The tardigrade damage suppressor protein binds to nucleosomes and protects DNA from hydroxyl radicals
- University of California, San Diego, United States
Figures
Figure 1
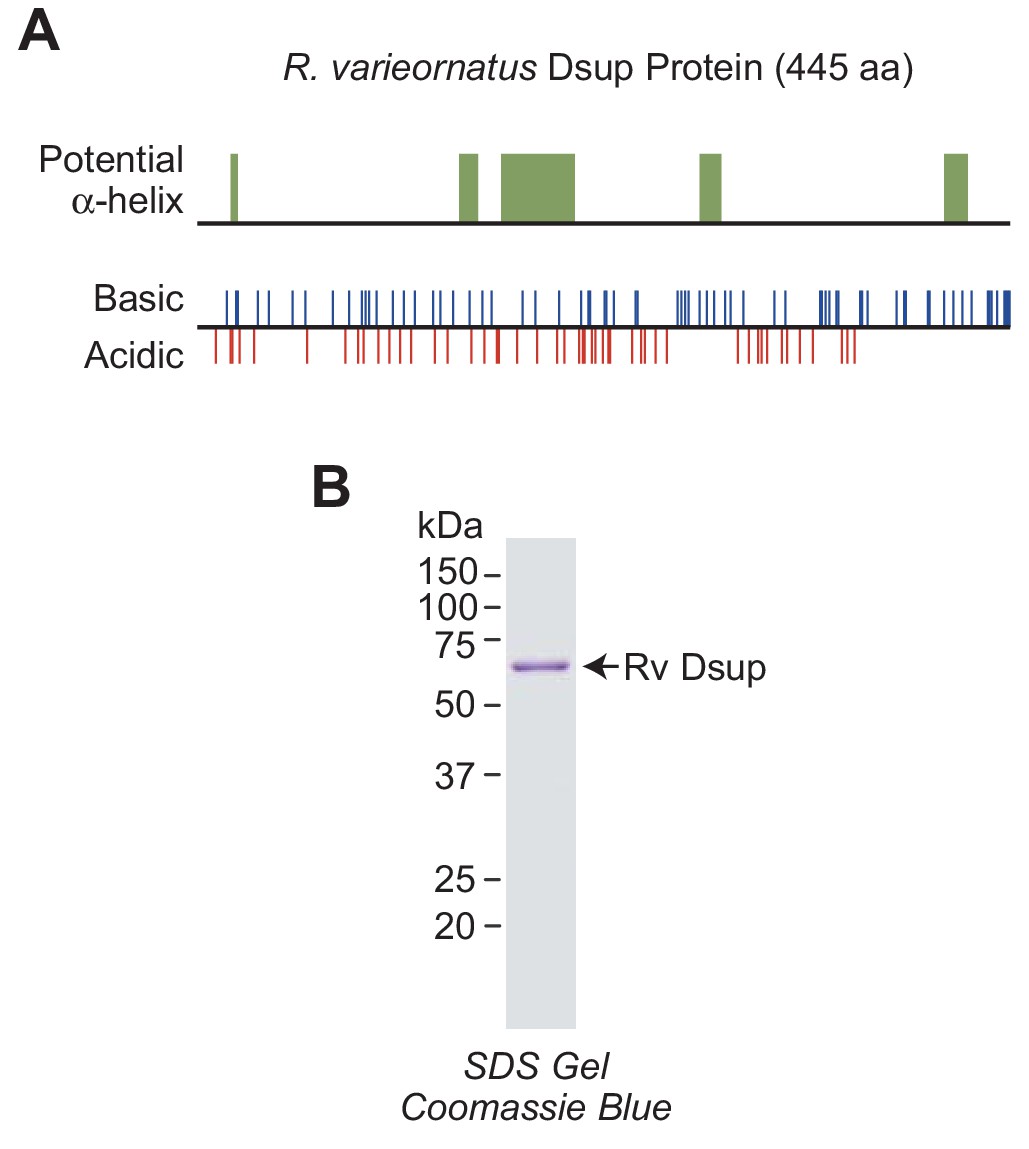
R.varieornatus damage suppressor protein (Rv Dsup).
(A) Rv Dsup is a highly charged protein. The protein was analyzed by using the MPI Bioinformatics Toolkit (Zimmermann et al., 2018), and it is predicted to be disordered except for some potentially alpha-helical regions (upper panel). The positions of basic and acidic amino acid residues are also shown (lower panel). (B) Purification of recombinant Rv Dsup protein synthesized in Escherichia coli. The recombinant Rv Dsup has an N-terminal His6-tag and a C-terminal FLAG tag. The purified protein was analyzed by 12% polyacrylamide-SDS gel electrophoresis and staining with Coomassie Brilliant Blue R-250. The sizes of molecular mass markers (in kDa) are indicated.
Figure 2 with 1 supplement
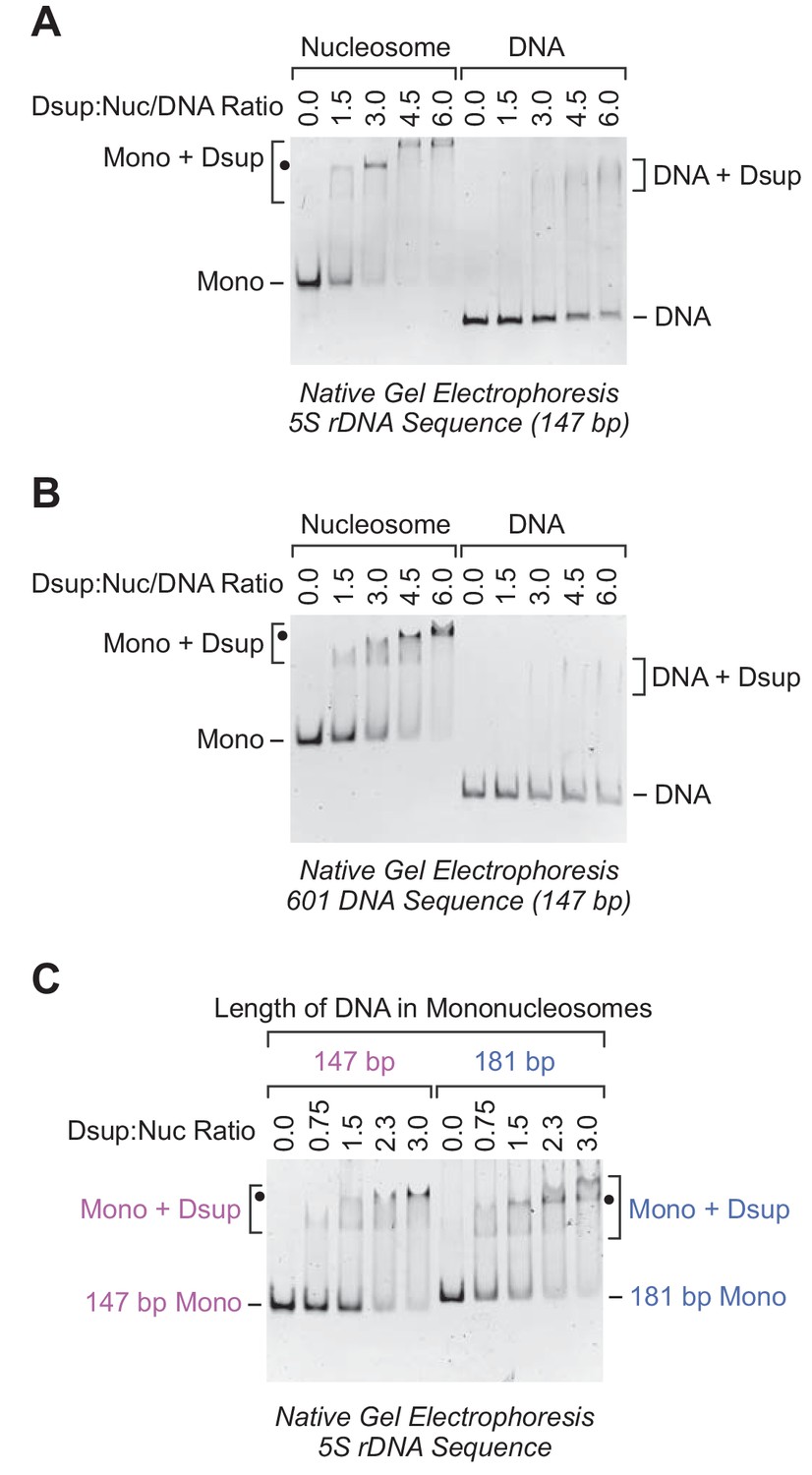
Rv Dsup binds preferentially to mononucleosomes relative to free DNA.
(A) Gel mobility shift analysis with the Xenopus borealis 5S rDNA (147 bp) as either mononucleosomes or free DNA. The indicated amounts of purified Rv Dsup was combined with the mononucleosomes or DNA, and the resulting samples were subjected to nondenaturing 4.5% polyacrylamide gel electrophoresis and staining with ethidium bromide. The positions of mononucleosomes (Mono), Rv Dsup-nucleosome complexes (Mono + Dsup; the major shifted distinct band is denoted by a black dot), free DNA fragments (DNA), and Rv Dsup-DNA complexes (DNA + Dsup) are shown. (B) Gel mobility shift analysis of Rv Dsup with the 601 synthetic DNA sequence (147 bp) as either mononucleosomes or free DNA. Reactions were performed as in A. (C) Effect of DNA length upon binding of Rv Dsup to mononucleosomes. Gel mobility shift assays were performed with mononucleosomes, as in A, with 5S rDNA sequences with a length of either 147 bp or 181 bp.
Figure 2—figure supplement 1
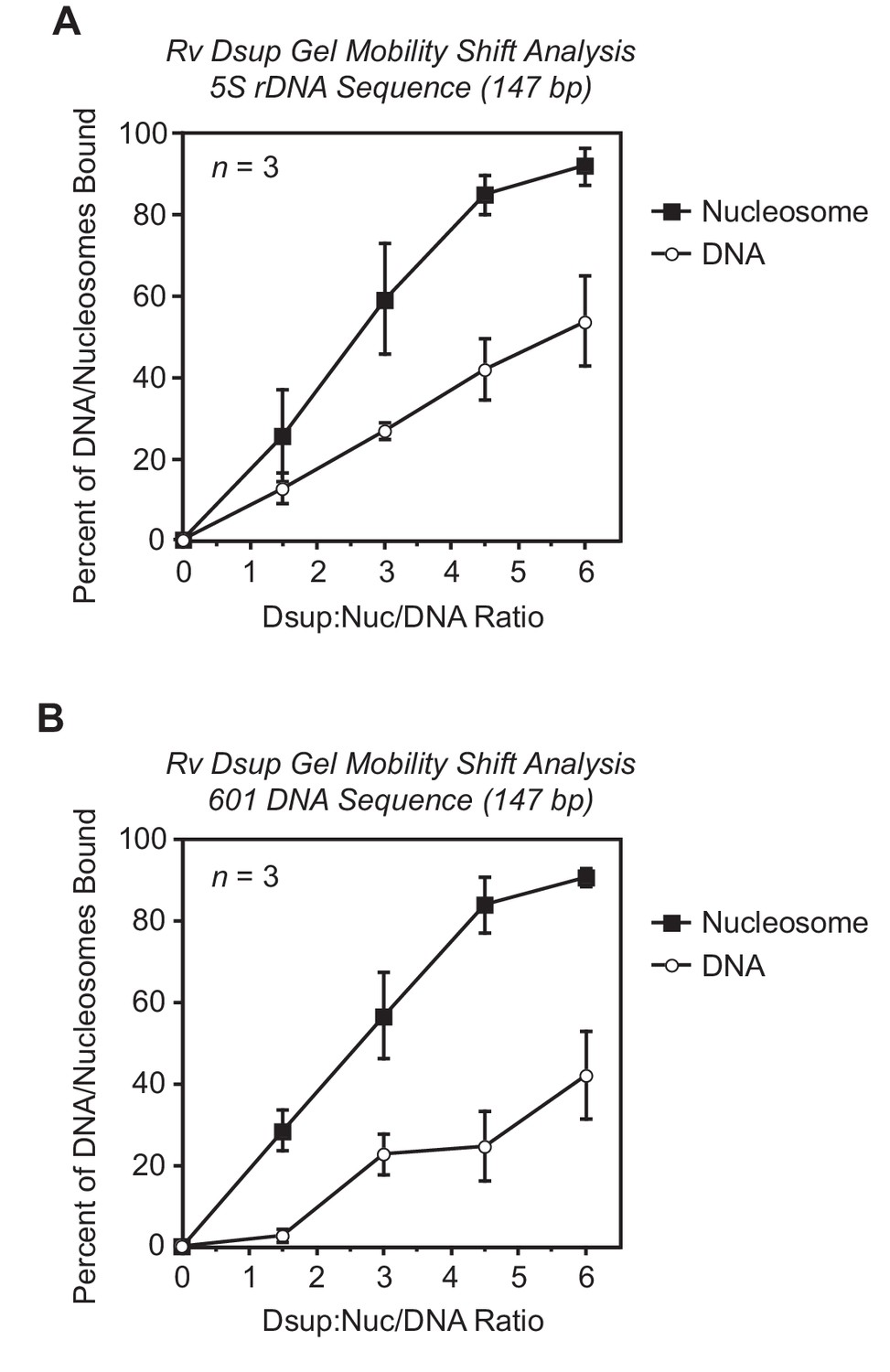
Quantitation of gel mobility shift analyses of Rv Dsup binding to mononucleosomes or free DNA.
Gel mobility shift experiments were performed with Rv Dsup as in Figure 2 of the main text. For each sample, the percent of the mononucleosomes or free DNA that was bound by Dsup was determined by quantitation of the unbound mononucleosomes or free DNA. The x-axis denotes the molar ratio of Dsup to mononucleosomes or free DNA. The results from three independent experiments (n = 3) are shown as the mean + /– standard deviation. (A) Gel mobility shift analysis of Rv Dsup with the Xenopus borealis 5S rDNA (147 bp) as either mononucleosomes or free DNA. (B) Gel mobility shift analysis of Rv Dsup with the 601 synthetic DNA sequence (147 bp) as either mononucleosomes or free DNA.
Figure 3

Rv Dsup can be incorporated into periodic nucleosome arrays.
The ATP-dependent assembly of periodic nucleosome arrays was carried out with purified ACF, dNLP, core histones, ATP, relaxed plasmid DNA, and topoisomerase I (Fyodorov and Kadonaga, 2003; Khuong et al., 2017) in the absence or presence of purified Rv Dsup. (A) DNA supercoiling analysis indicates that Rv Dsup does not affect the efficiency of nucleosome assembly. Chromatin assembly reactions were performed with the indicated amounts of purified Rv Dsup. The reaction products were deproteinized and subjected to 0.8% agarose gel electrophoresis, and the DNA species were visualized by staining with ethidium bromide. Samples of supercoiled DNA and relaxed DNA were included as references. The positions of nicked DNA (N), relaxed DNA (R), and supercoiled DNA (S) are indicated. The asterisk denotes a minor amount of linear DNA that is generated by a nuclease contaminant in the chromatin assembly factors. (B) Partial MNase digestion analysis reveals an increase in the nucleosome repeat length upon incorporation of Rv Dsup into chromatin. Chromatin assembly reactions were performed as in A, and the resulting samples were partially digested with two different concentrations of MNase, deproteinized, and subjected to 1.3% agarose gel electrophoresis. Reactions with purified Drosophila histone H1 were included as a reference. The yellow squares denote the DNA fragments derived from dinucleosomes, and the green dots correspond to the DNA fragments derived from trinucleosomes. The DNA size markers (M) are the 123 bp ladder (Invitrogen). (C) Native gel electrophoresis of mono- and dinucleosome particles obtained from extensive MNase digestion of chromatin that is assembled in the absence or presence of Rv Dsup. Reactions were performed as in A and B except that the products were subjected to extensive MNase digestion followed by nondenaturing 4.5% polyacrylamide gel electrophoresis. Reactions with histone H1 were included as a reference. The chromatin particles were detected by staining with ethidium bromide. The positions of mononucleosomes (Mono), dinucleosomes (Di), monucleosomes with Rv Dsup (Mono + Dsup), mononucleosomes with H1 (Mono + H1), and dinucleosomes with H1 (Di + H1) are indicated. (D) Dsup-mononucleosome particles generated by ACF assembly followed by MNase digestion migrate on native gels at approximately the same rate as Dsup-mononucleosome particles formed by the addition of Dsup to mononucleosomes. Native gel electrophoresis of chromatin particles was performed with salt dialysis-reconstituted mononucleosomes containing Rv Dsup (as in Figure 2A) as well as with mononucleosomes generated by MNase digestion of Rv Dsup-containing chromatin assembled with ACF (as in C). The positions of the mononucleosomes (Mono) and mononucleosome-Rv Dsup complexes (Mono +Dsup) are denoted.
Figure 4

Rv Dsup and histone H1 can bind simultaneously to nucleosomes.
(A) Native gel mobility shift analysis reveals the binding of both Rv Dsup and histone H1 to mononucleosomes. Experiments were performed with 5S rDNA mononucleosomes (181 bp DNA) and the indicated amounts of Rv Dsup and D. melanogaster histone H1. The samples were subjected to nondenaturing 4.5% polyacrylamide gel electrophoresis, and the DNA was visualized by staining with ethidium bromide. The positions of mononucleosomes (Mono), mononucleosome-H1 complexes (Mono + H1), mononucleosome-Rv Dsup complexes (Mono + Dsup), and mononucleosome-Rv Dsup-H1 complexes (Mono + Dsup + H1) are shown. The loss of signal at the highest concentration of H1 appears to be due to H1-mediated aggregation of the sample, as is typically seen with excess H1 (see, for example, Hashimoto et al., 2016). (B) DNA supercoiling analysis indicates that efficiency of nucleosome assembly is not significantly altered by the presence of Rv Dsup and histone H1. Chromatin assembly reactions were performed and analyzed as in Figure 3A. A minor amount of linear DNA is indicated by an asterisk. (C) Partial MNase digestion analysis shows that the nucleosome repeat length in the presence of histone H1 and Rv Dsup is similar to that seen with histone H1 alone. Chromatin assembly reactions were performed and analyzed as in Figure 3B. The yellow squares denote the DNA fragments derived from dinucleosomes, and the green dots correspond to the DNA fragments derived from trinucleosomes.
Figure 5
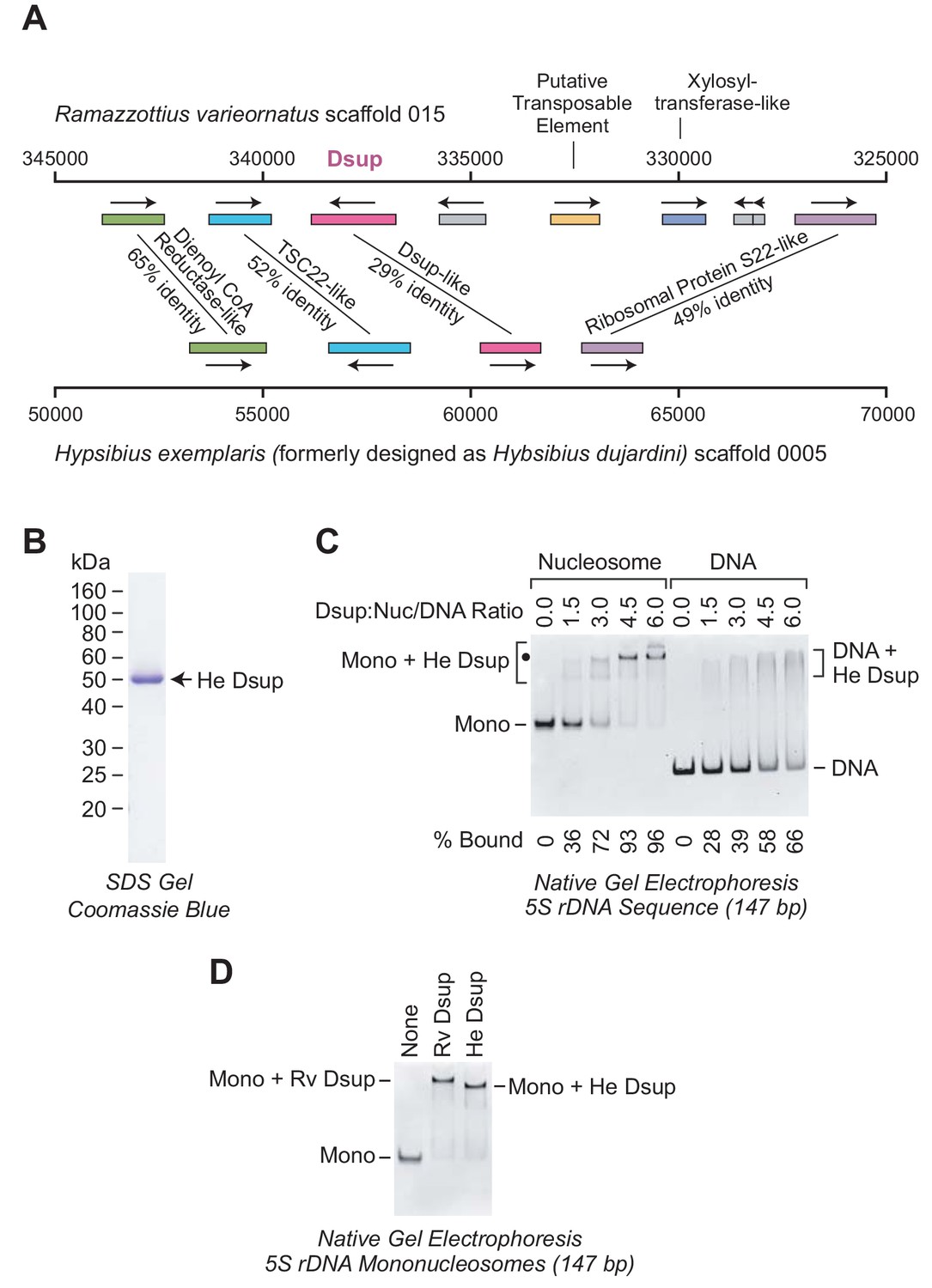
The Dsup-like protein from the tardigrade Hypsibius exemplaris is a nucleosome-binding protein.
(A) Comparison of the genomic regions in the vicinity of the genes encoding R. varieornatus Dsup (Rv Dsup) and H. exemplaris Dsup-like protein (He Dsup). This diagram is based on sequences from R. varieornatus strain YOKOZUNA-1 scaffold 015 (Hashimoto et al., 2016) and H. exemplaris strain Z151 scaffold 0005 (Yoshida et al., 2017). The numbers correspond to the nucleotide positions in the scaffolds. The genomic regions in the vicinity of the R. varieornatus Dsup gene (accession number BAV59442.1) and H. exemplaris Dsup-like gene (accession number OQV24709.1) are shown. The protein sequences were analyzed by pairwise alignment with BLAST (Altschul et al., 1997). The % amino acid identities between the predicted proteins encoded by the corresponding genes are denoted. For each pair of homologous proteins, the number of identical amino acid residues divided by the length (in amino acid residues) of the region of homology is as follows: dienol CoA reductase-like (187/288); TSC22-like (155/300); Dsup (124/434); ribosomal protein S22-like (168/341). The difference in the % amino acid identity between the Rv Dsup and He Dsup proteins in Hashimoto and Kunieda (2017) (26.4%) versus this study (29%) is due to the use of different methods for the pairwise alignments. (B) Purification of recombinant He Dsup protein synthesized in E. coli. The recombinant He Dsup has an N-terminal His6-tag and a C-terminal FLAG tag. The purified protein was analyzed by 12% polyacrylamide-SDS gel electrophoresis and staining with Coomassie Brilliant Blue R-250. The sizes of molecular mass markers (in kDa) are indicated. (C) He Dsup binds preferentially to mononucleosomes relative to free DNA. Gel mobility shift experiments were performed as in Figure 2A. The positions of mononucleosomes (Mono), He Dsup-nucleosome complexes (Mono + He Dsup; the major shifted distinct band is denoted by a black dot), free DNA fragments (DNA), and Dsup-DNA complexes (DNA + He Dsup) are indicated. For each sample, the percent of the mononucleosomes or free DNA that was bound by Dsup was determined by quantitation of the unbound mononucleosomes or free DNA. (D) Comparison of the binding of Rv Dsup and He Dsup to mononucleosomes. Gel mobility shift experiments were performed with 147 bp 5S rDNA mononucleosomes, as in Figures 2A and 5C, with 4.5 molecules of either Rv Dsup or He Dsup per nucleosome.
Figure 6
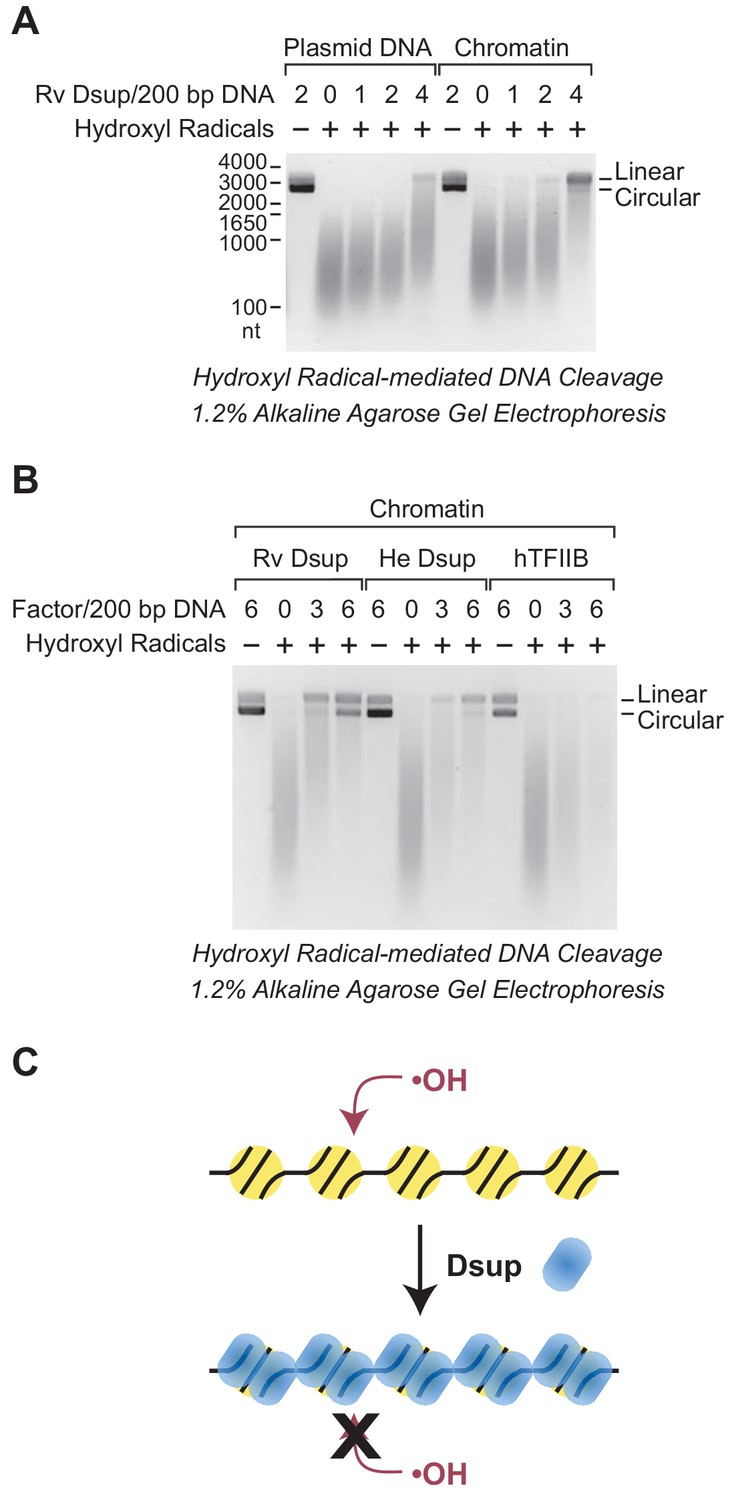
Dsup protects chromatin from hydroxyl radical-mediated DNA cleavage.
Hydroxyl radical-mediated cleavage of DNA was carried out with plasmid pGIE-0 (3.3 kb) as either free DNA or chromatin in the absence or presence of the indicated factors. Where noted, hydroxyl radicals were omitted to test for the possible presence of nuclease activity in the factors. The reaction products were deproteinized, subjected to denaturing alkaline agarose gel electrophoresis, and visualized by staining with GelRed (Biotium). (A) Rv Dsup protects free DNA and chromatin from hydroxyl radical-mediated DNA cleavage. Hydroxyl radical DNA cleavage reactions were each performed under identical conditions with the indicated amounts of Rv Dsup. The full-length linear and circular DNA species are denoted. In addition, the positions of the size markers (in nt) are shown. (B) Hydroxyl radical-mediated DNA cleavage reactions of chromatin in the presence of Rv Dsup, He Dsup, and human TFIIB. Reactions were performed with chromatin as in A. Human TFIIB, a nuclear protein that does not bind to nucleosomes, was included as a control. For reference, the calculated molecular masses of the wild-type (untagged) proteins are as follows: Rv Dsup, 42.8 kDa; He Dsup, 33.0 kDa; human TFIIB, 34.8 kDa. (C) Model for Dsup binding to nucleosomes and protecting chromatin from hydroxyl radicals. In this model, Dsup binds specifically to nucleosomes and protects the DNA from damaging agents such as hydroxyl radicals via coverage of the chromatin with its disordered regions that are enriched in SAGK residues.
Figure 7
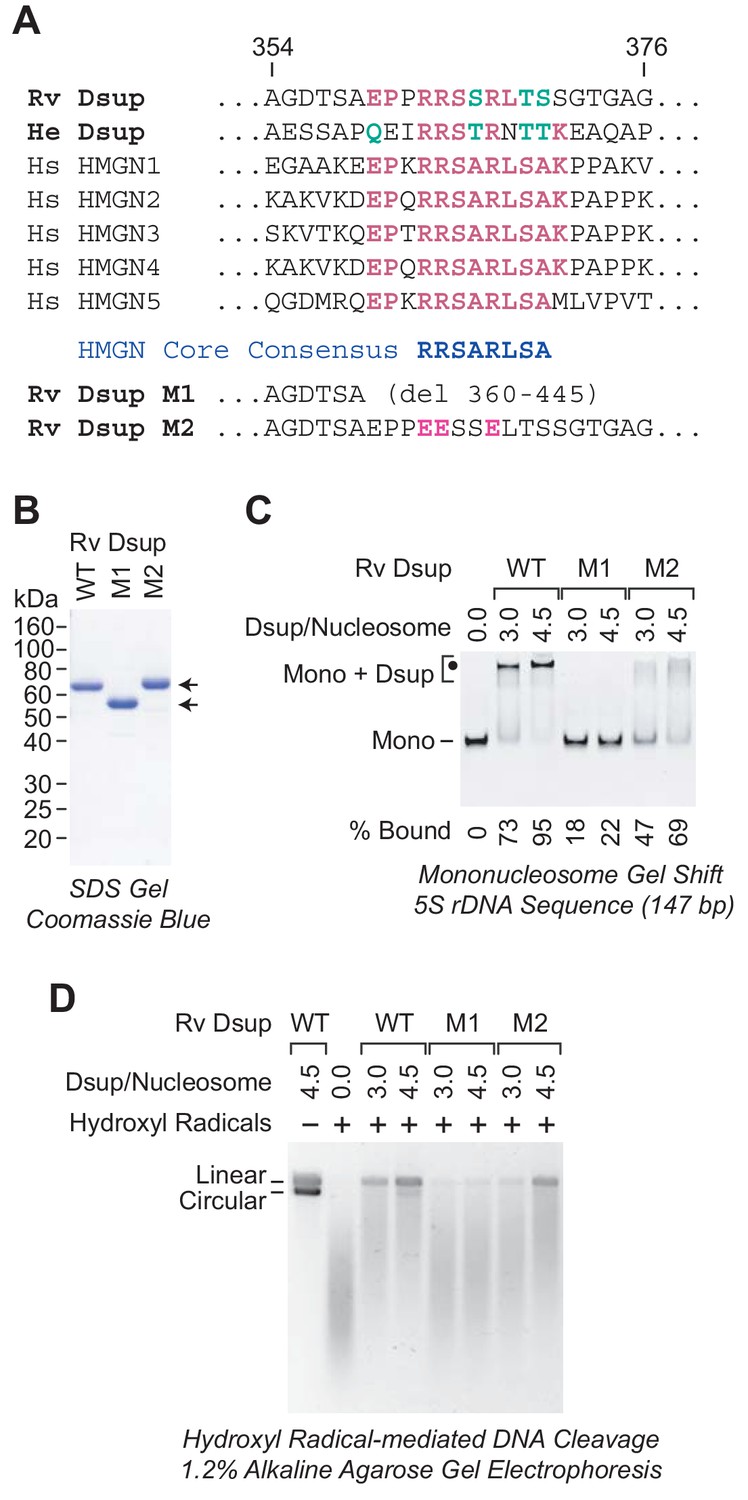
A region of sequence similarity between Dsup proteins and the nucleosome-binding domain of vertebrate HMGN proteins is important for the binding of Dsup to nucleosomes.
(A) Alignment of Dsup proteins with the five human HMGN proteins as representative members of the HMGN protein family. Partial sequences of Rv Dsup, He Dsup, and the five human HMGN proteins are shown. The conserved HMGN consensus in the nucleosome-binding domain (Ueda et al., 2008) is indicated. Identical amino acid residues are highlighted in red type, and conserved amino acid substitutions (as in, for example, Wu and Brutlag, 1996) are in green type. The numbers indicate the amino acid residues in Rv Dsup. The amino acid sequences of Rv Dsup mutant M1 and mutant M2 are shown at the bottom. The modified residues in M2 are highlighted in pink type. (B) Purification of Rv Dsup mutant proteins M1 and M2. The purified proteins were analyzed by 12% polyacrylamide-SDS gel electrophoresis and staining with Coomassie Brilliant Blue R-250. The sizes of molecular mass markers (in kDa) are indicated. (C) The M1 and M2 mutant Dsup proteins bind less efficiently to mononucleosomes than wild-type Dsup. Gel mobility shift experiments were performed as in Figure 2A. The positions of mononucleosomes (Mono) and Dsup-nucleosome complexes (Mono + Dsup; the major shifted distinct band is denoted by a black dot) are indicated. For each sample, the percent of the mononucleosomes or free DNA that was bound by Dsup was determined by quantitation of the unbound mononucleosomes or free DNA. (D) The M1 and M2 mutant Dsup proteins are less effective at protecting chromatin from hydroxyl radical-mediated DNA cleavage than wild-type Dsup. Hydroxyl radical-mediated DNA cleavage reactions were performed with chromatin as in Figure 6. The reaction products were deproteinized, subjected to denaturing alkaline agarose gel electrophoresis, and visualized by staining with GelRed (Biotium). The full-length linear and circular DNA species are denoted.
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.47682.011
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The tardigrade damage suppressor protein binds to nucleosomes and protects DNA from hydroxyl radicals
eLife 8:e47682.
https://doi.org/10.7554/eLife.47682




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}