Reporter-ChIP-nexus reveals strong contribution of the Drosophila initiator sequence to RNA polymerase pausing
- Stowers Institute for Medical Research, United States
- University of Kansas Medical Center, United States
Figures
Figure 1 with 2 supplements

Reporter-ChIP-nexus captures paused Pol II.
(A) Reporter-ChIP-nexus is performed by cloning Drosophila pseudoobscura promoters or synthetic promoters into a simple GFP reporter and transfecting into D. melanogaster Kc167 cells. The whole cell lysate from cross-linked cells is used to perform Pol II ChIP-nexus. Exonuclease stop bases are then mapped and shown on the positive strand in red above the line, while reads from the negative end are shown in blue below the line. (B) Results of reporter-ChIP-nexus reveal strong Pol II pausing at the synthetic super core promoter (SCP), which contains the core promoter elements TATA, Inr, MTE, DPE and PB (top). The position of transcriptional initiation is mapped by sequencing the 5’ end of the produced RNA (bottom). The results for the SCP promoter show that the vast majority of RNAs start at the expected site of initiation.
Figure 1—figure supplement 1
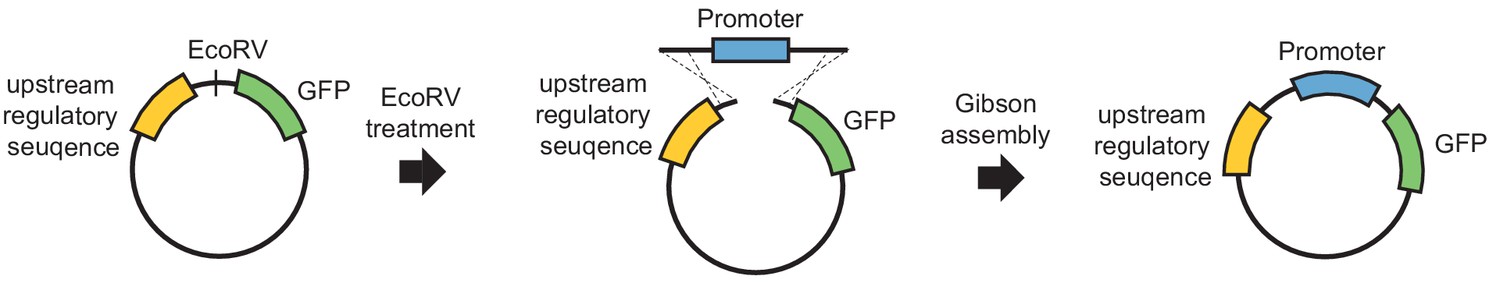
A simple GFP reporter that allows the fast insertion of any promoter sequence.
To perform reporter-ChIP-nexus, we constructed a simple GFP reporter plasmid with upstream regulatory sequences. An EcoRV cutting site was used to linearize the plasmid and insert a promoter of interest using Gibson assembly (see Materials and methods for more information).
Figure 1—figure supplement 2

Workflow for gene-specific 5’ RNA sequencing, a method similar to RNA amplification of cDNA ends (RACE).
RNA transcribed form the reporter was reverse transcribed using a primer that has 3’ sequences complementary to the GFP sequence. The 5’ of this primer contains a BamHI restriction cutting site and a partial reverse TruSeq P5 adapter sequence (purple). When the reverse transcriptase reaction is complete (usually stopping at the cap), each generated cDNA contains the TruSeq P5 adapter sequence. After purification, the single-stranded cDNA is circularized to bring the TruSeq P5 adapter sequence to the cDNA 3’ end, where the reverse transcriptase stops. The product is then linearized again by digesting with BamHI (scissors) after making the restriction enzyme cut site double-stranded by annealing it to a complementary oligos (red). The resulting DNA is then amplified in a PCR with a primer that contains the full-length P5 sequence (matching the partial P5 sequence), as well as a primer that primes on the GFP sequence (black) and has the full-length P7 adapter sequence at the 5’ end (yellow). The library is then sequenced with a P5 primer and after alignment to the plasmid, the first base is recorded. These bases correspond to the site where the reverse transcriptase stopped transcribing, which should occur at the 5’ end of the RNA, where the cap is found.
Figure 2 with 4 supplements
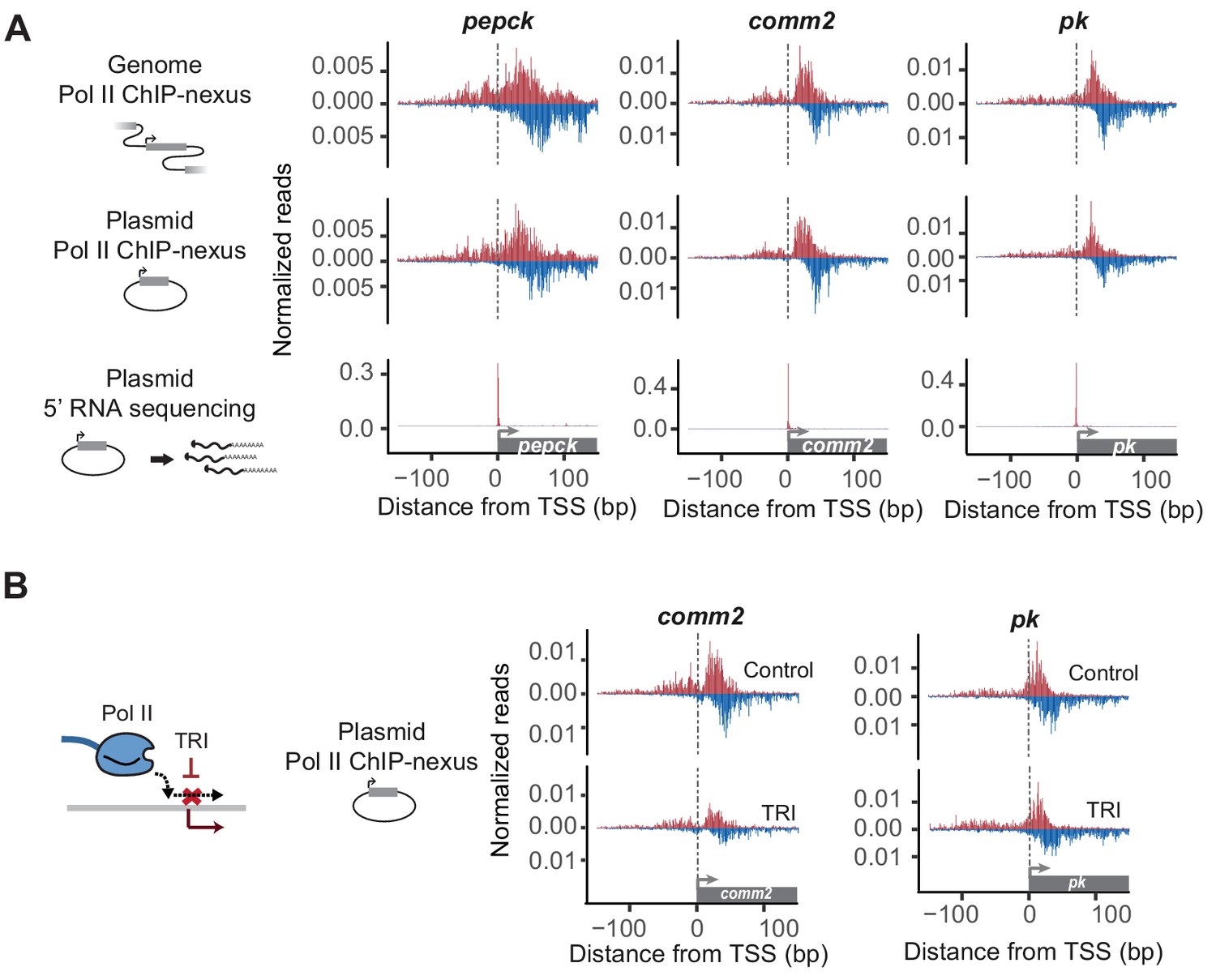
Reporter-ChIP-nexus recapitulates the endogenous Pol II pausing profile.
(A) To obtain the endogenous Pol II pattern of D. pseudoobscura promoters in the genome, ChIP-nexus was performed in a D. pseudoobscura cell line. Results are shown for the pepck, comm2 and pk promoters. The same promoters were then examined using reporter-ChIP-nexus in D. melanogaster Kc167 cells, which yielded patterns very similar to the endogenous Pol II profiles. The transcription initiation sites were confirmed by 5’ RNA sequencing. (B) To determine the stability of paused Pol II on the plasmid, transfected cells were treated with either DMSO (Control) or triptolide (TRI) for 1 hr. The results show a relative reduction in Pol II similar to that of the endogenous loci after treating the D. pseudoobscura cell line with TRI (see Figure 2—figure supplement 4). Therefore, reporter-ChIP-nexus reveals gene-specific Pol II pausing stability on a plasmid.
Figure 2—figure supplement 1
Paused Pol II stability measurements at eight D. pseudoobscura promoters.
Pol II ChIP-nexus was performed on the D. pseudoobscura cell line ML83-63. The stability of paused Pol II was analyzed by comparing the Pol II profile under control and triptolide (TRI) treated conditions. The eight D. pseudoobscura promoters shown in this figure have high Pol II signal and various degrees of Pol II pausing. The sequences of these promoters were cloned into our reporter and used for later mutagenesis experiments.
Figure 2—figure supplement 2

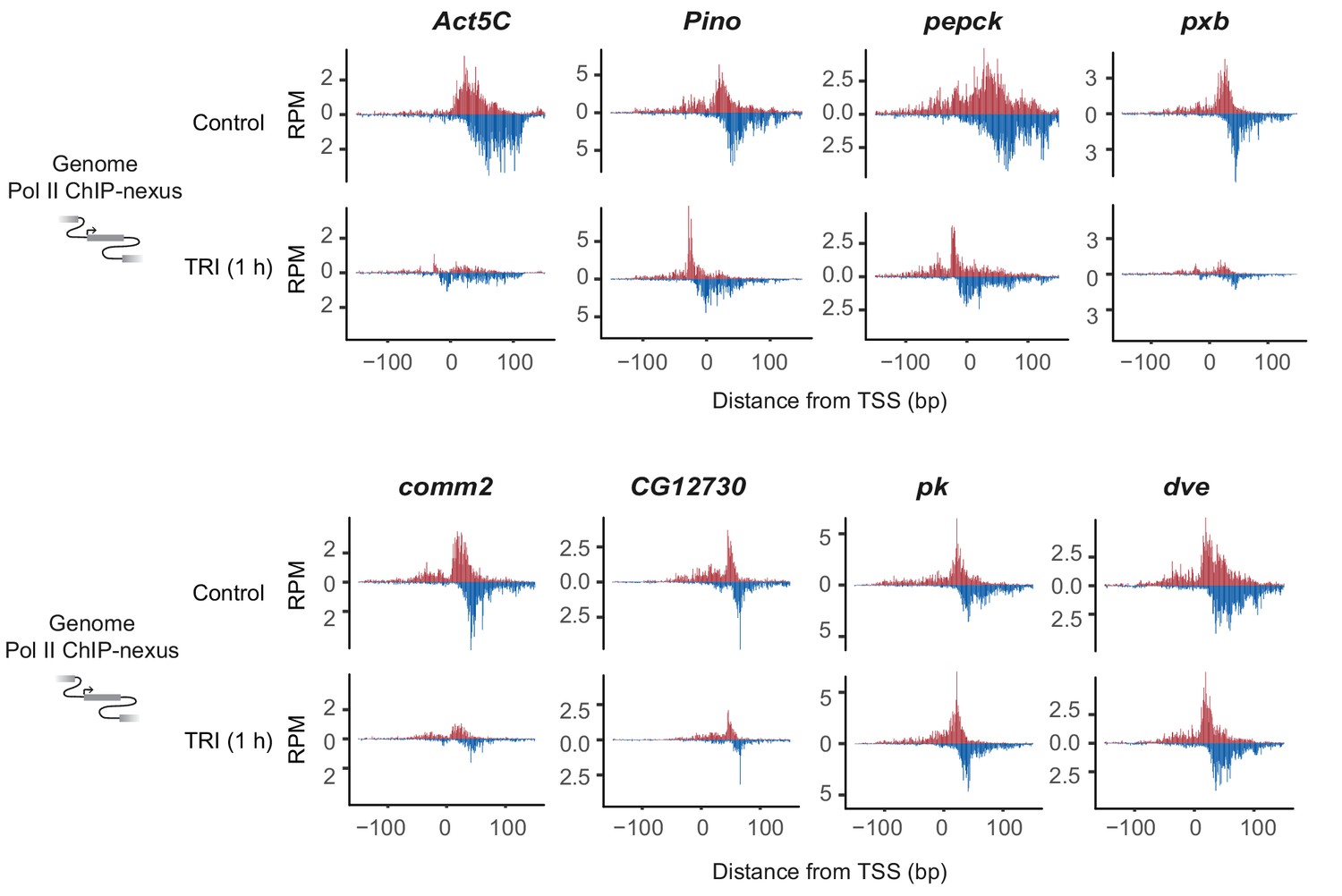
Reporter-ChIP-nexus recapitulates endogenous Pol II pausing.
The ChIP-nexus profile of endogenous D. pseudoobscura promoters (first row) strongly resembles the profile detected by reporter-ChIP-nexus after cloning the D. pseudoobscura promoters into the reporter and transfection of the plasmid into Kc167 cells (second row). The artificial super core promoter (SCP) also produces a strong Pol II pausing profile. The transcription start site on the reporter was mapped for each promoter using gene-specific 5’ RNA sequencing (third row) and was found to map to the expected location.
Figure 2—figure supplement 3
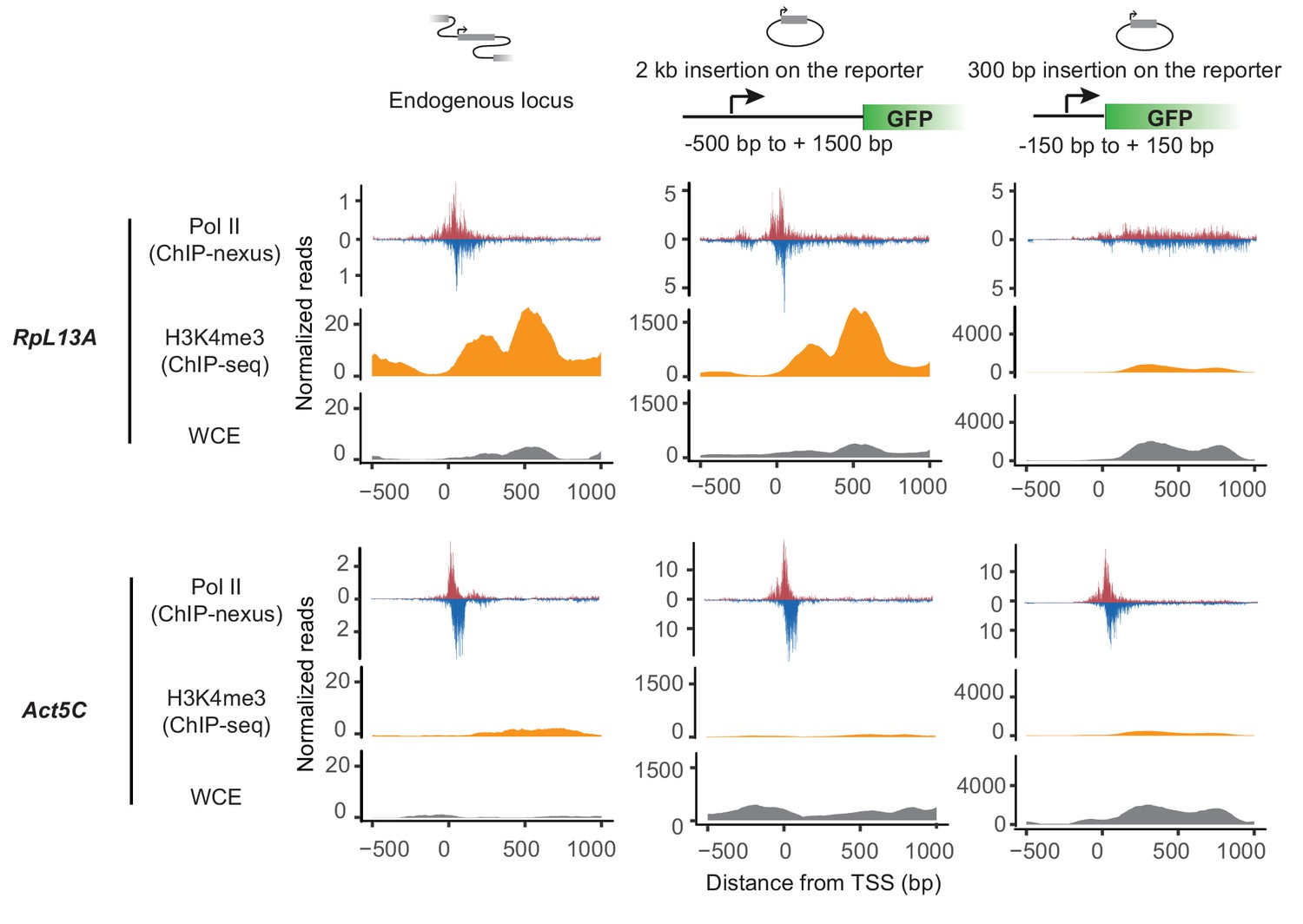
Larger promoter region insertion is required for recapitulating Pol II pausing at RpL13A on the plasmid.
Pol II ChIP-nexus and H3K4me3 ChIP-seq profiles at D. pseudoobscura RpL13A and Act5C endogenous loci and that on the plasmids with 300 bp or 2 kb promoter region insertion. Both RpL13A and Act5C show strong pausing profile at the endogenous loci. However, in contrast to Act5C, where Pol II pausing can be observed on the plasmid with both 300 bp and 2 kb promoter region insertion, RpL13A pausing profile can only be recapitulated on the plasmid when the larger 2 kb region was inserted. In addition, although Act5C show minimal enrichment of H3K4me3, high H3K4me3 signal can be observed at RpL13A endogenous locus, and this H3K4me3 signal was only recapitulated on the plasmid with the 2 kb promoter region insertion. This correlation between Pol II profile and H3K4me3 enrichment suggests that endogenous chromoatin context is important for establishing paused Pol II at Rpl13A.
Figure 2—figure supplement 4

Reporter-ChIP-nexus recapitulates gene-specific Pol II pausing stability.
The stability of paused Pol II was examined on the plasmids and at the endogenous loci by treatment with triptolide (TRI). After TRI treatment, paused Pol II is lost, and the degree of persistence relative to the DMSO-treated control condition is a measurement for the stability of paused Pol II. The examples of the comm2 and pk promoters show that the relative loss of Pol II on the plasmid is similar to that at the endogenous loci.
Figure 3 with 1 supplement
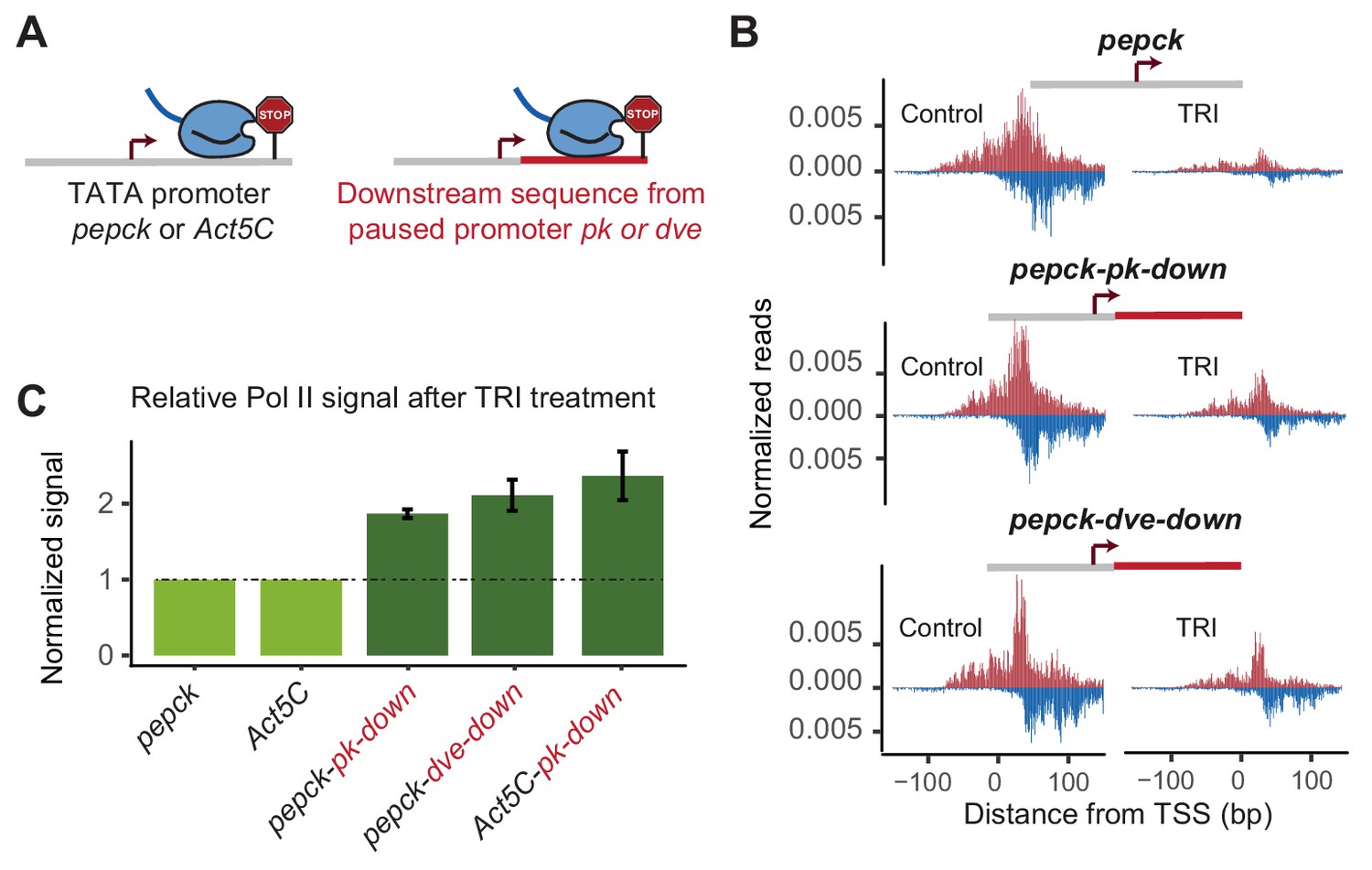
Changes in downstream promoter sequences alter paused Pol II stability.
(A) The pepck and Act5C downstream sequences were replaced with that from the stably paused promoter pk or dve (fusion site: 8 bp after the TSS). (B) Pol II ChIP-nexus data on the plasmids after transfection into Kc167 cells with or without treatment with triptolide (TRI) for 5 min. The wild-type pepck promoter shows a strong reduction of paused Pol II. At the pepck-pk-down and pepck-dve-down fusion promoters, the same TRI treatment did not reduce paused Pol II to the same extent as at the wild-type promoter, suggesting an increase in the paused Pol II stability as a result of changing the downstream promoter sequence. (C) To quantify the difference in paused Pol II stability between different constructs, the ratio of paused Pol II before and after TRI treatment is calculated using two biological replicates. The ratio for the wild-type promoter is then normalized to 1 (light green), and the relative change in this ratio for the fusion promoter is shown on the right (dark green). Error bars refer to the standard error of the mean (TRI treatment: 5 min for all promoters).
Figure 3—figure supplement 1
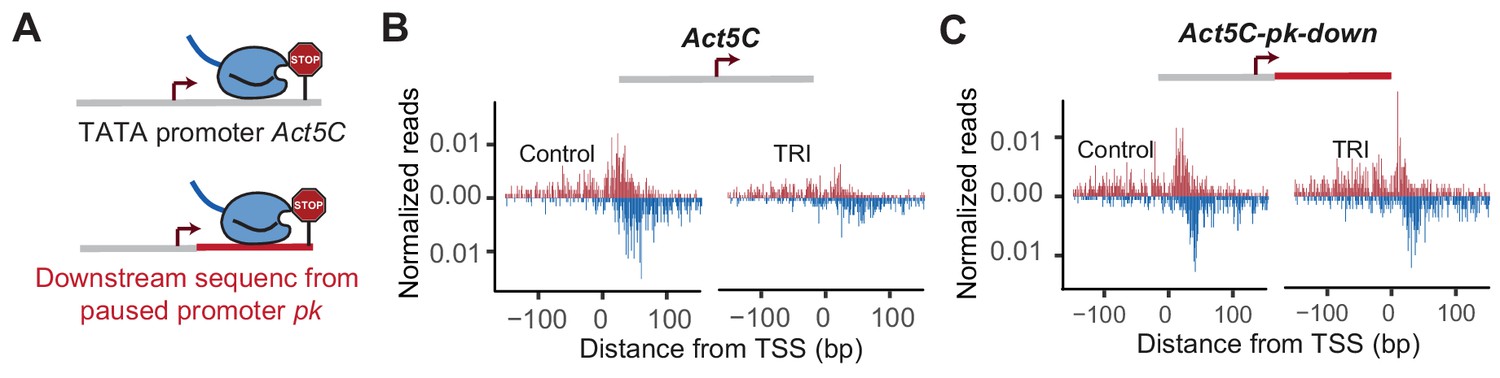
Downstream promoter sequences influences Pol II pausing at Act5C.
Pol II ChIP-nexus profile at Act5C and Act5C-pk-down fusion promoters under control condition and after 5 min Triptolide (TRI) treatment.
Figure 4 with 1 supplement
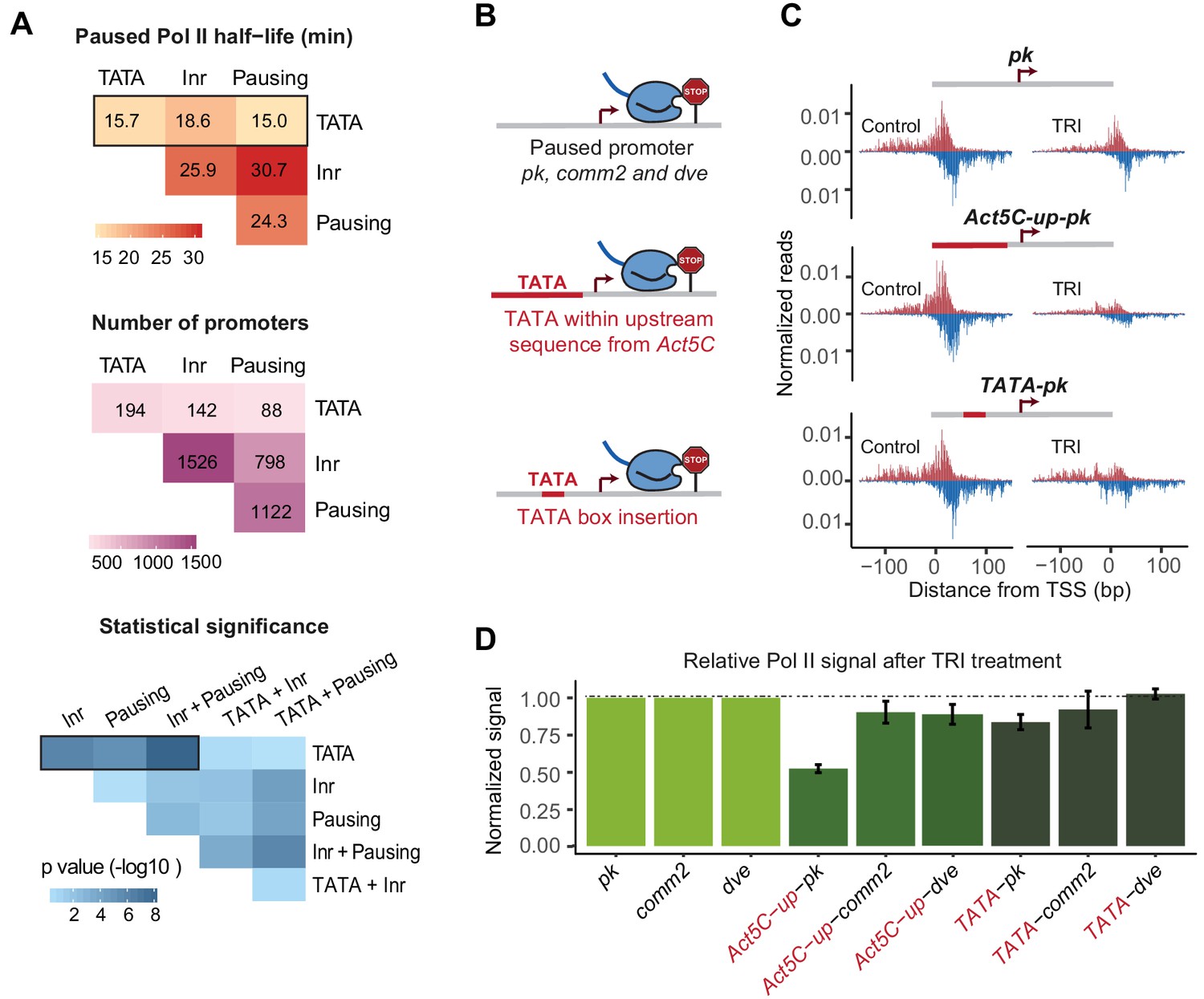
A TATA insertion may reduce Pol II pausing.
(A) Analysis of paused Pol II half-lives as a function of various core promoter element combinations. Median paused Pol II half-life of promoters with different combinations of core promoter elements (top) and the number of promoters classified (middle) are shown. A Wilcoxon rank sum test was then used to test whether the various promoter combinations have significantly different Pol II half-lives from each other (bottom). Core promoter elements other than those used for classification were allowed to occur in each group (e.g. Inr and pausing elements can occur at the 194 promoters with TATA box). Similar results were obtained when no promoter element other than those used for classification were allowed (mutually exclusive model Figure 4—figure supplement 1). (B) Experimental strategy to functionally test the role of TATA in Pol II pausing: the upstream promoter sequences of the paused promoter pk, comm2 and dve were replaced with that of the TATA promoter Act5C (fusion site: 7 bp before the TSS), or a canonical TATA box sequence (TATAAAA) was inserted at 31 bp upstream of the TSS. (C) The Pol II ChIP-nexus profiles for wild-type pk, the upstream region replacement (Act5C-up-pk) and the TATA insertion (TATA-pk) under control conditions (left) and after 1 hr treatment with triptolide (TRI, right) show that Pol II pausing is reduced after the pk promoter acquired a TATA box. (D) Quantification of the relative changes in paused Pol II stability for all fusion promoters relative to the wild-type promoters (TRI treatment: 1 hr for pk and dve based promoters, 40 min for comm2 based promoters). Note that only the pk promoter shows a clear reduction in Pol II pausing after insertion of a TATA box.
Figure 4—figure supplement 1
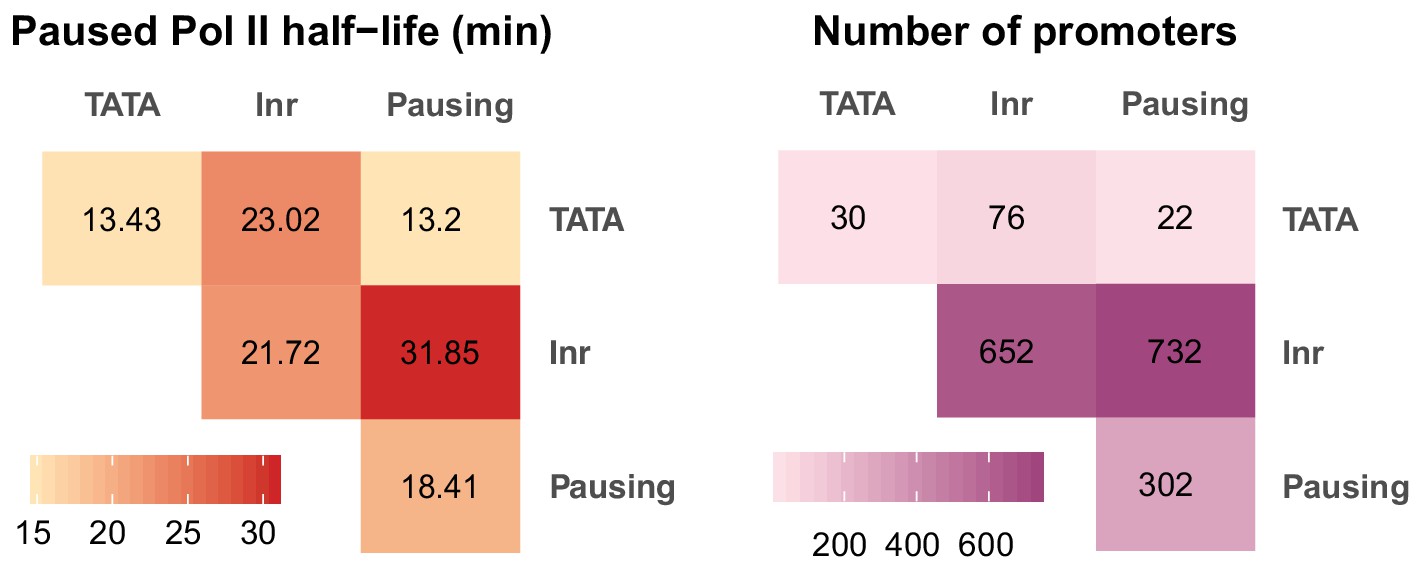
Correlation between paused Pol II half-life and core promoter elements using a mutually exclusive model.
To better understand the relationship between core promoter elements and the half-live of paused Pol II genome-wide, we calculated the median half-life for each element and its possible combination with other elements with a mutually exclusive model (See Materials and method for details). Left: median paused Pol II half-life. Right: number of promoters in each category.
Figure 5 with 1 supplement
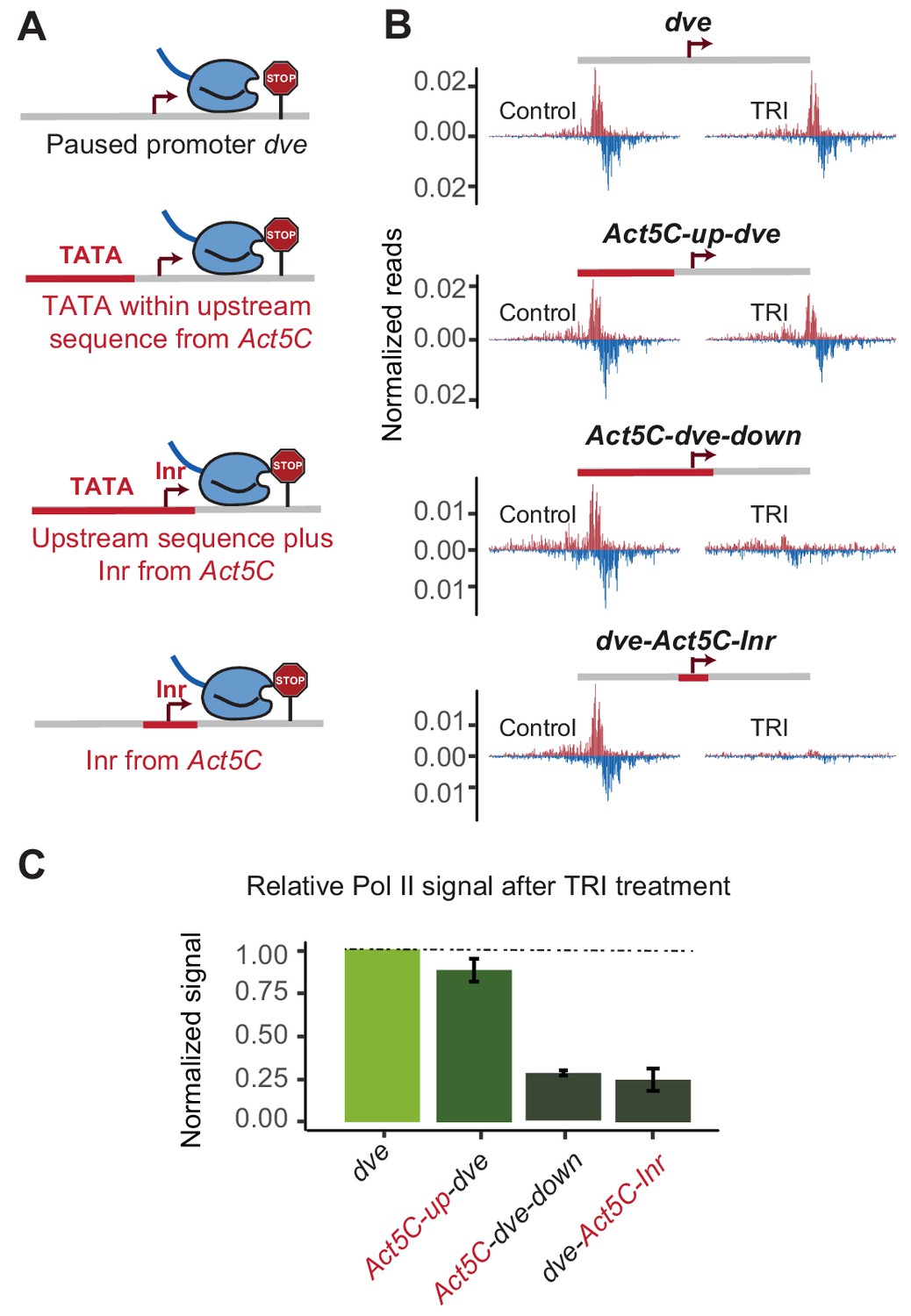
The initiator sequence plays an important role in Pol II pausing.
(A) Experimental series on the dve promoter testing the role of the Inr in conjunction with a TATA-containing region. Either only the upstream region was replaced with that of the Act5C promoter (fusion site 7 bp before the TSS), the upstream region and the Inr were replaced (fusion site 8 bp after the TSS), or only the Inr was replaced (16 bp region around the TSS). (B) Pol II ChIP-nexus profiles under control conditions and after treatment with triptolide (TRI) for 1 hr show a strong reduction of paused Pol II after replacing the Inr sequence. (C) Quantification of the relative changes in paused Pol II stability for all fusion promoters relative to the wild-type promoter (TRI treatment: 1 hr for all the promoters).
Figure 5—figure supplement 1
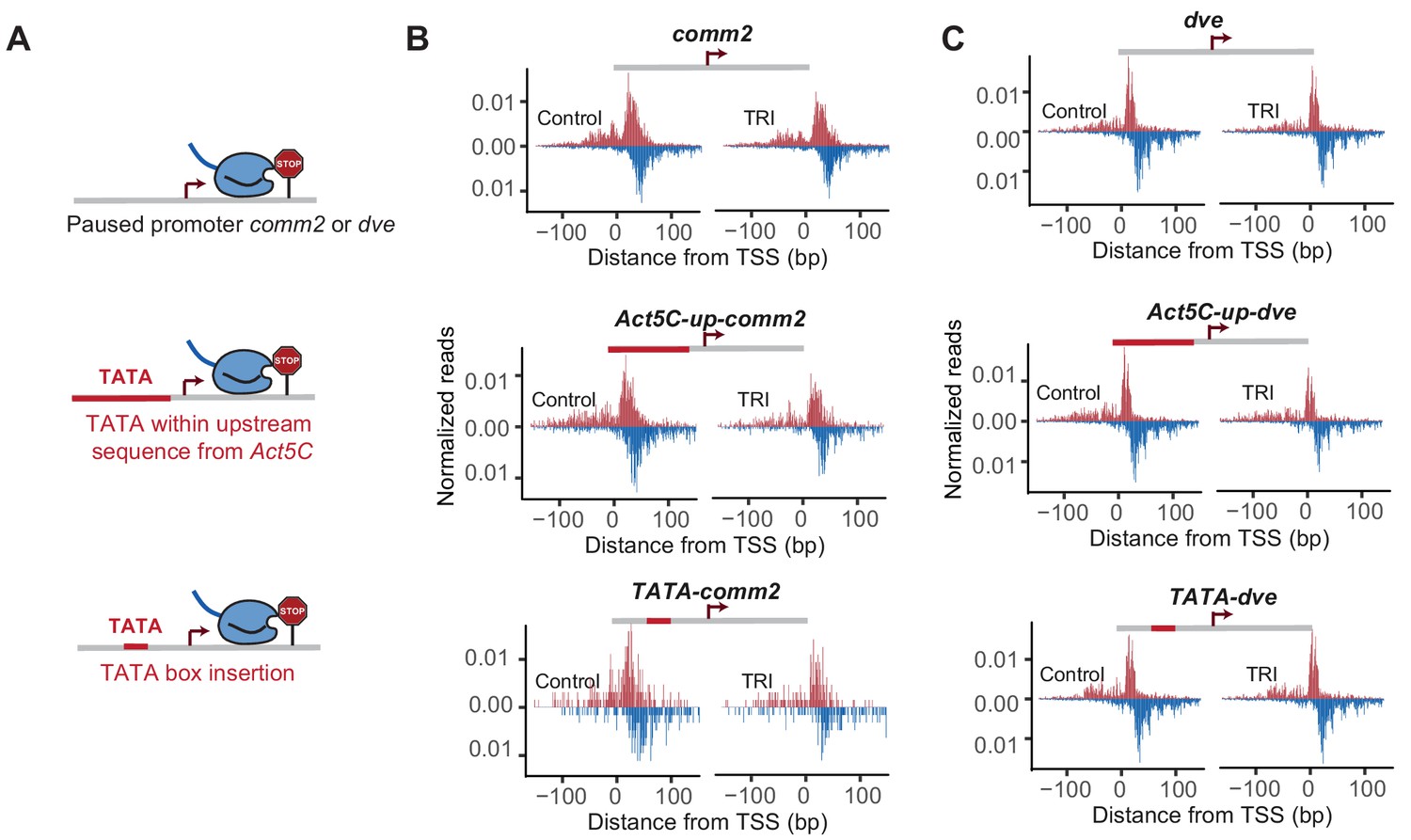
Effect of TATA insertion at comm2 and dve.
Pol II ChIP-nexus profile at comm2, Act5C-up-comm2, TATA-comm2, dve, Act5C-up-dve and TATA-dve promoters under control condition and after Triptolide (TRI) treatment (40 min for comm2 and 1 hr for dve).
Figure 6
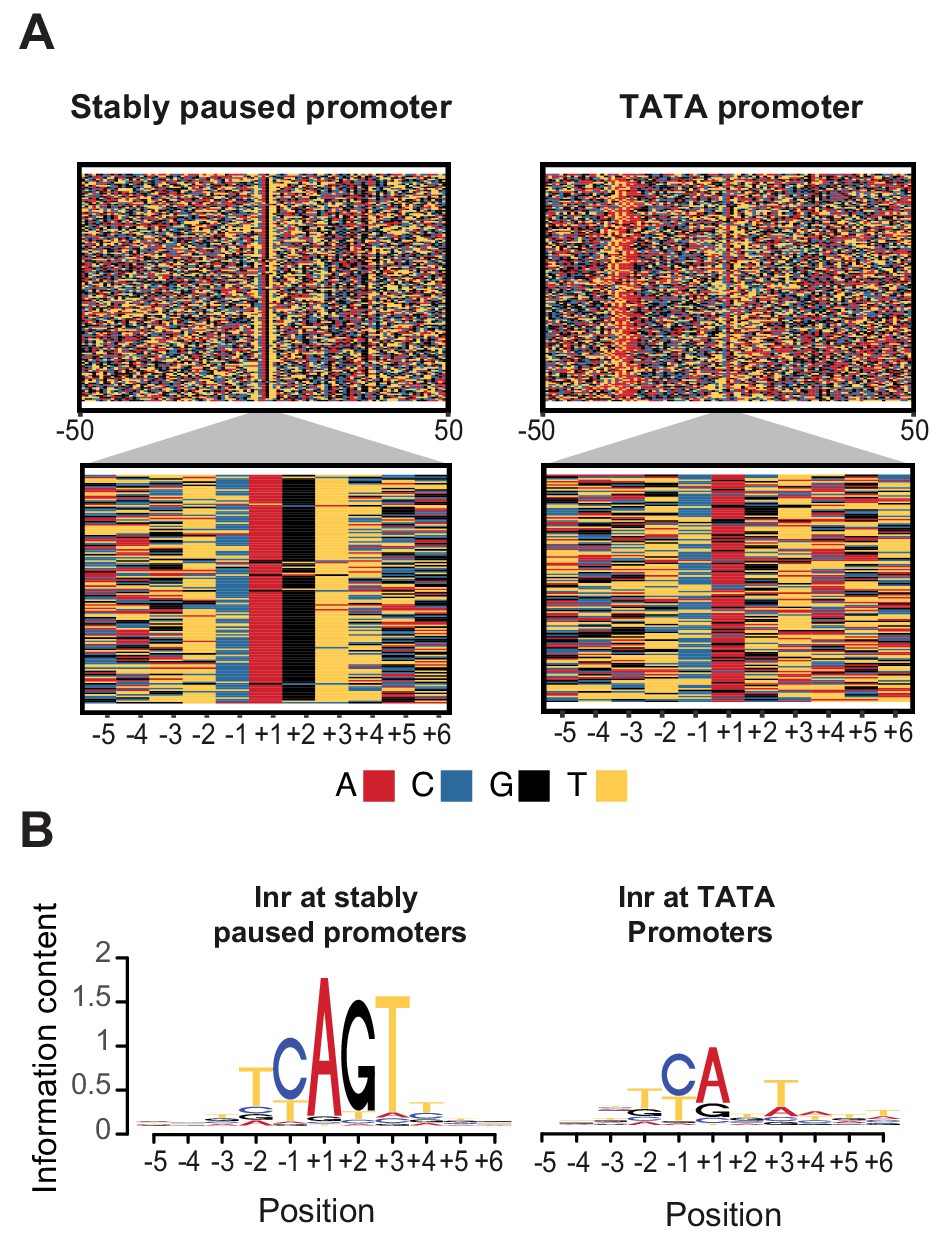
Initiator sequences differ between TATA and stably paused promoters.
(A) The sequence of stably paused promoters (132 randomly selected from the 490 promoters that did not have a TATA box and had a paused Pol II half-life longer than 60 min) are shown on the left as colored letters for a 100 bp window centered on the transcription start site (top). TATA promoters (132 promoters with paused Pol II half-lives shorter than 30 min) are shown on the right as comparison. A higher magnification of the initiator (Inr) sequences below shows clear differences between the two promoter types. (B) The consensus motif for the Inr sequences of stably paused promoters match more closely the canonical Inr consensus sequence (TCAKTY) and preferentially contains a G at the +2 position in most cases, whereas the Inr sequences of TATA promoters are more degenerate without enrichment of a G at the +2 position.
Figure 7 with 3 supplements
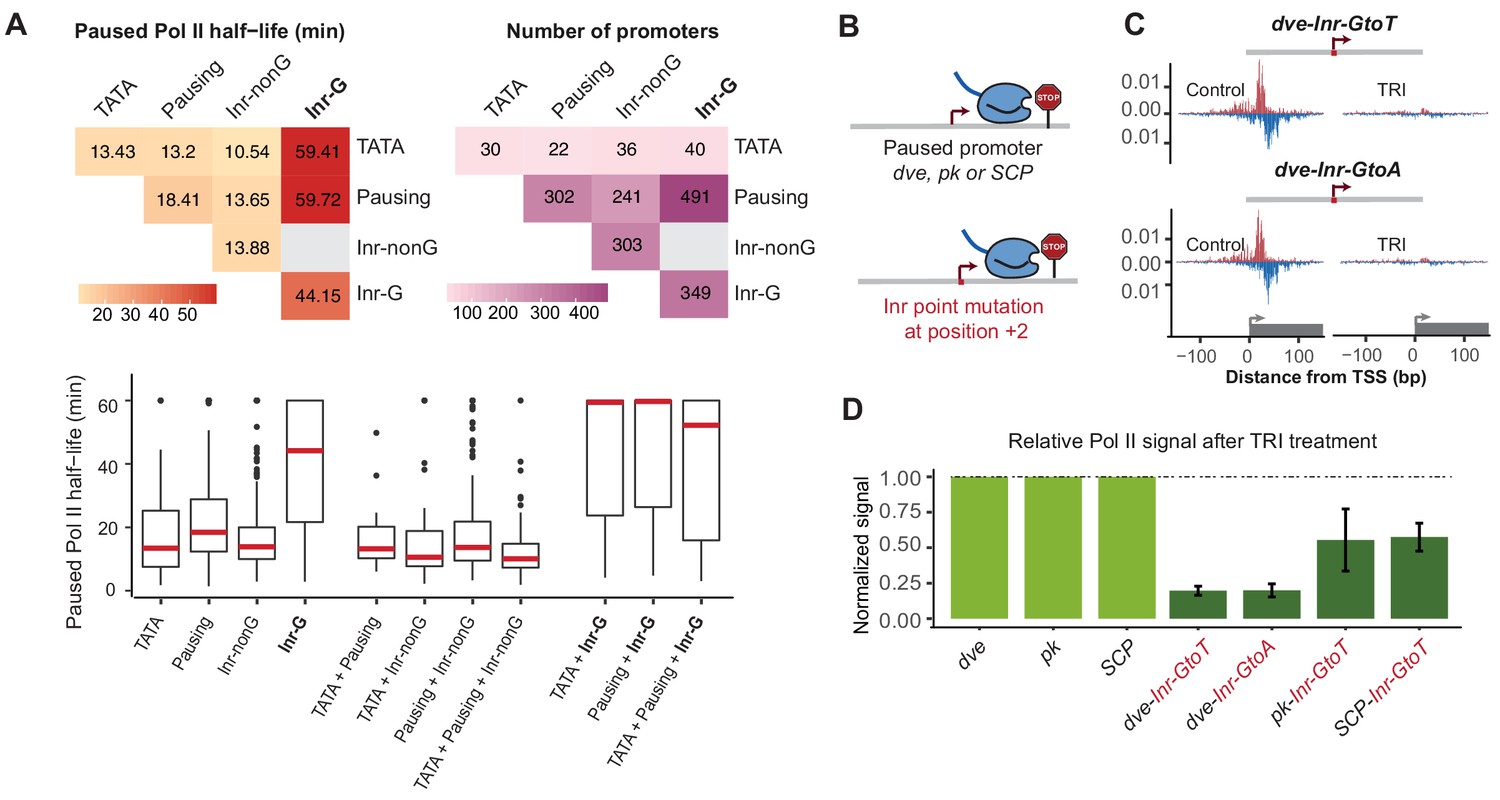
The G at Inr + 2 position is critical for stable Pol II pausing.
(A) Analysis of paused Pol II half-lives as a function of core promoter element combinations after separating the Inr sequences into those that contain a G at position +2 (Inr-G) versus those that do not contain a G at this position (Inr-nonG). Median paused Pol II half-life (left), promoter numbers (right) and boxplot of their distribution (bottom, median in red) are shown for promoters with different combinations of core promoter elements. The promoters only contain the indicated promoter element, excluding any other promoter element (e.g. the 491 promoters with pausing elements and Inr-G variant do not contain TATA box). Similar results were obtained using a non-mutually exclusive model (Figure 7—figure supplement 1). Note that strong Pol II pausing was observed in all combinations that contain the Inr-G variant. Results from testing between combinations using the Wilcoxon rank test are shown in Table S4. (B) Experimental strategy to test the effect of mutating the G at Inr + 2 position of the Inr to T or A at the stably paused promoters dve, pk and the synthetic Super Core Promoter (SCP). (C) Pol II ChIP-nexus profiles after treatment with triptolide (TRI) for 1 hr show a strong reduction in Pol II pausing after the G was mutated at the dve promoter. (D) Quantification of the relative changes in paused Pol II stability for all fusion promoters relative to the corresponding wild-type promoter. Error bars are from replicate experiments. The duration of TRI treatment was 1 hr for pk and dve derived promoters, 30 min for SCP derived promoters.
Figure 7—figure supplement 1
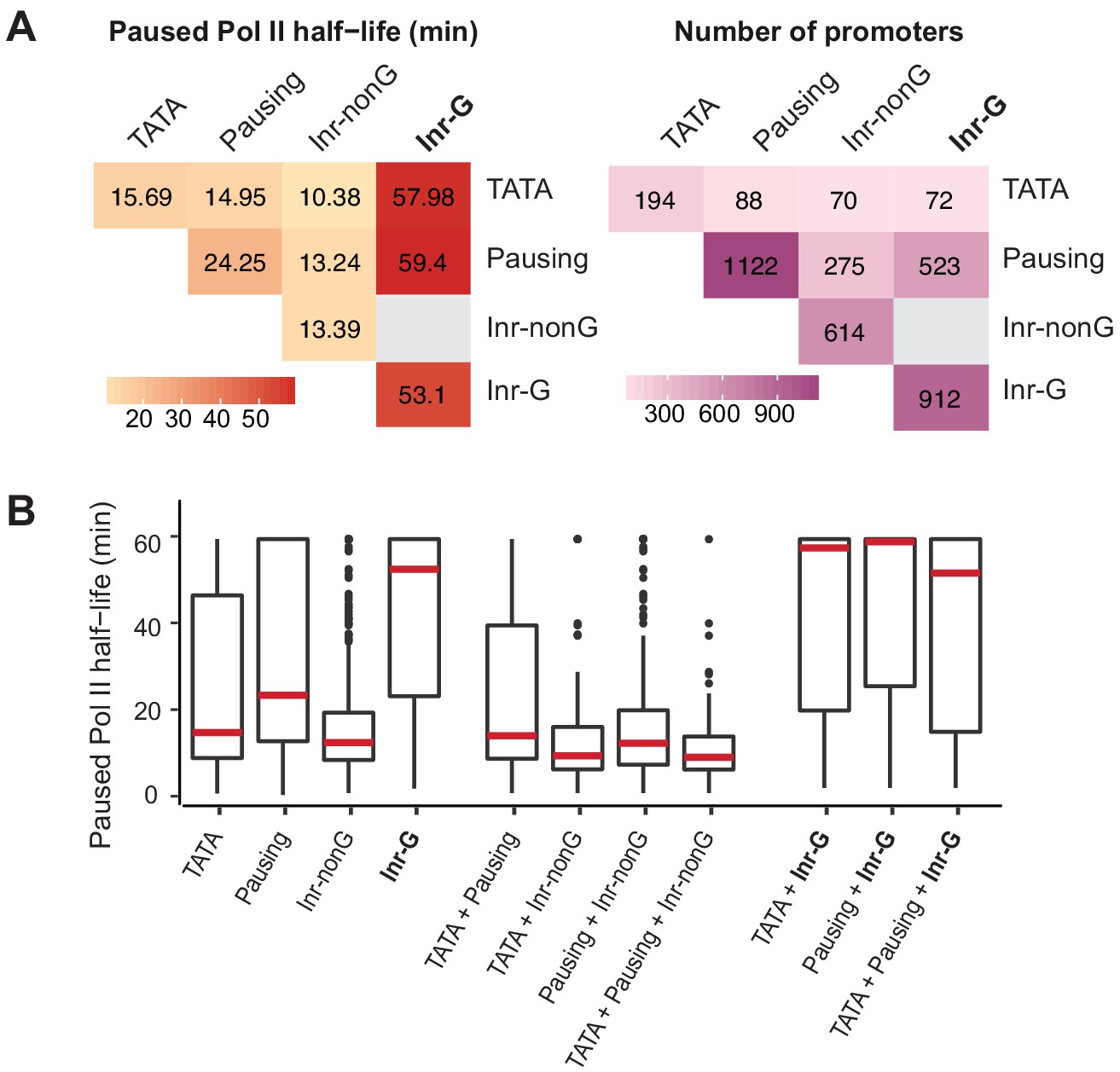
Correlation between paused Pol II half-life and Inr variants using a non-mutually exclusive model.
(A) Median paused Pol II half-life (left) and promoter numbers (right) at promoters with different combinations of core promoter elements using a non-mutually exclusive model. (B) Paused Pol II half-life at promoters with different combinations of core promoter elements using a non-mutually exclusive model.
Figure 7—figure supplement 2
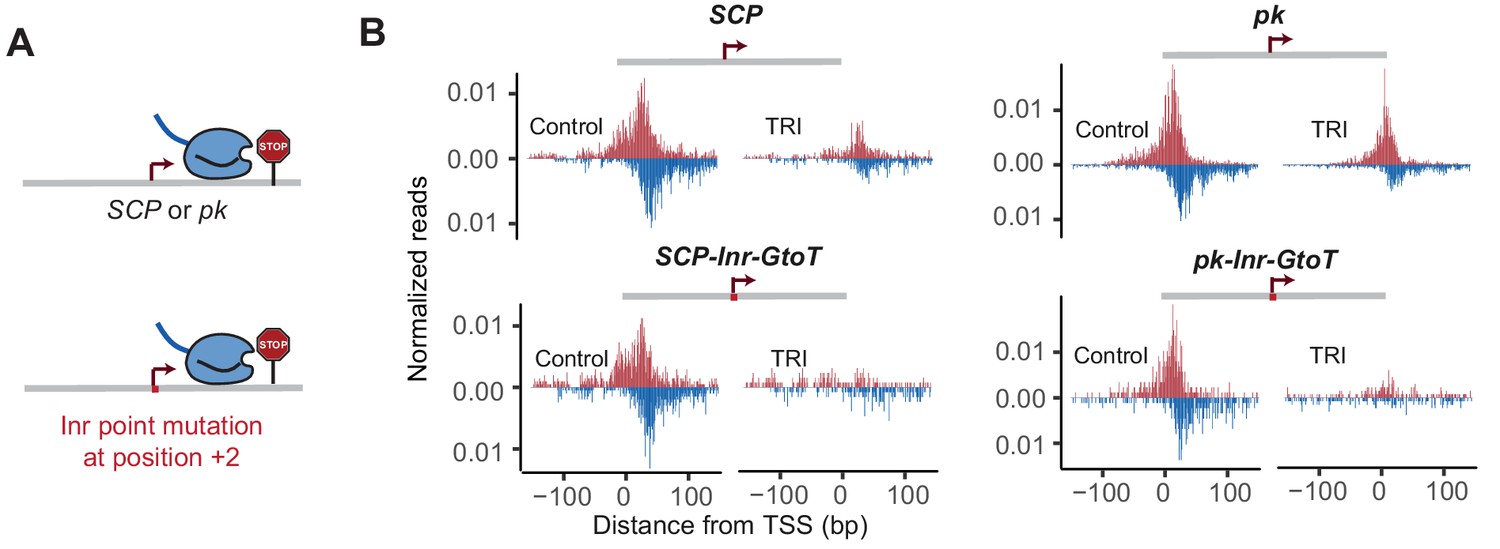
Mutating the G at Inr + 2 position reduces Pol II pausing at SCP and pk.
Pol II ChIP-nexus profile at SCP, SCP-Inr-GtoT, pk and pk-Inr-GtoT fusion promoters under control condition and Triptolide (TRI) treatment (30 min for SCP, 1 hr for pk).
Figure 7—figure supplement 3
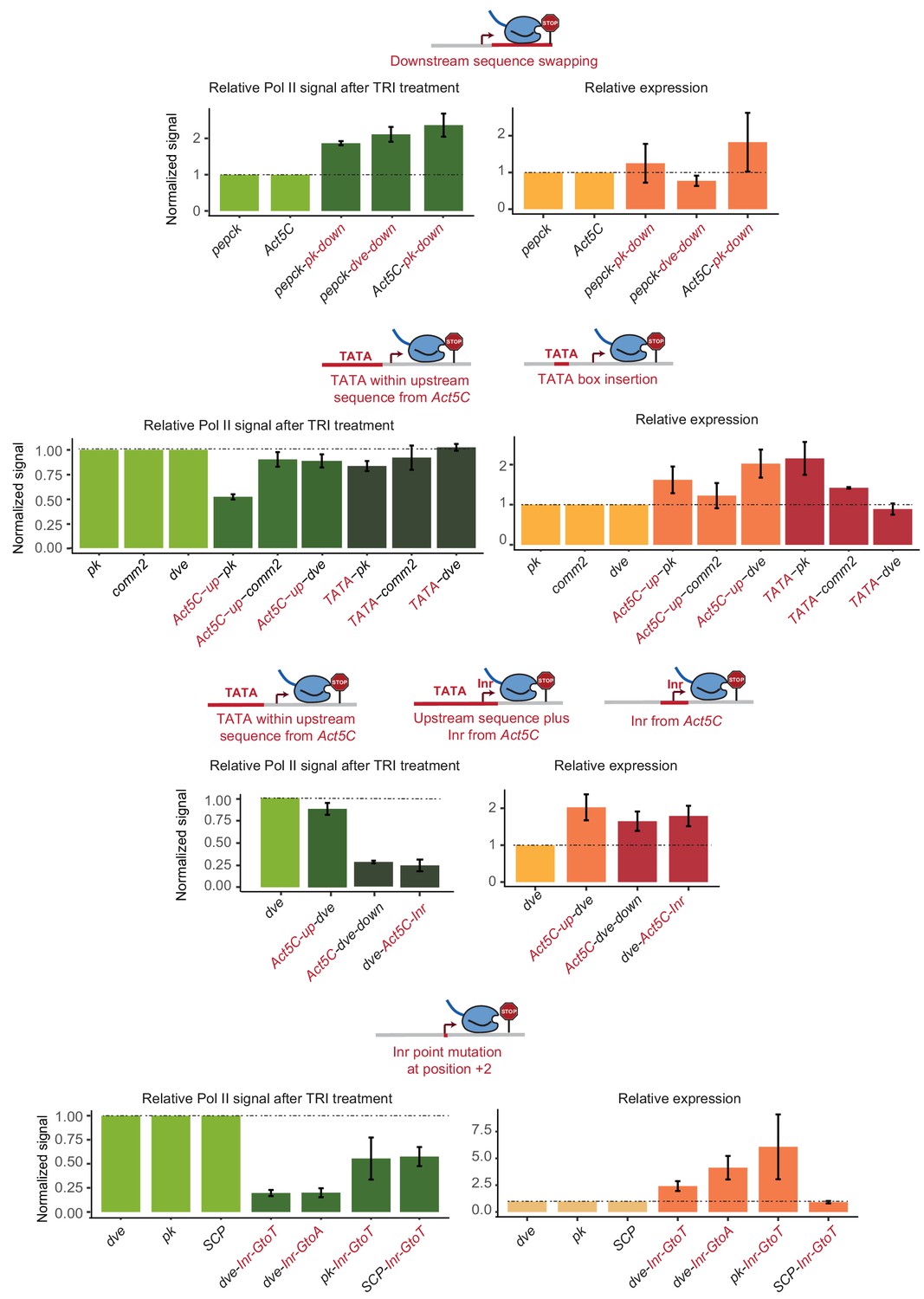
Transcript level changes after altering the promoter sequence.
To study the effect of altered promoter sequences on RNA expression, we performed RT-qPCR for all experiments. The relative Pol II pausing stability (green) and relative RNA expression levels (yellow) are shown. Measurements for the wild-type promoter are normalized to 1, and the relative values for the mutated promoters are shown next to it. Errors bars refer to standard error among two to three biological replicates. Note that the error is large due the variability in transfection efficiency between independent samples. Despite the variability, the change in Pol II pausing stability was in most cases inversely correlated with that of RNA transcription. Thus, a loss in Pol II pausing was usually associated with an increase in transcription. This is consistent with Pol II pausing having an inhibitory effect on transcription. We cannot exclude the possibility that the causality is reverse though, that is that an increase in promoter strength leads to a reduction in Pol II pausing. It is unclear, however, whether promoter strength can be uncoupled from Pol II pausing, and we cannot measure them separately. In our model, promoter strength and Pol II pausing are both determined by TFIID, which binds to both the upstream and downstream core promoter elements we studied.
Figure 8
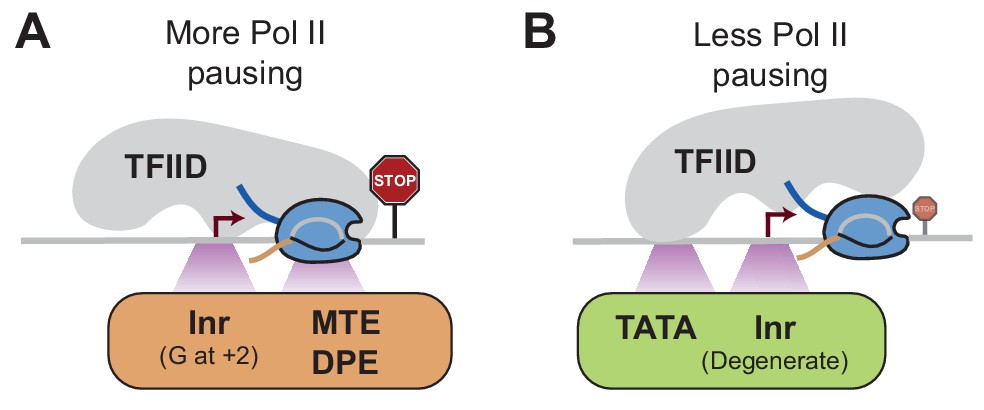
Model of how promoter sequences may influence Pol II pausing through TFIID Since the core promoter elements that we analyzed are bound by TFIID, we hypothesize that TFIID affects Pol II pausing dependent on the position it binds to the promoter.
(A)At stably paused promoters, pausing elements and consensus Inr sequences with a G at +2 position promote TFIID-downstream DNA interactions, which prolong Pol II pausing. (B)At TATA-containing promoters, the TATA box and a degenerate Inr sequence favor TFIID interactions with upstream DNA, which promote Pol II pause release.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Antibody | Rabbit polyclonal anti-Rpb3 | Julia Zeitlinger Lab | Zeitlinger Lab #163185–50 | 10 ug |
| Antibody | Rabbit polyclonal anti-H3K4me3 | Cell Signaling | #9727 | 10 ug |
| Strain, strain background (E. coli) | NEB 5-alpha Competent E. coli (High Efficiency) | New England Biolabs | #C2987H | |
| Strain, strain background (E. coli) | One Shot ccdB Survival 2 T1R Competent Cells | ThermoFisher Scientific | #A10460 | |
| Chemical | Chloroform | Sigma-Aldrich | #C2432 | |
| Commercial assay or kit | CircLigase ssDNA Ligase | Illumina (Epicentre) | #CL4115K | |
| Chemical compound | DMSO | Sigma-Aldrich | #276855 | |
| Chemical compound | Dynabeads Protein A | Life Technologies | #100-08D | |
| Chemical compound | Dynabeads Protein G | Life Technologies | #100-04D | |
| Commercial assay or kit | FastDigest BamHI | ThermoFisher Scientific | #FERFD0054 | |
| Chemical compound | FuGENE HD reagent | Promega | #E2311 | |
| Chemical compound | HyClone SFX-Insect Cell Culture Media | ThermoFisher Scientific | #SH3027802PM | |
| Commercial assay or kit | Klenow Fragment (3’ to 5’ exo-) | New England Biolabs | #M0212S | |
| Commercial assay or kit | Lambda Exonuclease | New England Biolabs | #M0262L | |
| Chemical compound | Opti-MEM Reduced Serum Medium | ThermoFisher Scientific | #31985062 | |
| Chemical compound | Phenol: Chloroform: iso-Amyl alcohol | VWR | #97064–824 | |
| Chemical compound | Protease Inhibitor Cocktail tablets EDTA-free | Roche Diagnostics Corporation | #5056489001 | |
| Commercial assay or kit | Proteinase K | Life Technologies | #25530–049 | |
| Commercial assay or kit | recJ | New England Biolabs | #M0264L | |
| Commercial assay or kit | Restriction emzyme AfeI | New England Biolabs | #R0652S | |
| Commercial assay or kit | Restriction emzyme EcoRV-HF | New England Biolabs | #R3195S | |
| Commercial assay or kit | Restriction emzyme SacI-HF | New England Biolabs | #R3156S | |
| Commercial assay or kit | RNase A | ThermoFisher Scientific | #EN0531 | |
| Commercial assay or kit | RNase H | New England Biolabs | #M0297S | |
| Commercial assay or kit | SuperScript II Reverse Transcriptase | ThermoFisher Scientific | #18064014 | |
| Commercial assay or kit | T4 DNA Polymerase | New England Biolabs | #M0203S | |
| Chemical compound | Triptolide | TOCRIS Bioscience | #3253 | |
| Chemical compound | TRIzol Reagent | ThermoFisher Scientific | #15596026 | |
| Commercial assay or kit | Direct-zol RNA MiniPrep kit | Genesee | #11–330 | |
| Commercial assay or kit | Fast SYBR Green Master mix | Life Technologies | #4385612 | |
| Commercial assay or kit | Gibson Assembly Master Mix | New England Biolabs | #E2611S | |
| Commercial assay or kit | High-Capacity RNA-to-cDNA Kit | ThermoFisher Scientific | #4387406 | |
| Commercial assay or kit | IBI High Speed Plasmid Mini Kit | MIDSCI | #IB47101 | |
| Commercial assay or kit | NEBNext dA-tailing module | New England Biolabs | #E6053L | |
| Commercial assay or kit | NEBNext end-repair module | New England Biolabs | #E6050L | |
| Commercial assay or kit | NEBNext Multiplex Oligos for Illumina | New England Biolabs | #E7335S | |
| Commercial assay or kit | Q5High-Fidelity 2X Master Mix | New England Biolabs | #M0492L | |
| Commercial assay or kit | Q5 Site-Directed Mutagenesis Kit | New England Biolabs | #E0554S | |
| Commercial assay or kit | Quick Ligation Kit | New England Biolabs | #M2200L | |
| Cell line (D. melanogaster) | Kc167 cells | DGRC | #1 | |
| Cell line (D. pseudoobscura) | ML83-63 cells | DGRC | #33 | |
| Sequence-based reagent | ChIP-nexus oligos | (He et al., 2015) | See Table S5 | |
| Sequence-based reagent | Gene-specific 5’ RNA sequencing oligos | This paper | See Table S6 | |
| Recombinant DNA reagent | Reporters used in this study | This paper | See Table S7 and S8 | |
| Software, algorithm | Analysis code | GitHub | https://github.com/zeitlingerlab/Shao_eLife_2019 |
Additional files
-
Supplementary file 1
2019 eLife Shao Manuscript Supplementary Information.
- https://doi.org/10.7554/eLife.41461.022
-
Transparent reporting form
- https://doi.org/10.7554/eLife.41461.023
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Reporter-ChIP-nexus reveals strong contribution of the Drosophila initiator sequence to RNA polymerase pausing
eLife 8:e41461.
https://doi.org/10.7554/eLife.41461




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}