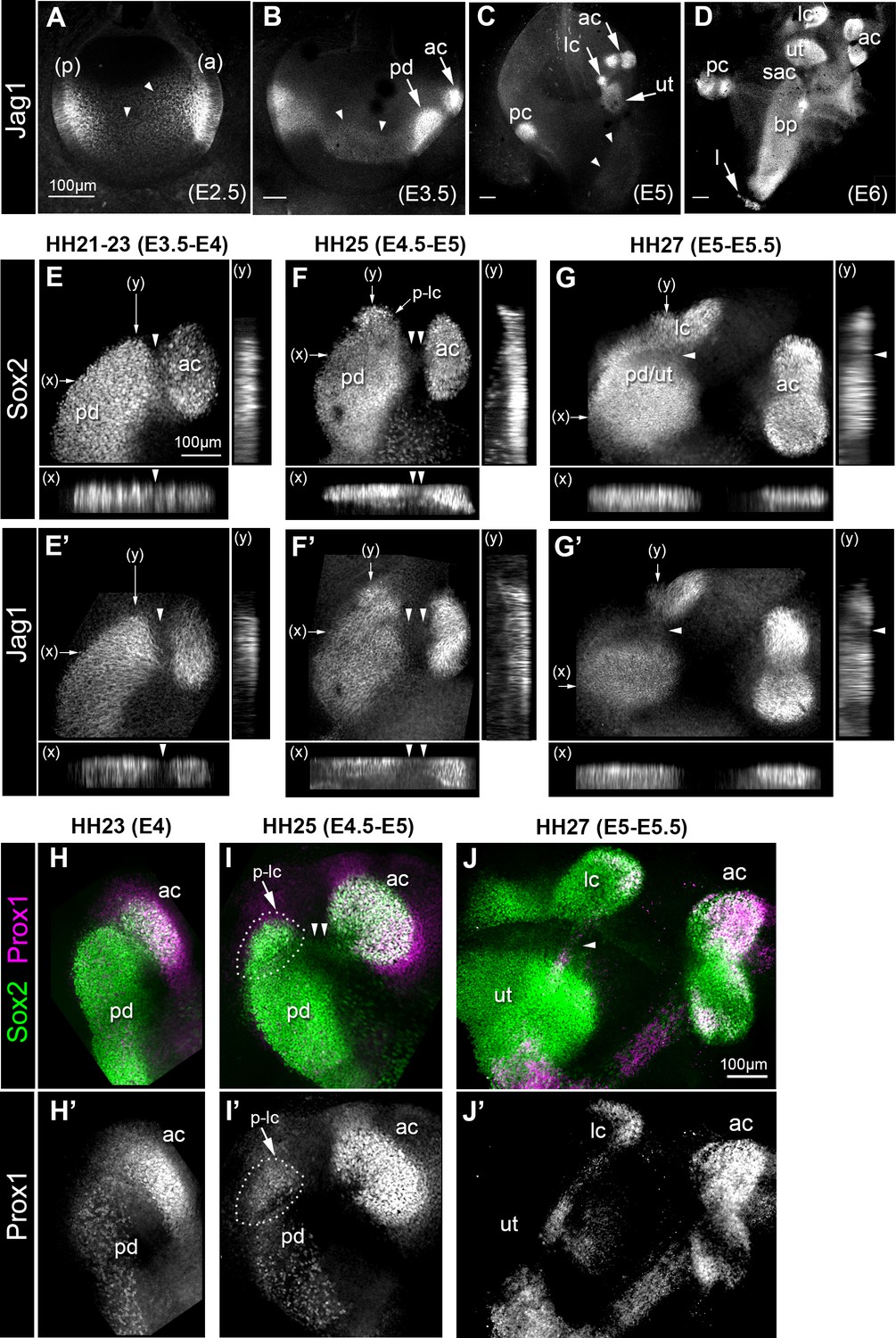
Expression of the Notch ligand Jag1,Sox2 and Prox1 during the formation and segregation of the chick inner ear sensory patches.
(A–D) Jag1 expression in the developing chick inner ear (whole-mount preparations). At E2.5 (A), two patches of high expression are present in the anterior (a) and posterior (p) poles of the otocyst, but lower levels of expression are also detected in the medial region along the antero-posterior axis (arrowheads). At E3.5 (B), two patches of high Jag1 expression are now present in the anterior domain (arrows), corresponding to the anterior crista (ac) and a larger prosensory domain (pd); the medial domain of lower expression remains clearly visible (arrowheads). At E5 (C), strong Jag1 staining is observed in the posterior (pc), anterior and lateral (lc) cristae and the utricle (ut); another fainter domain of expression extends ventrally within the developing cochlear duct (arrowheads). At E6 (D), the utricle, the three cristae, and the lagena (l) are well defined. Jag1 expression still forms a continuous domain of expression between the saccule (sac) and the basilar papilla (bp). (E–G’) Whole-mount surface views and transverse projections (along the x and y directions) of the anterior prosensory domains at different developmental stages, immunostained for Sox2 and Jag1 expression. At HH21-23 (E–E’), the prospective anterior crista becomes apparent at the edge of a large prosensory domain (pd); a group of Sox2-positive and Jag1-negative cells (arrowheads) is located at the interface of the two prosensory patches. At HH25 (F–F’), the distance separating the anterior crista from the large prosensory domain increases (double arrowheads); a distinct cluster of Sox2-positive cells becomes visible at the upper edge of the large prosensory domain, which corresponds to the location of the prospective lateral crista (p–lc). At HH27 (G–G’), the lateral crista starts to segregate from the prospective utricle (pd/ut); the interface domain is Sox2-positive, but exhibits a marked down-regulation of Jag1 expression (arrowheads). At HH23 (H–H’), Prox1 is strongly expressed in the prospective anterior crista (ac), which is still connected to the large prosensory domain (pd). At HH25 (I–I’), strong Prox1 immunostaining is present in the anterior crista; a new, denser cluster of Prox1-positive nuclei (dotted outline) is also present at the superior edge of the large prosensory domain, presumably corresponding to the prospective lateral crista (p–lc). Note that the cells located in between the anterior crista and the large prosensory domain still retain faint Sox2 expression (arrowheads in I). At HH27 (J–J’), the lateral crista (lc) segregates; note that only one of its halves contains Prox1-expressing cells at this stage. Scale bar for all panels = 100 μm.
Figure 1—figure supplement 1
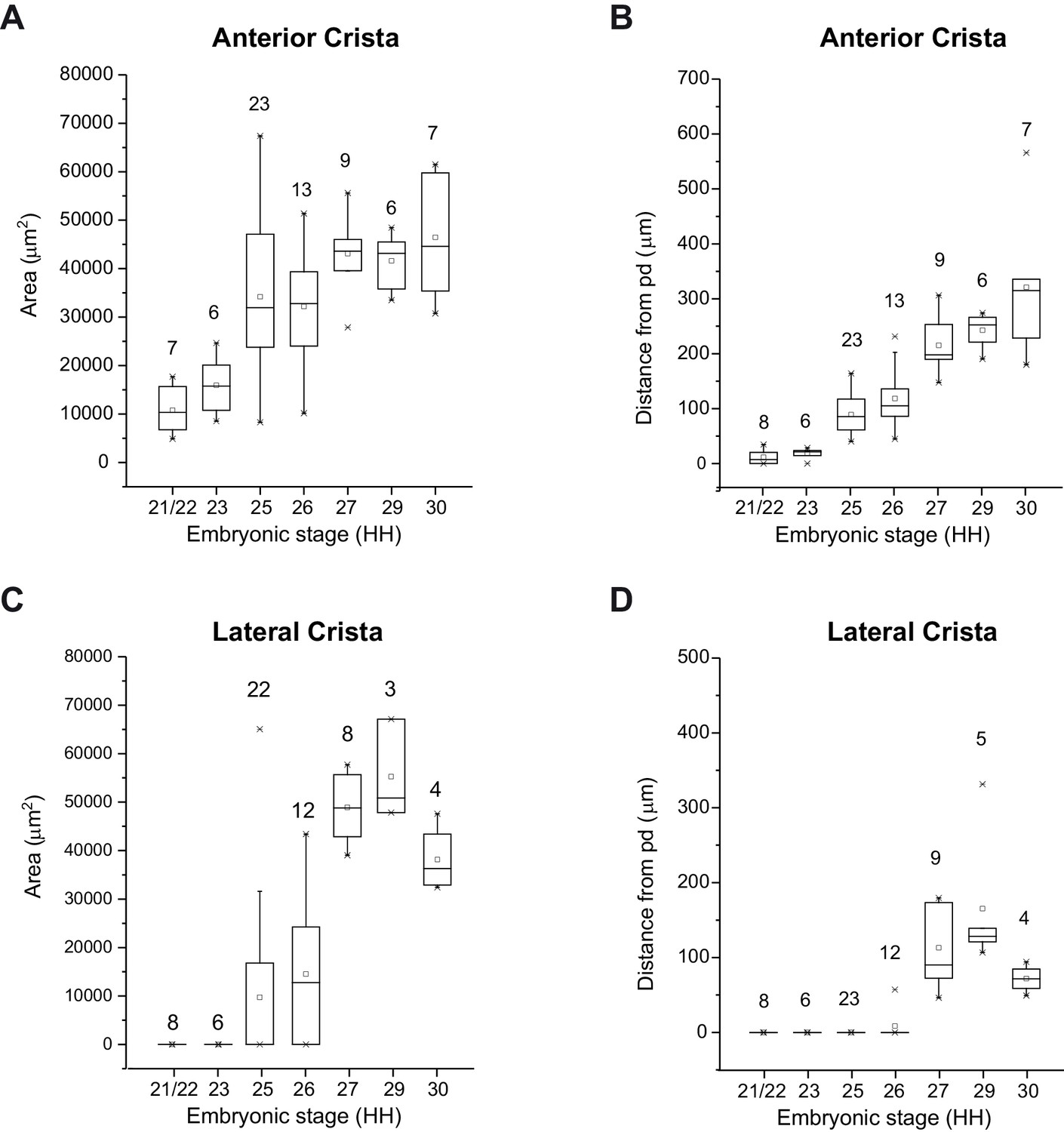
Quantification of anterior and lateral cristae segregation in the embryonic chick inner ear.
Box plots representing the surface area of the anterior (A) and lateral (C) cristae and the distance separating these from the large prosensory domain (pd) (B, D) at different developmental HH stages. The numbers indicated correspond to the number of samples analysed for each stage. Both cristae increase in size before physically separating from the large prosensory domain (prospective utricle).
-
Figure 1—figure supplement 1—source data 1
Measurements of the surface area of the anterior and lateral cristae and the distance separating them from the anterior prosensory domain as a function of the developmental stage (Hamburger and Hamilton stages) for each sample analyzed.
- https://doi.org/10.7554/eLife.33323.004
Figure 2 with 1 supplement
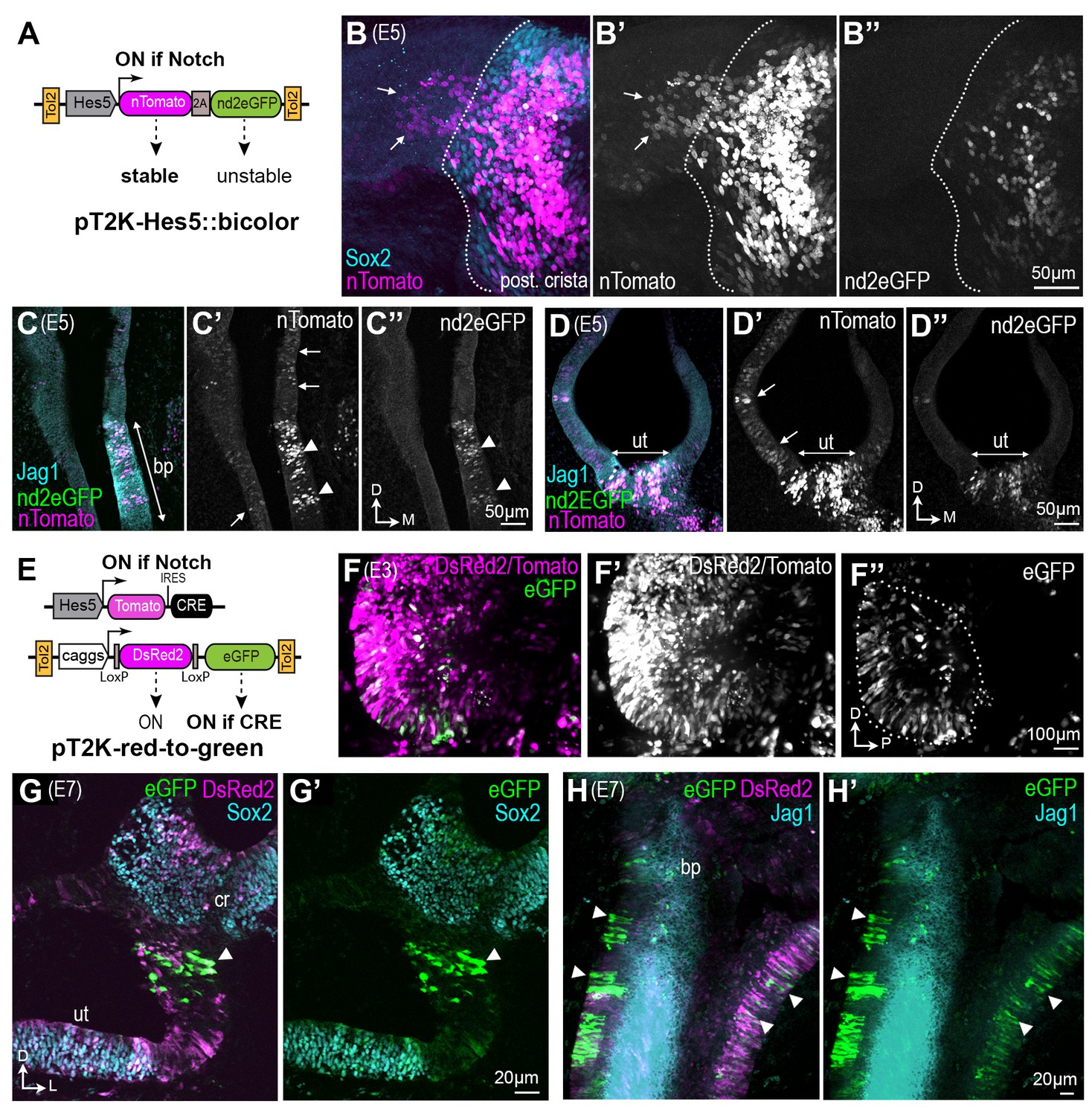
Some otic cells turn off Notch activity and adopt a non-sensory fate during sensory patch formation.
(A–D’’) Identification of ‘Notch-OFF’ cells using a bi-color Notch-sensitive Hes5 reporter as a molecular timer. (A) The Hes5::bicolor Tol2 construct contains the Hes5 promoter, regulating expression of a stable nTomato and a destabilized nd2eGFP. (B–B’’) Posterior crista (whole-mount) analysed 72 hr after electroporation with the Hes5::bicolor transposon and stained for Sox2 (cyan) to identify sensory progenitor cells. Note the presence of ‘Notch-OFF’ cells (arrows) with low levels of nTomato (magenta in B; gray in B’) but no nd2eGFP fluorescence (green in B, gray in B’’), in the non-sensory domain that flanks the ventral border of the crista (dotted line). Transverse section of an E5 cochlear duct (C–C’’) and (D–D’) utricle transfected with the Hes5::bicolor construct and immunostained for Jag1 (cyan). Notch-active cells (arrowheads) are located within the sensory patches (bp = basilar papilla; ut = utricle). Note the presence of ‘Notch-OFF’ cells, with nTomato fluorescence only, outside of the sensory patches (arrows in C’ and D’). (E–H’) Long-term genetic labelling of the early Notch-active cells using Cre-mediated recombination and a red-to-green Tol2 construct. (E) The Hes5::Tomato-IRES-Cre plasmid drives transient expression of the Tomato fluorescent protein and Cre recombinase in Notch-active cells. The pT2K-red-to-green Tol2 transposon drives constitutive expression of either DsRed2, or alternatively eGFP after Cre-mediated recombination. (F) Whole-mount view of an E3 chick otocyst 16 hr after co-electroporation with Hes5::Cre and PT2K-red-to-green. The majority of transfected cells are DsRed2 fluorescence only (F’), but a subset of cells, located in the anterior domain of the otocyst, exhibit eGFP fluorescence (outline in F’’). (G–G’) Transverse section through the vestibular system of a transfected chick inner ear at E7, immunostained for Sox2 (cyan). In this example, a group of eGFP-expressing cells (arrowhead) are present in the non-sensory domain separating the utricle (ut) from the crista (cr). (H–H’) Whole-mount preparation of the cochlear duct of a pT2K-red-to-green transfected chick inner ear at E7, immunostained for Jag1 (cyan). Note the presence of several groups of eGFP-expressing cells in the non-sensory domains surrounding the basilar papilla (arrowheads).
Figure 2—figure supplement 1
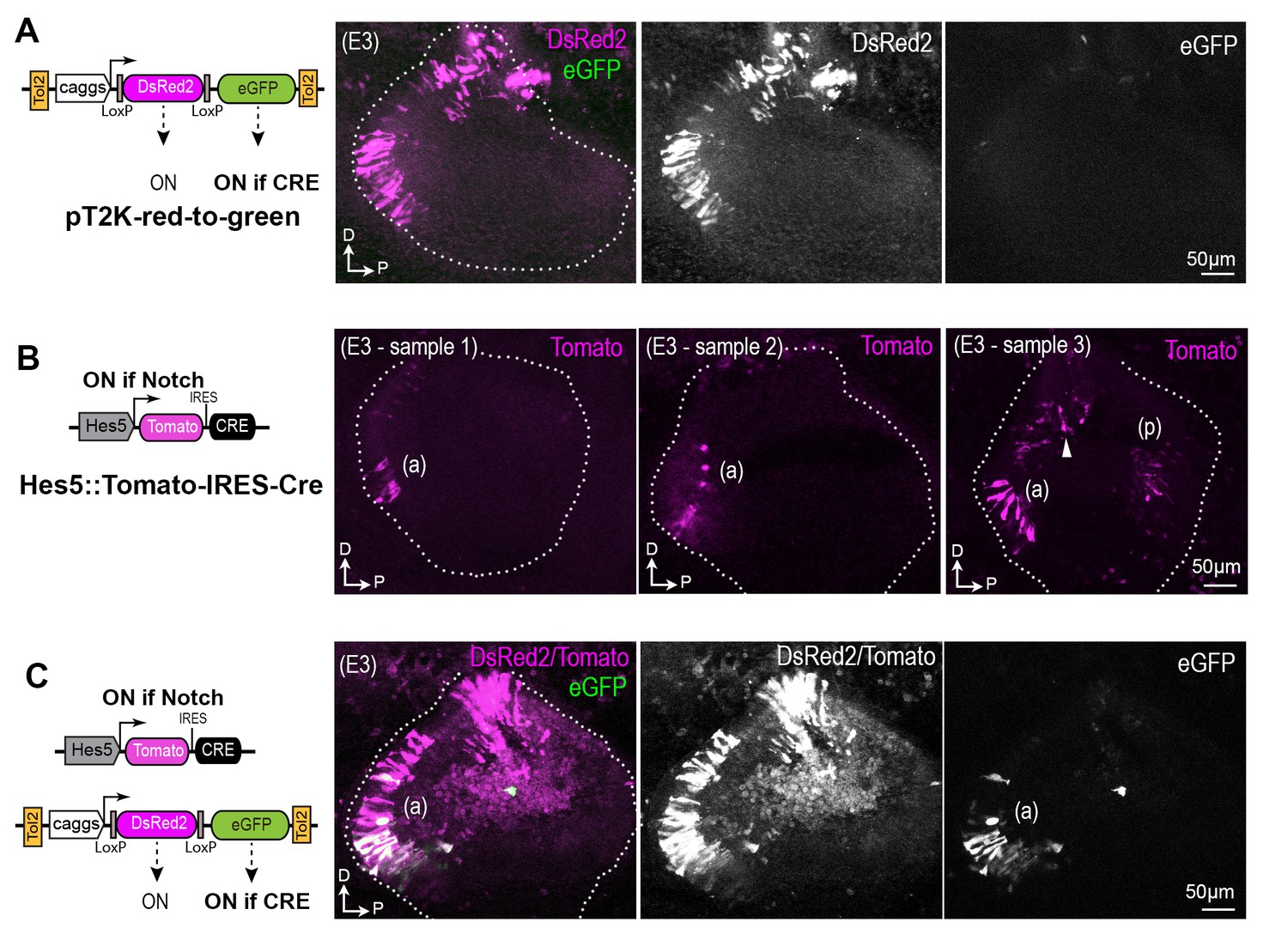
Representative whole-mount views of chicken otocysts 16 hr after electroporation with different combinations of pT2K-red-to-green and Hes5::Tomato-IRES-Cre constructs.
(A) Example of sample transfected at E2 with the pT2K-red-to-green Tol2 transposon, which drives constitutive expression of either DsRed2, or alternatively eGFP after Cre-mediated recombination. In the absence of Cre expression, there is no GFP fluorescence. (B) The Hes5::Cre plasmid drives transient expression of the tdTomato red fluorescent protein and Cre recombinase in cells experiencing Notch activity. Three examples of transfected samples are shown, with induction of strong Tomato expression in the anterior (a) and posterior (p) domain of the otocyst. Note that some fluorescent cells are also visible in a more dorsal and anterior location in sample 3, which could potentially be associated to the formation of the anterior crista. (C) After co-electroporation with both the Hes5::Tomato-IRES-Cre and the PT2K-red-to-green, the cells exhibiting GFP fluorescence are located in the anterior (a) domain of the otocyst, indicating successful recombination in a Notch-responsive manner.
Figure 3
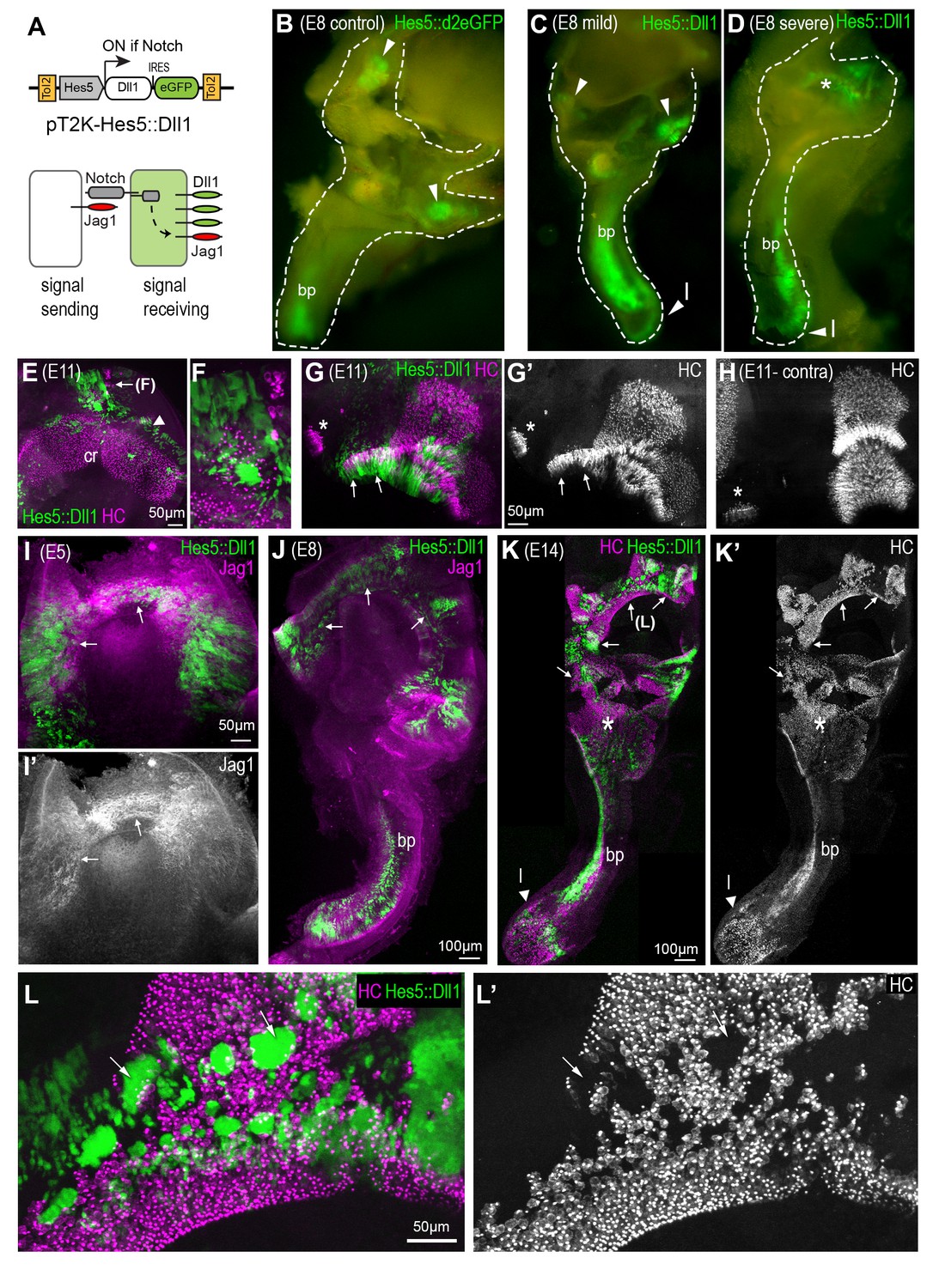
A gain of lateral induction disrupts the patterning and boundaries of inner ear sensory patches.
(A) Schematic of the pT2K-Hes5::Dll1 Tol2 construct used to induce an artificial gain of lateral induction. The Hes5 promoter drives expression of Dll1, along with eGFP, in transfected cells in which Notch is active. (B–D) Dissected E8 inner ears transfected with either a control Hes5::d2eGFP (B) or a Hes5::Dll1 (C–D) Tol2 construct at E2.5. In the control (B), eGFP fluorescence is detected in distinct sensory patches, in this case the basilar papilla (bp) and two cristae (arrowheads). Transfection with Hes5::Dll1 induces mild (C) or severe (D) defects in the morphogenesis of the vestibular system, and abnormal activation of eGFP expression in the dorsal regions (asterisk in D). (E–H) whole-mount preparations of E11 Hes5::Dll1 transfected cristae, immunostained with HCA and myosin 7A antibodies (in magenta) to identify hair cells. Note the formation of ectopic hair cells (E–F) or abnormal expansion of the posterior crista (arrows in G–G’) towards the macula neglecta (asterisk) compared to the contralateral untransfected posterior crista (H). (I–I’) Jag1 expression is induced (arrows) in the dorsal portion of an otocyst with strong activation of the Hes5::Dll1 construct. (J) Example of an E8 inner ear with ectopic induction of Jag1 and Hes5::Dll1 activity in the dorsal region of the vestibular system.(K–K’) Whole-mount of an E14 inner ear with widespread Hes5::Dll1 induction, immunostained with HCA and myosin 7A antibodies (in magenta). The vestibular system contains ectopic and fused sensory domains (arrows). Hair cells are also present in between the basilar papilla (bp) and the saccule-utricle region (asterisk), which appear to form a continuous sensory patch. (L–L’) Higher magnification view of the dorsal vestibular region of the sample shown in (K). Hair cell density is reduced inside the clusters of transfected cells with strongest eGFP fluorescence (arrows).
Figure 4
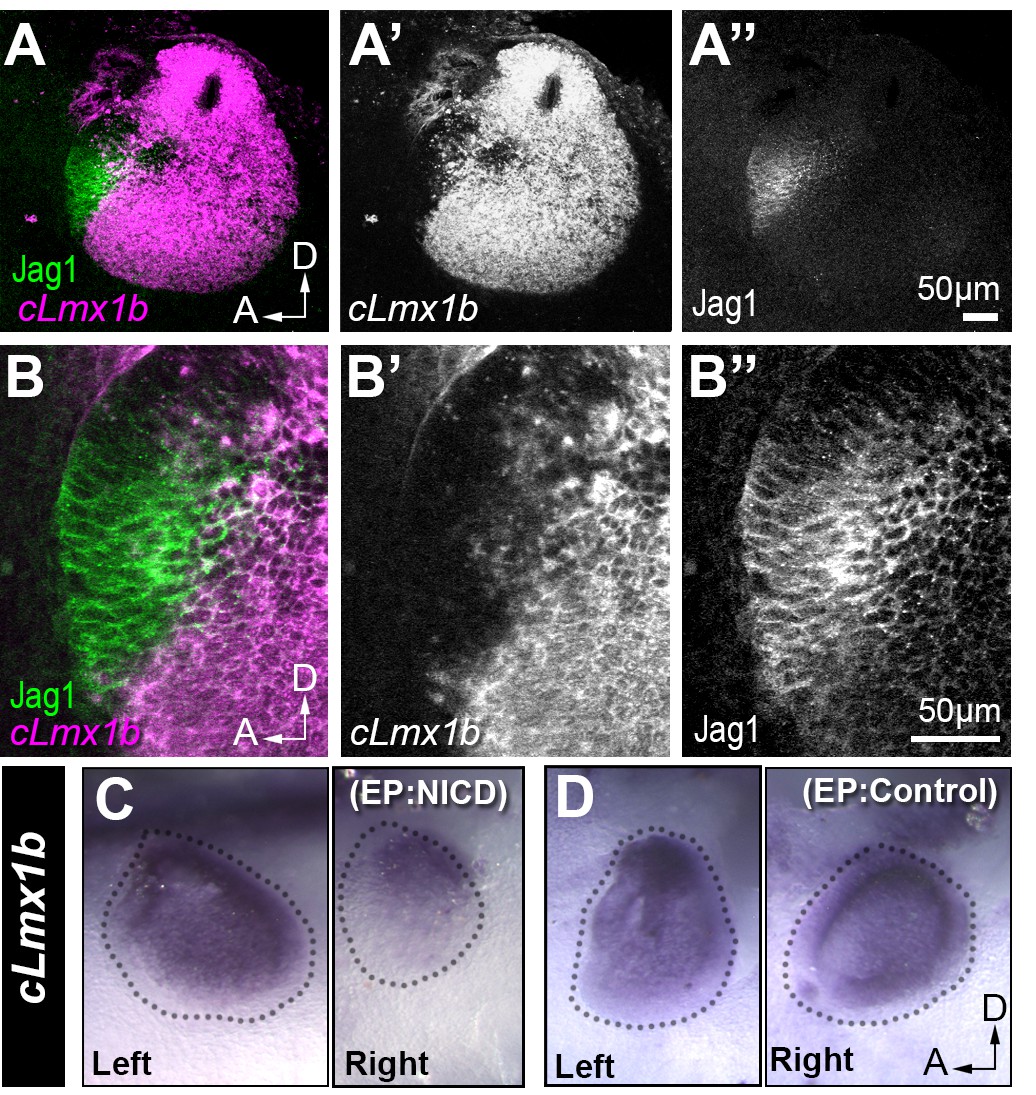
Jag1 and cLmx1b are expressed in complementary domains and Notch inhibits cLmx1b expression in the chick otocyst.
(A–B’’) Fluorescent in situ hybridization for cLmx1b (magenta) and immunostaining for Jag1 (green) in the E3 chick otocyst. (A, A’’) Jag1 is expressed in a complementary manner to cLmx1b. (B–B’’) High magnification view of the anterior region of the otocyst shown in (A). (C–D) Whole-mount in situ hybridization for cLmx1b 24 hr after in ovo electroporation with plasmids encoding either NICD (C) or a control red fluorescent protein (D) used at the same concentration. Note the reduction in cLmx1b expression in the NICD-transfected sample (C, right) in comparison to the untransfected contralateral otocyst (C, left) or to the otocyst transfected with a control plasmid (D, right).
Figure 5

Over-expression of cLmx1b down-regulates expression of Jag1.
(A–A’’) Whole mount preparation of an E4.5 chick utricle after electroporation with the pT2K-Lmx1-eGFP Tol2 construct and immunostaining for Jag1. Note the irregular contours of the utricular macula and the reduced expression of Jag1 in transfected cells (asterisk). (B–B’’) Whole mount view of an Lmx1-eGFP transfected chick inner ear at E7. Jag1 expression is reduced or completely absent in cLmx1b-overexpressing cells, resulting in abnormal positioning of the boundaries of the utricle (ut) and saccule (sac) (arrows in B’–B’’,) see also Video 1 for a 3D animation of a high magnification view of this region) and a large truncation (dotted line) of the basilar papilla (bp). (C) Surface view of an Lmx1-eGFP transfected E7 basilar papilla with several groups of cLmx1b-overexpressing cells (arrowheads). At higher magnification (D–D’’), note the rounded appearance of the clusters of transfected cells (D’) and the absence of Jag1 expression in those cells (D’’). (E–E’’) Surface view of an eGFP (control) transfected E7 basilar papilla. There is no disruption in the expression pattern of Jag1 or the positioning of its lateral borders.
Figure 6
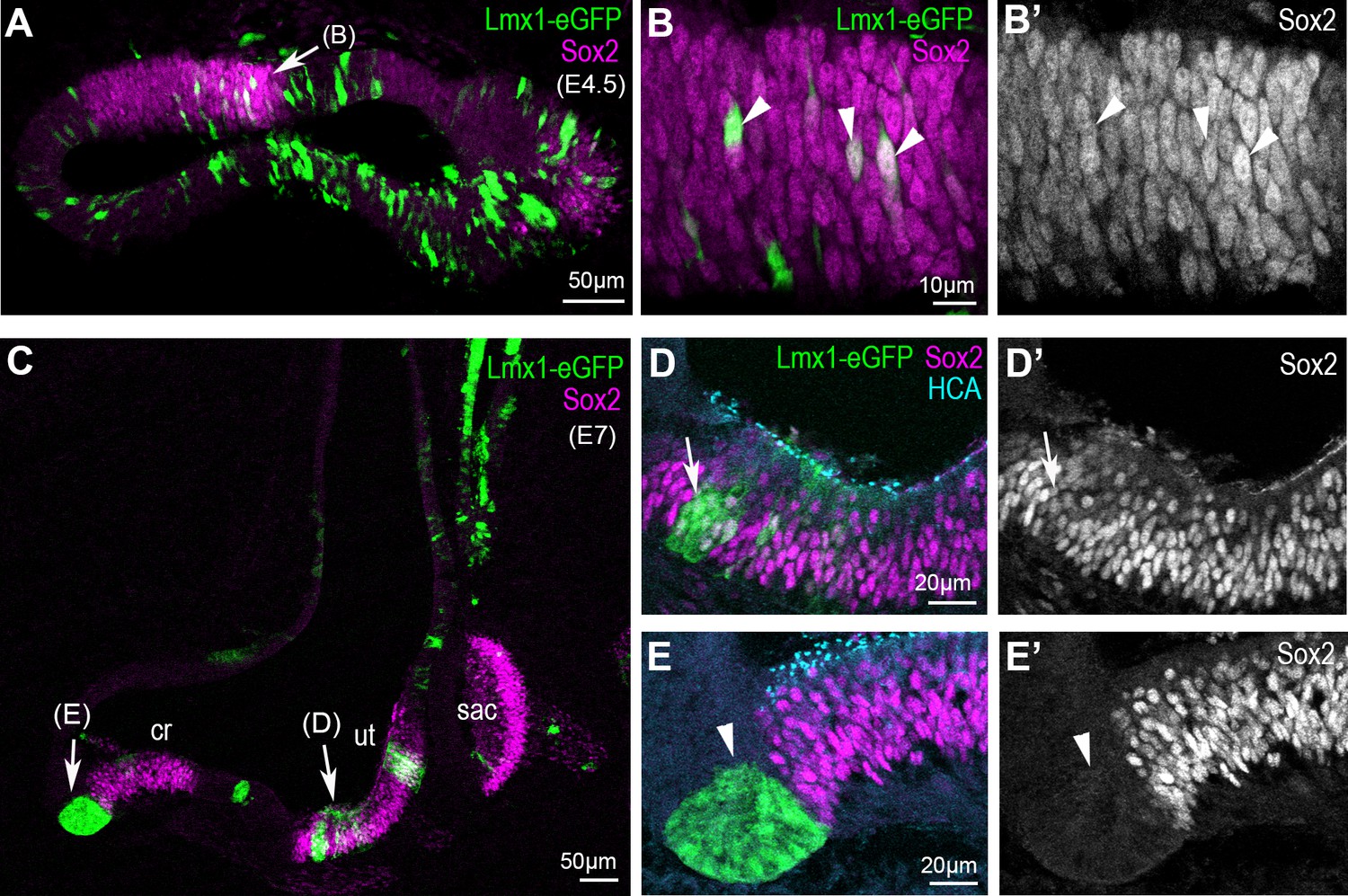
Overexpression of cLmx1b down-regulates Sox2 expression in a context-dependent manner.
(A–E’) Transverse views of the developing chick inner ear following electroporation with pT2K-Lmx1-eGFP and immunostaining for Sox2 (magenta) and HCA (cyan). (A) An E4.5 sample with two vestibular sensory patches containing transfected cells. At high magnification (B–B’’), note that the levels of Sox2 are unchanged in cLmx1b-overexpressing cells (arrowheads) compared to neighbouring untransfected cells (B–B’). (C) Low magnification view of an E7 sample, with the utricle (ut), saccule (sac) and anterior crista (cr) visible. (D–D’) High magnification view of the utricle, containing cLmx1b-overexpressing cells that retain Sox2 expression (arrows), although at a reduced level compared to neighbouring untransfected cells. (E–E’) A cluster of transfected cells is abutting the anterior crista, and show a complete absence of Sox2 expression. Note also the apical constriction of the transfected cells (arrowheads).
Figure 7
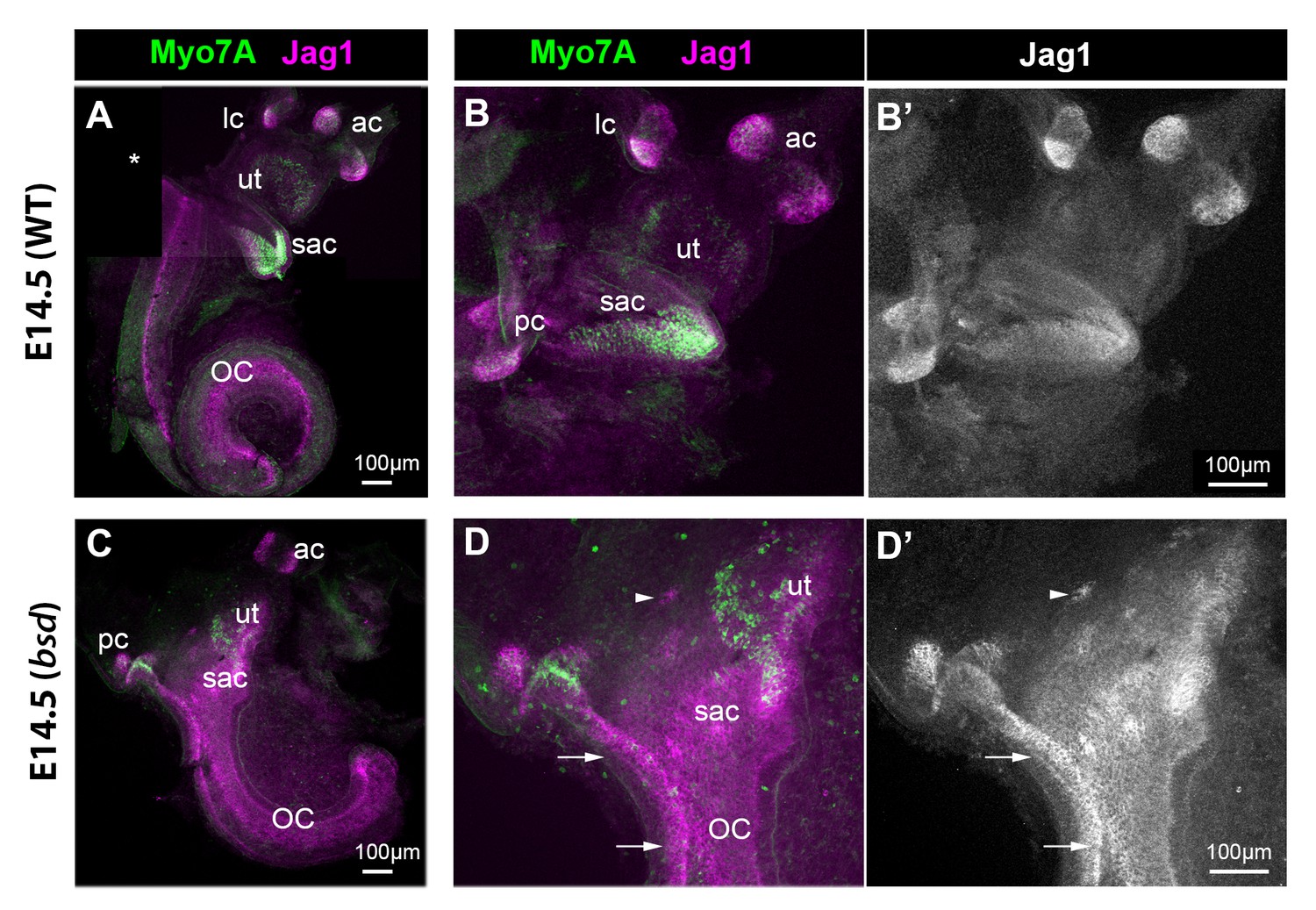
Jag1 expression is expanded in the inner ear of E14.5 bsd mutant mice.
Whole-mount preparations of the inner ear of E14.5 wild-type and bsd mice, immunostained for Jag1 and the hair cell marker myosin 7A (Myo7A). In the wild-type (A–B’), several patches of jag1 expression corresponding to the distinct vestibular organs and the organ of Corti are visible. Highest levels of expression are found in the cristae. In the bsd mutant (C–D’), Jag1 expression is expanded throughout the vestibular system, with occasionally patches of cells exhibiting higher expression levels (arrowhead in D–D’). Individual patches are difficult to identify, with the exception of the anterior and posterior cristae due to their position and partial segregation. Note the continuity in Jag1 expression between the posterior crista and the organ of Corti (arrows in D–D’) and the absence of segregation between the saccule and utricle domains. Abbreviations: lateral (lc), anterior (ac) and posterior (pc) cristae; utricle (ut); saccule (sac); organ of Corti (OC).
Figure 8 with 1 supplement

Sox2 expression in the developing inner ear of Lmx1aGFP/+ mice.
Whole-mount preparations of the developing inner ear of Lmx1aGFP/+ mice. At E8, Sox2 expression is present in the anterior (a) and posterior (p) prosensory domains of the otocyst, which also exhibit reduced Lmx1a/GFP expression (A–A’’). At higher magnification, cell mixing between GFP+ and Sox2+ cells (arrows in B–B’’) is visible at the posterior border of the anterior prosensory domain. At E10.5 (C–C’’), Sox2 expression is present in a large anterior prosensory domain (the prospective utricle, labelled pd/ut) and more ventrally in a group of cells that could correspond to the prospective saccule (sac). Some Lmx1a/GFP cells with reduced Sox2 expression are present in between the utricle and the saccule (arrows). By E12.5 (C–C’’), Sox2 is expressed in distinct sensory patches in the vestibular system, while GFP is present in between the sensory patches. In the utricle (ut), residual GFP expression is present in the anterior-most domain (star in C’) and in the cells facing the cristae; at the posterior edge of the utricle, Sox2+ cells are GFP-negative, and directly abut the non-sensory domain with strongly GFP+ cells (arrowheads in C’). Note the overlap between Sox2 and GFP expression at the borders of the anterior (ac) and lateral (lc) cristae (arrows in C) and in the saccule region (sac).
Figure 8—figure supplement 1
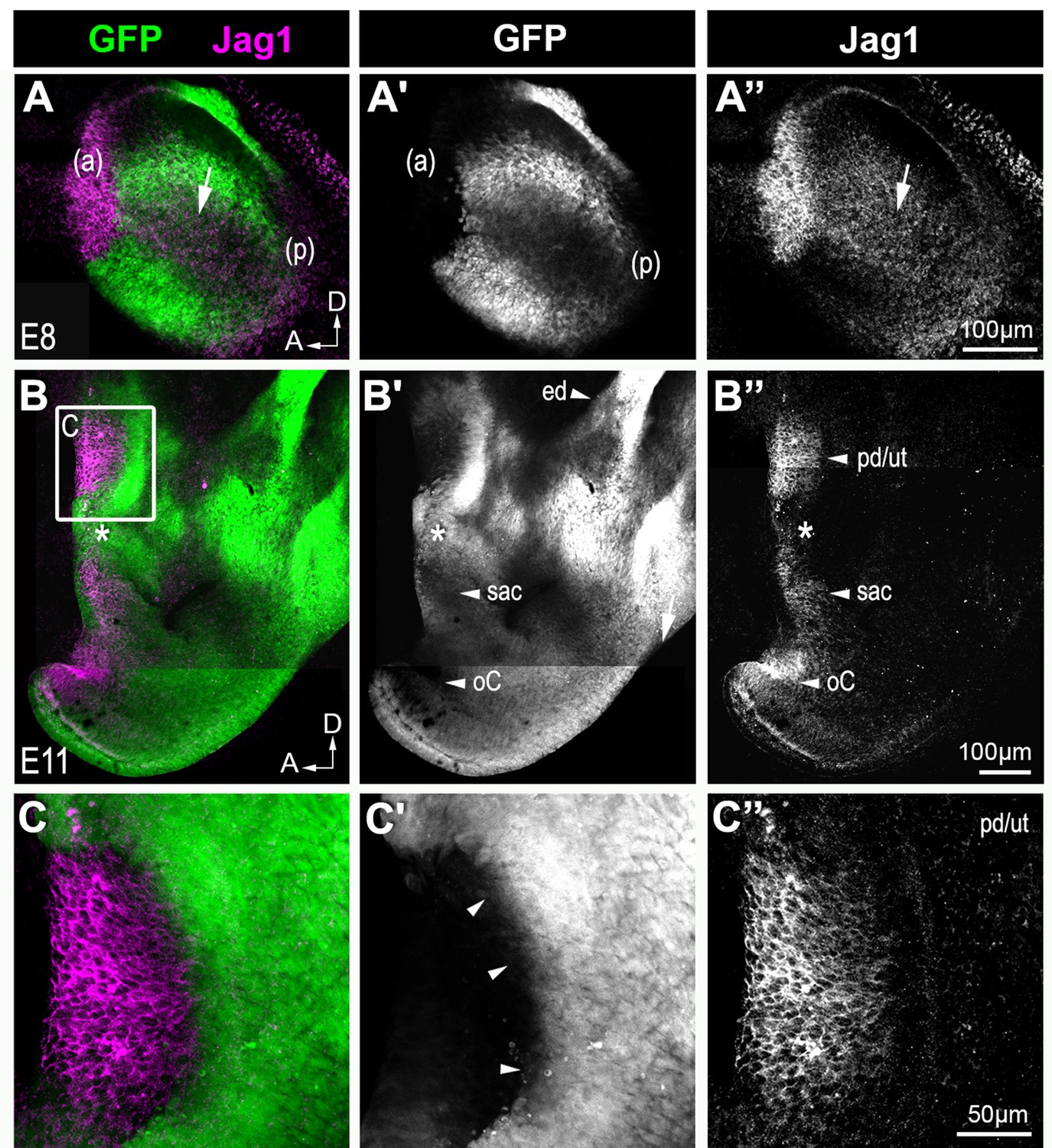
Jag1 expression in the developing inner ear of Lmx1aGFP/+ mice.
Whole-mount preparations of the inner ears from Lmx1aGFP/+ mice at E8 (A–A’’) and E11 (B–C’’). In the E8 otocyst, Jag1 is strongly expressed in the anterior prosensory domain (ap), where no GFP fluorescence is detected. Jag1 is expressed in a mode diffuse manner in the medial region (arrow in A’’) and posterior prosensory domain (pd) of the otocyst, where there is partial overlap with GFP expression. At E11 (B–C’’), several patches of Jag1 expression, corresponding to the prospective utricle (pd/ut), the prospective saccule (sac) and the organ of Corti (oC) are visible and separated by GFP+ cells. Note that GFP expression overlaps partially with Jag1 expression in between the prospective utricle and saccule (stars in B–B’’). There is also strong GFP expression in the endolymphatic duct (ed in B’), in the dorsal most region of the inner ear. In the prospective utricle (C–C’’), there is no GFP expression in the Jag1+ domain and the boundary of the GFP+ non sensory domain is very sharp (arrowheads in C’).
Figure 9
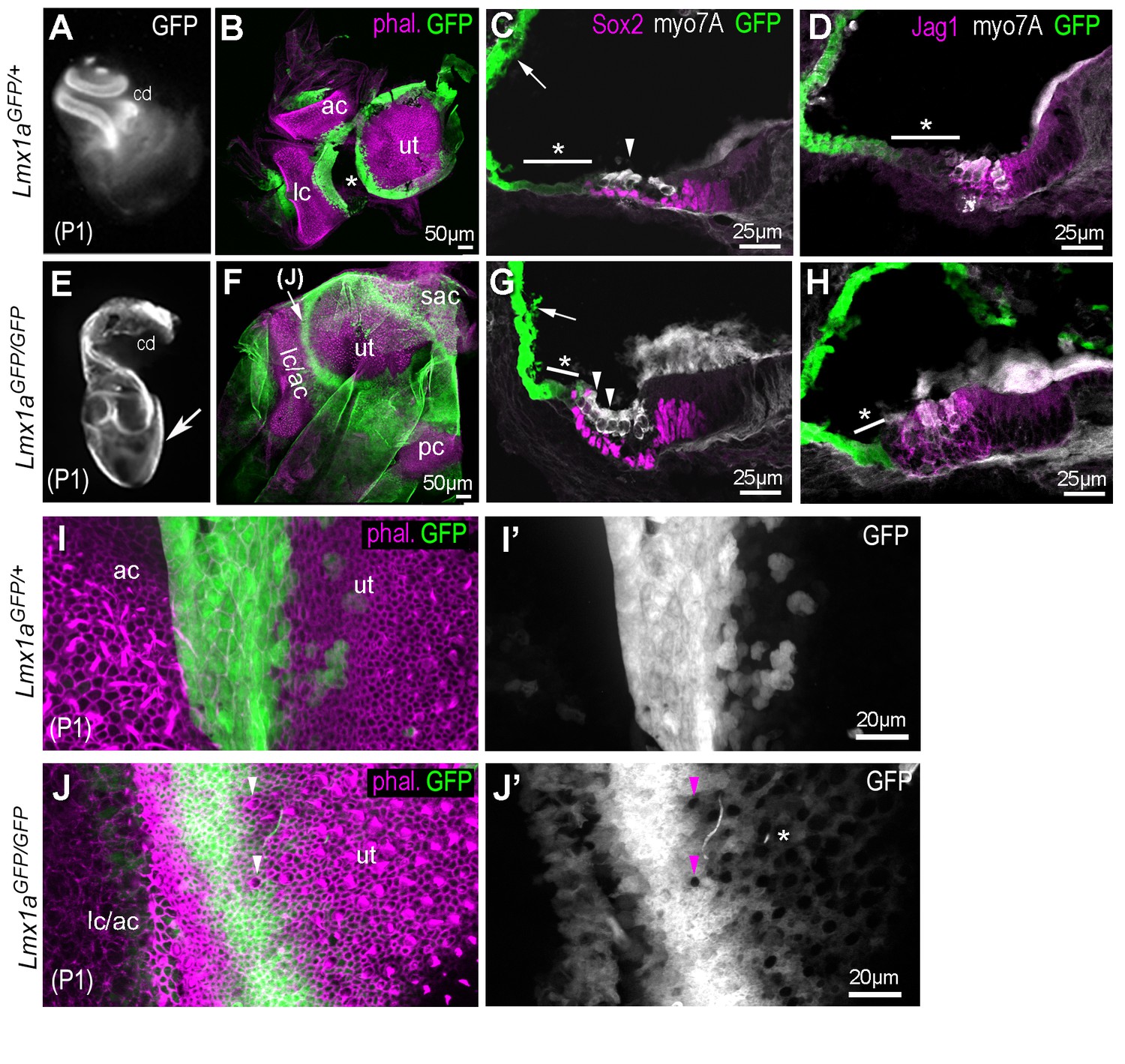
Inner ear morphogenesis is severely disrupted in homozygous Lmx1aGFP/GFP knock-in mice.
In heterozygous P1 Lmx1aGFP/+ (A–D), inner ear morphology is normal and GFP expression is found at known sites of Lmx1a expression: in the lateral wall of the cochlea (A, arrow in C), in the domains separating the utricle (ut) from the anterior (ac) and lateral (lc) cristae (B; asterisk indicate artefactual interruption of GFP due to dissection). (C–D) Transverse section through the cochlear duct of P1 Lmx1aGFP/+mice. The lateral domain of the organ of Corti (asterisk in C–D), composed of the Hensen and Claudius cells, exhibit reduced levels of GFP fluorescence compared to the lateral wall. In the Lmx1aGFP/GFP double knock-in mice (E–H), inner ear morphology is severely disrupted: the vestibular system forms a cyst-like structure exhibiting high levels of GFP fluorescence (arrow in E) and the cochlear duct is reduced in size. (F) Some GFP-expressing domains are still present around the utricular macula (ut), the fused anterior and lateral cristae (lc/ac) and the posterior crista (pc) regions. (G–H) The organ of Corti contains additional rows of Myo7a-positive hair cells in the lateral domain (arrowheads in G), but the expression of Sox2 and Jag1 remains confined to the GFP-negative domain. The region abutting the lateral border of the sensory domain (asterisk in G–H) exhibits lower levels of GFP than the lateral wall (arrow in G), but is smaller in comparison to that of the Lmx1aGFP/+ cochlea (C–D). (I–J’) higher magnification surface view of the region separating the utricle from the lateral crista in Lmx1aGFP/+ (I–I’) and in Lmx1aGFP/GFP (J–J’) mice. Hair cells are identifiable by their actin-rich bundle of stereocilia. In the Lmx1aGFP/GFP vestibular system, some hair cells devoid of GFP fluorescence (arrowheads in J–J’) are present inside the GFP-positive domain in between the utricle and the lateral/anterior cristae region. Note also the presence of weakly GFP fluorescent cells intermingled with hair cells in the lateral part of the utricle (asterisk in J’).
Figure 10 with 1 supplement
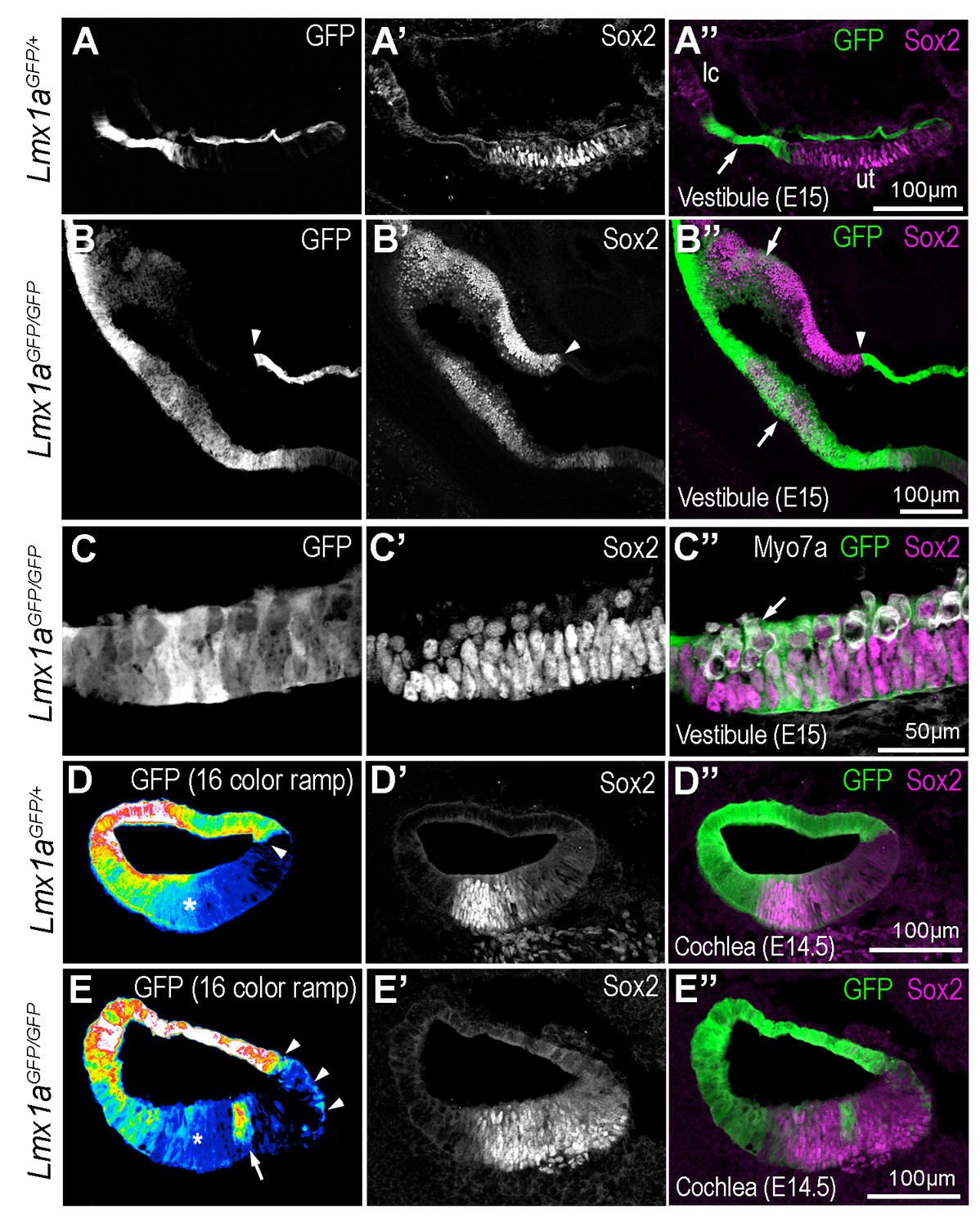
Some cells of the Lmx1a lineage are diverted towards a sensory fate in the absence of Lmx1a function.
Transverse section through the vestibular system and cochlear duct of embryonic Lmx1aGFP/+ and Lmx1aGFP/GFP mice, immunostained for Sox2 and myosin 7A. (A–A’’) cross-section through the vestibular system of E15 Lmx1aGFP/+ samples. The GFP-positive cells are located in between the utricle (ut) and lateral crista (lc) expressing Sox2, and in the roof of the utricle. In Lmx1aGFP/GFP littermates (B–C’’), Sox2 expression is expanded but the identity of vestibular patches is difficult to ascertain. At least some of the boundaries between Sox2+ and Sox2- domains coincide with that of GFP expression (arrowheads in B–B’’). On the other hand, there is some overlap between GFP and Sox2 expression (arrows in B’’) in some sensory territories. (C–C’’) High magnification view of a vestibular patch with expression of GFP and Sox2. Some hair cells (arrows) are also present. (D–D’’) Sox2 expression in the cochlear duct of an E14.5 Lmx1aGFP/+ mouse. At the medial border of the developing organ of Corti (arrowhead), cells with either high or low levels of GFP fluorescence (displayed as a 16-color ramp in D–E) form a clear interface. At the lateral border, GFP fluorescence decreases progressively, with high levels in the lateral wall and low levels within the Sox2-positive prosensory cells (asterisk). In Lmx1aGFP/GFP littermates (E–E’’), the lateral gradient of GFP fluorescence is irregular (asterisk) and some cells with fairly high levels of GFP fluorescence are present inside the Sox2-positive domain (arrow in E). At the medial border, GFP cells are also intermingled with GFP-negative cells (arrowheads in E).
Figure 10—figure supplement 1
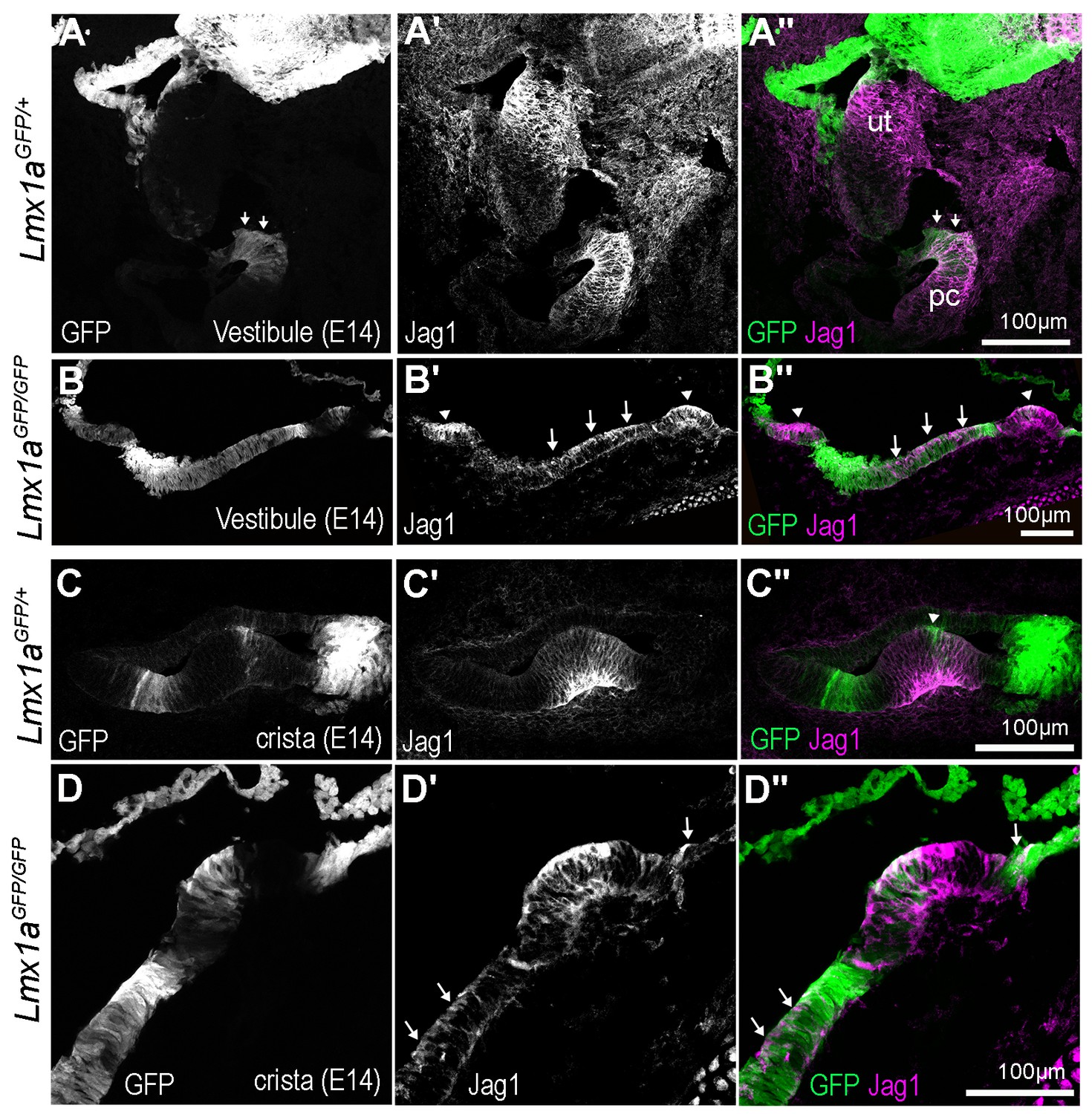
Jag1 expression in the vestibular system of E14 Lmx1aGFP/+ and Lmx1aGFP/GFP mice.
(A–A”) Cross section through the vestibular system of an E14 Lmx1aGFP/+ mouse. Jag1 immunostaining is elevated in the utricle and anterior crista region, whilst GFP is present in the roof of the utricle (ut), and in between the crista and utricle (arrows). (B–B”) In the Lmx1aGFP/GFP mice, two crista-like patches (arrowheads) with strong Jag1 expression are detected, but the region with medium/low levels of GFP fluorescence separating them contains Jag1-expressing cells (arrowheads). (C–C”) High magnification view of a crista in Lmx1aGFP/+ mice, showing the clear separation between GFP-positive and Jag1-positive cells, with the exception of the eminentia cruciatum (arrowheads), a region that is devoid of hair cells. (D–D”) Higher magnification view of the sample shown in (B–B’’). Note the ectopic expression of Jag1 within the GFP-positive domains bordering this crista.
Figure 11
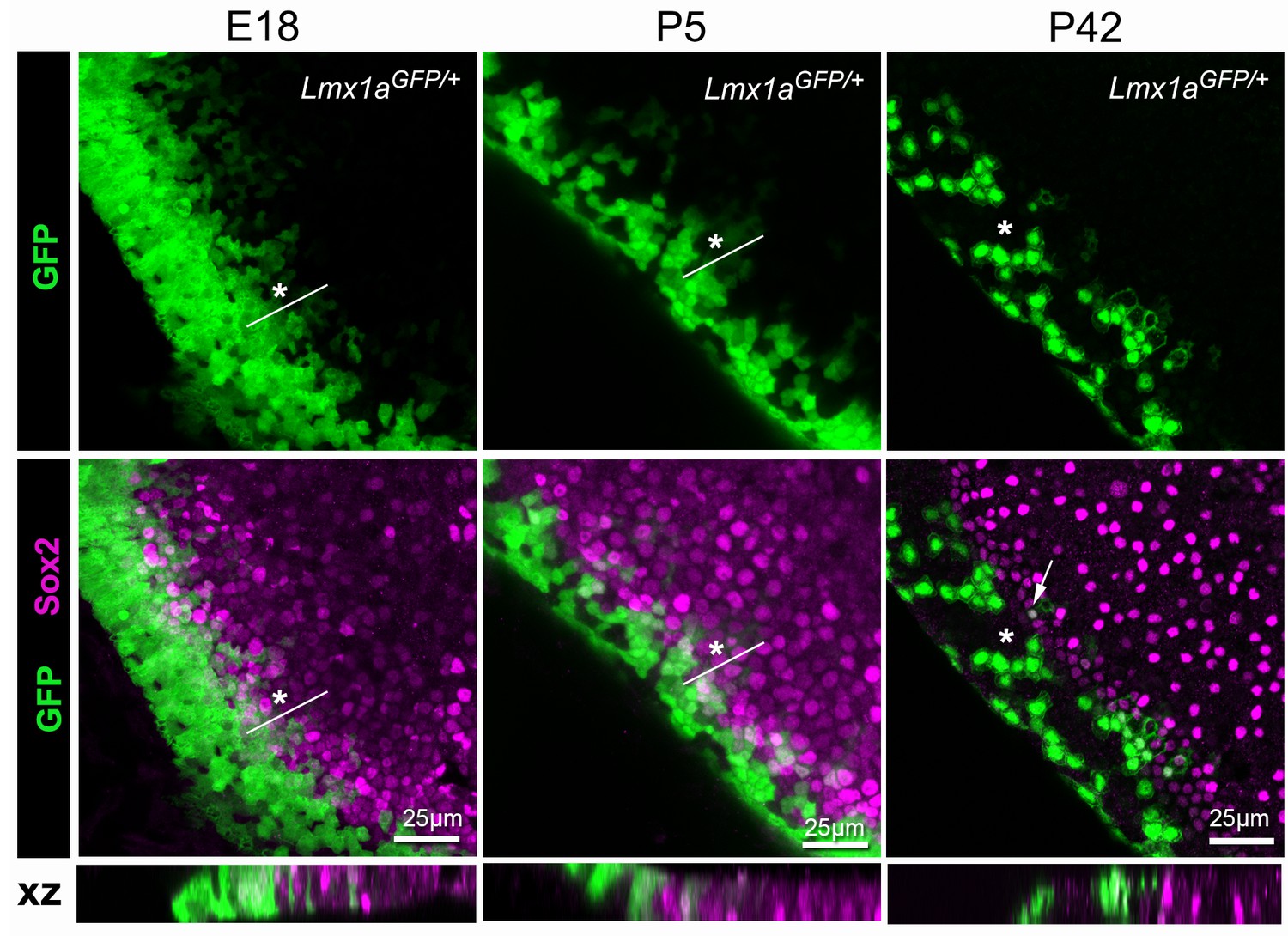
Lmx1a expression is progressively down-regulated at the lateral border of the developing utricle.
Maximum intensity z-projections of whole-mount utricles from Lmx1aGFP/+ mice at different developmental ages (E18, P5 and P42), immunostained for Sox2 expression (magenta). Note the mosaic pattern of GFP fluorescence at the lateral border of the utricle, and the overlap between GFP+ and Sox2+ cells at E18 and P5 (asterisk). In the adult (P42) utricle, only a few cells retained low levels of GFP fluorescence and Sox2 expression (arrow) in the sensory domain, whilst mosaic expression of GFP is still observed in the transitional cell region (asterisk). The XY images are orthogonal projections taken from the same preparations.
Figure 12
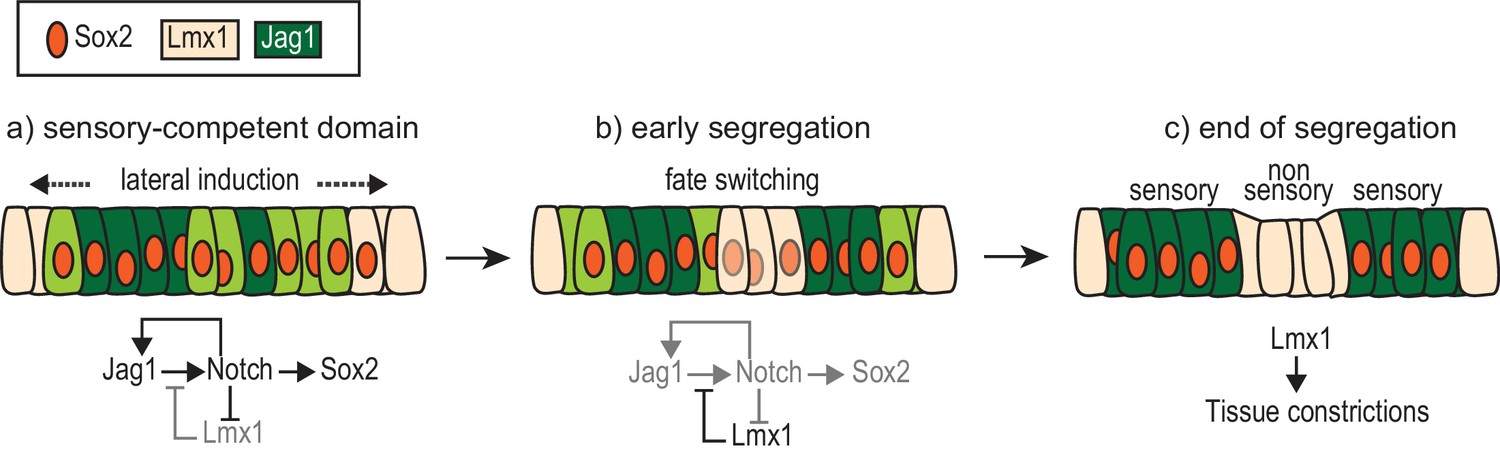
A model of the interactions between Notch signalling and Lmx1a during sensory patch segregation.
Initially (a), the cells of the ‘pan’ sensory-competent domain express Sox2 as well as Jag1 (strong = dark green or low = light green), which promotes its own expression and adoption of the prosensory fate (and therefore Sox2 expression) by lateral induction. Within the pan-sensory domain, Lmx1a expression is repressed by Notch activity. At its border, a dynamic competition takes place between lateral induction/prosensory specification and the factors promoting Lmx1a expression. (b) The segregation of a prosensory domain could be due to a localized reduction in lateral induction within the pan-sensory domain; this could gradually lead to an upregulation of Lmx1 and a conversion (fate switching) of sensory-competent cells into non-sensory cells at the interface of segregating prosensory patches. (c) As cells commit to their definitive fate, Lmx1 could lead to the formation of the tissue constrictions that separate adjacent sensory organs. The regulatory interactions depicted are likely to involve intermediate factors.
Videos
Video 1
3D Volocity rendering of an E7 chicken inner ear electroporated at E2 with cLmx1b-eGFP and immunostained for Jag1 expression (magenta).
https://doi.org/10.7554/eLife.33323.010Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.33323.020
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Shaping of inner ear sensory organs through antagonistic interactions between Notch signalling and Lmx1a
eLife 6:e33323.
https://doi.org/10.7554/eLife.33323




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}