In vitro reconstitution of branching microtubule nucleation
- University of Exeter, United Kingdom
Figures
Figure 1 with 1 supplement
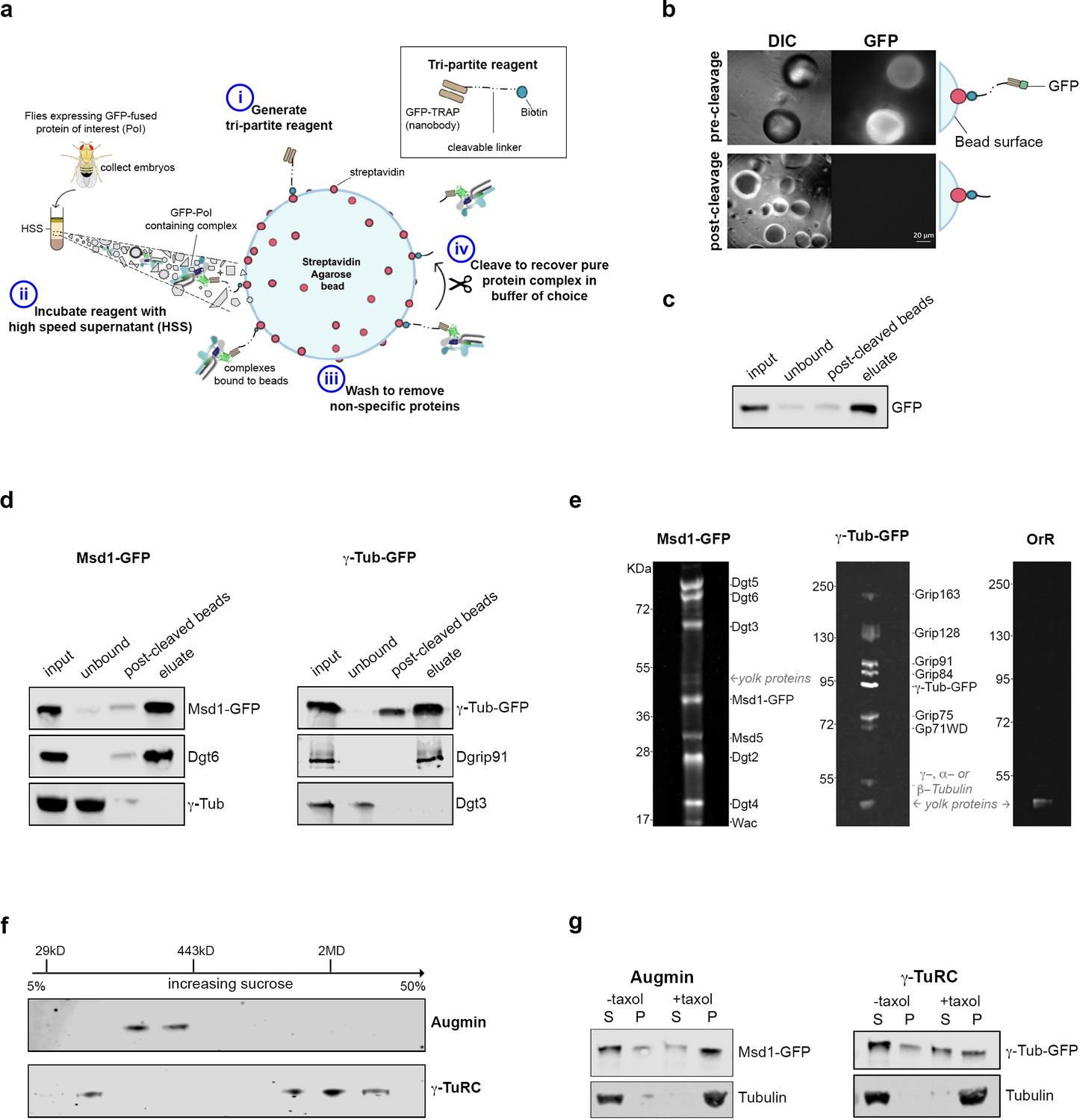
Isolation of functional γ-TuRC and Augmin using cleavable affinity purification.
(a) Sketch of purification methodology. (b) Images of GFP-TRAP-Sulfo beads after incubation with His-GFP, pre- and post-cleavage by 50 mM DTT. (c) Western blot demonstrating the effective isolation and cleavage of soluble His-GFP. (d) Western blots of cl-AP of Msd1-GFP and γ-Tubulin-GFP. The fusion proteins, present in embryo extracts (input), are efficiently depleted upon incubation with GFP-TRAP-Sulfo beads (unbound), released in 50 mM DTT (post-cleaved beads) and present in the eluate. Subunits of Augmin, but not γ-TuRC, co-elute with Msd1-GFP. Subunits of γ-TuRC, but not Augmin, co-elute with γ-Tubulin-GFP. (e). SYPRO-ruby stained gels of post-cleaved eluates from control embryos (OrR), or MG132-treated (mitotic) embryos expressing the Augmin subunit Msd1-GFP or γ-Tubulin-GFP. (f) Western blot of sucrose gradient fractionation of purified mitotic Augmin or γ-TuRC. Complexes sediment as expected for their molecular weights. (g) Western blots of in vitro MT co-sedimentation assays. In the absence of taxol (-), Tubulin, pure Augmin (Msd1-GFP) and pure γ-TuRC (γ-Tubulin-GFP) remain in the supernatant (S). In the presence of taxol (+), Tubulin polymerises and is present in the pellet (P). Augmin and γ-TuRC co-sediment.
Figure 1—figure supplement 1
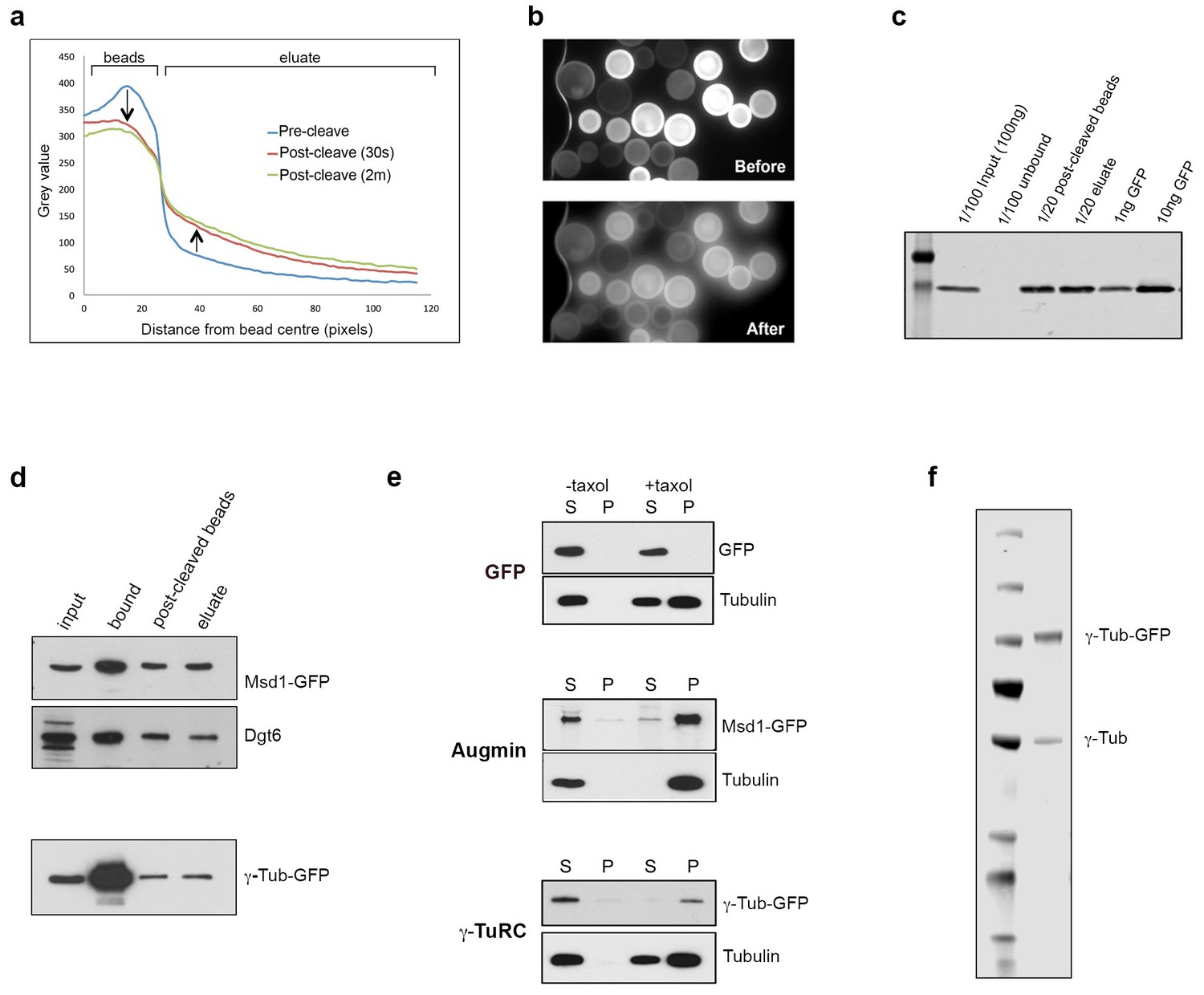
Isolation of functional γ-TuRC and Augmin using GFP-TRAP-PC beads.
(a) Graph showing the comparative decrease in GFP fluorescence on GFP-TRAP-PC beads, and increase in the surrounding eluate, following exposure to UV light (UV filter block on TE2000U fluorescence microscope). (b) Images of GFP-TRAP-PC beads after incubation with GFP, pre- and post-cleavage by 30 s exposure to UV light, used to generate data for (a). (c) Western blot to quantify the binding and release capacity of GFP-TRAP-PC beads. 100 ng of GFP is completely immobilised on the beads. In this case,~60% of GFP is cleaved via UV exposure, corresponding to 3 ng x 20 = 60 ng GFP. (d) Western blots demonstrating the isolation and cleavage of Msd1-GFP and γ-Tubulin-GFP using GFP-TRAP-PC beads. The proteins, present in embryo extracts (input), are efficiently captured onto beads (bound), with ~50% released following UV exposure (post-cleaved beads and eluate). (e) Western blots of in vitro MT co-sedimentation assays using GFP-TRAP-PC beads. In the absence of taxol (-), Tubulin, pure Augmin (Msd1-GFP) and pure γ-TuRC (γ-Tubulin-GFP) remain in the supernatant (S). In the presence of taxol (+), Tubulin polymerises and is present in the pellet (P). Augmin and γ-TuRC co-sediment. GFP alone does not co-sediment with MTs in this assay. (f) Western blot of purified γ-Tubulin-GFP eluate, probed with anti-γ-Tubulin antibody. Endogenously expressed, untagged γ-Tubulin is co-purified with γ-Tubulin-GFP, presumably as part of active γ-TuRCs, at a ratio of ~3:1 γ-Tubulin-GFP: γ-Tubulin (as quantified by LiCOR analysis).
Figure 2 with 2 supplements
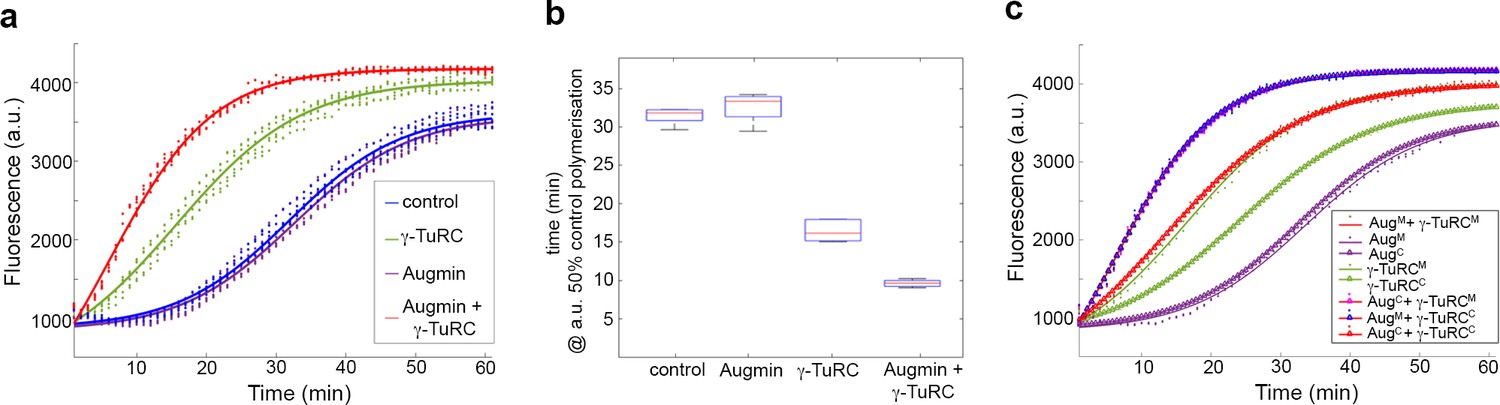
Pure Augmin enhances γ-TuRC-dependent MT nucleation in a cell cycle dependent manner.
(a) Tubulin polymerisation assays, where fluorescence is directly related to the amount of Tubulin polymer present. The curves are a sigmoidal fit to six replicates of the experiment (dots); three independent purification experiments, each undertaken in duplicate. (b) Plot of the x50 in relation to control polymerisation assay, showing the median (red line), interquartile ranges (blue box) and 95% confidence intervals of the median (notches) for each condition. The differences in the time taken for 50% polymerisation between all conditions differs significantly at p=0.001, except when comparing control to Augmin (ANOVA). (c) Fluorescent tubulin polymerisation assays undertaken with complexes isolated from MG132-treated (mitotic) or cycling embryos.
Figure 2—figure supplement 1

The synergistic effect of pure Augmin on γ-TuRC-dependent MT nucleation in vitro is dependent on the Augmin-γ-TuRC interface.
(a) Representation of the interactions between Augmin subunits, as determined by cross-linking mass spectrometry. The N-terminus of Dgt3 and Dgt5, and the C terminus of Dgt6 have all been previously shown to be responsible for localising the γ-TuRC to the mitotic spindle in Drosophila embryos. (b) Western blot showing the purification of MBP-Dgt3N, MBP-Dgt5N and MBP-Dgt6C, expressed in bacteria. (c) Fluorescent tubulin polymerisation assays undertaken with GFP-Augmin and GFP-γ-TuRC isolated from MG132-treated embryos, in the presence and absence of Dgt3N, Dgt5N and Dgt6C. Inclusion of the Dgt regions completely abrogates the ability of Augmin to affect γ-TuRC-dependent MT nucleation, without affecting γ-TuRC-dependent MT nucleation itself. (d) Individual datasets from (c). Note that inclusion of the Dgt regions with γ-TuRC alone does not affect γ-TuRC-dependent MT nucleation but does slightly reduce the total amount of MT polymer formed (middle graph). (a and b reproduced from Chen et al., 2017).
Figure 2—figure supplement 2
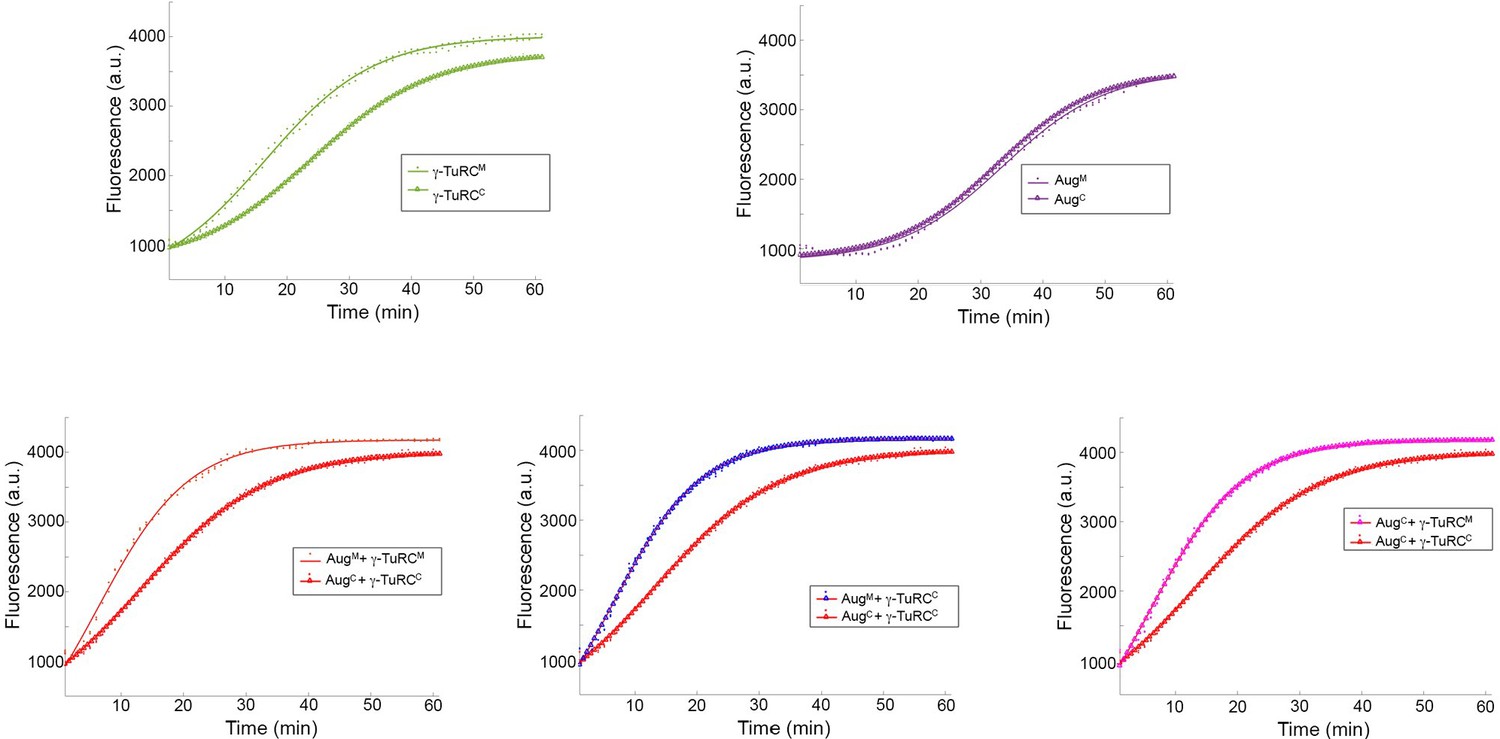
Individual sets of polymerisation curves using Augmin and γ-TuRC purified from either mitotic (M) or cycling (C) embryo extracts, taken from Figure 2c.
Mitotic γ-TuRC stimulates MT nucleation/polymerisation to a greater extent than γ-TuRC isolated from cycling embryo extracts. Neither cycling nor mitotic Augmin affects MT nucleation/polymerisation. Addition of mitotic Augmin to mitotic γ-TuRC stimulates γ-TuRC-dependent MT nucleation. This effect is reduced when cycling Augmin and cycling γ-TuRC are used. However, if either Augmin or γ-TuRC is mitotic, Augmin-dependent MT nucleation is active, even if the other complex is isolated from cycling embryos.
Figure 3 with 1 supplement
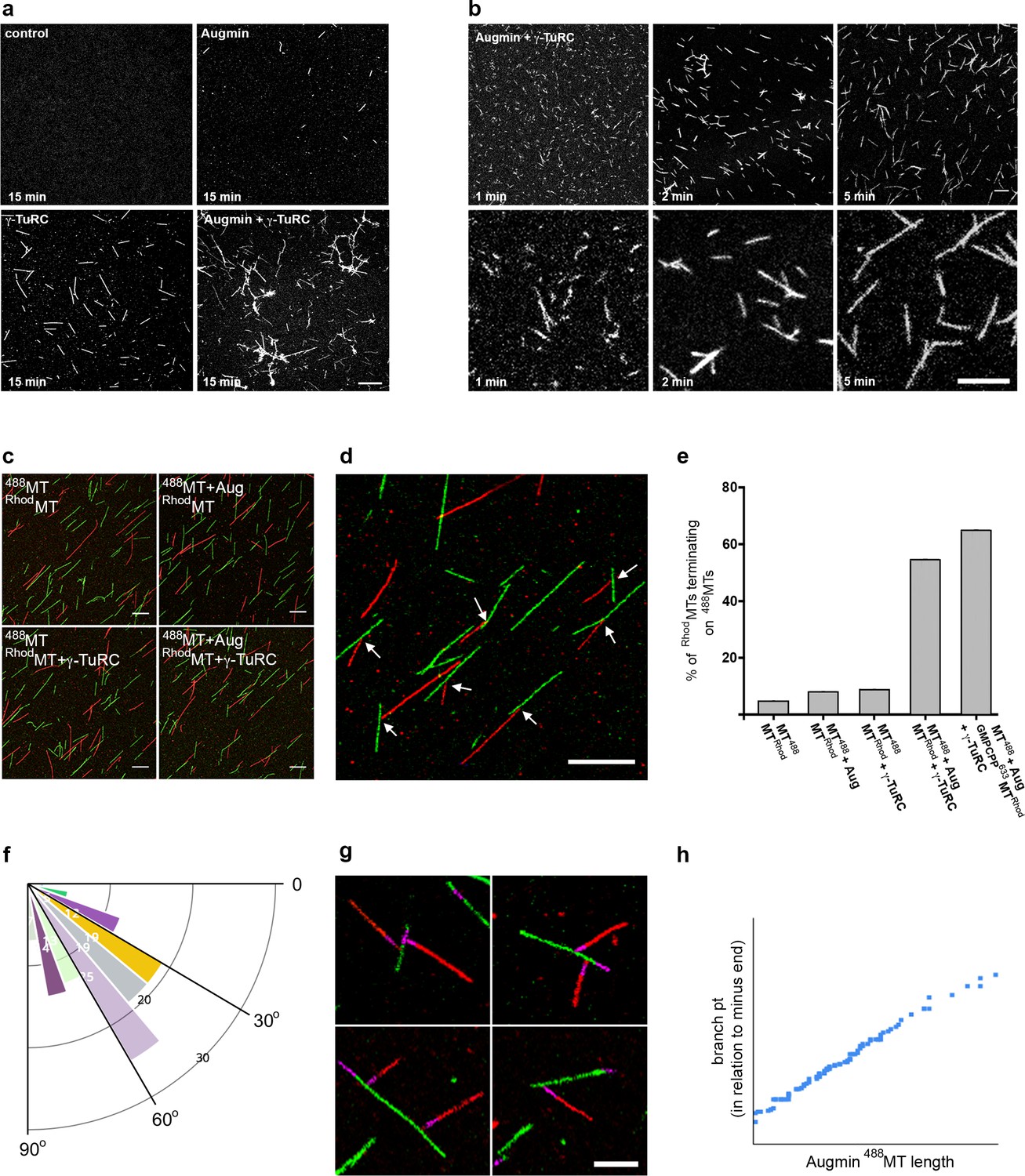
Reconstitution of the MT-Augmin-γ-TuRC-MT junction.
(a) Confocal images of fixed, fluorescent polymerisation assays at t = 15 mins for each condition. (b) Confocal images of fixed, fluorescent polymerisation assays at t = 1, 2 and 5 mins in the presence of purified Augmin and γ-TuRC. (c) Confocal images of taxol stabilised MTs, formed in the absence or presence of purified Augmin and γ-TuRC, and co-incubated. Incubation of Augmin-488MTs (green) with γ-TuRC-RhodMTs (red) leads to MT branches. (d) Higher magnification of the MT-Augmin-γ-TuRC-MT junctions (arrows). (e) Histogram of the percentage of RhodMTs that terminate precisely at a 488MT, generating a branchpoint. (f) Distribution of γ-TuRC-RhodMTs junction angles, in relation to Augmin-488MTs. (g) Confocal images of junctions, using GMPCPP seeds (purple) to distinguish the MT minus ends. Minus ends of γ-TuRC-RhodMTs (red) interact with Augmin-488MTs (green). (h) Pearson correlation coefficient plot demonstrating a strong positive correlation between the length of Augmin-488MTs and the position of γ-TuRC-RhodMT branch points. Scale bars, a-d, 5 μm; scale bar, g, 2 μm.
-
Figure 3—source data 1
Excel spreadsheets of the individual data points corresponding to Figure 3b (number and length of MTs per field of view at t = 2 and t = 5 mins); Figure 3d (Histogram of % of Rhod-MTs terminating on a 488-MT – i.e. % of γ-TuRC activity); Figure 3e (diagram of branch angles); Figure 3f (branchpoint distance from MT minus end, as assessed by GMPCPP seed).
- https://cdn.elifesciences.org/articles/49769/elife-49769-fig3-data1-v1.xlsx
Figure 3—figure supplement 1
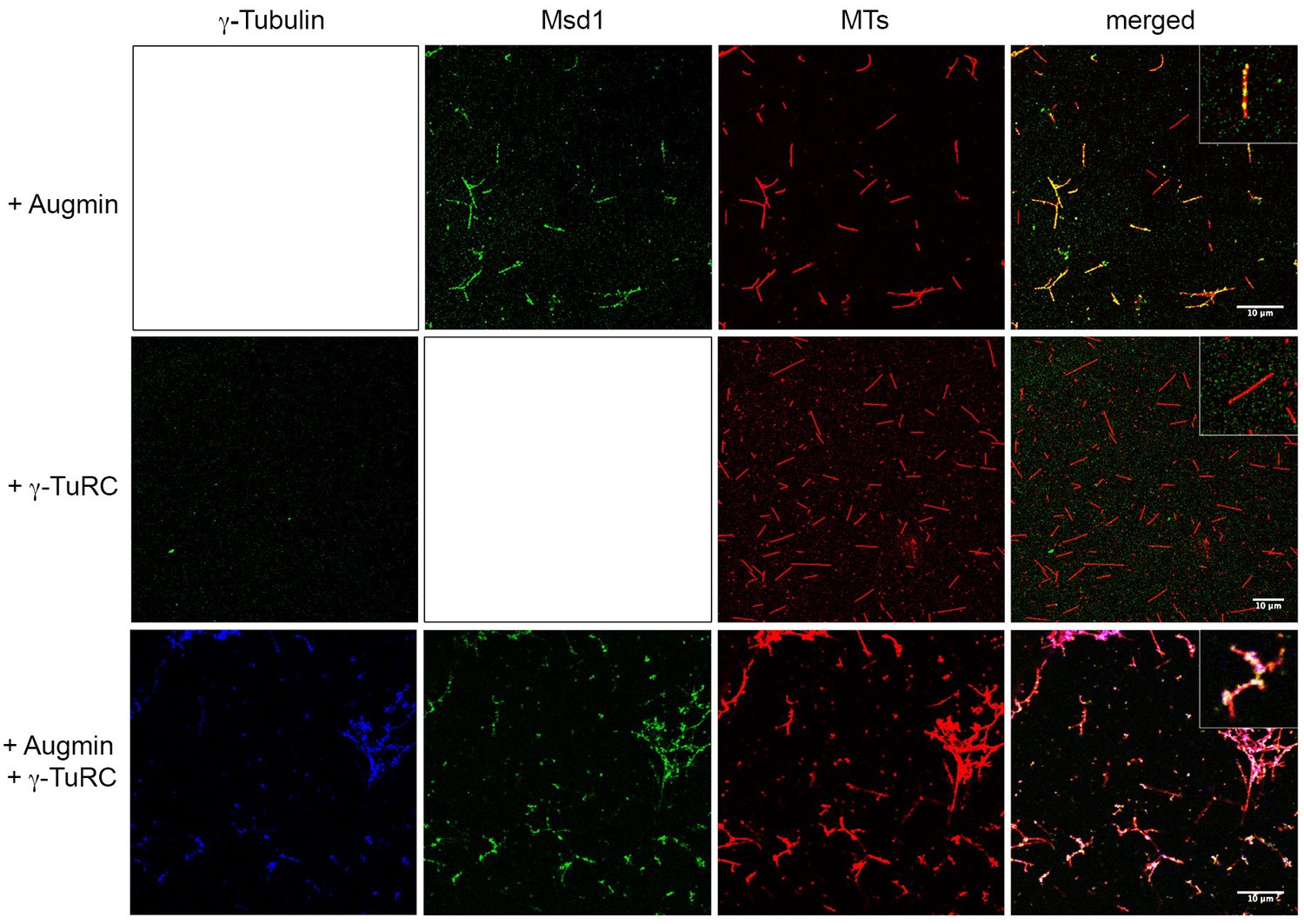
Augmin recruits γ-TuRC to MTs.
Samples were taken mid-way through in vitro polymerisation reactions, fixed and stained for γ-Tubulin, the Augmin Msd1 subunit or MTs. When present on its own, Augmin localises along the length of MTs. Note this localisation is not uniform; rather Augmin is present as punctae. When γ-TuRC is present on its own, γ-Tubulin is not observed on MTs, even though it stimulates MT nucleation (presumably the levels on the minus ends of MTs are too low to be resolved). However, when γ-TuRC is present with Augmin, γ-Tubulin co-localises with Augmin on MTs. Scale bar, 10 μm. See insets for higher magnified images.
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
In vitro reconstitution of branching microtubule nucleation
eLife 9:e49769.
https://doi.org/10.7554/eLife.49769




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}