RNA-binding proteins distinguish between similar sequence motifs to promote targeted deadenylation by Ccr4-Not
- MRC Laboratory of Molecular Biology, United Kingdom
Figures
Figure 1 with 1 supplement
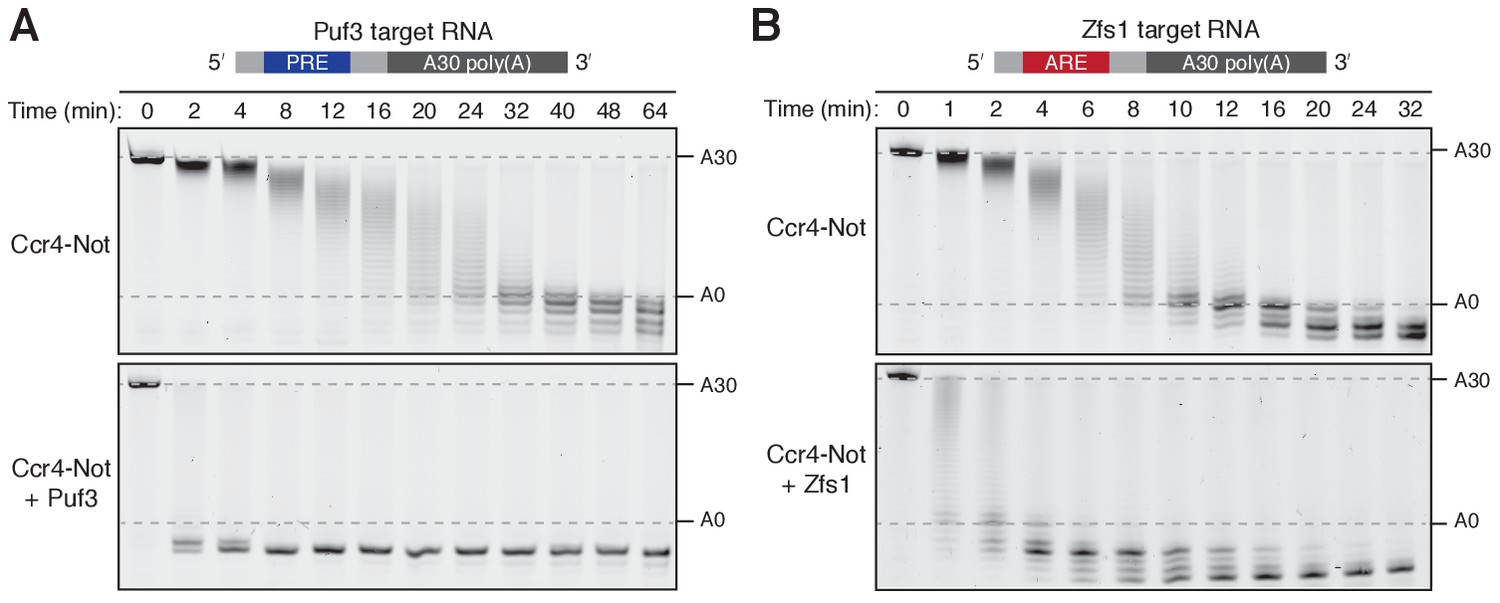
Puf3 and Zfs1 accelerate deadenylation by Ccr4-Not in vitro.
Deadenylation by purified Ccr4-Not in the absence and presence of (A) Puf3 and (B) Zfs1. Synthetic RNA substrates were designed to contain a 30-adenosine poly(A) tail downstream of 23 non-poly(A) ribonucleotides including the recognition elements of Puf3 (PRE) or Zfs1 (ARE). RBP-RNA complexes were prepared at a 1:1 molar ratio before the addition of Ccr4-Not. Reactions were stopped at the indicated times and analyzed by denaturing PAGE. The sizes of the RNA substrate with and without the poly(A) tail are indicated by dashed lines.
Figure 1—figure supplement 1
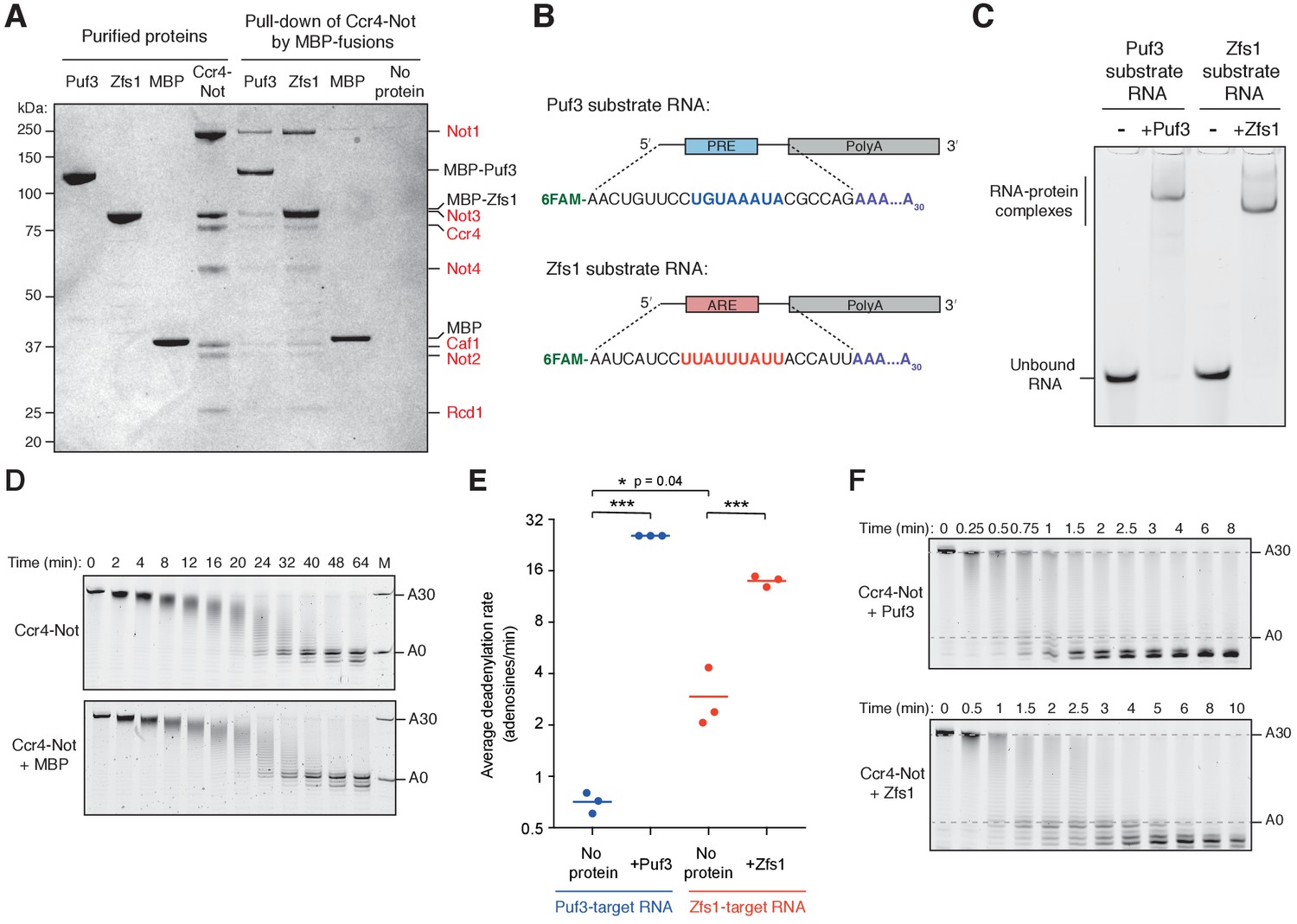
Acceleration of deadenylation by Puf3 and Zfs1.
(A) Coomassie-stained SDS-PAGE of pull-down assay showing binding of Ccr4-Not (subunits labelled in red) to immobilized MBP-Puf3 and MBP-Zfs1. (B) Schematic diagram of RNA substrates showing 5ʹ 6-FAM fluorescent label, position of Pumilio-response element (PRE; blue), AU-rich element (ARE; red), and 30-adenosine poly(A) tail. (C) Electrophoretic mobility shift assay (EMSA) showing the protein-RNA complexes used as substrates for deadenylation assays. Puf3 or Zfs1 (250 nM) was incubated with labelled RNA (200 nM) for 15 min at room temperature before resolving on a native polyacrylamide gel. (D) Control deadenylation assay with Puf3-target RNA showing that MBP alone does not have an effect on the deadenylation activity of Ccr4-Not. M is marker for RNA with and without a poly(A)30 tail. (E) Average rates of Ccr4-Not-mediated deadenylation in the presence of Puf3 or Zfs1. Reaction rates were calculated by densitometric analysis of denaturing polyacrylamide gels. The most abundant RNA size decreased linearly with time, and average reaction rates in number of adenosines removed/min were determined by linear regression on triplicate measurements. Each experiment is presented as a single data point, and the mean of each triplicate experiment is plotted as a horizontal line. Statistical significance was calculated by one-way ANOVA in GraphPad Prism. *p=0.04; ***p=0.001. (F) Fully-deadenylated and non-deadenylated RNA exist simultaneously in reactions with Ccr4-Not and Puf3 or Zfs1. Experiments were performed as in Figure 1 with shorter time increments.
Figure 2 with 2 supplements
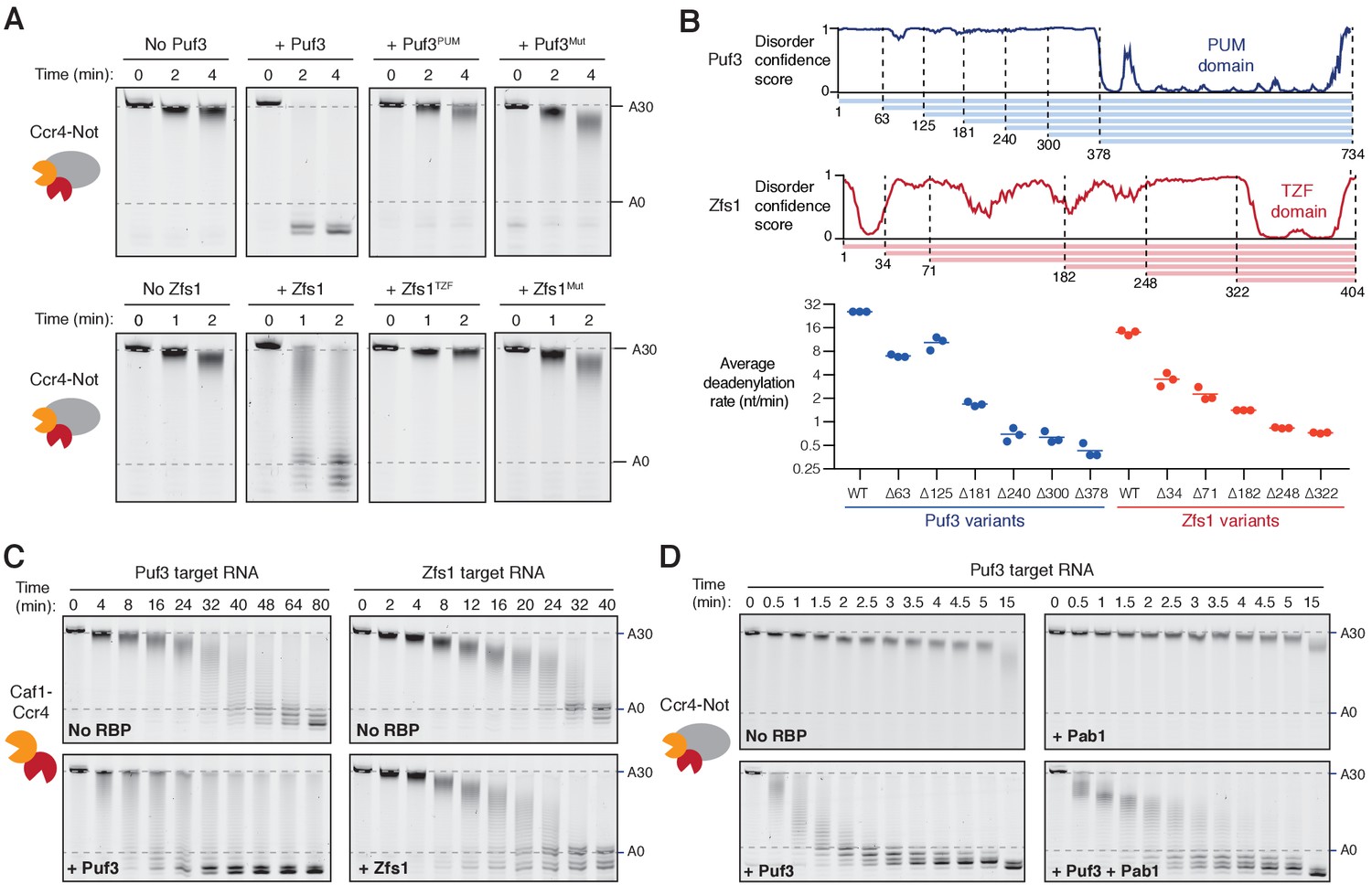
Recruitment of Ccr4-Not by Puf3 and Zfs1.
Accelerated deadenylation by RNA binding proteins requires both a low-complexity region and RNA-binding activity. (A) Deadenylation by purified Ccr4-Not in the presence of the indicated RNA-binding protein variants was analyzed by denaturing PAGE. Puf3PUM and Zfs1TZF each comprise the RNA-binding domain alone; Puf3Mut, Zfs1Mut are full-length proteins with point mutations that abolish RNA binding. Panels for no Puf3, no Zfs1, and wild-type proteins are reproduced from Figure 1. (B) The low-complexity domains of Puf3 and Zfs1 contain multiple regions that contribute to deadenylation by Ccr4-Not. The upper panel shows diagrams of Puf3 and Zfs1 with regions of predicted disorder and truncated construct lengths. The bottom panel shows deadenylation by purified Ccr4-Not in the presence of the indicated protein variants, analyzed by denaturing PAGE. The most abundant RNA size decreased linearly with time, and average reaction rates in number of adenosines removed/min were determined by linear regression on triplicate measurements. Each experiment is plotted as a point and the mean is shown as a horizontal line. Statistical significance is shown in Figure 2—figure supplement 1G. (C) Deadenylation by the Caf1-Ccr4 nuclease subcomplex in the absence (No RBP) and presence of Puf3 and Zfs1. (D) Puf3-stimulated deadenylation by Ccr4-Not in the absence or presence of Pab1 (1:1 molar ratio to RNA).
Figure 2—figure supplement 1
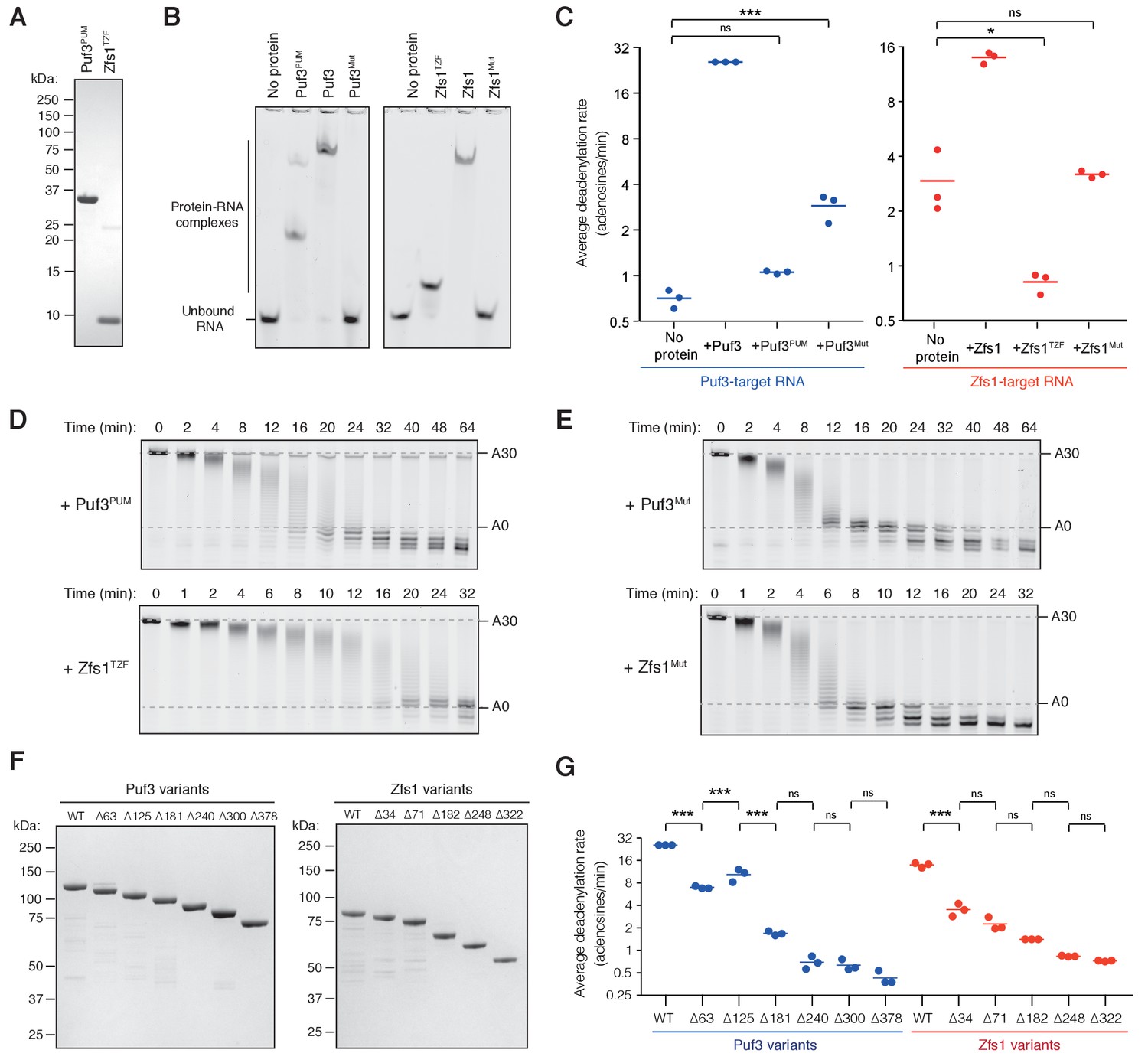
Characterization of Puf3 and Zfs1 domains required for accelerated deadenylation.
(A) Coomassie-stained SDS-PAGE of purified RNA-binding domains: Puf3 residues 378–714 (Puf3PUM) and Zfs1 residues 322–392 (Zfs1TZF). (B) Electrophoretic mobility shift assay (EMSA) showing binding of the indicated RNA-binding proteins to poly(A)-containing RNAs used as substrates in deadenylation assays. (C) Average rates of Ccr4-Not-mediated deadenylation in the presence of the indicated Puf3 or Zfs1 variants. Reaction rates were calculated by densitometric analysis of denaturing polyacrylamide gels. The most abundant RNA size decreased linearly with time, and average reaction rates in number of adenosines removed/min were determined by linear regression on triplicate measurements. Each experiment is presented as a single data point, with the mean (n = 3) shown as a horizontal line. Statistical significance was calculated by one-way ANOVA in GraphPad Prism. *, p=0.05; ***, p=0.001, n.s., not significant. (D, E) Full deadenylation reactions performed in the presence of the indicated Puf3 or Zfs1 variants as in Figure 2A. (F) Coomassie-stained SDS-PAGE of purified Puf3 and Zfs1 N-terminal truncation variants. (G) Average rates of Ccr4-Not-mediated deadenylation in the presence of the indicated Puf3 or Zfs1 variants. The p-values are as in panel (C).
Figure 2—figure supplement 2
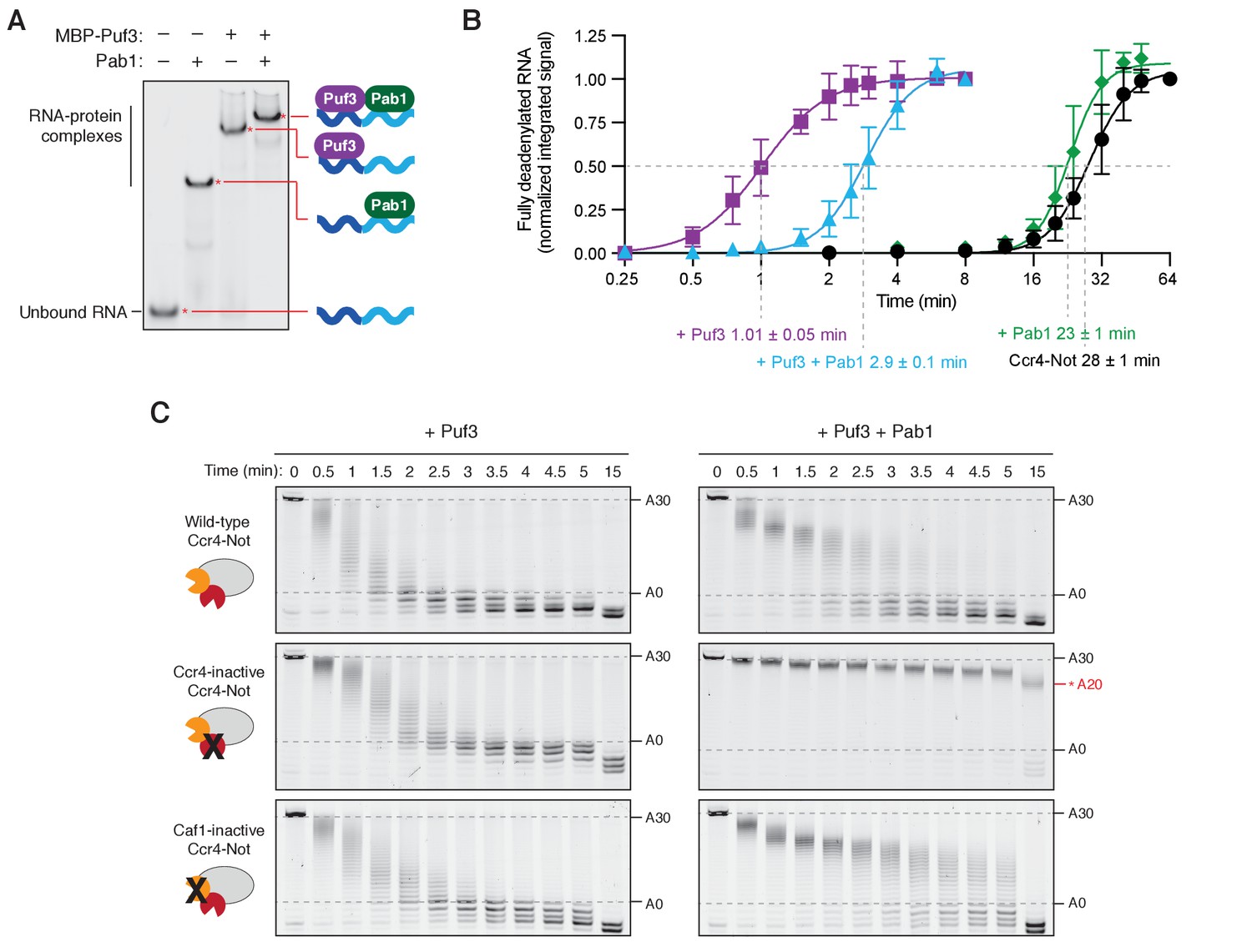
Puf3 accelerates deadenylation in the presence of Pab1.
(A) Electrophoretic mobility shift assay showing the binding of MBP-Puf3 and Pab1 to the deadenylation substrate containing a Puf3 motif upstream of a 30-adenosine poly(A) tail. (B) Puf3 accelerates release of Pab1 during deadenylation by Ccr4-Not. Densitometric analysis of previously published data (Webster et al., 2018) and the reactions shown in Figure 2D to calculate the amount of fully deadenylated RNA. A quadratic fit of the data was used to determine the time taken for half of the RNA to be completely deadenylated. Reactions were performed in triplicate and the standard error is given. (C) Deadenylation by Ccr4-Not and variants with mutations in the active site of either Ccr4 (Ccr4-inactive) or Caf1 (Caf1-inactive) with Pab1 and Puf3. Reactions with wild-type Ccr4-Not are reproduced from Figure 2D. In the presence of Pab1, Ccr4-inactive Ccr4-Not stalls when ~ 20 As remain, as indicated.
Figure 3 with 1 supplement
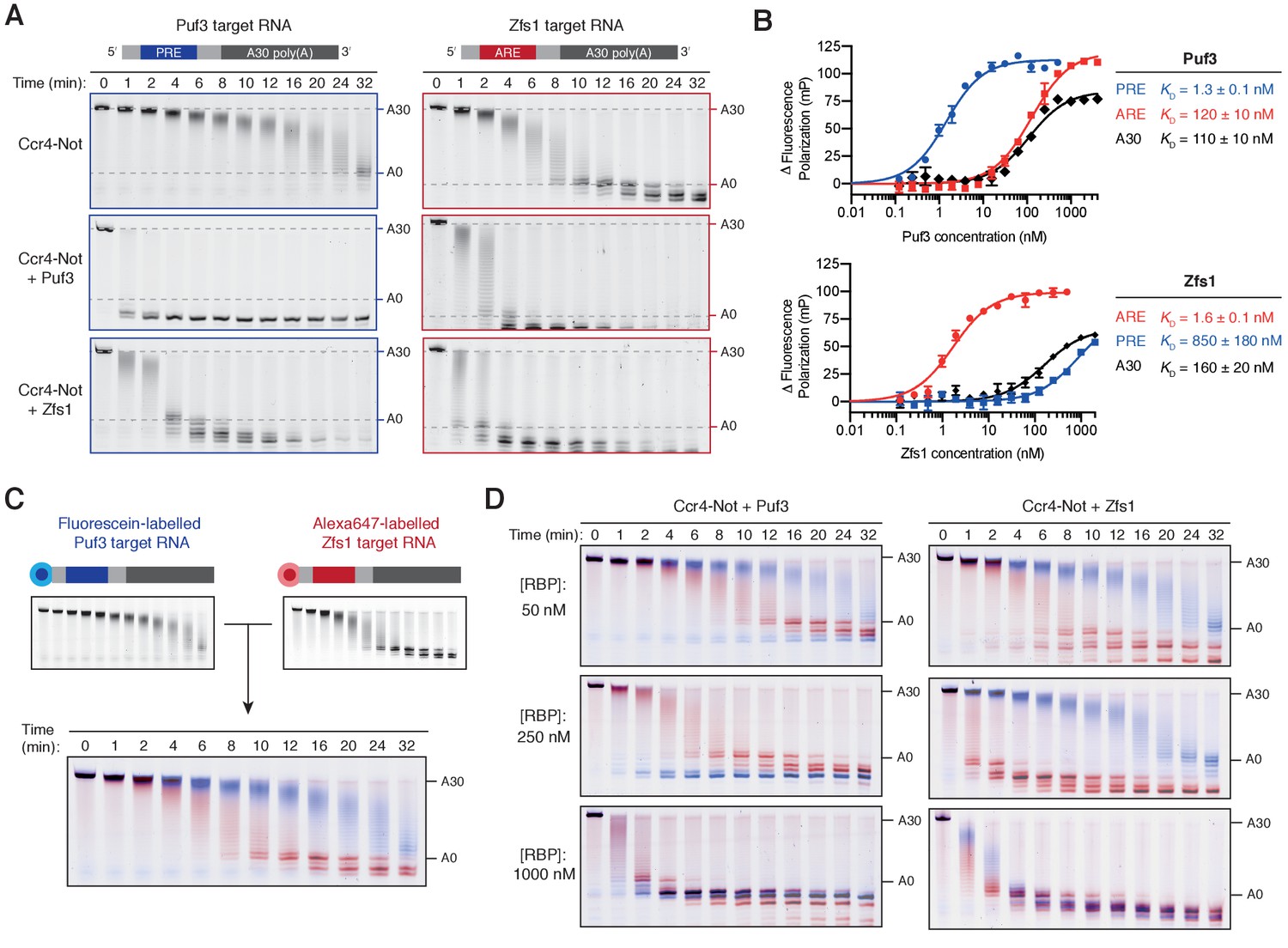
Puf3 and Zfs1 mediate selective deadenylation by Ccr4-Not.
(A) Puf3 and Zfs1 accelerate deadenylation of RNAs with or without a target recognition motif. Puf3 or Zfs1 was added to PRE-containing RNA (Puf3-target) or ARE-containing RNA (Zfs1-target) before addition of Ccr4-Not. Deadenylation was analyzed by denaturing PAGE. Top right and bottom right panels are reproduced from Figure 1B. (B) Fluorescence polarization experiments assaying the binding of full-length Puf3 (top) or Zfs1 (bottom) to 5ʹ−6FAM labelled RNAs. Error bars represent the standard error in triplicate measurements. (C) Deadenylation of a mixture of 100 nM fluorescein-labelled Puf3-target RNA (blue) and 100 nM Alexa647-labelled Zfs1-target RNA (red) analyzed by denaturing PAGE. Gels were scanned to detect each fluorophore before signals were overlaid. (D) Puf3 and Zfs1 promote selective deadenylation. Reactions were performed as in (C) but with the addition of Puf3 (left) or Zfs1 (right) at the indicated concentrations.
Figure 3—figure supplement 1
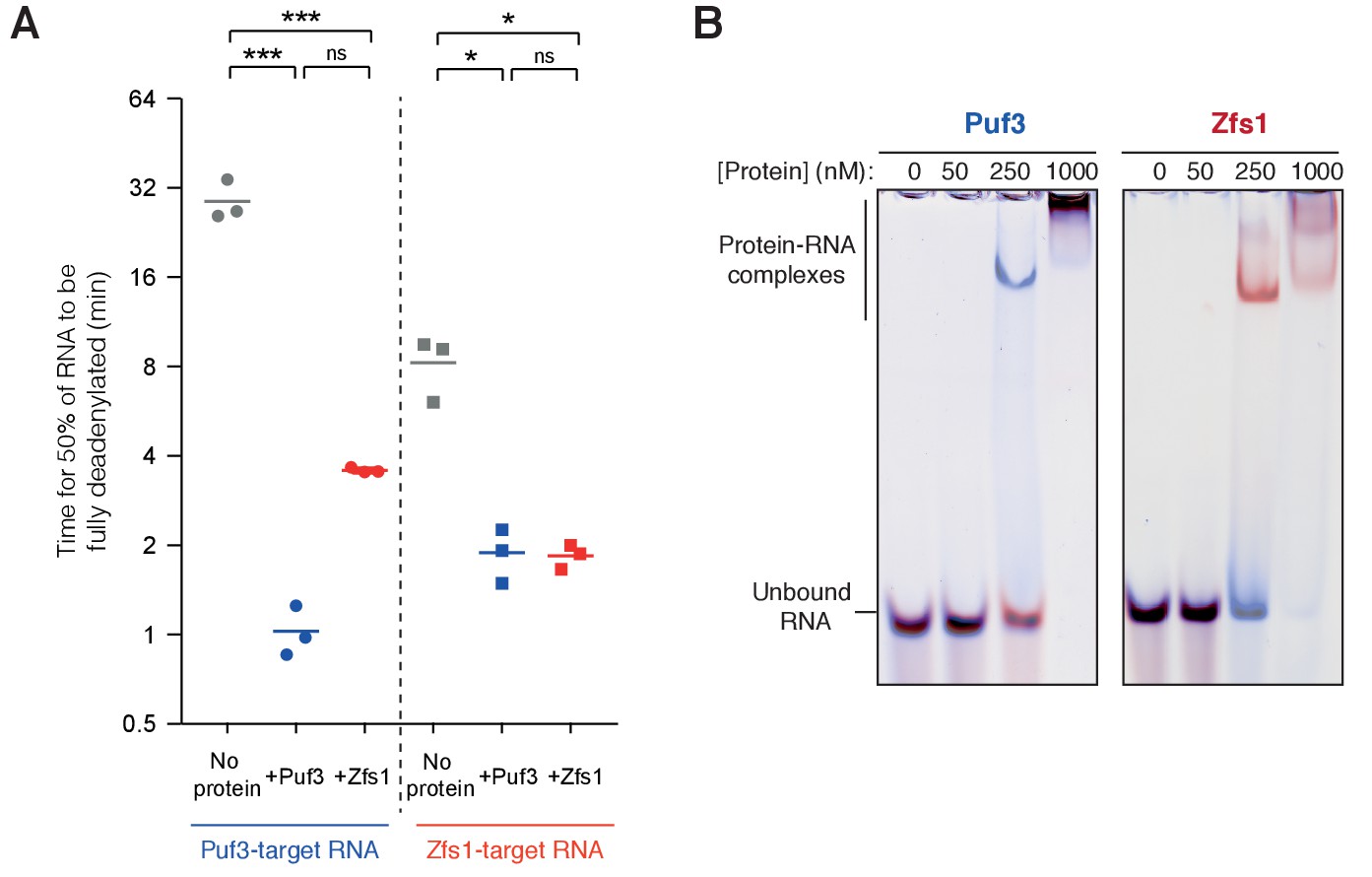
Analysis of RNA selectivity during deadenylation.
(A) Quantification of on-target and off-target deadenylation rates. Densitometric analysis of the amount of fully deadenylated RNA was performed on gels shown in Figure 3A. Each experiment is presented as a single data point, and the mean of triplicate experiments is plotted as a horizontal line. Time for complete deadenylation, rather than change in modal poly(A) tail length, was calculated because these reactions contained low levels of intermediates between non-deadenylated and fully-deadenylated RNA. ns, not significant; *p=0.05; ***p=0.001. (B) Electrophoretic mobility shift assays showing selective RNA binding of Puf3 and Zfs1. A mixture of 100 nM fluorescein-labelled Puf3-target-A30 RNA (red) and 100 nM Alexa647-labelled Zfs1-target-A30 RNA (blue) was incubated with the indicated concentration of Puf3 or Zfs1. Gels were scanned with a Typhoon FLA-7000 with excitation at 478 nm (fluorescein detection) and 633 nm (Alexa647 detection), and images were superimposed with color overlay.
Figure 4
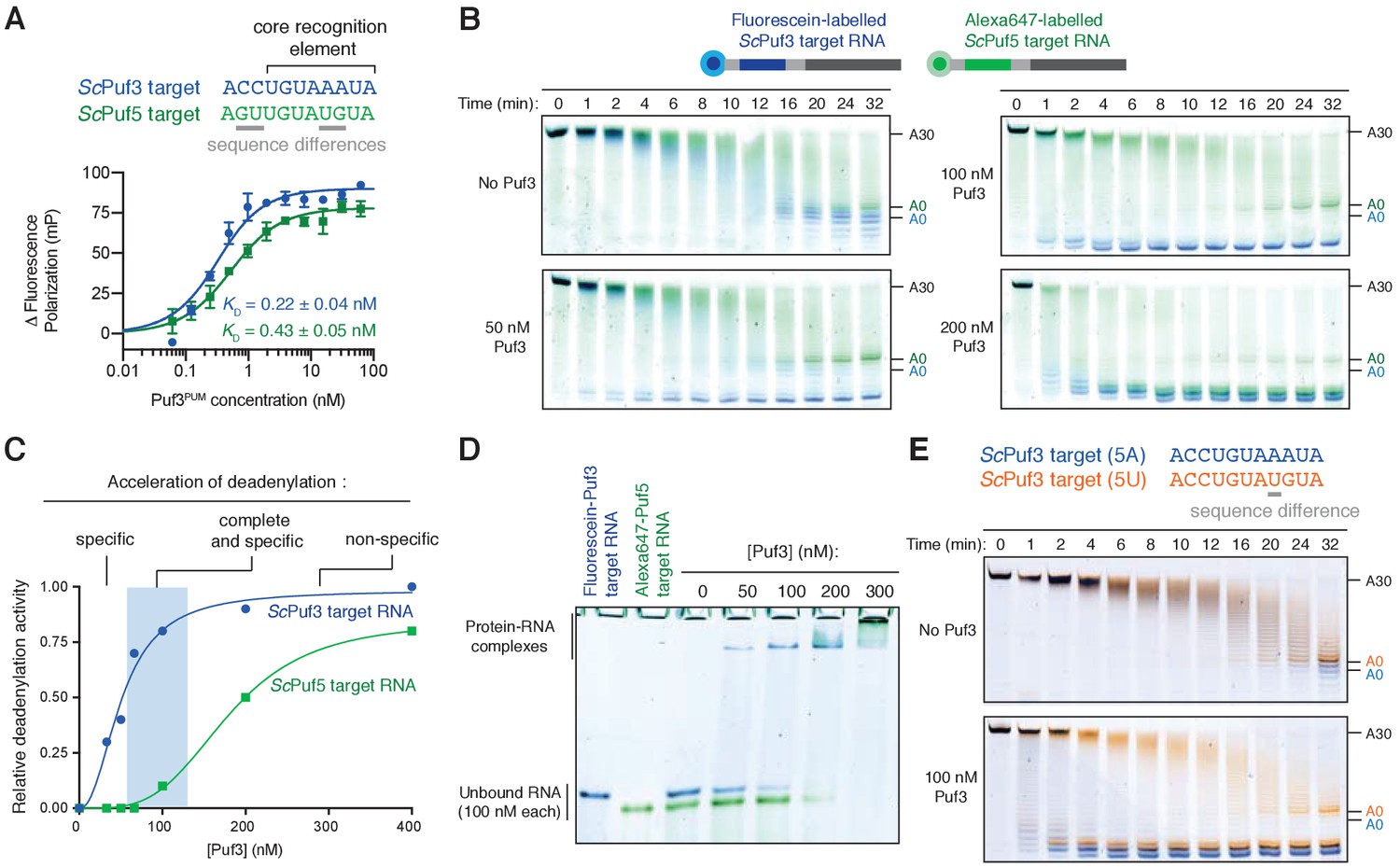
Puf3 distinguishes between similar RNA motifs.
(A) Fluorescence polarization experiments assaying the binding of Puf3 RNA-binding domain to RNA containing a ScPuf3 motif (blue) or a ScPuf5 motif (green). Error bars represent the standard error in triplicate measurements. (B) Deadenylation of a mixture of fluorescein-labelled ScPuf3-target RNA (blue) and Cy3-labelled ScPuf5-target RNA (green) analyzed by denaturing PAGE. Gels were scanned to detect each fluorophore before signals were overlaid. (C) Selective deadenylation occurs within a narrow range of Puf3 concentrations. Plot of relative deadenylation rates of ScPuf3-target RNA and ScPuf5-target RNA in reactions as in (B) at a series of Puf3 concentrations. Reactions were quantified based on the fraction of RNA fully deadenylated within 8 min. (D) Electrophoretic mobility shift assay showing the binding of Puf3 to a mixture of fluorescein-labelled ScPuf3-target RNA (blue) and Cy3-labelled ScPuf5-target RNA (green). The concentration of each RNA was 100 nM. (E) Deadenylation of a mixture of fluorescein-labelled ScPuf3(5A)-target RNA (blue) and Alexa647-labelled ScPuf3(5U)-target RNA (orange) analyzed by denaturing PAGE. Gels were scanned to detect each fluorophore before signals were overlaid.
Figure 5

RNA motif quality affects Puf3 binding kinetics.
(A) The kinetic rates of association (kon) of purified Puf3PUM with RNA containing the indicated S.cerevisiae sequence motifs were determined by SwitchSENSE. Error bars are the standard error. (B) Representative SwitchSENSE sensograms showing the dissociation of Puf3PUM from the indicated S. cerevisiae RNA sequences. dru, dynamic response units. In (A) and (B), rate constants and half-lives are shown with standard error for measurements performed in triplicate.
Figure 6 with 1 supplement
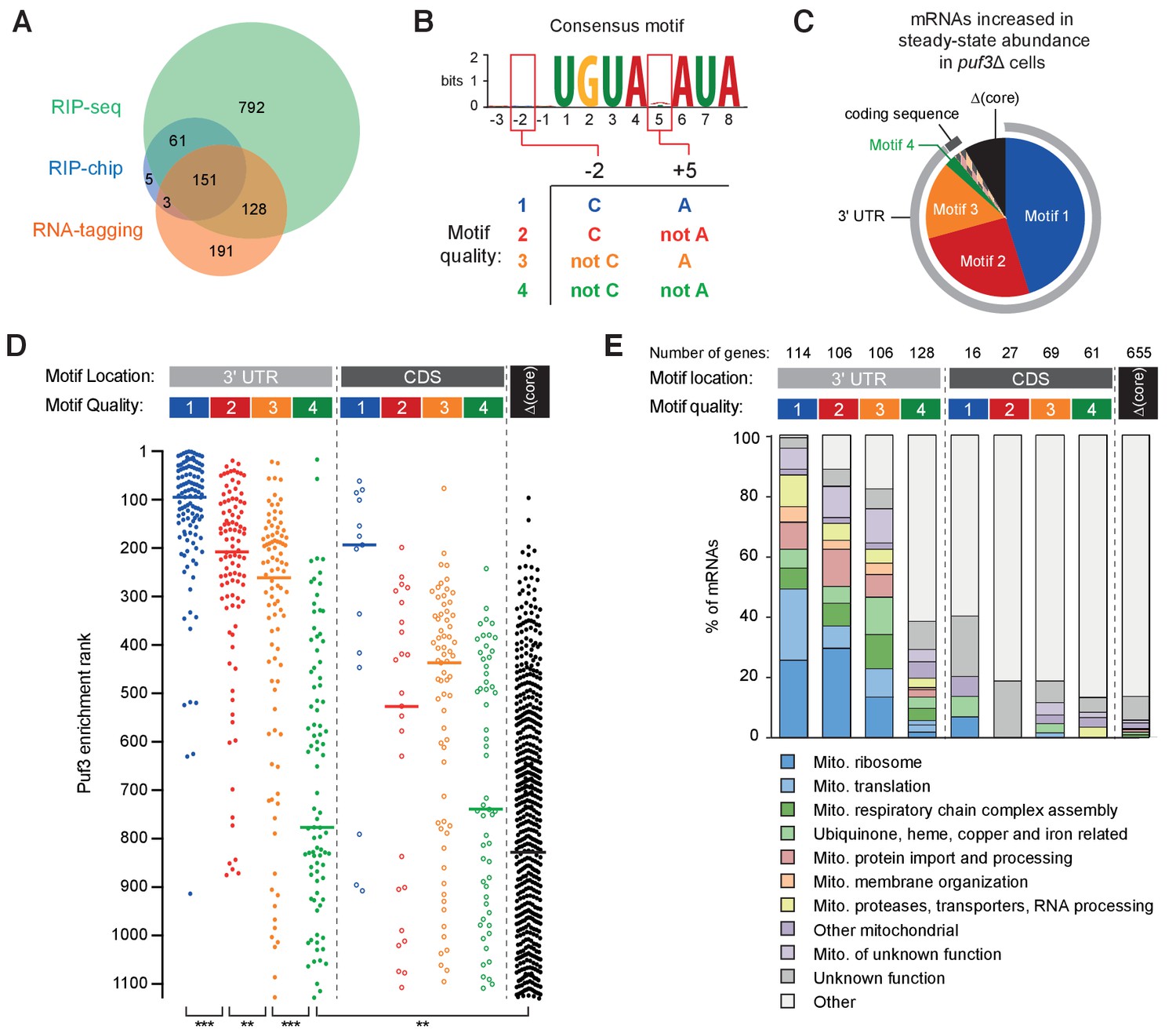
RNA motif quality defines a coherent mRNA regulatory network.
(A) Overlap between the putative mRNAs bound by S.cerevisiae Puf3 determined by RIP-seq (Kershaw et al., 2015), RIP-chip (Gerber et al., 2004) and RNA-tagging (Lapointe et al., 2015). The numbers of mRNAs identified in each study is given. (B) Consensus motif identified from the complete set of putative ScPuf3 mRNA targets. Definition of categories of Puf3 motif quality is shown below. (C) Diagram showing RNA motif quality and location in mRNAs that increase in abundance upon PUF3 deletion (Kershaw et al., 2015). (D) ScPuf3 binding versus RNA motif quality and location. Median enrichment rank is indicated with horizontal line. Statistical significance was evaluated with a Mann-Whitney rank comparison test. **, p=0.01; ***, p=0.001. (E) mRNAs with high quality ScPuf3-target motifs encode proteins related to mitochondrial biogenesis.
Figure 6—figure supplement 1
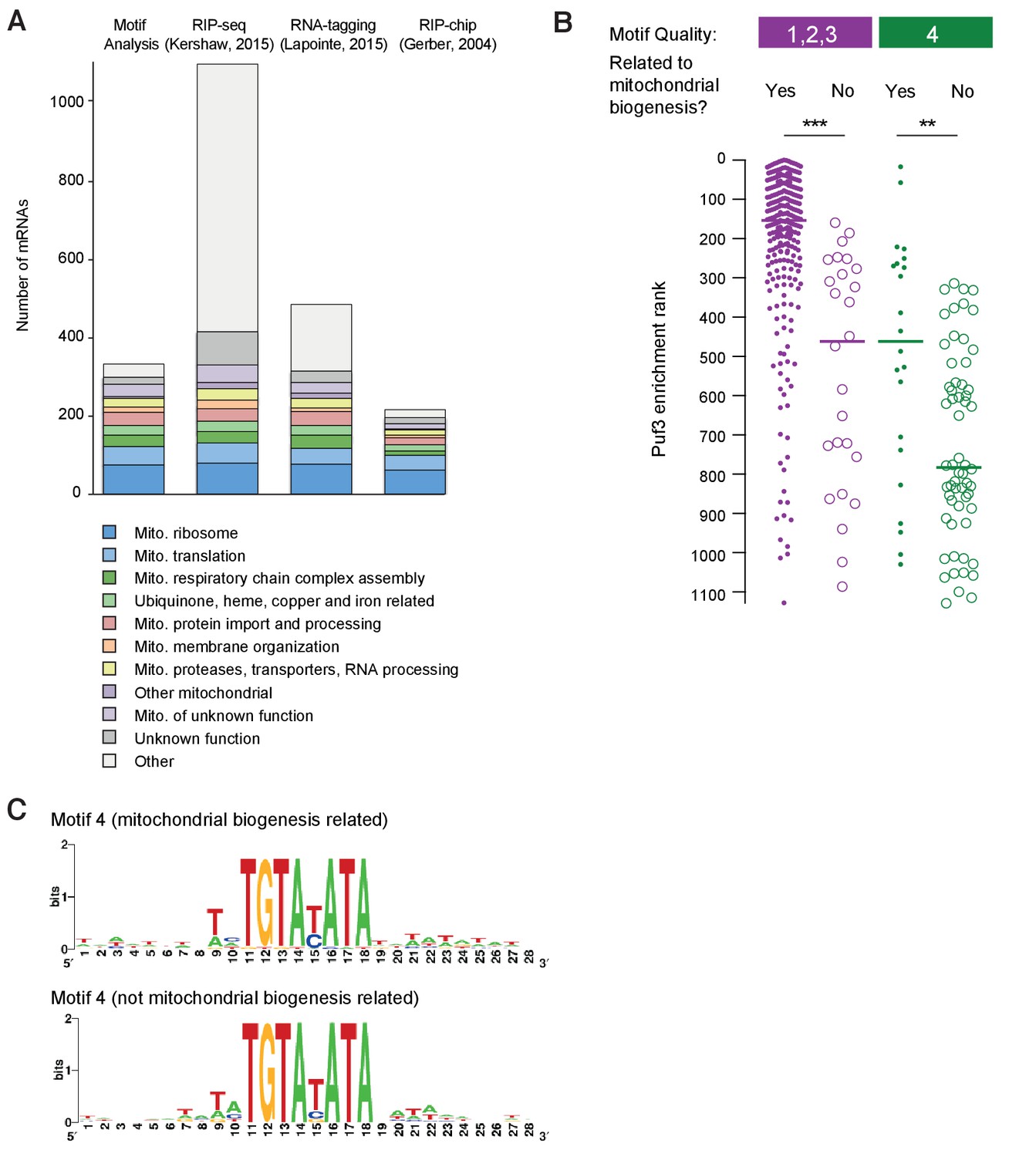
Evaluation of mRNA target prediction by motif quality analysis.
(A) Comparison of the encoded functions of mRNAs identified as targets of ScPuf3 by different methods. (B) Within a single motif category, mRNAs encoding proteins related to mitochondrial biogenesis are more enriched in ScPuf3 binding. Statistical significance was evaluated with a Mann-Whitney rank comparison test. **p=0.01; ***p=0.001. (C) Comparison of RNA sequence context of motifs in category 4 between mRNAs that encode mitochondrial biogenesis proteins and those that do not.
Figure 7
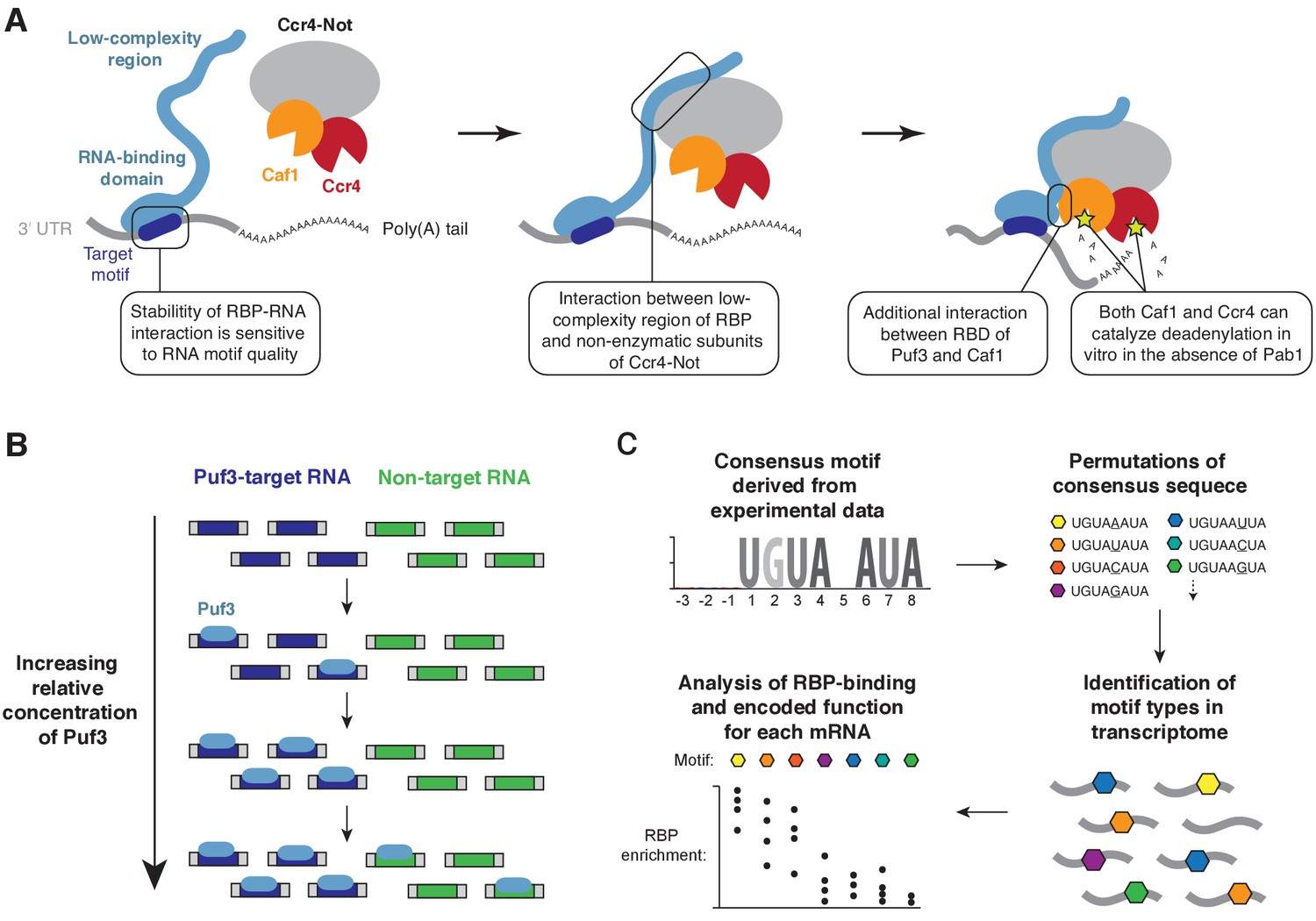
Models for substrate selectivity in deadenylation.
(A) RNA-binding proteins (RBPs) promote targeted deadenylation by simultaneously binding to RNA via an RNA-binding domain (RBD) and to Ccr4-Not via a low-complexity region. These interactions can be regulated by post-translational modifications. (B) Model for priority-based binding of Puf3 to RNA. (C) Scheme for defining target sequences of RNA-binding proteins.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Escherichia coli) | E. coli BL21 star (DE3) | Thermo Fisher Scientific | C601003 | |
| Strain, strain background (Escherichia coli) | E. coli DH5α (DE3) | Thermo Fisher Scientific | 18258012 | |
| Cell line (Spodoptera frugiperda) | Sf9 | Thermo Fisher Scientific | 11496015 | |
| Peptide, recombinant protein | Ccr4-Not | PMID: 27851962 | Purified recombinant protein expressed in Sf9 cells following infection with viruse produced from LP_Ccr4Not | |
| Peptide, recombinant protein | Ccr4-Not (Ccr4-inactive) | PMID: 27851962 | Purified recombinant protein with Ccr4(E387) expressed in Sf9 cells following infection with virus produced from LP_P22-60 | |
| Peptide, recombinant protein | Ccr4-Not (Caf1-inactive) | PMID: 27851962 | Purified recombinant protein with Caf1(D53A) expressed in Sf9 cells followinginfection with virus produced from LP_P22-61 | |
| Peptide, recombinant protein | MBP-Puf3 | this paper | Purified recombinant protein expressed inE. coli BL21star (DE3) following transformation with LP_P22-27 | |
| Peptide, recombinant protein | MBP-Puf3(Δ63) | this paper | Purified recombinant protein expressed inE. coli BL21star (DE3) followingtransformation with LP_P22-28 | |
| Peptide, recombinant protein | MBP-Puf3(Δ125) | this paper | Purified recombinant protein expressed inE. coli BL21 star (DE3) following transformation with LP_P22-29 | |
| Peptide, recombinant protein | MBP-Puf3(Δ181) | this paper | Purified recombinant protein expressed in E. coli BL2 1 star (DE3) following transformation with LP_P22-30 | |
| Peptide, recombinant protein | MBP-Puf3(Δ240) | this paper | Purified recombinant protein expressed in E. coli BL21 star (DE3) following transformation with LP_P22-31 | |
| Peptide, recombinant protein | MBP-Puf3(Δ300) | this paper | Purified recombinant protein expressed in E. coli BL21star (DE3) following transformation with LP_P22-32 | |
| Peptide, recombinant protein | MBP-Puf3(Δ378) | this paper | Purified recombinant proteinexpressed inE. coli BL21 star (DE3) following transformation with LP_P22-33 | |
| Peptide, recombinant protein | MBP-Puf3(Mut) | this paper | Purified recombinant protein expressed inE. coli BL21star (DE3) following transformation with LP_P22-34 | |
| Peptide, recombinant protein | Puf3(PUM) | this paper | Purified recombinant protein expressed in E. coli BL21 star (DE3) following transformation with LP_P22-35 | |
| Peptide, recombinant protein | MBP-Zfs1(FL) | this paper | Purified recombinant proteinexpressed in E. coli BL21 star (DE3) following transformation with LP_P22-36 | |
| Peptide, recombinant protein | MBP-Zfs1(Δ34) | this paper | Purified recombinant protein expressed in E. coli BL21star (DE3) followingtransformation with LP_P22-37 | |
| Peptide, recombinant protein | MBP-Zfs1(Δ71) | this paper | Purified recombinant protein expressed in E. coliBL21 star (DE3) following transformation with LP_P22-38 | |
| Peptide, recombinant protein | MBP-Zfs1(Δ182) | this paper | Purified recombinant protein expressed inE. coli BL21 star (DE3) followingtransformation with LP_P22-39 | |
| Peptide, recombinant protein | MBP-Zfs1(Δ248) | this paper | Purified recombinant protein expressed in E. coli BL21 star (DE3) following transformation with LP_P22-40 | |
| Peptide, recombinant protein | MBP-Zfs1(Δ322) | this paper | Purified recombinant proteinexpressed in E. coli BL21 star (DE3) following transformation with LP_P22-41 | |
| Peptide, recombinant protein | MBP-Zfs1(Mut) | this paper | Purified recombinant protein expressed in E. coli BL21 star (DE3) following transformation with LP_P22-42 | |
| Peptide, recombinant protein | Zfs1(TZF) | this paper | Purified recombinant protein expressed in E. coli BL21star (DE3) following transformation with LP_P22-43 | |
| Peptide, recombinant protein | Pab1 | PMID: 29932902 | ||
| Commercial assay or kit | Quikchange Lightning Multi Site-Directed Mutagenesis Kit | Agilent Technologies | 210513 | |
| Commercial assay or kit | In-Fusion HD Cloning Kit | Takara Bio | 121416 | |
| Commercial assay or kit | Phusion High-Fidelity DNA Polymerase | New England BioLabs | M0530S | |
| Recombinant DNA reagent | MultiBac-Ccr4Not | PMID: 27851962 | LP_Ccr4Not | MultiBac expression vectorfor S. pombe Ccr4-Not |
| Recombinant DNA reagent | MultiBac-Ccr4Not(Ccr4 E387A) | PMID: 27851962 | LP_P22-60 | MultiBac expression vector forS. pombe Ccr4-Not (Ccr4 E387A) |
| Recombinant DNA reagent | MultiBac-Ccr4Not(Caf1 D53A) | PMID: 27851962 | LP_P22-61 | MultiBac expression vector forS. pombe Ccr4-Not (Caf1 D53A) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(FL) | this paper | LP_P22-27 | pMAL-c2x expression vector forS. pombe Puf3(FL) (res1-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Δ63) | this paper | LP_P22-28 | pMAL-c2x expression vector for S. pombe Puf3(Δ63) (res63-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Δ125) | this paper | LP_P22-29 | pMAL-c2x expression vector forS. pombe Puf3(Δ125) (res125-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Δ181) | this paper | LP_P22-30 | pMAL-c2x expression vector forS. pombe Puf3(Δ181) (res181-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Δ240) | this paper | LP_P22-31 | pMAL-c2x expression vector for S. pombe Puf3(Δ240) (res240-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Δ300) | this paper | LP_P22-32 | pMAL-c2x expression vector for S. pombe Puf3(Δ300) (res300-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Δ378) | this paper | LP_P22-33 | pMAL-c2x expression vector for S. pombe Puf3(Δ378) (res378-734) |
| Recombinant DNA reagent | pMAL-c2x-Puf3(Mut) | this paper | LP_P22-34 | pMAL-c2x expression vector for S. pombe Puf3(Mut) (res1-734; Y449A, Y671A) |
| Recombinant DNA reagent | pGEX6P-2-Puf3(PUM) | this paper | LP_P22-35 | pGEX6P-2 expression vector for S. pombe Puf3(PUM) (res378-714) |
| Recombinant DNA reagent | pMAL-c2x-Zfs1(FL) | this paper | LP_P22-36 | pMAL-c2x expression vector forS. pombe Zfs1(FL) (res1-404) |
| Recombinant DNA reagent | pMAL-c2x-Zfs1(Δ34) | this paper | LP_P22-37 | pMAL-c2x expression vector forS. pombe Zfs1(Δ34) (res34-404) |
| Recombinant DNA reagent | pMAL-c2x-Zfs1(Δ71) | this paper | LP_P22-38 | pMAL-c2x expression vector forS. pombe Zfs1(Δ71) (res71-404) |
| Recombinant DNA reagent | pMAL-c2x-Zfs1(Δ182) | this paper | LP_P22-39 | pMAL-c2x expression vector for S. pombe Zfs1(Δ182) (res182-404) |
| Recombinant DNA reagent | pMAL-c2x- Zfs1(Δ248) | this paper | LP_P22-40 | pMAL-c2x expression vector for S. pombe Zfs1(Δ248) (res248-404) |
| Recombinant DNA reagent | pMAL-c2x- Zfs1(Δ322) | this paper | LP_P22-41 | pMAL-c2x expression vector for S. pombe Zfs1(Δ322) (res322-404) |
| Recombinant DNA reagent | pMAL-c2x-Zfs1(Mut) | this paper | LP_P22-42 | pMAL-c2x expression vector for S. pombe Zfs1(Mut) (res1-404; F349A, F387A) |
| Recombinant DNA reagent | pGEX6P-2-Zfs1(TZF) | this paper | LP_P22-43 | pGEX6P-2 expression vector forS. pombe Zfs1(TZF) (res699-769) |
| Recombinant DNA reagent | pGEX6P-2 -Pab1 | PMID: 29932902 | LP_P22-11 | pGEX6P-2 expression vector forS. pombe Pab1 (res80-653) |
| Software, algorithm | ImageJ | NIH | https://imagej.nih.gov/ij/ | |
| Software, algorithm | GraphPad Prism 6 | GraphPad | https://www.graph pad.com/scientific- software/prism/ | |
| Software, algorithm | switchANALYSIS | Dynamic Biosensors | https://www.dynamic- biosensors. com/software/ | |
| Software, algorithm | ImageQuant | GE Healthcare | TL 5.2 | |
| Other | Insect-XPRESS protein-free insect cell medium withL-glutamine | Lonza | 12-730Q | |
| Other | Protease Inhibitor Cocktail | Sigma-Aldrich | 11836170001 | |
| Other | Desthiobiotin | IBA | 2-1000-001 | |
| Other | Imidazole | Sigma-Aldrich | I5513 | |
| Other | Formamide | Sigma-Aldrich | 11814320001 | |
| Other | TEMED | Sigma-Aldrich | T9281 | |
| Other | Ammonium persulfate (APS) | Sigma-Aldrich | A3678 | |
| Other | SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| Other | SYBR Green II RNA Gel Stain | Thermo Fisher Scientific | S7586 | |
| Other | Ni-NTA Agarose | QIAGEN | 30210 | |
| Other | Glutathione Sepharose 4B | GE Healthcare | 17075601 | |
| Other | Amicon Ultra Centrifugal Filter Units | Millipore | UFC901096 | |
| Other | TWEEN 20 | Sigma-Aldrich | P9416 |
Table 1
Sequences of RNAs used in this study.
https://doi.org/10.7554/eLife.40670.015| Name | Sequence | Experiment |
|---|---|---|
| Zfs1 target A30 | AAUCAUCCUUAUUUAUUACCAUU[A]30 | Deadenylation |
| Puf3 target A30 / Puf3 target (5A) A30 | AACUGUUCCUGUAAAUACGCCAG[A]30 | Deadenylation |
| Puf3 target (5U) A30 | AACUGUUCCUGUAUAUACGCCAG[A]30 | Deadenylation |
| Puf5 target | AACUGUUGUUGUAUGUACGCCAG[A]30 | Deadenylation |
| ARE/Zfs1 target | AUUAUUUAUU | Fluorescence polarization |
| PRE/Puf3 target (5A) | ACCUGUAAAUA | Fluorescence polarization |
| Puf3 target (5U) | ACCUGUAUAUA | Fluorescence polarization |
| Puf5 target | AGUUGUAUGUA | Fluorescence polarization |
| A30 | [A]30 | Fluorescence polarization |
| Puf3 target (5A) | ACCUGUAAAUAGGCG-[*DNA] | SwitchSENSE |
| Puf3 target (5U) | ACCUGUAUAUAGGCG-[*DNA] | SwitchSENSE |
| Puf5 target | AGUUGUAUGUAGGCG-[*DNA] | SwitchSENSE |
| Poly(A) | [A]15-[*DNA] | SwitchSENSE |
-
*DNA for hybridization to nanolever:
ATCAGCGTTCGATGCTTCCGACTAATCAGCCATATCAGCTTACGACTA
Additional files
-
Supplementary file 1
Analysis of Puf3-motif quality in putative mRNA targets of Puf3.
This table includes the S. cerevisiae mRNAs that are candidates for Puf3 regulation, obtained from published sources as detailed in Materials and methods. The highest quality motif present in each mRNA is indicated with the category numbers defined in Figure 6B (1–4), and 5 for Δcore. Functional categories were defined in this study based on published data obtained from the SGD database.
- https://doi.org/10.7554/eLife.40670.016
-
Supplementary file 2
Gene Ontology analysis of mRNA groups defined by Puf3-motif quality and location.
Statistically significant gene ontology over-representation was identified with PANTHER as detailed in Materials and methods. Terms were identified for the mRNA groups defined by the presence of motifs 1, 2 or 3 within the 3' UTR (see Figutre 6B for motif quality definitions). No significantly enriched GO terms were identified for motif 4 in the 3' UTR or for any groups with motifs in the coding sequence. Significantly enriched GO terms for the mRNAs defined by a mismatch within the seven core nucleotides (Δcore) were identified and are shown here.
- https://doi.org/10.7554/eLife.40670.017
-
Supplementary file 3
Plasmids and Oligonucleotides.
Plasmids and oligos used in this study are listed.
- https://doi.org/10.7554/eLife.40670.018
-
Transparent reporting form
- https://doi.org/10.7554/eLife.40670.019
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
RNA-binding proteins distinguish between similar sequence motifs to promote targeted deadenylation by Ccr4-Not
eLife 8:e40670.
https://doi.org/10.7554/eLife.40670




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}