Point of View: Predictive regulation and human design
- Perelman School of Medicine, University of Pennsylvania, United States
- Article
- Figures and data
- Abstract
- Introduction
- Cellular origins of human computational capacity
- Multi-cellularity expanded resources and computational capacity
- To occupy the world H. sapiens required a large, efficient brain
- Expanding the community’s computational capacity
- What led to our present difficulties?
- Predictive regulation in a regime of shrinking positive surprise
- Alternative hypothesis to explain the planetary rise in obesity
- What level to treat?
- Conclusions
- References
- Decision letter
- Author response
- Article and author information
- Metrics
Abstract
Organisms evolving toward greater complexity were selected across aeons to use energy and resources efficiently. Efficiency depended on prediction at every stage: first a clock to predict the planet’s statistical regularities; then a brain to predict bodily needs and compute commands that dynamically adjust the flows of energy and nutrients. Predictive regulation (allostasis) frugally matches resources to needs and thus forms a core principle of our design. Humans, reaching a pinnacle of cognitive complexity, eventually produced a device (the steam engine) that converted thermal energy to work and were suddenly awash in resources. Today boundless consumption in many nations challenges all our regulatory mechanisms, causing obesity, diabetes, drug addiction and their sequelae. So far we have sought technical solutions, such as drugs, to treat complex circuits for metabolism, appetites and mood. Here I argue for a different approach which starts by asking: why does our regulatory system, which evolution tuned for small satisfactions, now constantly demand 'more'?
https://doi.org/10.7554/eLife.36133.001Introduction
Homo sapiens is now beset by multiple difficulties. Atmospheric carbon dioxide is rising and destabilizing the climate. Obesity is increasing – although multitudes still live in extreme poverty – accompanied by increases in cases of hypertension and type 2 diabetes. Addiction to opioids and other drugs is also rising, resulting in more deaths from overdose. Medical science continues to advance but, at the same time, the costs of treating an increasingly unhealthy population continue to rise. These problems seem, prima facie, separate. After all, what could possibly connect climate change to obesity, or obesity to drug addiction?
Here I suggest that these difficulties arise from one profound tension: a conflict between how we evolved to live versus how we live now. For 200,000 years we were governed by a close matching of resources to needs. We, like all our progenitors, were shaped by nature’s broadest constraint: natural selection favors organisms that gather and use energy efficiently. Everything, from proteins and cells up to organs and the behavior of whole organisms, evolved to be efficient – and to be efficiently regulated by an efficient brain (Sterling and Laughlin, 2015). But then in a flash – the past 250 years – everything changed and many of us now live amidst resources that greatly exceed our biological needs. Although such wealth might seem an unalloyed blessing, it may be also problematic.
What aspect of the design of H. sapiens has delivered us to our present state? The short answer, I argue here, is our preeminent computational capacity (that is, our superior intelligence). Once unleashed, it led to a particular event that promptly derailed our species. If we could grasp the enormity of that event and its connection to our present ills, we might see broadly what recovery would require. Here I sketch the growth of the computational capacity H. sapiens to show how it has been constrained at every stage by energy and to identify the key adaptations to that constraint.
Cellular origins of human computational capacity
Human computational capacity began to unfold nearly four billion years ago with tiny cells – prokaryotes – containing DNA whose structure encoded information for assembling proteins that catalyze the chemical reactions that supply energy and synthesize all the materials needed to reproduce the cell. In essence, the cell is an analogue computer where all information is processed by chemistry (Bray, 2009). Analogue is most efficient because the processes are graded and thus matched to avoid waste: just the right concentration of each reactant to produce products that will be just right to serve as new reactants – to make the next products, and so on. Prokaryotes accomplish these functions at an energy cost approaching the lower limit set by physical law (Noor et al., 2010).
Cellular chemistry is powered by a small molecule, adenosine triphosphate (ATP), that transfers a fixed packet of energy to another molecule, thereby activating its function. The energy boost provided by ATP is small, about 20 kBT (where kB is the Boltzmann constant, and T is temperature) in one millisecond, and modestly exceeds the mean energy of thermal noise. Yet, because a protein molecule is continually energized by thermal noise, the energy boost provided by ATP is enough to tilt the protein from 'almost certainly not active' to 'almost certainly active' (Astumian, 1997; Astumian, 2015; Motlagh et al., 2014). A larger energy boost would be wasteful, so the size of the packet of energy provided by ATP is near optimal. This may explain why prokaryotes adopted ATP as the universal energy donor.
Prokaryotes produced ATP by burning sugars to drive an extraordinary nanoscale turbine (ATP synthase) embedded in the cell membrane. This turbine spins at 9,000 rpm, and each revolution spits out three ATP molecules (He et al., 2018). The turbine is 90% efficient, meaning that 90% of the energy that goes into the turbine is 'captured' by ATP (Kinosita et al., 2000). In short, the proteins in a cell operate with maximal economy: they are excited by no-cost thermal noise, boosted by energy packets of optimal size that are synthesized with near optimal efficiency.
Yet prokaryotes remained tiny – just a few micrometers in diameter. Their genomes also remained small and did not accumulate information. They had encoded all the genes they could afford energetically, so when new genes were needed to adapt to novel conditions, cells were forced to shed their nonessential genes (Lane, 2014). What restrained these early cells, both physically and computationally, was the location of their power plant in the cell membrane. Were a cell to enlarge, its membrane would increase as the square of its diameter, but its volume would increase as the cube. Thus, until a cell could expand its power plant, it could not afford to enlarge (Lane, 2014).
Then occurred an event so improbable that it was never repeated (Lane, 2014). One bacterium invaded another and took up residence. The host provided nutrients and the guest supplied energy-bearing compounds needed to support its own information system. As long as the host and guest remained independent there was no advantage to the host because the guest used all its energy to reproduce. But gradually the guest transferred most of its genes to the host genome, retaining only key genes needed for substrate oxidation. Now that the guest was a power plant with no household to support, it could multiply without limit and increase the host’s energy capacity – by up to 100,000-fold.
The guest, having ceded its passport (most of its genome) to the host cell, became a permanent resident, an obligatory cell organelle: the mitochondrion. Cells powered by mitochondria could now balloon from 3 micrometers in diameter to 300 micrometers, 100-fold greater diameter and 10,000-fold greater volume. These cells – eukaryotes – added 3,000 new gene families along with a more complex and expensive system of gene splicing. Moreover, eukaryotes were under less selective pressure to prune genetic material added by gene duplication or viruses. Consequently, odd sequences accumulated – like bits of wire and old screws in a capacious toolbox. Such fragments were exploited over the next 1.5 billion years by eukaryotes relieved of the prokaryote’s energy constraint (Lane, 2014).
Unicellular organisms eventually reached hard limits with respect to the capture of resources and information. First, they were constrained from foraging widely by the viscosity of water. For Paramecium, rowing through pond water is like H. sapiens swimming through molasses (Purcell, 1977). Second, despite the invention of subcellular compartments by eukaryotes, there was a limit to how many different proteins could coexist without hindering function (Zhang et al., 2008). Third, they grew too large to communicate internally by chemical diffusion (which is fast over short distances but slow over long ones). Fourth, a unicellular organism could send electrical signals across its membrane, but this approach allowed only a single communication channel. These problems were all solved when cells assembled as a multicellular organism.
Multi-cellularity expanded resources and computational capacity
An animal assembled from many cells could enlarge by orders of magnitude and thus overcome the viscosity of water that had restricted foraging. Multi-cellularity also solved the limit on protein diversity because now cells could specialize their proteins for particular functions such as contraction (muscle cells), metabolic storage (liver cells), and so on. Muscle cells could specialize still further to, for example, contract slowly (gut), rapidly (limbs) or rhythmically (heart). Moreover, as more cells specialized for rapid signaling, an animal could greatly expand its circuits for processing information. This was essential to manage rising complexity.
By about 500 million years before the present, there arose from the jellyfish line a marine worm termed the urbilaterian because it was the progenitor of all subsequent bilaterally symmetrical animals, such as insects and vertebrates (Arendt et al., 2008). From this worm, thought to resemble its closest surviving descendent, Platynereis dumerilii, we inherited some key building blocks: bilateral symmetry, which was efficient for attaching legs; sensors in the head, which was efficient for detecting what is coming and where we are going; a brain in the head, which shortens the input wires; and a brain extended as a nerve cord, which places motor circuits near their effectors and shortens the output wires. We also received various additional principles of efficient neural design (Sterling and Laughlin, 2015).
One key design principle was to regulate in a predictive manner. Internal sensors collect detailed information regarding nutrient levels, osmolarity, sexual state and so on, while external sensors collect detailed information from the environment, such as time, temperature, pH, light, the danger of predators, and opportunities for food, shelter and sex. The brain processes all this information, prioritizes needs, establishes efficient trade-offs, and weighs opportunities against dangers. Then it predicts what should serve best and chooses a behavior, plus all the metabolism and physiology needed to support it (Figure 1). Predictive regulation, termed allostasis, minimizes the frequency and size of errors; thus it is intrinsically more efficient than homeostasis, which waits for errors to occur and then corrects them by negative feedback (Sterling, 2012).
Figure 1
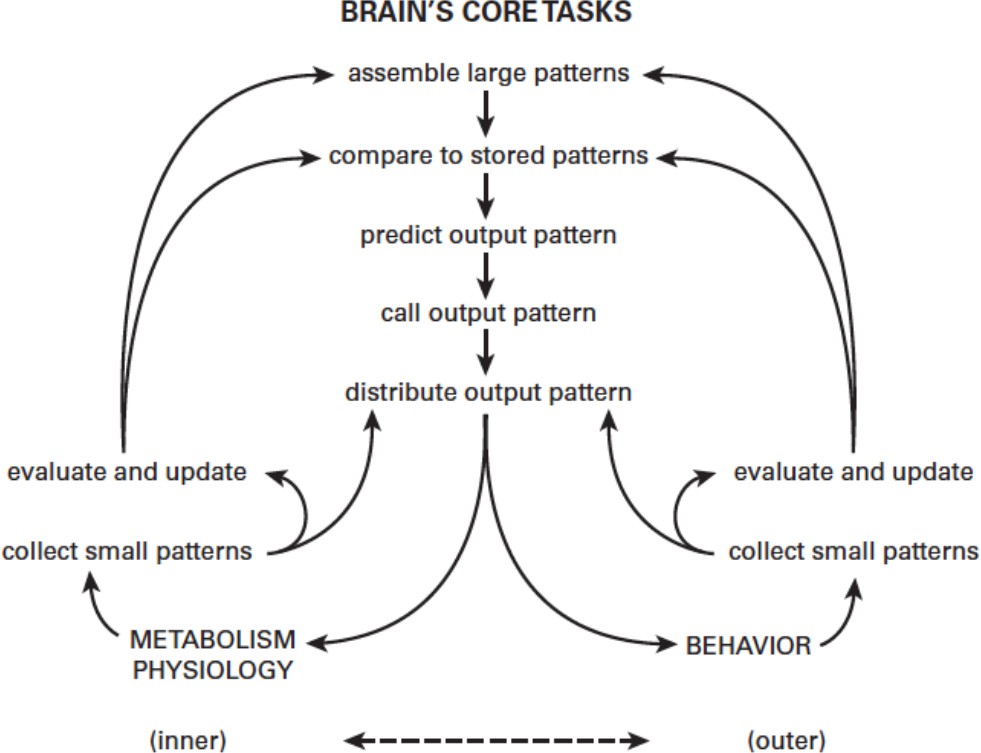
Predictive regulation (allostasis).
The brain’s fundamental challenge is to match the inner needs of metabolism and physiology (bottom left) with the outer needs of behavior. Small input patterns directly drive low-level output mechanisms to produce a rapid response (known as a 'reflex'). Small input patterns are also processed and combined to form larger input patterns that allow informed decisions to be made (for example, if gut is empty, send blood from gut to muscle; otherwise, send blood from kidney to muscle). The brain compares these larger input patterns to stored patterns for historical context (what happened last time?) before deciding on a course of action. The list on the top right shows innate needs served by predictive regulation in the earliest bilaterians, exemplified by Platynereis; H. sapiens has additional innate needs (bottom right). However, both species use the same 'choosing circuit' (which learns by reinforcement of positive reward-prediction errors).
© 2015 MIT Press. Figure 1 reprinted from Sterling and Laughlin, 2015 with permission.
The earliest key to predictive regulation was a circadian clock. Predicting when to forage, it sets metabolism and physiology to catabolic mode, and predicting when to grow and repair, it resets them to anabolic mode. The central clock governs separate clocks in each tissue so that they can predict locally when to turn on; for example, turn on liver before meals and muscle before exercise (Gerhart-Hines and Lazar, 2015). This hierarchy of clocks also exists in the fruit fly, so it too was probably present in our last common ancestor, the urbilaterian worm.
Another principle of efficient neural design was to send signals at the lowest possible rate (that is, the fewest bits per second) because it is cheaper and conserves resources. For example, hormones distribute information wirelessly at low rates without taking up space. Moreover, the final transfer of information by a hormone (via a G protein) takes about the same amount of energy as is provided by one molecule of ATP (Sterling and Laughlin, 2015). To allow signals to be sent at higher rates the early brain added electrical signals, but the energy needed to send a sodium current through a single membrane channel for one millisecond to create an electrical 'spike' or signal takes 2,000 times the energy needed for a hormone signal (Perge et al., 2012; Sterling and Laughlin, 2015). All this means that the early brain used electrical signals sparingly, and this principle has been preserved during evolution.
Another principle was to learn. By storing experiences an animal can predict what behaviors it should repeat. Thus, when a behavior delivers a result better than predicted, certain neurons reward the 'choosing circuits' by secreting a pulse of dopamine. This encourages the brain to store the memory and repeat the behavior (Glimcher, 2014; Schultz, 2015). This type of reward learning is mathematically optimal, and since it is present in the fruit fly, it was probably already present in the brain of our last common ancestor the urbilaterian worm. Reward learning emerged early as a core principle of efficient regulation, and H. sapiens took it to a whole new level partly because we experience the pulse of dopamine as a pulse of satisfaction – a brief uplift in mood.
To occupy the world H. sapiens required a large, efficient brain
Fast forward to primates. Our nearest living relative, the chimpanzee, forages for visible edibles but lacks the imagination to harvest what it cannot see. Human foragers learned that tubers concealed beneath the ground store carbohydrates that are highly nutritious if properly processed and cooked. Moreover, humans remember where the tubers grow and when. The chimpanzee hunts cooperatively for small animals, but the dominant male immediately appropriates the prize. Human hunters learned that effective cooperation requires fairness, so they manage more restraint and complex schemes needed to capture larger game. The chimpanzee ranges over about 6 km2 of forest, but a human hunter learns to hunt effectively and safely over 12,000 km2 (Kaplan and Robson, 2002).
Human foragers collect and process roughly five-fold more calories than a chimpanzee – and do so anywhere on earth. The requirement is a three-fold larger brain that matures gradually. Whereas a chimpanzee has learned all it needs to know for caloric production by age five, a human forager’s passage from net consumer to net producer requires 20 years. Moreover, its learning curves for gathering and hunting continue rising until around age 45. By then a hunter will have quadrupled their productivity compared to their entry level. Such prolonged learning indicates that gathering and hunting are challenging careers that require life-long study and practice (Kaplan and Robson, 2002; Gurven et al., 2017).
The expanded brain continued to follow a number of principles in the interests of efficiency:
to specialize, because two parts are more efficient than one part doing two jobs: for example, visual cortex carves out separate circuits for color, motion, faces, and objects.
to only express circuits that are needed, because this saves space and energy: for example, the visual cortex expands circuits to analyze a tiny, high-resolution patch of the central retina (0.1%) for identifying faces, and it reduces circuits for analyzing the extensive low-resolution regions of peripheral retina. This design shrinks space and energy costs by1000-fold (Akbas and Eckstein, 2017).
to separate the neural circuits, because this reduces wire length: for example, visual cortex separates circuits for the two eyes because interlacing them would require each to route its wires around the other (Chklovskii and Koulakov, 2004).
Ultimately cerebral cortex of H. sapiens subdivided into about 180 different areas (Glasser et al., 2016). These include large areas for early sensory processing and motor control, but also various small, narrowly specialized areas, such as six patches for recognizing faces (Tsao et al., 2008). Every square millimeter of cortex is occupied with some specific computation. Moreover, the areas retain considerable plasticity. For example, as we learn to read, even in adulthood, a particular area in the left hemisphere shifts from recognizing objects to recognizing written words (Nakamura et al., 2012).
Once an individual brain reaches a set volume and cortical expanse, its computational capacity can still increase. First, it specializes the corresponding areas in each hemisphere. For example, auditory areas on the left emphasize language, and on the right they emphasize music (Garcea et al., 2017). Second, it programs cortical areas to mature and regress at different stages of the life cycle, thereby matching expensive cerebral resources to the needs and capacities of each stage. For example, visual and motor tracts mature by our mid-30s and slowly regress, but prefrontal and temporal tracts mature around age 45 and regress much later (Yeatman et al., 2014).
Why would H. sapiens postpone its final brain development for so long? These late-developing tracts in prefrontal cortex – which support insight, planning, impulse control and choice – ripen in foragers as their economic productivity is peaking. This timing means that a family benefits from a family member during the last few decades of his or her 'three score and ten'. It surprises us to realize that early humans managed to survive so long without modern medicine. So here’s the math: each child, on its way to caloric self-sufficiency at age 20, accumulates an enormous energy debt to the family. Partial repayment occurs during the 20s and 30s via calories transferred to the children; but that does not suffice to retire the debt, so repayment continues from ages 40 to 70 via transfers to grandchildren (Kaplan and Robson, 2002).
Grandparental foraging allows parents to bear their next child sooner without endangering the existing offspring. If most foragers were to die before repaying their caloric debt, the reproductive capacity of H. sapiens would plummet and threaten its extinction. In summary, to occupy the world required a large brain which, in turn, required a long life-span to allow substantial energy transfers from grandparents. Chimpanzees, by the way, repay their smaller debt sooner and are dead at 45 (Blurton Jones et al., 2002; Kaplan and Robson, 2002; Kim et al., 2012; Gurven et al., 2017).
Expanding the community’s computational capacity
Once the raw computational capacity of an individual brain reached its limits of space and energy, our species could still expand the computational capacity of the group. Just as the brain specialized the hemispheres for different tasks, it also specialized people, providing each with a different set of circuits and thus a different bundle of innate talents. The learning system encourages the exercise of talents, because that is most rewarding, and soon this generates a community of experts – hunter, healer and so on. A community of experts can easily outcompete a community where everyone is the same, but it can also create profound psychological stresses and interpersonal conflicts.
Therefore the 'community-of-experts' design also includes innate capacities to release tensions and preserve social cohesion. Such behaviors might collectively be termed sacred practices, where 'sacred' means 'reverence for what cannot be expressed in words'. These practices include sex, music, dance and a multitude of ceremonies surrounding birth, puberty, marriage and death. They also included stories, jokes and, eventually, literature. Sacred practices elicit intense emotions such as awe, joy and grief – which somehow bring relief. Accepting the principle 'to use expensive circuits only as needed', the investment of H. sapiens in circuits for producing and processing music and art suggest their importance to our design.
Sacred practices emerged early in the evolution of H. sapiens. A skilled artist stenciled hands on a cave wall near the southern tip of South America nine millennia ago; a musician played a bone flute in northern Europe 40 millennia ago; sculptors carved abstract objects and ornaments in southern Africa between 70 and 100 millennia ago. Paintings of similar antiquity in a Spanish cave are attributed to H. neanderthal, suggesting its capacity for sacred practice (Hoffmann et al., 2018). Thus, our design – extreme individuality coupled to extreme sociality and relieved by sacred practice – was apparently inherited from our last shared ancestor with H. neanderthal.
All these activities could be termed 'culture'. But here I consider them more narrowly – as neurally-generated, species-specific behaviors. The neural circuits that generate and respond to language, music and art required numerous cortical areas to expand at great metabolic cost. That required a richer diet and one that was rendered more digestible by cooking. As these requirements were met, the intestine was reduced. This allowed the brain, which resembles the intestine in metabolic cost, to expand without an overall increase in metabolic rate (Aiello and Wheeler, 1995). How language circuits would have paid their way seems obvious – but painting and music? How did the investments in our neural endowment for the arts pay off nutritionally? The answer is that it probably facilitated long-term cooperation between unrelated specialists (Muthukrishna and Henrich, 2016).
What led to our present difficulties?
For roughly two million years members of the genus Homo used fire to warm a cave and roast a root. But in 1769 James Watt patented the first efficient machine that used fire to perform mechanical work. This invention coincided with the final stages of 'enclosure' that moved English peasants from their traditional common lands into cities (Overton, 1996). The new urban labor pool was immediately set to work at new machines.
Of course, Watt’s engineers already had 'slide rules' to calculate his machine’s critical specifications; and agricultural productivity had already increased to feed the urban workers. Water-driven mills had already created centers for production, and so on. Yet, the invention that first oxidized carbon to generate power instantly transformed our capacity to exploit resources across the whole planet. Soon the burning of coal enabled steam shovels to dig canals, steam locomotives to cross deserts, steam ships to cross oceans, and steam factories to maintain production, even when rivers froze or went dry. Watt’s engine was the most significant development in the generation of power since the mitochondrion.
The steam engine also led to the rise in atmospheric carbon dioxide that drives global warming today (Figure 2). Our species’ ability to change the climate followed inexorably from the preeminent computational capacity of certain individual brains – like Watt’s – and their preeminent capacity to cooperate. But what now drives our consumption of manufactured goods that factories keep pumping out? And what drives our consumption of rich foods and intoxicants? Since biology matched resources to predicted need for billions of years why, in a time of abundance, does human 'need' continue to grow apparently without bound?
Figure 2
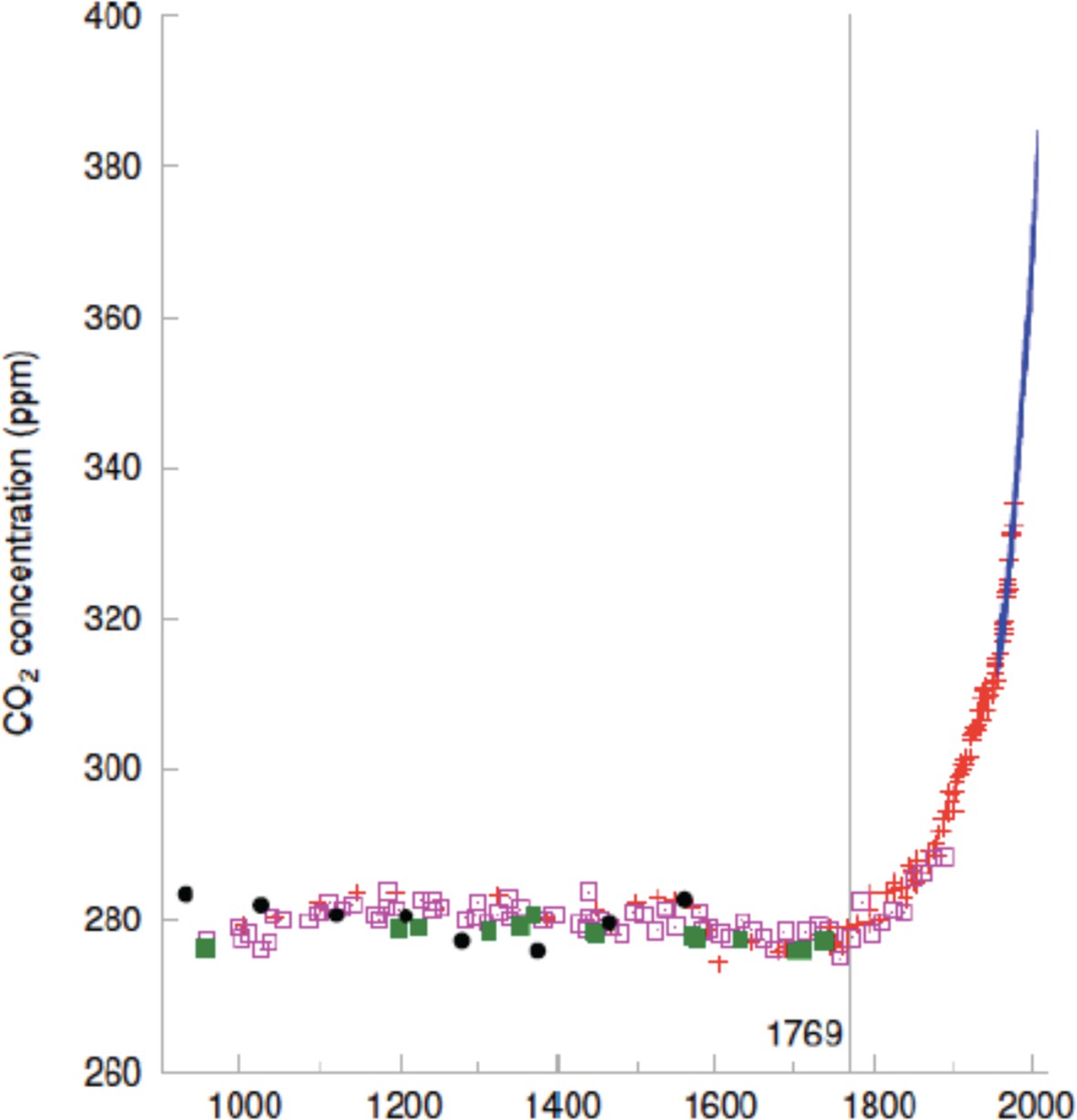
The steam engine initiated a sharp rise in atmospheric carbon dioxide.
Concentration of carbon dioxide in the atmosphere (y-axis) as a function of year. The steam engine was patented in 1769.
© 2008 Mackay. Figure 2 reprinted from Mackay, 2008 under a CC BY-NC-SA 2.0 UK license.
The steam engine removed men, women, and children from challenging, integrated lives in the countryside to jobs that yoked them to a machine or sent them down a mine for twelve hours a day or more (Engels, 1845). Adam Smith had anticipated the consequence, writing in the very year that his friend’s machine went on line: "The man whose life is spent in performing a few simple operations has no occasion to exert his understanding or to exercise his invention in finding out expedients for difficulties which never occur. He naturally loses, therefore, the habit of such exertion and generally becomes as stupid and ignorant as it is possible for a human creature to become" (Smith, 1776).
Many factory and agricultural tasks still fit Adam Smith’s description. As do many 'post-industrial' tasks that confine descendants of large-brained foragers within temperature-controlled offices to sit still and stare at screens. Of course many professions – science among them – remain challenging. Yet, it should concern us that simple tasks learned quickly disallow the continuing rewards from prolonged mastery; that time pressure and isolation disallow the rewards from companionate work and community approval; that reduced physical exertion attenuates the rewards from rest; that vicarious sports, music art, and sex lose the rewards from directly exercising wit and skill. Overall, it seems that in 'controlling nature', we shrink the rewards that accompany relief from its small discomforts (Sterling, 2016).
What happens when the abundance, comfort and social isolation of modern life shrink the sources of positive surprise required by our ancient circuit to deliver its reinforcing pulse of dopamine? According to computational theories of reward learning, the system is compelled to seek reinforcement from their intensification (Sutton and Barto, 1998). Thus, for many people the main source of positive surprise becomes 'more'. In predicting a Mac, the next surprise must be a Big Mac, and then a Whopper. Notice that when the surprise is more, the next surprise must be even more.
Predictive regulation in a regime of shrinking positive surprise
Unbounded societal consumption of manufactured goods has been explained by theories that an economy must 'grow or die'. But for a system driven by the need to intensify, 'grow and die' is more likely – because unbounded consumption drives diverse, mutually reinforcing pathologies. At the planetary scale, rising carbon dioxide increases atmospheric temperature that melts sea ice. That increases absorption of solar radiation – a cause of further warming that melts permafrost, releasing sequestered greenhouse gases, and so on. The consequent environmental pathologies are innumerable and familiar (Kolbert, 2015).
Similarly, unbounded individual consumption of rich food drives metabolic circuits awry. Normally the brain predicts the glucose level needed to fuel, say, a game of tennis and sets the level accordingly by modulating the secretion of insulin and many other hormones. But, when the reward system drives consumption of carbohydrate and fat far beyond metabolic need, the regulatory circuits chronically predict a need to elevate insulin (Kleinridders et al., 2014). Insulin receptors in many tissues, including the brain, adapt by reducing their sensitivity to insulin ('insulin resistance'), and this eventually causes cells to need more insulin, which evokes further resistance. The process leads finally to type 2 diabetes, whose complex pattern of endocrine signaling contributes to hypertension and vascular inflammation – thus elevating cardiovascular mortality (Figure 3; American Heart Association, 2017).
Figure 3
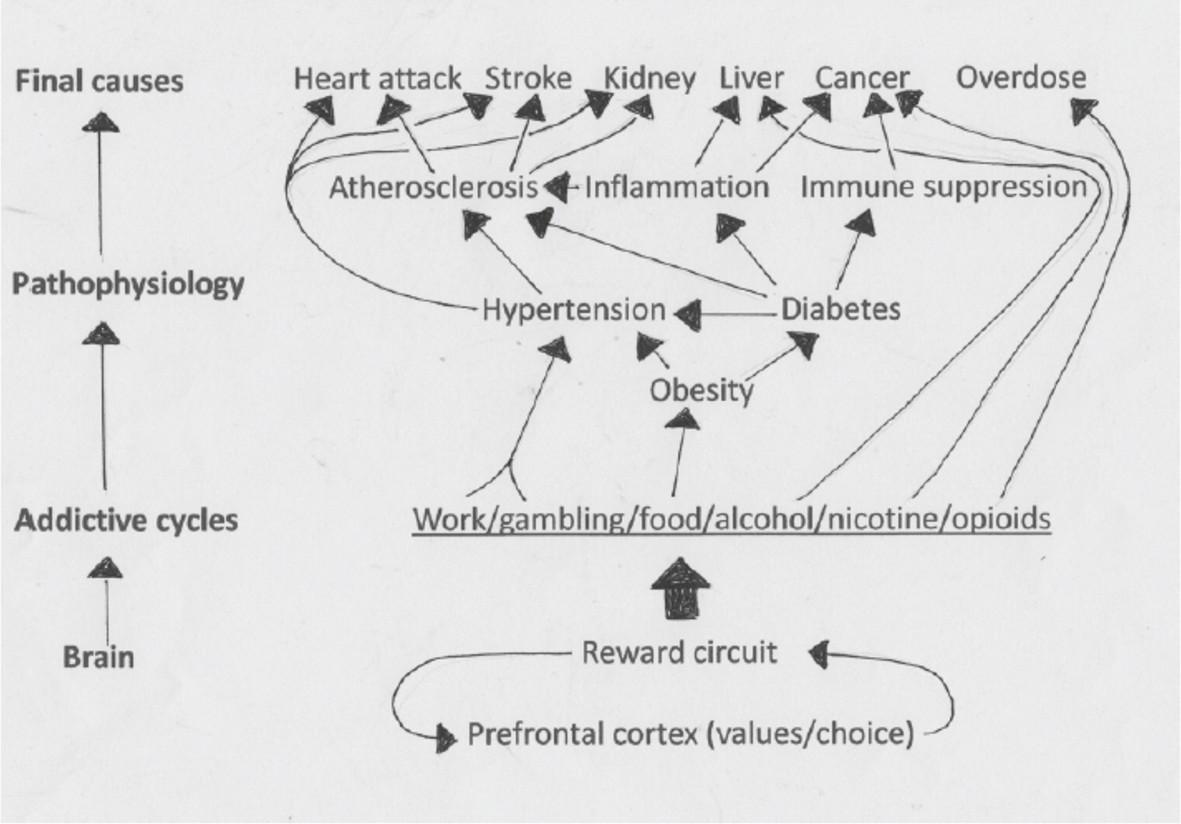
The unbounded consumption of rich food drives metabolic circuits awry.
The brain drives consumptive behaviors that mobilize many different hormones (not shown) from the brain, gut, liver, pancreas, bone, fat, muscle and other tissue. These regulatory hormones eventually require further elevation: for example, sustained high levels of insulin eventually leads to a need for even higher levels of insulin. The end result can be obesity, diabetes, hypertension and a range of other medical conditions.
Unbounded consumption of drugs (such as nicotine, alcohol, cocaine, amphetamine and opiates) easily drive the reward circuit into an addictive cycle because they directly release dopamine or prolong its presence at synapses (Keiflin and Janak, 2015). Various loops in the reward circuit adapt to these drugs and demand higher doses. Investigators of addiction now recognize that compulsive consumption of rich foods and dopamine-elevating drugs share the same circuits (Murray et al., 2014). And suspicion grows that the same is true of other compulsive behaviors (such as addiction to gambling, porn and shopping). Individuals who are able to exercise their diverse skills, especially those skills for which they have an innate talent, are more likely to obtain sufficient rewards naturally and thus avoid becoming trapped in addictive behaviors.
One doubt raised against the present hypothesis is that, after all, the 'market' has produced wealth undreamed of by earlier generations. It is true that the 'masses' now have access to immense varieties of processed food, processed entertainment, and opportunities for programmed travel. However, these activities are largely passive. Moreover, since they require neither exercise, creativity or courage, they cannot deliver surprises equivalent to the same activities carried out actively. Each one of the ten thousand items in a modern supermarket might potentially deliver a positive reward signal, but these items are all basically predictable – you know exactly what you will find in a particular aisle in a supermarket. Moreover, the ten different brands of, say, olive oil are nearly indistinguishable. Thus the supermarket stimulates boundless consumption precisely because it offers few positive surprises.
Alternative hypothesis to explain the planetary rise in obesity
Some observers suggest that, when food was more difficult to obtain, we were mostly hungry, so the best survival strategy would have been to gorge whenever possible and store fat via metabolic pathways programmed by 'thrifty genes'. Yet, contemporary foragers (hunter-gatherers) are not mostly hungry. Rather they employ mathematically optimal foraging strategies to satisfy their nutritional needs with relatively few hours per week, leaving considerable free time for sacred practices. Put another way, foragers reliably obtain sufficient food to support their cultural activities (Kelly, 2014).
Feeding is regulated meticulously by myriad signals from gut, liver, muscle, fat, bone and brain (Garfield et al., 2015). Neural circuits include various 'push-pull' mechanisms that slacken the drive to eat as satiety approaches and restore the drive to forage even as a meal is winding down. Consequently animals in the wild, including H. sapiens, tend neither to overeat nor over-forage. They just find other activities. Foraging, remember, carries risk of being foraged. Where animals do get fat is in a zoo, and that is where we seem to have placed ourselves.
The 'thrifty gene' hypothesis has largely collapsed. Now, except for regions of periodic famine and warfare, essentially all human populations across the planet – whatever their genotype – are rapidly depositing fat (GBD 2015 Obesity Collaborators et al., 2017). Moreover, fruit fly larvae fed a high-carbohydrate diet also develop hyperglycemia, insulin resistance, type 2 diabetes and high plasma levels of triglycerides and free fatty acids (Musselman et al., 2011). Thus human mechanisms of metabolic regulation were apparently inherited from our shared urbilaterian ancestor, suggesting that all animals are vulnerable to obesity.
When reward diversity is restricted by socio-economic inequalities, which include poor education and unrewarding 'jobs', eating remains. This may explain why obesity is highest in countries with greatest inequality of income, with the United States literally taking the cake (Wilkinson and Pickett, 2010). And within the United States, obesity is highest in the most unequal states, and highest among the least educated (Ogden, 2010).
What level to treat?
The standard medical model views much pathology as biochemical 'dysregulation' – too much of one molecule or too little of another. Consequently therapies focus on drugs to treat biochemical circuits at the lowest levels. For example, the resemblance of binge eating to opioid addiction suggests an antagonist of the µ opioid receptor as a therapeutic agent (Ziauddeen et al., 2013). More broadly, newly discovered molecular regulators of carbohydrate and lipid metabolism are often mentioned as possible therapeutic targets for diabetes, heart disease and the panoply of interconnected pathologies shown in Figure 3.
Where a molecule is demonstrably missing, such as insulin in type 1 diabetes, the approach has a clear logic and has achieved great triumphs. But where the approach targets a complex circuit that is incompletely known and not obviously broken, the logic is less compelling (Sterling, 2014).
Consider, for example, essential hypertension, a major killer that has been treated pharmacologically for half a century. Children entering school are separated from their families and confined in large groups where they must remain still, silent and attentive to the voice of authority. Children gifted with those abilities are well rewarded with praise and gold stars. But for children gifted with a compulsion to move and a strong inner voice, school can be stressful, and soon blood pressures start to rise. By age eighteen nearly 50% of US boys have systolic pressures of 130 mm Hg or higher (Falkner, 2005). The brain, regularly predicting a need for higher pressure, drives all components of the circuit (Figure 4). Gradually each component learns its role in sustaining elevated pressure (like skeletal muscles learning to anticipate exercise). This circuit is not 'broken', nor are the components behaving 'inappropriately'; they are simply responding to the brain’s predictions. Eventually, however, the circuit enters a pathological cycle: as arteries adapt to higher pressure by thickening and stiffening, they need more pressure to achieve the required output – but higher pressure further narrows and stiffens the arteries. Finally the system loses the ability to resume normal pressure as it should, for example, during sleep. Chronically high pressure causes inflammation, which encourages atherosclerosis and eventually heart attack or stroke.
Figure 4
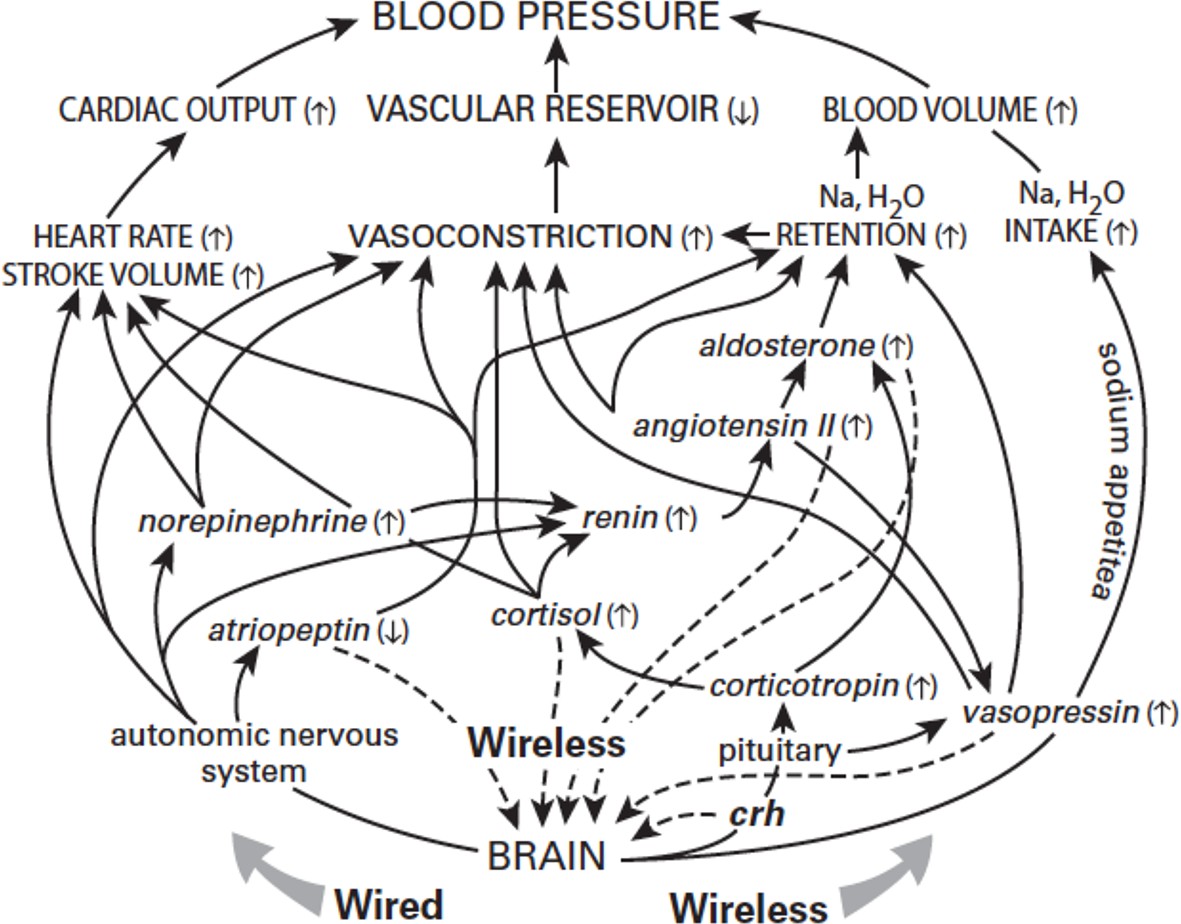
The origins of high blood pressure.
The brain predicts what blood pressure will be needed and then relies on multiple mechanisms to adjust blood pressure accordingly. These mechanisms, operating on different time scales, are shaped by an interconnected network that employs wired signals (sent along neurons), wireless signals (transmitted by hormones), and motivated behaviors, such as an increased appetite for salt. Again, unbounded individual consumption can lead to high-blood pressure.
© 2015 MIT Press. Figure 4 reprinted from Sterling and Laughlin, 2015 with permission.
In treating hypertension medicine commonly targets the lowest level mechanisms (Figure 4). Since one cause of high blood pressure is excessive fluid for the vascular reservoir, hypertension is often treated with a diuretic to shrink blood volume. But then the brain, predicting a need for high pressure, compensates by shrinking the reservoir. To prevent that, a calcium antagonist is added to relax vascular smooth muscle. Still the brain insists that pressure should be high, and again it compensates by increasing cardiac output. To prevent that, a beta blocker is added, thus antagonizing the last pathway capable of raising the pressure. Unfortunately, this also renders the patient unable to exercise – a core need for every aspect of physical, emotional, and cognitive health.
Similar scenarios can be drawn for other epidemic problems, such as obesity, type 2 diabetes and drug addictions. All of our control systems are designed with multiple compensatory loops because that allows for efficient trade-offs. This leads me to doubt that any 'magic pill' will heal the global epidemics whose simple cause is unbounded consumption and whose deeper cause is loss of reward diversity. A more promising strategy would be to re-expand the opportunities for small satisfactions and thus rescue the reward system from its pathological regime. Individuals satisfied by their work and active extracurricular activities are less likely to rely on food and other substances to quell their restless searching.
To reach this point would require a substantial reorientation of attitudes and politics. However, since the result would reduce consumption of everything from fossil fuels to medical care for chronic disease, economic resources should present no obstacle.
Conclusions
Just as single cells are designed to operate just above thermal noise, single humans are designed to operate just above the roiling currents of their myriad needs. This means that we are constantly reaching out and struggling to stay above the waves. However, to me at least, the core of the design of H. sapiens – extreme individuality coupled to extreme sociality – suggests some principles for healing.
First, recognize individual clustering of abilities and deficits as fundamental to our design. A deficit is not prima facie a 'disorder'; it may simply be a gap to be filled by a neighbor, whose own gap may in turn be filled by someone else. We need to develop constructive niches for individuals with different clusters. To start, we should cease confining all children into the same classroom and treating those who tolerate it poorly with amphetamines. Instead, we should identify the special gifts conferred on each child and support their early practice. The resources needed to accomplish this are now being squandered, as later down the line they are spent to incarcerate aimless, angry, drug-addicted young adults.
Second, recognize that pharmacology cannot be the primary route to health. The brain has been regulating the body since Platynereis, and the systems are far too complicated to be managed primarily with pills.
Third, re-diversify activities that can offer unpredicted rewards. Much public discourse concerns the need for more jobs. But the inexorable trend toward automation will be reducing the number and quality of jobs. Masses of humans will need something interesting to do besides consuming stuff and traveling from place to place snapping 'selfies'. We will need somehow to reverse the trend that Adam Smith identified in 1776 toward losing our skills and alertness from oversimplifying work.
Fourth, renew and re-diversify the sacred practices upon which H. sapiens depends as a species to relieve the tensions caused by our innate strangeness. This included music, art, dance, literature and monumental constructions that engaged large numbers of people over decades. Of course, we are already rich in the products of sacred practice – we can enjoy a concert, play, museum, or novel – for an hour or so. But it is the artists who practice their sacred skills over years and decades who benefit daily from the small, unpredicted rewards of improving a skill. Vicarious activity cannot substitute for individual engagement.
References
-
Object detection through search with a foveated visual systemPLoS Computational Biology 13:e1005743.https://doi.org/10.1371/journal.pcbi.1005743
-
Heart disease and stroke Statistics–2017 UpdateCirculation 135:e146–e603.https://doi.org/10.1161/CIR.0000000000000485
-
The evolution of nervous system centralizationPhilosophical Transactions of the Royal Society B: Biological Sciences 363:1523–1528.https://doi.org/10.1098/rstb.2007.2242
-
Antiquity of postreproductive life: are there modern impacts on hunter-gatherer postreproductive life spans?American Journal of Human Biology 14:184–205.https://doi.org/10.1002/ajhb.10038
-
Maps in the brain: what can we learn from them?Annual Review of Neuroscience 27:369–392.https://doi.org/10.1146/annurev.neuro.27.070203.144226
-
A neural basis for melanocortin-4 receptor-regulated appetiteNature Neuroscience 18:863–871.https://doi.org/10.1038/nn.4011
-
Health effects of overweight and obesity in 195 countries over 25 yearsNew England Journal of Medicine 377:13–27.https://doi.org/10.1056/NEJMoa1614362
-
Circadian metabolism in the light of evolutionEndocrine Reviews 36:289–304.https://doi.org/10.1210/er.2015-1007
-
Understanding the hows and whys of Decision-Making: from expected utility to divisive normalizationCold Spring Harbor Symposia on Quantitative Biology 79:169–176.https://doi.org/10.1101/sqb.2014.79.024778
-
The Tsimane health and life history project: integrating anthropology and biomedicineEvolutionary Anthropology: Issues, News, and Reviews 26:54–73.https://doi.org/10.1002/evan.21515
-
BookThe Lifeways of Hunter-Gatherers: The Foraging SpectrumNew York: Cambridge University Press.
-
Increased longevity evolves from grandmotheringProceedings of the Royal Society B: Biological Sciences 279:4880–4884.https://doi.org/10.1098/rspb.2012.1751
-
A rotary molecular motor that can work at near 100% efficiencyPhilosophical Transactions of the Royal Society B: Biological Sciences 355:473–489.https://doi.org/10.1098/rstb.2000.0589
-
Bioenergetic constraints on the evolution of complex lifeCold Spring Harbor Perspectives in Biology 6:a015982.https://doi.org/10.1101/cshperspect.a015982
-
Hormonal and neural mechanisms of food reward, eating behaviour and obesityNature Reviews Endocrinology 10:540–552.https://doi.org/10.1038/nrendo.2014.91
-
A high-sugar diet produces obesity and insulin resistance in wild-type DrosophilaDisease Models & Mechanisms 4:842–849.https://doi.org/10.1242/dmm.007948
-
Innovation in the collective brainPhilosophical Transactions of the Royal Society B: Biological Sciences 371:20150192.https://doi.org/10.1098/rstb.2015.0192
-
BookAgricultural Revolution in England: The Transformation of the Agrarian Economy 1500–1850Cambridge: Cambridge University Press.https://doi.org/10.1017/CBO9780511607967
-
Why do axons differ in caliber?Journal of Neuroscience 32:626–638.https://doi.org/10.1523/JNEUROSCI.4254-11.2012
-
Neuronal reward and decision signals: from theories to dataPhysiological Reviews 95:853–951.https://doi.org/10.1152/physrev.00023.2014
-
BookPrinciples of Neural DesignCambridge, MA: The MIT Press.https://doi.org/10.7551/mitpress/9780262028707.001.0001
-
Allostasis: a model of predictive regulationPhysiology & Behavior 106:5–15.https://doi.org/10.1016/j.physbeh.2011.06.004
-
BookThe Spirit Level: Why Greater Equality Makes Societies StrongerNew York: Bloomsbury Press.
-
Lifespan maturation and degeneration of human brain white matterNature Communications 5:4932.https://doi.org/10.1038/ncomms5932
Decision letter
-
Peter RodgersReviewing Editor; eLife, United Kingdom
In the interests of transparency, eLife includes the editorial decision letter and accompanying author responses. A lightly edited version of the letter sent to the authors after peer review is shown, indicating the most substantive concerns; minor comments are not usually included.
Thank you for submitting your manuscript "Neurobiology of Homo sapiens: what went wrong?" to eLife for consideration as a Feature Article. Your manuscript has been reviewed by three peer reviewers, and the evaluation has been overseen Peter Rodgers as the eLife Features Editor.
The full reports from the reviewers are below. The following individuals involved in review of your manuscript have agreed to reveal their identity: Jeremy Gunawardena (Reviewer #2) and Bruce McEwen (Reviewer #3).
While the reviewers found the manuscript to be interesting and provocative, they also found it to be "speculative and superficial" in places. It is vital, therefore, that you fully address all the concerns of the reviewers in a revised version of the manuscript. In particular, you need to address the following points:
– Referee 1. Point 1 (which argues against the conclusion that "modern life shrinks reward diversity.")
– Referee 1. Point 2 (which argues against the conclusion that something has "gone wrong"/that we face a "crisis"). Referee 2 makes a similar point in paragraph 7 of his report.
– Referee 2. The comments in paragraph 4 (about extreme teleological language).
– Referee 2. The comments in paragraph 8 (about language and culture).
– Referee 2. The comments in paragraph 9.
– Referee 3. The comments in paragraph 2 (about the role of the human prefrontal cortex)
Reviewer #1:
This article discusses the origin of various "crises" impacting the human race (climate change, obesity) and argues that they all originate from the drive for positive reward prediction error, which in a society of abundance results in excessive consumption. The article is interesting and provocative in a popular science way, and presents many intriguing connections between evolution, neuroscience, history, and behavior that will stimulate thought among readers. But it is also extremely speculative and superficial in its treatment of these topics.
Major points
1) This is a strange article in that it is extremely broad, speculating about everything from the evolution of life on earth to the modern opioid epidemic. I am not sure that something like this belongs in a scientific journal that publishes original research such as eLife, or at least I am not sure by what criteria it should be reviewed. The article skips through so many different areas of biology at such a superficial level that it is unclear where to begin with criticism. For example, a central conclusion is that "modern life shrinks reward diversity." Is this true? I could make a counterargument that life as a hunter-gatherer was likely repetitive, and that my modern existence as a scientist living in San Francisco provides a much greater diversity of interesting and unexpected stimuli. But in any case the author provides no evidence for his position beyond examples and anecdotes.
2) The article starts from the premise that something has "gone wrong" with the human race and that we face a "crisis". But by virtually every measure the human race is better off now than it has been at any point in history – longer lifespans, better health, greater freedom and security, etc. The "crisis" that the author describes consists of a few cherry-picked examples that support his thesis. It is reasonable to suggest that obesity or drug addiction may be caused by dysregulated reward systems, but the author instead wants to propose an overarching theory for life on earth.
Reviewer #2:
1) This is a fascinating and ambitious paper, which contains many interesting ideas that should be of broad interest. However, it also contains suggestions that are poorly worked out, which get in the way of appreciating the former.
2) The author attempts something that is increasingly rare, which is to integrate our scientific understanding across the scales from the molecular to the physiological to the social. I can think of few other scientists with the chutzpah to attempt this, with the exception perhaps of Jean-Pierre Changeux ("Climbing brain levels of organisation from genes to consciousness", TICS 21:168-181 2017). This kind of integration is sorely needed, because the rest of us are too scared to attempt it, and I vigorously applaud the author for trying.
3) The most interesting and most important ideas centre around "allostasis", or the re-evaluation of the classical homeostasis of Claude Bernard and Walter Cannon to incorporate the central nervous system, prediction, motivation and feed-forward regulation (Figures 3 and 11). This draws on the author's own seminal contributions (Sterling, 2012), as well as the powerful synthesis of neural "design principles" in the author's collaboration with Simon Laughlin (Principles of Neural Design, MIT Press 2015). These ideas suggest that the organism is not just responding to its environment, as in classical homeostasis, but actively, if largely unconsciously, exploring and internalising it. The organism acquires thereby an unconscious autonomy over aspects of its functioning that has previously been overlooked (Figures 10 and 12). This is an extremely provocative viewpoint and I quite understand why the author feels it ought to have profound implications for how we think about the human organism in the context of upbringing, education, illness and participation in society.
4) Allostasis is described within an account of how increasing information-processing complexity has emerged from the first primordial cells to us (first 3 sections after the Introduction). This would be engaging and entertaining were it not for its extreme teleological language. For example, "Cells began to accumulate odd sequences that could serve some future challenge", "Moreover, an animal could specialize many cells for rapid signaling and thus greatly expand its channels for processing information". What?! Evolution could look ahead, could it, to see what it would need in the future? It will be anticipating the emergence of humans next. Who needs intelligent design when a biologist writes like this? Of course, the author knows better, which only makes it worse. Evolutionary biologists must despair of the rest of us.
5) The clinical context is where allostasis should have the most traction, by providing an alternative to traditional pharmacology's "magic bullet", as trenchantly described in "Treating the lowest levels". Working out a strategy for reaching the "higher levels" and convincing clinicians of its value would be an immense contribution.
6) However, the paper does not take this route. It seeks instead to stretch much further and here the ice starts to crack, if not disappear altogether. According to the author, the human species "went wrong" by discovering capitalism (Figure 8), which "shrank reward diversity" (Figure 9), so that our allostatic biology led inexorably to global warming and a pandemic of ignorance, stupidity, Big Macs, obesity, diabetes, hypertension and addiction. Furthermore, we are all so different that mental disease should be placed on a continuum, rather than treated categorically ("Understanding human differences"). I do not find these assertions compelling for many reasons.
7) First, each of them is worthy of a paper, if not a book, in its own right, which, to be credible, would have to take on board a formidable body of existing literature. In the case of what "went wrong", if, in fact, anything did, the discussion would have to go back at least as far as Rousseau's "noble savage". As they stand, these assertions come across as the author's personal view of the World.
8) Second, there is a long, and mostly unhappy, history of biologists trying to contribute to the human sciences. The author is attempting to do this at a very ambitious level but there is a striking failure to mention the key concepts which differentiate us from other animals and which form the intellectual currency of the humanities – language and culture. Allostatic biology is shared with many other animals. Language and culture are not. The former appears once in the paper, as an example of brain specialisation, and the latter appears not at all. How can they not be part of the explanation? Sociobiology at least attempted to say something about culture, with the "gene" as the unit of explanation, however unwelcome that was to most social scientists, but the author ignores culture, as if to say that allostatic "design", in place of "gene", is now sufficient to explain human behaviours and societies. There is an enormous gaping chasm here, to put it mildly.
9) Finally, these assertions lead to suggestions of what we should do to "resolve the tension between our biology and how we now contrive to live", which are, as listed in the "Conclusions", to "develop constructive niches for individuals", "reverse the trend toward becoming ever more stupid and ignorant", "identify each child's special gifts", "renew sacred practices" and be accommodating of sociopaths. I am afraid these strike me as platitudes.
10) In summary, there is an important and interesting paper here but it needs sharper focus and ruthless editing to extricate it from the author's personal beliefs. The updated view of the organism that the author is putting forward is worthy of serious scientific attention. If it becomes broadly influential, as it should, then perhaps in time it may help lead to the changes in society that the author wants to see. If so, it will be others who accomplish such change, not the author. He does himself, and us, a disservice by seeking a final cause, rather than allowing evolution to take its course.
Reviewer #3:
1) Thoughtful, informative of the science, and enjoyable to read. Clearly the author is very passionate about this topic and rightly so!
2) One possible addition, to what is implied but not stated, is to add a discussion of the variable role of the human prefrontal cortex in self-regulation of impulses, mood and other behaviors as well as proactive planning. The PFC develops after birth in formative years of childhood, adolescence and into young adulthood, and it develops more quickly on the average in females. (Hence, why for young males, auto insurance rates are higher than for females!) The variability of results of the famous "marshmallow test" on children indicates how variable among individuals is "self-regulation"; tests on children turn out to be predictive of self-regulations 40 years later! Early life adversity redirects the PFC development and is likely to be a contributor to the variability in impulsiveness, quarrelsomeness, proactive planning (e.g. for the future of our planet) as well as the pursuit of consumption and hedonic rewards.
3) Clearly education is an important factor. Our prolonged developmental period as humans allows the fortunate to develop the perspective needed to do what the author advocates. So many people are not so lucky!
[Editors' note: further revisions were requested prior to acceptance, as described below.]
Thank you for submitting the revised version of your article "Predictive regulation and human design" to eLife. I have received comments on the revised version of article from two of the reviewers who reviewed the original version, and discussed the revised version and these comments with another editor.
We agreed that you need to address some of the points made by Jeremy Gunawardena, and that we will accept the article for publication if you address these points plus a number of editorial points.
Re the comments from Jeremy Gunawardena: Please address the following points:
– point 3
– points 4 and 5 (which are related)
– points 6 and 7 (which are related)
– point 8. (Addressing some of the editorial comments should help address this point)
1) The author has made substantial changes to the original draft but not really in the way I was hoping for. Some of the interesting scientific details have been lost (previous Figures 5, 6, 10, for instance) but the personal opinions largely remain. However, the changes have made the logic of the argument clearer and I will focus on that.
2) To summarise, the author's claim is that the dopamine-based, reinforcement learning system which we share with other animals entered a regime of "positive feedback" in which "consumption grows explosively" because "modern life shrinks reward diversity", and this led to our "present difficulties" of "climate change, obesity, drug addiction".
3) Like reviewer 1, I am not convinced that modern life has lower reward diversity than that of an agricultural labourer in 1700 CE or a stone carrier undertaking the "monumental construction" of the pyramid of Cheops in ~2500 BCE. I expect it is quite the reverse. The author does not tell us how reward diversity can be measured and the issue tested, so this remains a matter of opinion. That is part of the problem, especially when personal opinion is not clearly stated as such (see point 8).
4) Even if we give the author the benefit of the doubt over point 3, I do not understand the claim that reinforcement learning systems lead to positive feedback and explosive growth. As the author points out, these learning algorithms have properties of optimality within a specified mathematical context (Niv, J Math Psych, 53:139-54 2009). Positive feedback and explosive growth are pathologies, which would destroy optimality. So, how do they arise?
5) The author suggests that they do so through habituation: "all neurons and circuits adapt. As a result repeats and becomes predictable, its targets at the molecular and circuit levels reduce their sensitivity". That is well and good but habituation leads to levelling off, not positive feedback, and levelling off is what a reinforcement learning system would do if it kept receiving the same reward. No instability arises. The assertion that reinforcement learning systems become pathological is not justified.
6) It is an enormous leap to implicate dopamine reward pathology as the cause of our present crises (I have no problem calling them that). I previously pointed out that culture is the more conventional culprit. I did not mean by culture, "art, music, etc.", but rather the societal infrastructure – language, laws, Government, economy, industry, media, institutions – which has allowed us to dominate the planet. The author has not convinced me that biology is a better explanation for the crises than culture, understood in this sense.
7) To clarify the difficulty, let us consider drug addiction because this is an area in which dopamine-based reward systems may play an important role in individual pathology (Berridge, Psychopharmacology, 191:391-431 2007). But is the reward system the cause of the addiction crisis? During the Opium Wars of the 19th century, the British East India Company used gunboat diplomacy to force the Chinese Qing dynasty to license opium sales in China, resulting in an addiction crisis that affected a large part (some reports suggest 25%) of the population. Was the crisis due to an "explosive" dopamine system in the Chinese or to the British creating the world's first drug cartel? I think the latter is "the cause" and I suspect most commentators would do the same. This is not to say that the biology is unimportant but it seems more plausible that its role is in deciding who becomes addicted, and who escapes, rather than being the root of the crisis, which lies in culture and society (here, British imperialist trade policy).
8) As I said previously, the author has put forward some fascinating and important scientific ideas that deserve wider attention but I fear these are getting lost in the speculation and personal opinion. This is a perspective, of course, and speculation and personal opinions can sometimes be hugely beneficial in stimulating fresh thinking. I have no problem with that but speculation needs to be clearly seen as such and not dressed up as if it were science. The author states in his rebuttal letter that he has recast his views as a hypothesis and seems to feel that his personal opinions have been adequately segregated. I do not agree. I think these are still very much present in the issues discussed above. The main claim is more clearly a "hypothesis" in the rebuttal than in the revision. In the latter, "hypothesis" is used directly only to refer to "alternatives" and only once used, indirectly, to refer to the "present hypothesis". The speculative nature of what is being claimed and the extent to which it reflects personal opinion is not adequately stated in the new draft.
https://doi.org/10.7554/eLife.36133.008Author response
While the reviewers found the manuscript to be interesting and provocative, they also found it to be "speculative and superficial" in places. It is vital, therefore, that you fully address all the concerns of the reviewers in a revised version of the manuscript. In particular, you need to address the following points:
– Referee 1. Point 1 (which argues against the conclusion that "modern life shrinks reward diversity.")
– Referee 1. Point 2 (which argues against the conclusion that something has "gone wrong"/that we face a "crisis"). Referee 2 makes a similar point in paragraph 7 of his report.
– Referee 2. The comments in paragraph 4 (about extreme teleological language).
– Referee 2. The comments in paragraph 8 (about language and culture).
– Referee 2. The comments in paragraph 9.
– Referee 3. The comments in paragraph 2 (about the role of the human prefrontal cortex)
I heartily thank all three reviewers for their thoughtful comments and vigorous wake-up call. Trying to read the essay through their eyes, I find many instances where rhetorical intensity replaced solid explanation and have revised extensively to fix that. I have also followed reviewer 2’s request for “sharper focus and ruthless editing” – as detailed below. The text plus legends is shorter by about 1000 words.
1) Reviewers 1 and 2 objected to the term “crisis” and to the suggestion that something “has gone wrong”. Accordingly, the Introduction now describes the rise of atmospheric CO2, obesity, and drug addictions as “difficulties” caused by excessive consumption.
The Abstract, Introduction, and final section of the essay mentioned “mass migrations” as one of the “crises”. Since migrations are not directly caused by excessive consumption (the main focus), that material is now deleted. Also deleted is the material on human differences and mental disorder.
2) The essay now traces the evolution of computational capacity constrained by energy efficiency (the topic of my book with Laughlin) to what I identify as a critical event. It then hypothesizes a connection between this event and subsequent “difficulties”. Reviewers 1 and 2 may remain unconvinced, but at least the hypothesis now rests more clearly on evidence from the scientific literature and not my personal opinions.
3) The section, now headed “Multi-cellularity expanded resources and computational capacity”, contained the sentences to which reviewer 2 objected as “extreme teleology”, and I have re-phrased them to meet his concern. I also reviewed the rest of the text with this concern in mind, but I am clearly less sensitive to this issue than reviewer 2, so if he finds additional objectionable phrasings, I’m certainly willing to accommodate.
This section describes several principles of efficient design adopted by early brains, including predictive regulation (allostasis), where a clock governs a primitive neuroendocrine system and hypothalamus. One principle, learn, is introduced as key to predictive regulation along with its efficient mechanism, dopamine reward of better-than-predicted results. This mechanism appeared 500 million years ago with worms, a point that becomes relevant in the essay’s last section.
4) The next section retitled, “To occupy the world H. sapiens required a large, efficient brain”, is reorganized to better explain the degree of computational capacity that humans required to inhabit the entire natural world: a brain 3-fold larger than that of our nearest living relative, the chimpanzee. To grow such a large brain we assume a deep caloric debt that requires a lifetime to repay (70 years). Foraging and hunting skills require such extended learning that productivity doesn't peak until age 45, when foragers are grandparents. Our long life span and economic contributions as grandparents (ages 45-70) are essential to support a birth rate high enough to prevent extinction. This section relies on current anthropology – detailed economic measurements and analysis of modern foragers on three continents with supporting citations. This material represents the current consensus in this field -- not my personal conjecture.
These anthropological data help make sense of the recent MRI finding that tracts for prefrontal cortex that support judgment, choice, planning, and impulse control -- all those responsible behaviors – finally mature during our middle to late 40s and are the last to regress in old age. It seems plausible that foragers would live long enough to benefit from their final brain maturation. The evidence cited does not romanticize the “noble savage”, but neither does it support Hobbes’ description, ‘nasty, brutish, and short”. Reviewer 3 requested additional comment on this prefrontal expansion, and I have added some as part of the section on late-maturing tracts.
5) The next section is also retitled for sharper focus: Expanding the community’s computational capacity. In four paragraphs it notes that specializing individual brains extends our species computational capacity – but entails additional mechanisms for cooperation and relief of tensions. This is where “culture” enters. I address it, as reviewer 2 requested, noting that neural circuits for culture belong to the overall problem of efficient computing – neural circuits for music and art must pay their way. I suggest what seems fairly obvious, that they do so by facilitating long-term cooperation between unrelated specialists. There are undoubtedly many other ways to say this, but mine is at least economical – in the spirit of reviewer 2’s request for ruthless editing.
Various investigators, Boyd and Richerson, for example, discuss co-evolution of culture and genes, but I haven’t found specific reference to the role of culture in enhancing computational capacity by facilitating brain specialization. If reviewer 2 knows a good reference here, that would be great.
6) The next section, “What led to our present difficulties?”, presents the core hypothesis. Since that will be controversial, at least the historical facts should be well accepted – no cherry picking allowed – and for that I draw on a recent authoritative history (Overton). The key effect of factory work was captured by the succinct observation of a contemporary observer, Adam Smith, the father of modern economics. His description contrasts sharply to the life of foragers presented in the preceding section. So far, no personal opinion.
Then comes the essay’s core hypothesis, that modern life shrinks the diversity of better-than-predicted rewards and thus drives the reward learning circuit into positive feedback. The problem is presented more carefully than in the earlier version and hopefully the reasoning is clearer. Even if reviewers 1 and 2 are not ultimately convinced, I believe that the idea is broad and original enough that it deserves consideration – especially since it potentially explains a host of health issues. Thanks to reviewers 1 and 2, the idea now emerges as a hypothesis rather than as sheer opinion larded with a host of other issues.
The next section, “Predictive regulation under positive feedback”, explicitly connects the “difficulties” of CO2, obesity, and drug addiction as systems gone haywire due to positive feedback. Same facts as before, but organized better to address the broad hypothesis.
Finally, I moved the material on hypertension to the end, where it is useful in explaining problems of treating the lowest regulatory levels with drugs.
Specific comments.
i) Reviewer 1 suggests that a forager’s life was repetitive and that his own “modern existence as a scientist living in San Francisco provides a much greater diversity of interesting and unexpected stimuli”. Reviewer 1 objects that I provide no evidence beyond anecdotes.
The essay now offers quantitative evidence of the long-term challenge of foraging and hunting – activities complex enough to require learning into middle age. A hunter, for example, covering a territory of 12,000 km2 seems unlikely to repeat very much – except possibly upon reaching the familiar track to home -- and that would be rewarding.
Of course, rewarding stimuli are numerous for a scientist in San Francisco – and that is exactly my point. Research scientists are indeed rich in diverse activities – probably competing well with a forager in this regard. And correspondingly, our rates of obesity and drug addiction are low. But, if reviewer 1 traded his profession for the job of a supermarket checker, his opinion might change. The essay notes that obesity is most prevalent among the least educated who, if they are employed, have the least rewarding jobs
ii) Reviewer 1 says, “it is reasonable to suggest that obesity or drug addiction may be caused by dysregulated reward systems, but the author instead wants to propose an overarching theory for life on earth”.
If we agree that obesity and drug addiction are caused by a disturbed reward system, we can agree further on the importance of asking what causes that “dysregulation”. The essay presents my hypothetical answer.
The essay is not meant to present “an overarching theory for life on earth”. Rather, it simply tries to explain why consumption is rising in the midst of plenty, where normally one might expect satiety. Since the reward system is a core principle of all animal behavior from worms onward, and since it influences every choice under control of the prefrontal cortex, naturally its dysregulation in humans would have broad effects.
iii) Reviewer 2 [point 8] criticizes the failure to consider genes and culture as contributions to the difficulties of concern here. I do discuss the ‘thrifty gene’ hypothesis and its inadequacies. Beyond that I omit these vast topics – because, unlike Sociobiology, the essay does not attempt to unify all behavior, genetics, and culture. To the contrary, my point is more focused: essentially all cultures and all genetic stocks are struggling with the same difficulty––unbounded consumption––so I don't see where culture would apply.
iv) Reviewer 2 criticizes the failure to distinguish our species from animals. I cannot see a reason to make this distinction because the problem lies, according to my hypothesis, with a mechanism that we share with animals: the reward system. Rats and fruit flies become obese and drug-addicted just like people; for example, a new paper in Current Biology shows that a male fruit fly, restricted in reward diversity by optogenetic inhibition of its ejaculation, drinks more ethanol. All animals are wired for multiple small rewards, and when the possibilities shrink, they all seek ways to get their hits of dopamine.
I cite no evidence that culture and education protect against obesity and addiction because they don’t – except as they promote opportunities for reward diversity, which I do cite.
v) Reviewer 1 argues that we are better off today than at any time in human history. Maybe so for research scientists in San Francisco, but there are now 8 billion members of the “human race”, and a great proportion are living miserably – even in our great cities. However, I prefer not to enter this debate, which has been treated exhaustively by Steven Pinker (among others), so it does not appear in my essay.
vi) Reviewer 2 objects to the conclusions as platitudes. I claim to the contrary that these social policy suggestions emerge from basic principles of human design set forth in the essay. For one example, the principle: enhance group computational capacity by specializing individual brains, cries out for individualized education. Yet public education is anything but that. Furthermore, the low tolerance of many children for a one-size-fits-all classroom evokes behavior diagnosed as ADHD that is widely treated with amphetamines, which of course are also widely consumed as drugs of abuse. A reform that re-individualized education would require a substantial social investment – so this principled reversal of current social policy goes well beyond a platitude.
More generally, the computational richness accumulated in neural circuits over 500 million years leads me to doubt that the “disorders of consumption” will be cured by new drugs that tweak one or another protein receptor. Since that is the only approach expressed these days in leading journals, a principled critique seems overdue. My critique generally provokes outrage; whereas platitudes provoke a yawn.
[Editors' note: further revisions were requested prior to acceptance, as described below.]
We agreed that you need to address some of the points made by Jeremy Gunawardena, and that we will accept the article for publication if you address these points plus a number of editorial points.
Re the comments from Jeremy Gunawardena: Please address the following points:
– point 3
– points 4 and 5 (which are related)
– points 6 and 7 (which are related)
– point 8. (Addressing some of the editorial comments should help address this point)
The attached manuscript now:
i) includes your revisions to the Abstract – except for “treating neural circuits […]” because many of the treatments are somatic as well. My point is that the circuits are complex, so I added that word. Also accepted are your revisions to the Introduction.
ii) attends to all the points that you inserted with bold type. These changes also address the reviewer’s concerns (4 and 5) about the terms “positive feedback” and adaptation, which I have deleted. That section now simply describes what is observed, a “pathological cycle”.
iii) omits the figure regarding reward diversity and adds explanation to figure legends where requested.
iv) adds the carbon dioxide curve with a simple legend and attribution.
v) accepts the reviewer’s request (point 8) to better identify my opinions; have done so at all the points that you kindly suggested.
That leaves the reviewer’s three related points. His point 6 states “It is an enormous leap to implicate dopamine reward pathology as the cause of our present crises.” But that is not my claim and does not accurately represent my argument. I doubt that the neural circuits are pathological. They simply respond to the situation, and when that leads to unfortunate behaviors, it is not their fault.
My concern is unbounded consumption. That, undeniably, is what causes the rise in CO2; it is what causes obesity (and its sequelae); and it perfectly characterizes the cycle of addiction to various intoxicants. So my question is, what causes unbounded consumption?
Researchers in the field of reward learning agree that the various intoxicants drive dopamine release or increase its dwell time at synapses. They also agree that it is for this effect on dopamine that the drugs are consumed without bound. Many researchers also accept that rich foods are consumed to caloric excess for the same reason––to drive the dopamine system. In fact, several studies propose to treat obesity with opioid antagonists. So the connections between obesity, drugs of addiction, and the dopamine reward system are clear and uncontroversial.
It is also widely recognized that unbounded participation in other activities, such as work, gambling, sex, and so on, also release dopamine and can drive an addictive cycle. Hence the expression “workaholic” and the 12-step programs and support groups for diverse unbounded behaviors.
Authorities on reinforcement learning agree that learning by machines and animals, starts with a constitutive seeking of reward. A computation predicts a behavior, and when the result is better than predicted (positive reward-prediction error), the machine gets a “reward” signal, and the animal gets a pulse of dopamine. It is also agreed that, when sources of positive reward-prediction error shrink, the machine and the animal will seek a familiar reward for which the only positive prediction error is greater intensity; i.e., more. This is in the math and in its recognized biological instantiation.
The above generally accepted points lead me to hypothesize that unbounded consumption in modern society arises from reduced reward diversity that leaves more as the main source of positive reward-prediction error. The reviewer disputes this (point 3) and complains that I do not explain how to measure the effects of shrinking reward diversity. A nice example of such a measurement has just appeared: male fruit flies, deprived optogenetically of the reward from ejaculation, drink more ethanol (Zer-Krispil et al. 2018 Ejaculation induced by the activation of Crz neurons is rewarding to Drosophila males Current Biol. 28:1445-1452).
Regarding humans, it seems as obvious today, as it did to Adam Smith, that simplified tasks, which constitute many of the ‘jobs’ in modern society, do not nearly approach the challenges posed by foraging – for which our brain evolved. I have expanded slightly the text to explain this. In short, the chain of reasoning about the reward system and the consequence of shrinking reward diversity is well founded in current thinking.
The reviewer’s comment about a laborer in Cheops’ pyramid (point 3) is not relevant because I do not try to characterize every type of labor across 150 millennia. I simply compare foraging to modern life. However, to reduce a possible source for minor dispute, I have deleted my minor comment about “monumental constructions”. Regarding the reviewer’s comment regarding the Opium Wars (point 7): of course, the reward system can be manipulated for devious ends. Provide cheap opium and many will become addicted; the same is true today for cheap fructose syrup. But that is not relevant to my hypothesis, namely that the current cause of unbounded consumption is the loss of reward diversity.
https://doi.org/10.7554/eLife.36133.009Article and author information
Author details
Acknowledgements
I thank the following individuals for their encouragement and comments on various drafts of this essay: Dean Astumian, Sally Zigmond, Simon Laughlin, Michael Mullin, Alan Pearlman, Robert Collins, Price Peterson, Irving Seidman, Wolfram Schultz, Gino Segre, Dost Ongur, Scott Poethig, Jonathan Demb, Olivia Demb, Andy Barto, Ann Stuart, Guy Lanza, Jose Tapia Granados, Nica Borgese, Stan Schein, Richard Masland, Bart Borghuis, Kai Kaila, Marla Feller and Adrianne Fairhall. I also thank Iris Broudy for preparing the manuscript.
Publication history
- Received:
- Accepted:
- Version of Record published:
Copyright
© 2018, Sterling et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 5,502
- views
-
- 572
- downloads
-
- 17
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 17
- citations for umbrella DOI https://doi.org/10.7554/eLife.36133
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Point of View: Predictive regulation and human design
eLife 7:e36133.
https://doi.org/10.7554/eLife.36133




{kind=link}
{kind=link}
{kind=link}
{kind=link}