Exchange of water for sterol underlies sterol egress from a StARkin domain
- Weill Cornell Medical College, United States
Figures
Figure 1 with 3 supplements

Different modes of Lam4S2-membrane association deduced from 'Stage 1' molecular dynamics simulations.
(A) Structural elements of the Lam4S2 domain used to construct collective variables (CVs) for time-structure independent components analysis (tICA). Residues Q121, S181, D61 are labeled. Also highlighted are the locations of the Ω1 loop (purple), the C-terminal helix (yellow), the N-terminus (green), and the C-terminus (red). Cholesterol is shown in space-filling representation colored cyan except for the oxygen atom in red. (B) Initial positioning of Lam4S2 (cartoon) near the membrane. In this configuration, the distance between any atom of the protein and any atom of a lipid molecule was ≥10 Å. The cholesterol molecule bound to Lam4S2 is shown in space-fill representation. The water box including solution ions is omitted for clarity. (C) For each residue of Lam4S2, the fraction of trajectory frames from Stage 1 simulations in which the residue is in contact with the membrane was determined and plotted. A residue was considered to be in contact with the bilayer if the z-coordinate of the Cα atom of this residue was within 1 Å of the average z-position of the neighboring lipid phosphate atoms (identified as those located within 10 Å of this Cα atom). The relevant protein segments are labeled and colored using the color-code used in panel (A). (D, E) Two modes of Lam4S2-membrane association. The lipids in the membrane are shown as lines. The relevant protein segments are colored using the color-code used in panel (A). For completeness, panels (D, E) also show the protein-bound cholesterol molecule in space-fill, as well as residues K163, K167, K62, K90, K98, K203, and Q121.
-
Figure 1—source data 1
Frequency of contacts between Lam4S2 residues and the membrane from MD simulations.
- https://cdn.elifesciences.org/articles/53444/elife-53444-fig1-data1-v2.zip
Figure 1—figure supplement 1
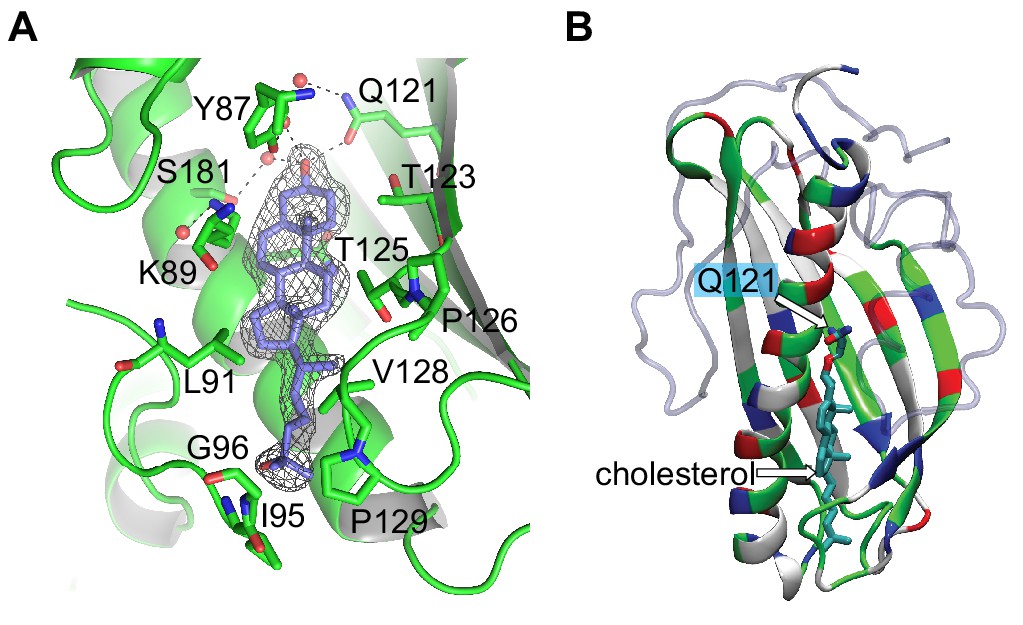
Lam4S2 binding pocket.
(A) Lam4S2 with bound 25HC. This figure shows an ergosterol molecule in the binding site, surrounded by its electron density, as well as key residues that engage the sterol molecule. (B) Polarity of the Lam4S2 binding pocket. The structure shows a bound cholesterol molecule (taken from Figure 1A) and the key Q121 residue. The color code used for the protein ribbon indicates positive (blue), negative (red), polar (green), and hydrophobic (white) residues.
© 2018, The American Society for Biochemistry and Molecular Biology Inc. All Rights Reserved. Panel A adapted from Jentsch et al. (2018), with permission. It is not covered by the CC-BY 4.0 license and further reproduction of this panel would need permission from the copyright holder.
Figure 1—figure supplement 2
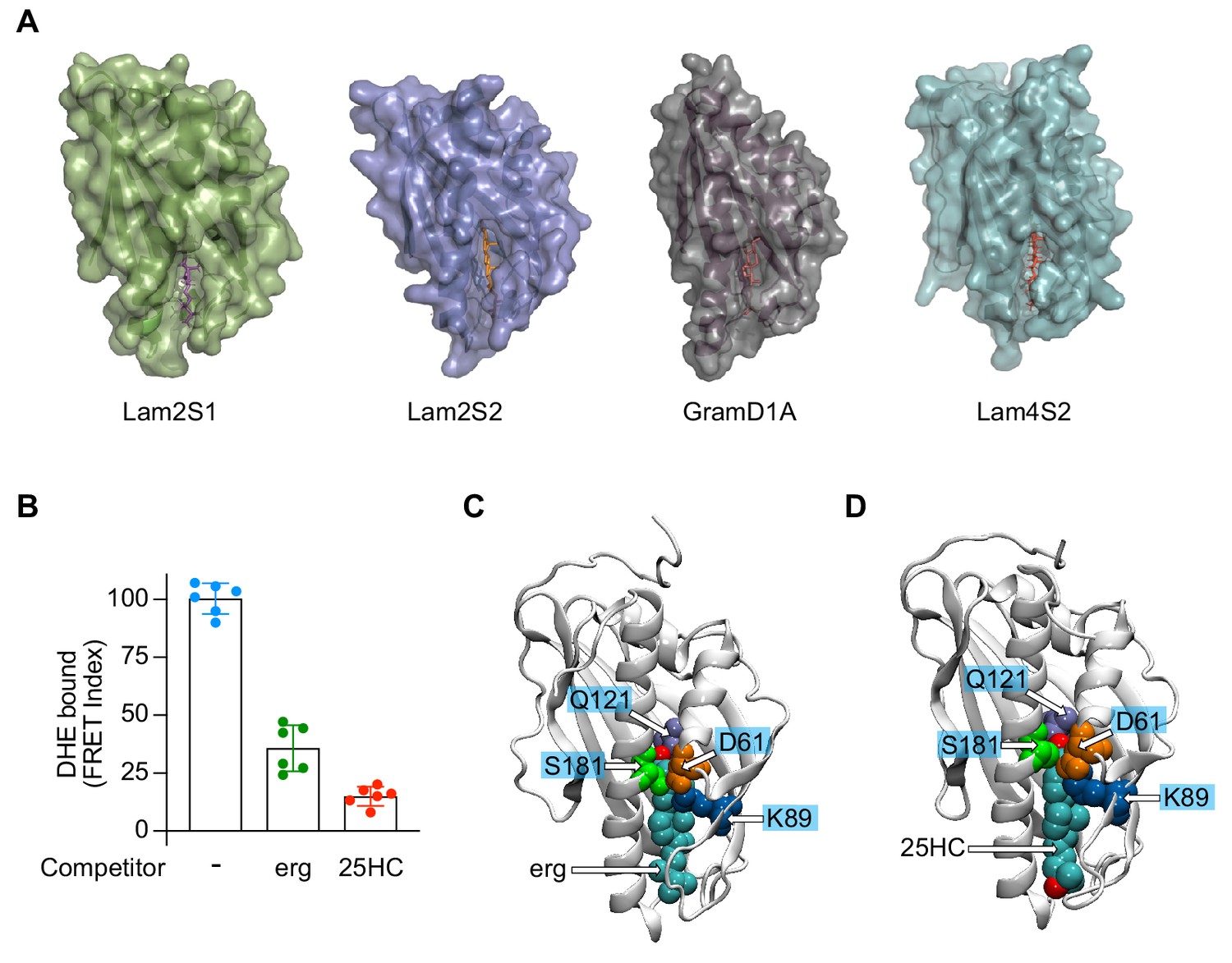
Side opening in Lam/GramD1 structures and sterol binding specificity.
(A) Crystal structures (surface representation) of Lam/GramD1 domains with bound sterol. The structures (PDB ID: 6CAY (Lam2S1), 5YS0 (Lam2S2), 6GQF (GramD1a), 6BYM (Lam4S2)) are oriented with their sterol entry/exit site at the bottom, displaying the lateral fracture (side-opening) through which the bound sterol is visible from the bulk solvent. (B) Competition assay showing that both ergosterol and 25HC compete with DHE for binding to Lam4S2. The assay, performed as described by Gatta et al. (2015), reports Förster resonance energy transfer (FRET) between a tryptophan residue in Lam4S2 and bound DHE. Lam4S2 (1 μM) was mixed with 3 μM DHE (in the absence or presence of 3 μM ergosterol or 25HC [all sterols were added from stock solutions in methanol]) and incubated for 30 min at room temperature. A fluorescence emission spectrum was obtained over the range 300–450 nm (λex = 290 nm) and, after background subtraction, the ratio of intensities at 373 nm and 340 nm was used as an index of FRET. Results were normalized by setting the average FRET Index for the Lam4S2–DHE complex in the absence of competitor as 100. (C) Model of ergosterol bound to Lam4S2 from an atomistic MD simulation. The protein is shown in cartoon, and the sterol as well as selected protein residues are shown in space fill and are labeled. The ergosterol–Lam4S2 complex was built by docking an ergosterol molecule into Lam4S2 using the cholesterol–Lam4S2 structure, which was the starting point for the Stage 2 ensemble simulations. The resulting ergosterol–Lam4S2 system was then subjected to MD simulations using NAMD 2.13. The model was first energy-minimized and then equilibrated with unbiased MD for ~23 ns. (D) Model of 25HC bound to Lam4S2 from the X-ray structure with PDBID 6BYM. The protein is shown in cartoon, and the sterol as well as selected protein residues are shown in space fill and are labeled.
Figure 1—figure supplement 3
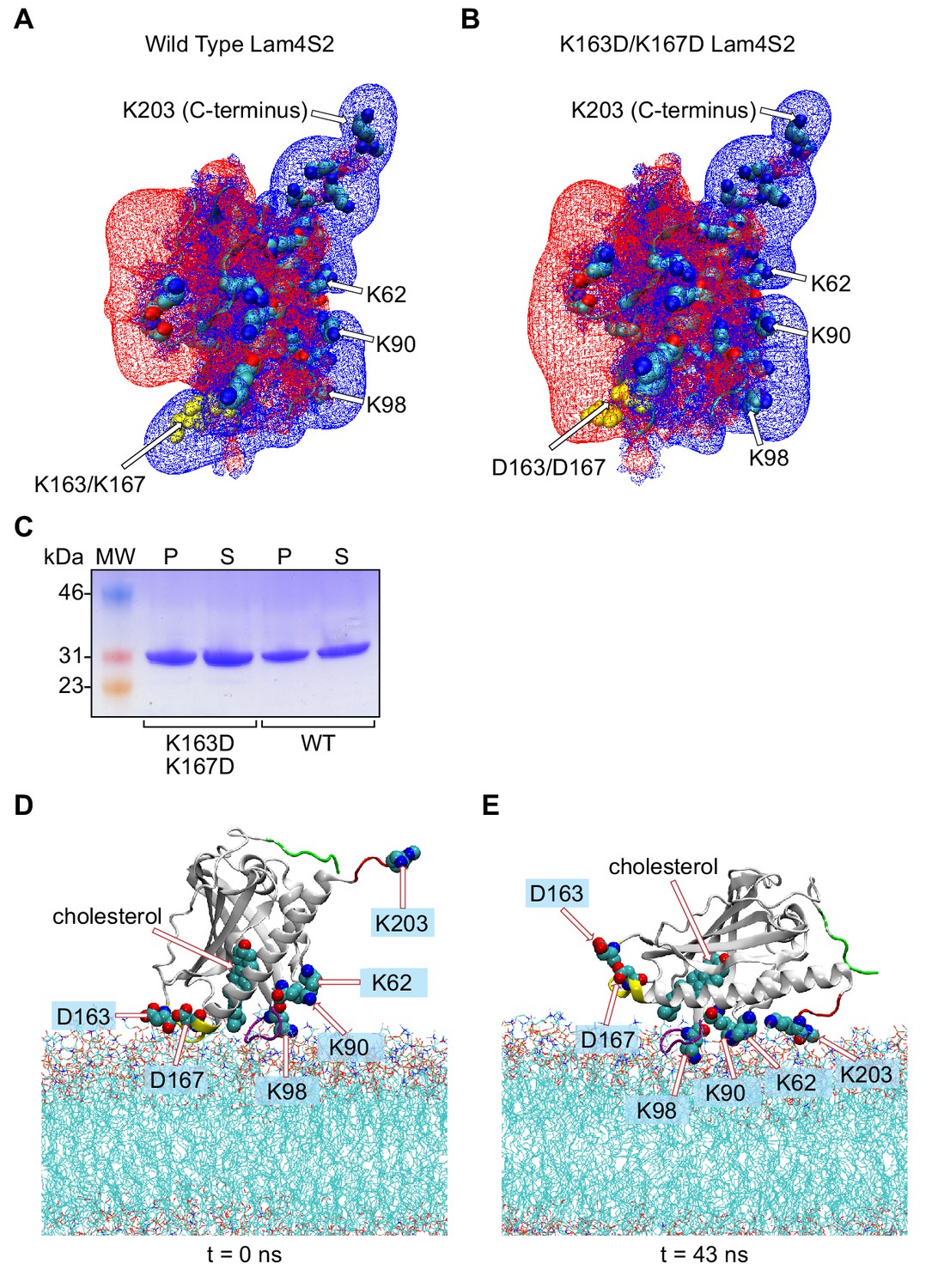
Charge-mediated modes of membrane binding by Lam4S2.
(A, B) Electrostatic potential of wild type Lam4S2 (A) and the K163D/K167D double mutant (B). The blue and red wireframes represent +1kt/e and −1kt/e electrostatic potential iso-surfaces, respectively, and were obtained by solving the non-linear Poisson-Boltzmann (NLPB) equation with APBS (see Methods). The protein is shown in cartoon and all the basic residues are illustrated in space-fill representation. Residues K62, K90, K98, and K203 are labeled, and the amino acids at positions 163 and 167 are highlighted in yellow and are labeled. (C) Binding of wild-type and K163D/K167D Lam4S2 to membranes. The binding assay was performed as described in ‘Materials and methods (Sterol extraction assay)’, and the relative amount of protein in the supernatant (S) and pellet (P) fractions was quantified by Coomassie-stained SDS-PAGE analysis of sample equivalents. A representative gel is shown (molecular weight [MW] markers are indicated). The percentage of protein bound was (mean ± SD, n = 3) 48.5 ± 0.5 (wild-type) and 46.0 ± 2.0 (K163D/K167D mutant). (D, E) Positioning of K163D/K167D Lam4S2 at the start (D) and end (E) of a 43 ns MD simulation. The protein is engaged with the membrane via Mode 2 at the start of the simulation and re-orients over time to interact with the membrane via Mode 1. The lipids in the membrane are shown as lines. The relevant protein segments are colored using the following color-code: Ω1 loop (purple), C-terminal helix (yellow), N-terminus (green), and C-terminus (red). The cholesterol molecule, as well as selected protein residues, are shown in space fill and are labeled.
Figure 2 with 6 supplements
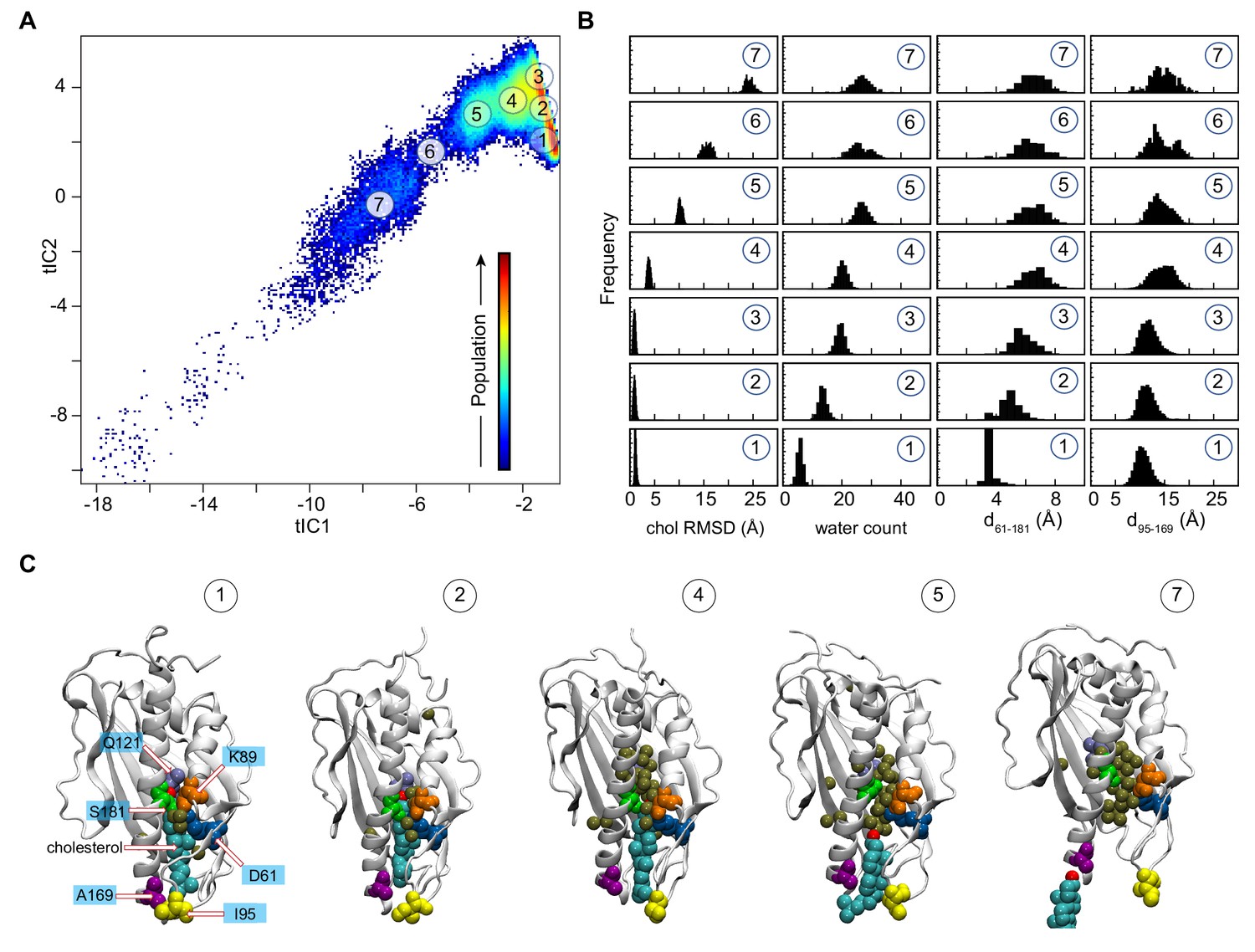
Mechanistic steps of cholesterol release from Lam4S2 revealed by tICA analysis.
(A) 2-D landscape representing all of the Stage 2 MD trajectories mapped with the tICA transformation in the space of the first two tICA eigenvectors (tIC 1 and tIC 2). The lighter shades (from red to light green to yellow) indicate the most populated regions of the 2D space (see the color bar). Microstates representing the most populated states in these simulations are indicated by the numbered circles (1-7) and represent various stages in the lipid translocation process. (B) Characteristics of the selected microstates. The columns record the probability distributions of the cholesterol RMSD, number of water oxygens in the sterol-binding pocket, and distances between residues 61 and 181 (d61–181) and between residues 95 and 169 (d95–169). (C) Structural models representing selected microstates. In these snapshots, Lam4S2 is shown in cartoon, and cholesterol as well as selected protein residues (Q121, D61, K89, S181, I95, A169) are shown in space fill (the residues are labeled in the snapshot of Microstate 1). Water oxygens in the sterol-binding site are drawn as gold spheres.
-
Figure 2—source data 1
Characteristics of different microstates from tICA approach.
- https://cdn.elifesciences.org/articles/53444/elife-53444-fig2-data1-v2.zip
Figure 2—figure supplement 1

tICA analysis of Stage 2 simulations.
(A) Contribution of each tIC vector to the total fluctuation of the system in Stage 2 simulations. (B) The full set of Stage 2 trajectories mapped on the 2D landscape of the first two tICA eigenvectors (tIC 1 and tIC 2). Also shown are the locations of the 100 microstates (red squares) obtained from the k-means clustering analysis of the conformational space. The lighter shades on the two-dimensional space indicate the most populated regions (see also Figure 2).
Figure 2—figure supplement 2
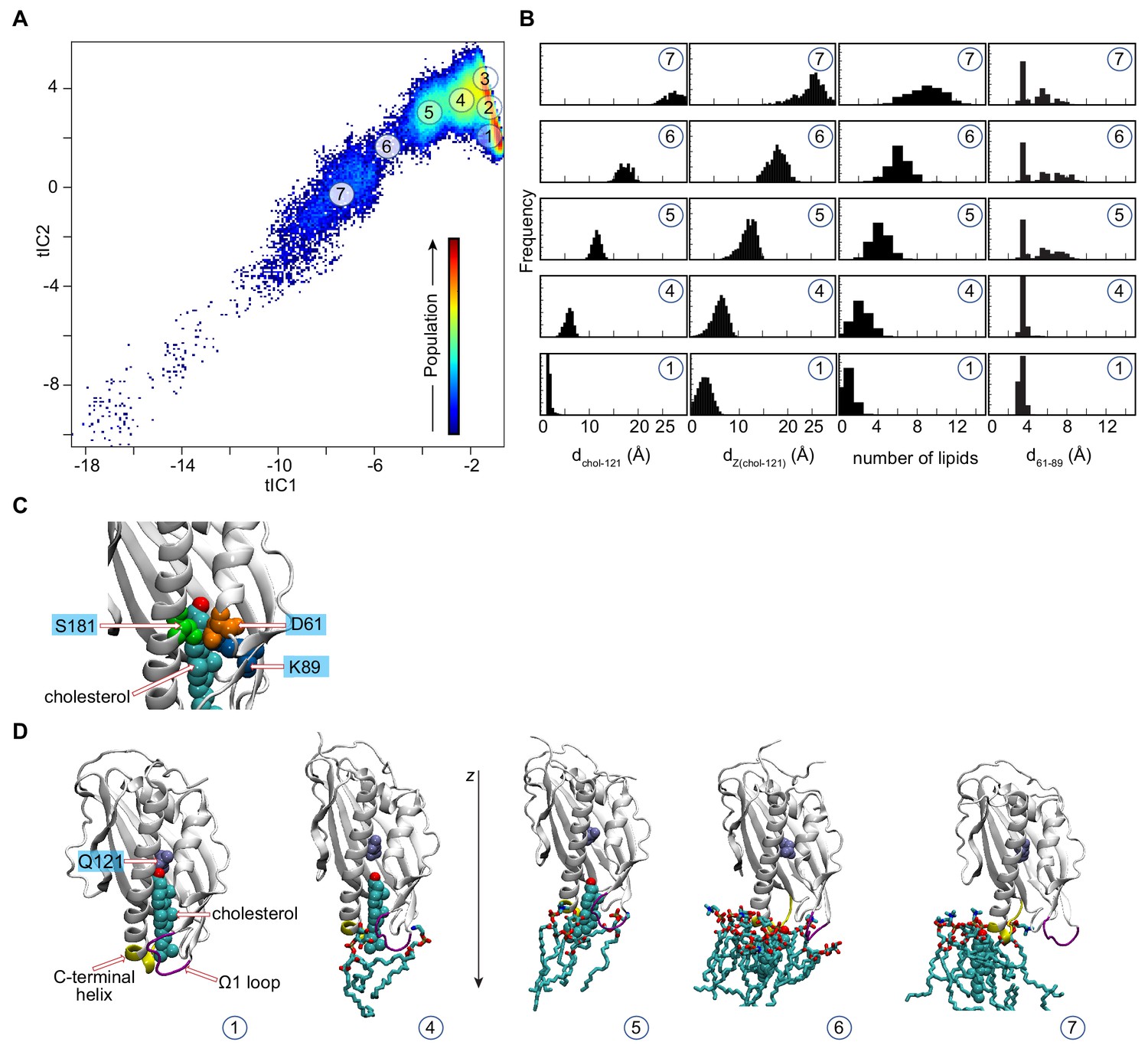
Mechanistic steps of cholesterol release from Lam4S2 revealed from tICA analysis.
(A) Two-dimensional (2-D) landscape (reproduced from Figure 2A for ease of reference) representing all of the Stage 2 MD trajectories mapped with the tICA transformation in the space of the first two tIC vectors. The lighter shades (from red to light green to yellow) indicate the most populated regions of the 2D space (see the color bar). Microstates representing the most populated states in these simulations are indicated by the numbered circles (1-7) (Maxfield and van Meer, 2010; Menon, 2018; Holthuis and Menon, 2014; Wong et al., 2019; Wong and Levine, 2016; Alpy and Tomasetto, 2005; Gatta et al., 2015) and represent various stages in the lipid translocation process. (B) Structural characteristics of selected microstates. The columns from left to right record the probability distributions of dchol–121 distance, dZ(chol–121) distance, number of lipids within 3 Å of cholesterol, and d61–89 distance. (C) Structural snapshot of Microstate 1 zoomed at the side-entrance to the sterol binding pocket highlighting the juxtaposition of D61, K89, and S181 residues (labeled). For completeness, the cholesterol molecule is also shown in space-fill representation and is labeled. (D) Structural models representing microstates from panel (B). These snapshots illustrate gradual immersion of the cholesterol molecule into the membrane as it exits the binding site. In these models, lipids within 3 Å of cholesterol are shown in licorice, cholesterol in space-fill, and residue Q121 is colored in ice blue and labeled. In addition, the C-terminal helix and the Ω1 loop are depicted in yellow and purple, respectively, and the z-axis direction is highlighted.
Figure 2—figure supplement 3
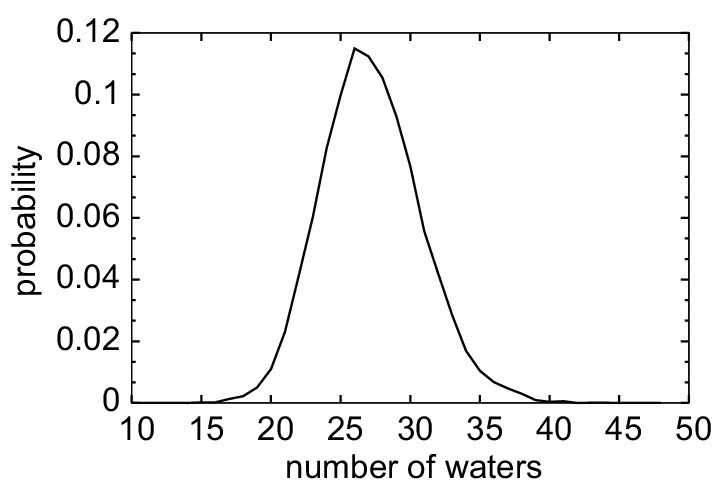
Probability distribution of the number of water oxygens in the sterol-binding pocket calculated from analysis of Stage 1 ensemble MD simulations of apo Lam4S2 (based on PDB ID 6BYD).
The high degree of hydration of the binding pocket seen for the apo protein recapitulates the level of solvation observed in the MD simulations of the cholesterol-bound Lam4S2 after sterol exit (see Microstates 5–7 in Figure 2B).
Figure 2—figure supplement 4

Sterol release sampled in multiple trajectories.
Projection on the 2D tICA landscape from Figure 2A of each individual trajectory from Stage 2 simulations. The 2D tICA map is drawn as in Figure 2A but using smaller-size transparent dots. The colors of the larger dots indicate the time-frames in the evolution of the trajectories: darker colors (blue, cyan) represent the initial stages of the simulations, lighter colors dots (yellow, green) correspond to the middle part of the trajectories, and shades of red show the last third of the trajectories. The tICA landscapes boxed in red represent simulations in which the cholesterol translocation process was sampled in its entirety. The landscapes marked with a green star are simulations in which the system evolved from Microstate 1 to Microstate 5 but did not progress further.
Figure 2—figure supplement 5

Selection of wild-type trajectory frames to initiate simulations of D61A, S181A, and K89A mutants.
Time-evolution of d61–181 distance (A), minimum distance between cholesterol and Q121 residue (dchol–121) (B), and number of water oxygens in the sterol-binding site (C) during one of the Stage 2 simulations of the wild-type protein in which sterol release was observed. The two trajectory frames that were selected for initiating simulations of the mutants are marked by arrows (120 ns and 150 ns time-points). Simulation of the D61A, S181A, and K89A mutants is shown in Figure 2—figure supplement 6.
Figure 2—figure supplement 6
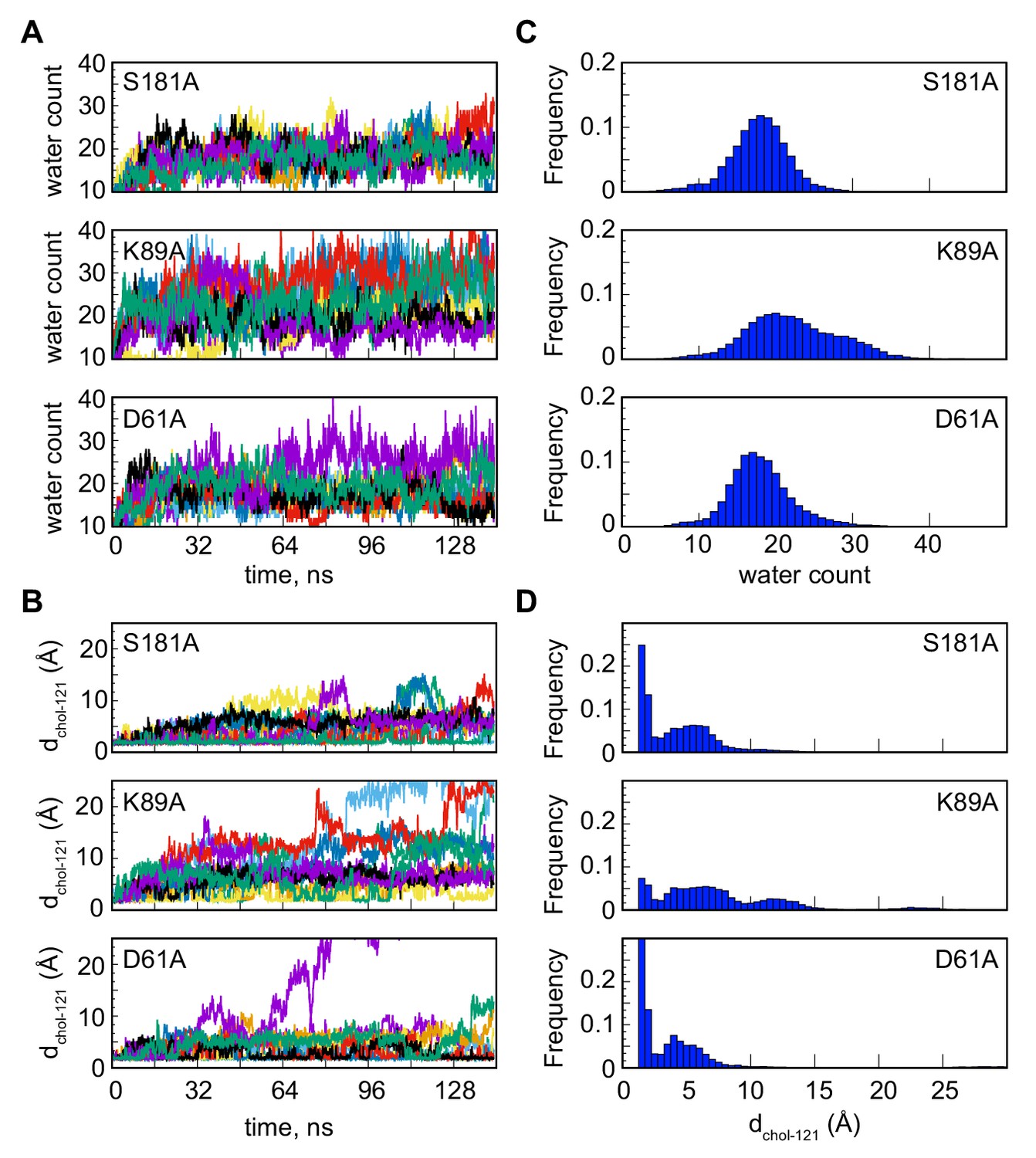
Cholesterol destabilization during unbiased ensemble MD simulations of the S181A, K89A and D61A Lam4S2 mutants.
(A) Number of water oxygens in the sterol-binding site as a function of time in simulations of S181A, K89A, and D61A Lam4S2 (top, middle, and bottom panels, respectively). The results for 10 statistically independent replicates in the corresponding ensemble of trajectories are shown in different color traces. (B) Minimum distance between cholesterol and residue Q121 (dchol–121) as a function of time in simulations of S181A, K89A, and D61A Lam4S2 (top, middle, and bottom panels, respectively). The results for 10 statistically independent replicates in the corresponding ensemble of trajectories are shown in different color traces. (C) Histograms of number of water oxygens corresponding to the time-traces in panel (A) (data from the entire time-period were used to construct the histograms). (D) Histograms of dchol–121 corresponding to the time-traces in panel (B) (data from the entire time-period were used to construct the histograms).
Figure 3 with 1 supplement
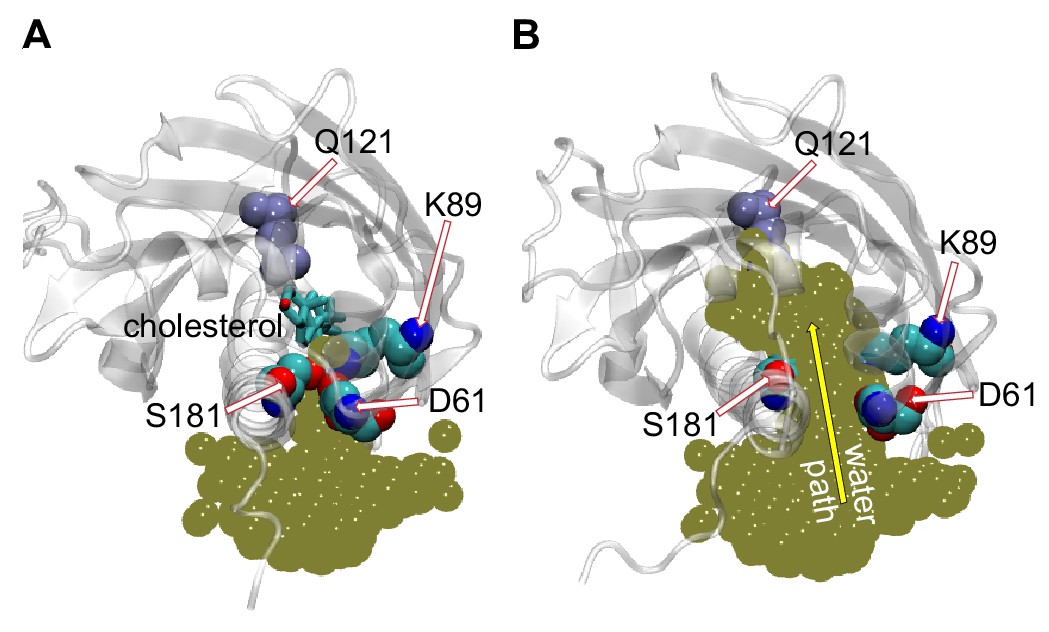
Penetration of water into the binding pocket through the side-opening is a key step in the sterol-release process.
(A, B) Top view of the sterol-binding pocket in Lam4S2, illustrating closed (A) and open (B) conformations of the side-opening to the binding site (the protein models are representative structures from Microstates 1 and 7, respectively). In both snapshots, residues D61, K89, and S181 (which line the side-opening) and residue Q121 (which coordinates the cholesterol hydroxyl group) are highlighted (in space-fill and labeled). The gold spheres in panels (A) and (B) represent the superposition of water oxygens in the binding site and near the side-entrance from one of the Stage 2 trajectories before (panel A) and after (panel B) the side-entrance opens. In panel (A), the cholesterol is shown as licorice. A water pathway to the binding pocket, formed when the side-entrance is open, but not when it is closed, is illustrated in panel (B) by the yellow arrow.
Figure 3—figure supplement 1
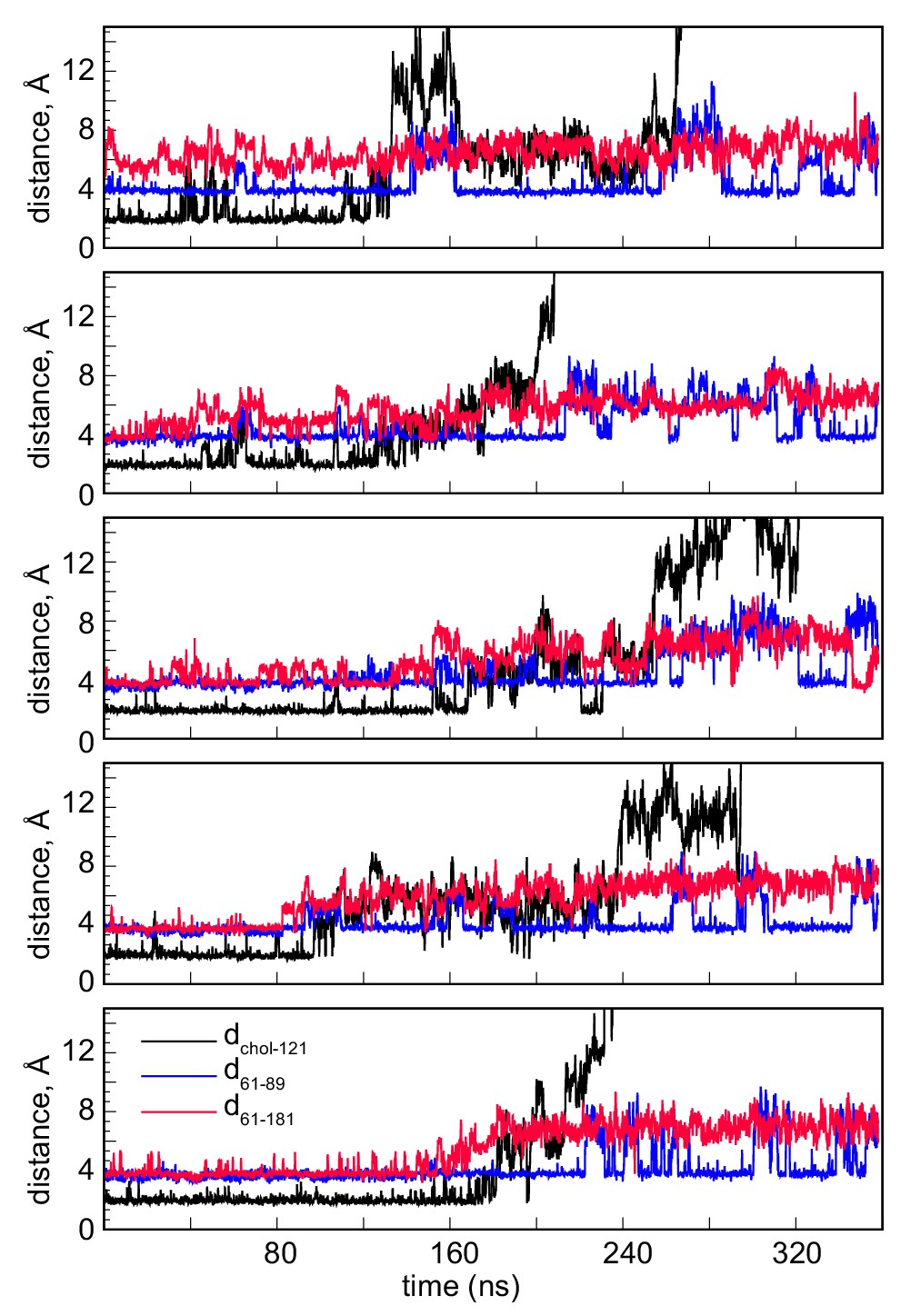
Minimum distances between cholesterol and residue Q121 (dchol–121, black), between residues D61 and K89 (d61–89, blue), and between residues D61 and S181 (d61–181, red) as a function of time in the five MD simulation trajectories from Stage 2 in which full release of the sterol molecule was observed.
The time-traces show that cholesterol destabilization is preceded by increases in the d61–181 distance (>4 Å) and that its departure is followed by larger fluctuations in the d61–89 distance.
Figure 4 with 3 supplements
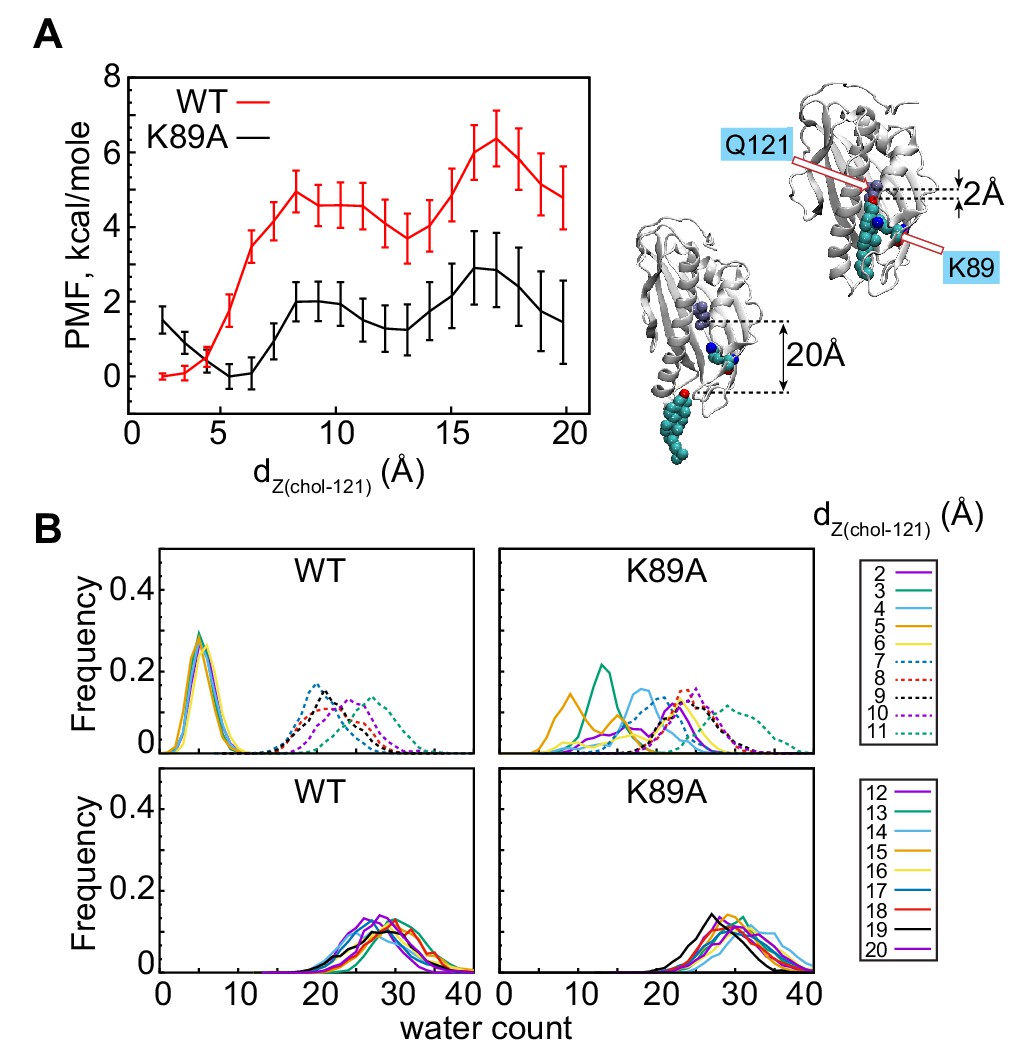
The K89A mutation reduces the energy barrier for cholesterol release.
(A) Potential of mean force (PMF) as a function of dZ(chol–121) distance for wild-type (red) and K89A (black) Lam4S2, calculated from umbrella sampling MD simulations at each dZ(chol–121). The structural representations on the right side of the panel illustrate locations of cholesterol corresponding to dZ(chol–121) ~2 Å (top) and dZ(chol–121) ~20 Å (bottom). Residues Q121 and K89 in these snapshots are also shown. (B) Histograms of the number of water oxygens in the sterol-binding site constructed from analysis of trajectories representing various windows in the range of dZ(chol–121)∈ (2Å; 20Å) from the umbrella MD simulations of the wild-type (left panels) and K89A (right panels) systems.
-
Figure 4—source data 1
Potential of mean force for wild-type and K89A Lam4S2.
- https://cdn.elifesciences.org/articles/53444/elife-53444-fig4-data1-v2.zip
-
Figure 4—source data 2
Water counts in wild-type and K89A Lam4S2 corresponding to different umbrella windows.
- https://cdn.elifesciences.org/articles/53444/elife-53444-fig4-data2-v2.zip
Figure 4—figure supplement 1

The K89A mutation promotes opening of the side-opening to the binding pocket and the influx of water.
Histograms of dchol–121 (A, B) and d61–181 (C, D) distances constructed from analysis of trajectories representing various windows in the range of dZ(chol–121)∈ (2Å; 11Å) from the umbrella MD simulations of the wild type (A, C) and K89A (B, D) systems.
Figure 4—figure supplement 2
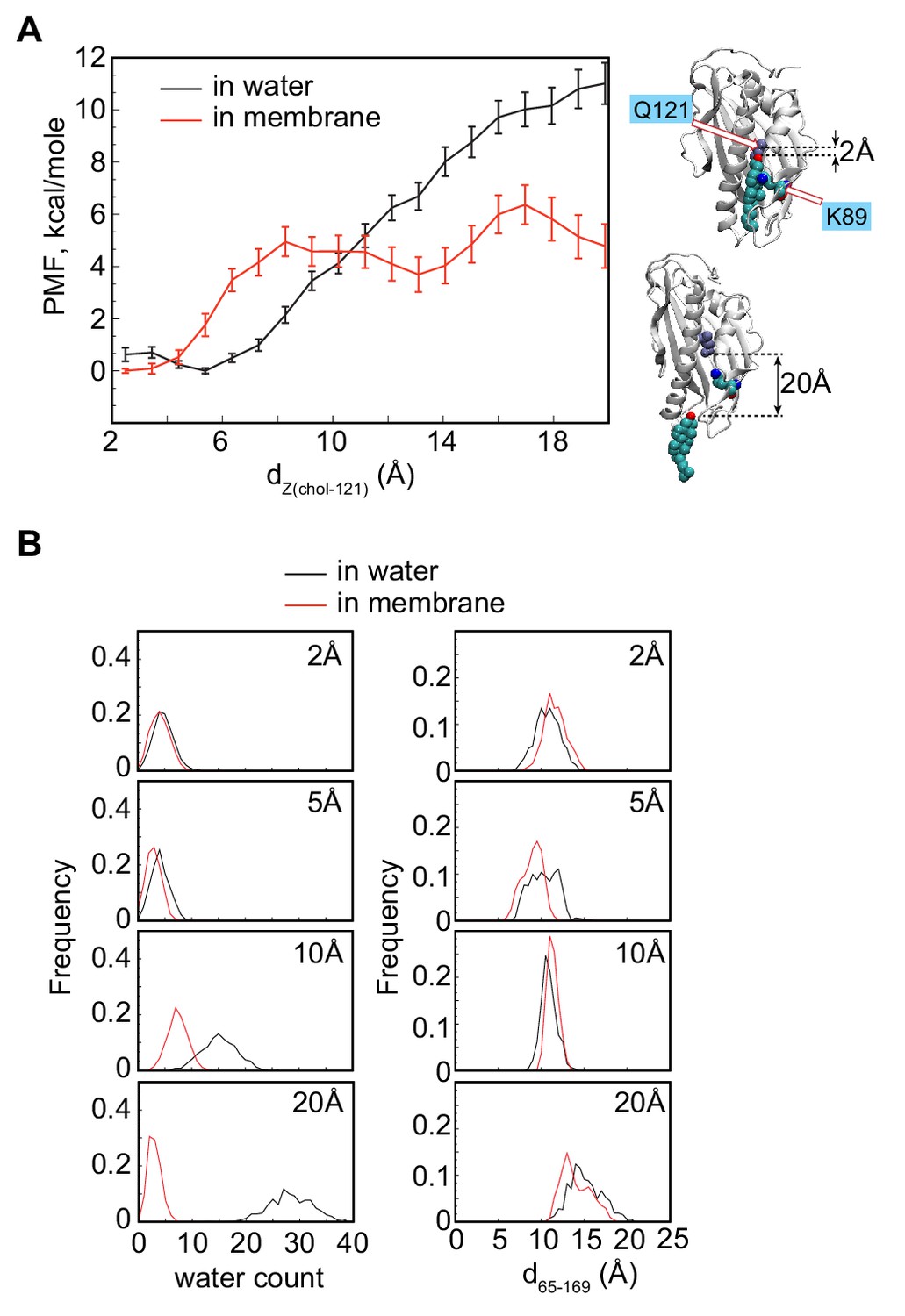
Potential of mean force for sterol egress into water versus the membrane.
(A) Potential of mean force (PMF) as a function of dZ(chol–121) distance for the membrane-bound (red) and soluble (black) wild-type Lam4S2 calculated from umbrella sampling MD simulations at each dZ(chol–121). The structural representations on the right side of the panel illustrate locations of cholesterol corresponding to dZ(chol–121) ~2 Å (top) and dZ(chol–121) ~20 Å (bottom). Residues Q121 and K89 in these snapshots are also shown for reference. (B) Graphs of the number of water oxygens within 3 Å of cholesterol (left panels) and of d65–169 distance (right panels) constructed from analysis of trajectories representing various windows in the range of dZ(chol–121)∈ [2 Å–20 Å] from the umbrella MD simulations of the membrane-bound (red) and soluble (black) wild-type Lam4S2 systems.
Figure 4—figure supplement 3
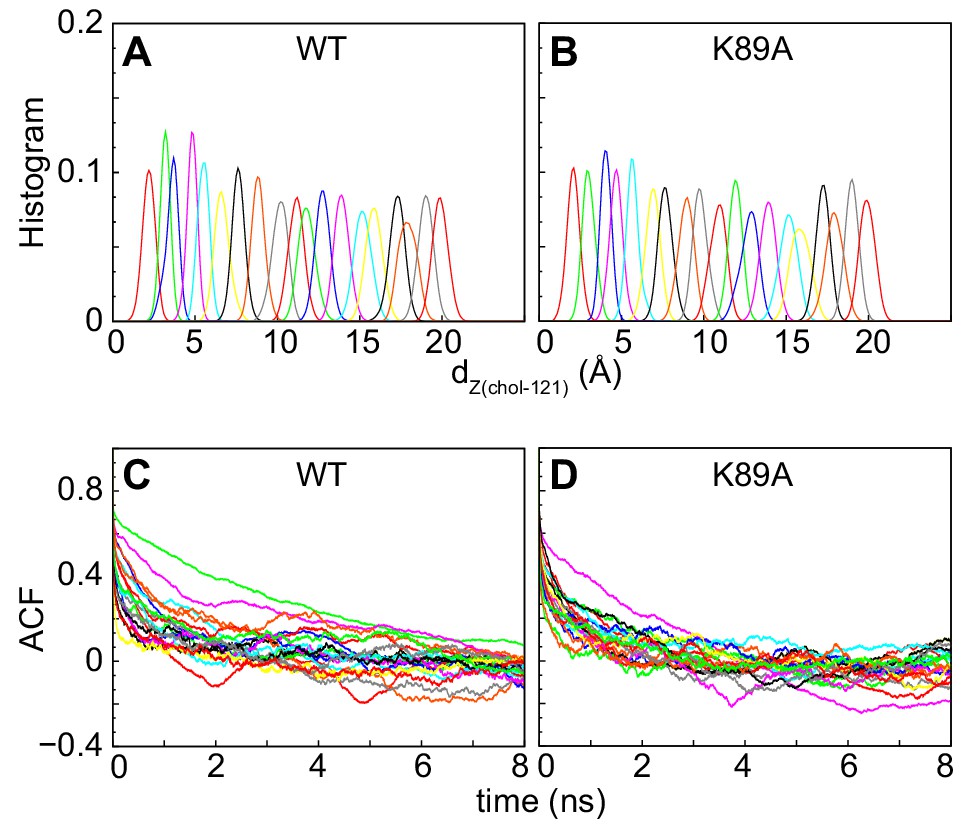
The potential of mean force calculations with the weighted histogram analysis method (WHAM) (Grossfield, 2013).
(A, B) Probability distributions of dZ(chol–121) distance in different windows sampled during the umbrella MD simulations for the wild-type (A) and the K89A mutant (B) systems. The data for the different umbrella windows are represented in different colors. (C, D) Auto-correlation function (ACF) vs. time in different windows sampled during the umbrella MD simulations for the wild-type (C) and the K89A mutant (D) systems. The color-code is the same as in panels (A) and (B).
Figure 5
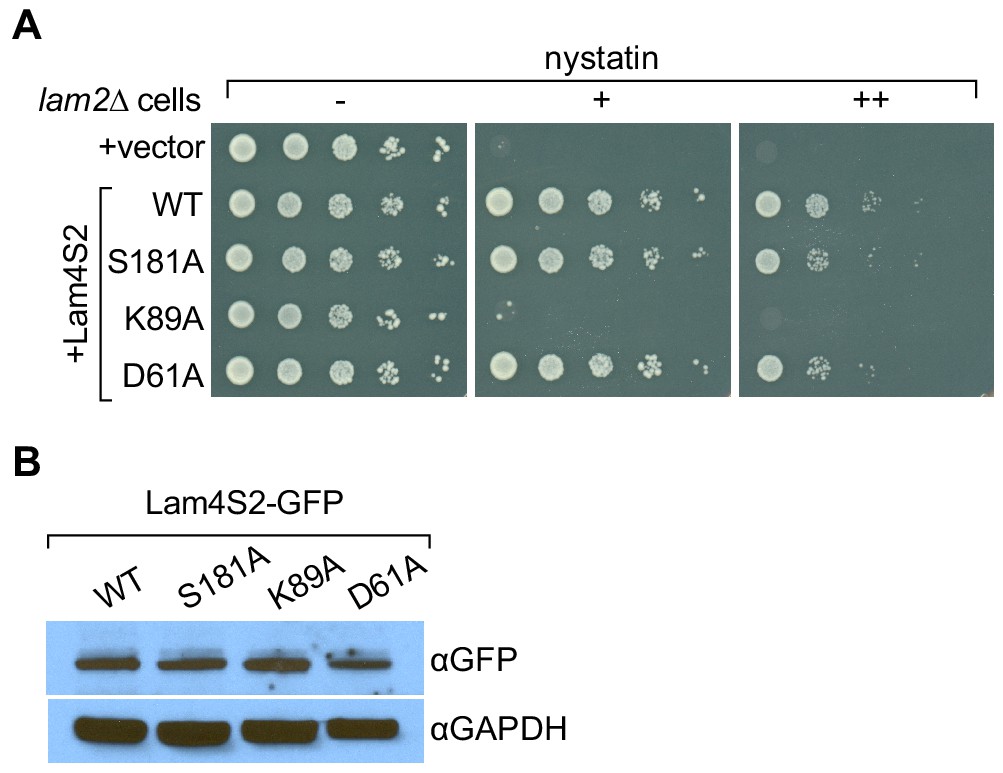
Lam4S2(K89A) does not rescue the nystatin sensitivity of lam2Δ cells.
(A) Cells (lam2Δ) were transformed with an empty vector (top row) or with a vector for the expression of GFP–Lam4S2 wild-type (WT) or point mutants as indicated. Serial 10-fold dilutions were spotted onto agar plates containing defined minimal media (lacking [–] or containing 2 μg/ml [+] or 8 μg/ml [++] nystatin). The plates were photographed after 72 hr at room temperature. (B) Cell-equivalent amounts of cytosol from lam2Δ cells expressing GFP–Lam4S2 wild-type or mutants were analyzed by SDS-PAGE and immunoblotting using anti-GFP antibodies to detect the fusion proteins and anti-GAPDH as a loading control.
Figure 6
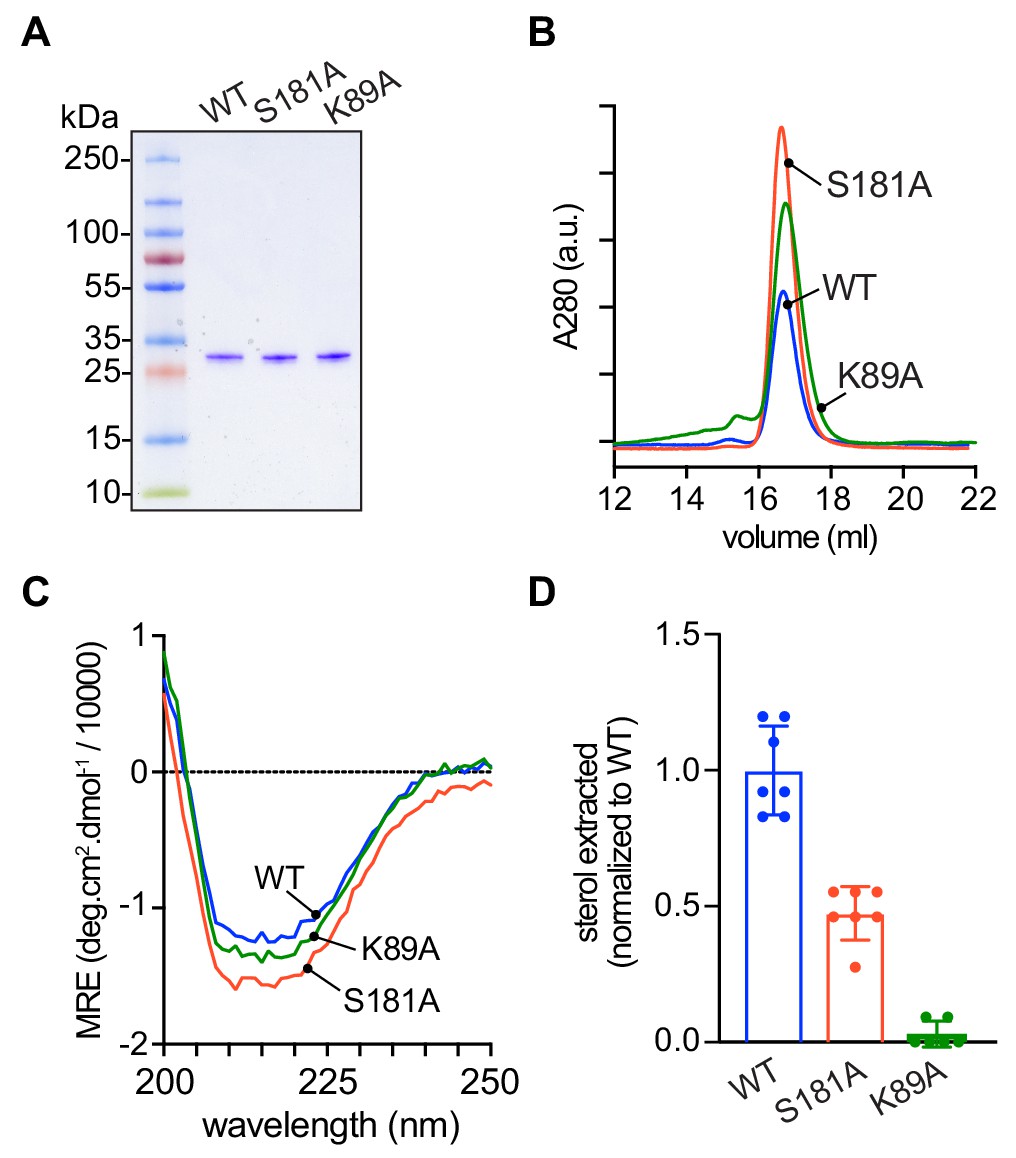
Purification and characterization of Lam4S2 mutants.
(A) Lam4S2 wild-type and the S181A and K89A mutants were purified as His-tagged proteins via affinity chromatography and size-exclusion. The purified proteins were analyzed by SDS-PAGE (4–20% gradient gel) and Coomassie staining. (B) Size-exclusion analysis of purified proteins. (C) Circular dichroism spectra of purified proteins. Protein samples were 12 μM and the spectra shown are the average of three scans per sample. (D) Sterol extraction by purified Lam4S2 and mutants. Sucrose-loaded liposomes (DOPC:DOPE:DOPS:cholesterol, 49:23:23:5 mol %, doped with [3H]cholesterol) were incubated with 750 pmol of purified proteins for 1 hr at room temperature. After ultracentrifugation, the radioactivity and the protein amount in the supernatant were determined, and the stoichiometry of binding was calculated. Data are represented as mean ± SEM (error bars; n = 5–7). Data are normalized to the average value obtained for the wild-type protein (0.11 ± 0.02 pmol cholesterol/pmol protein [mean ± standard deviation (n = 6)]). The ability of the three proteins to interact with membranes was comparable (percentage bound to membranes: 42 ± 7.2 (WT), 43 ± 3.0 (S181A), 49 ± 4.5 (K89A) (mean ± standard deviation [n = 8]).
Figure 7
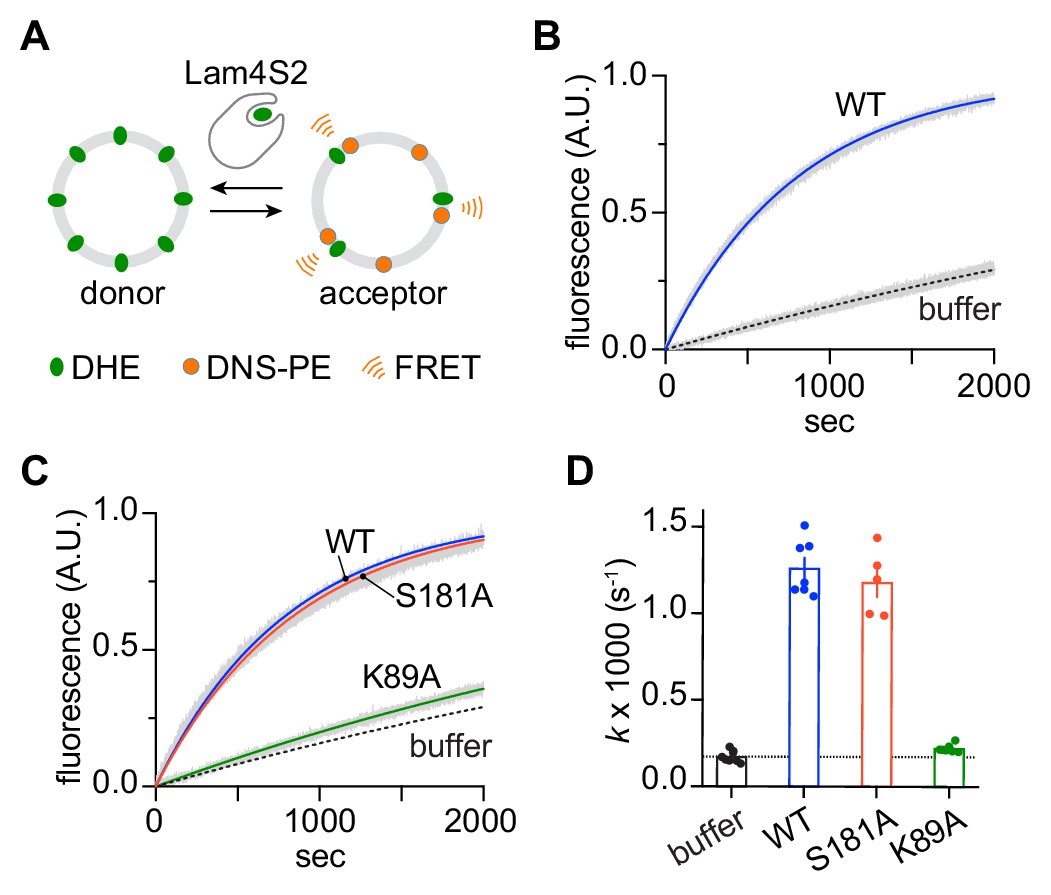
Sterol transfer activity of Lam4S2 mutants.
(A) Schematic of the sterol transport assay. (B) Spontaneous sterol exchange between vesicles, and transport catalyzed by wild-type Lam4S2 (0.05 μM). Traces (n = 7–8) were acquired from three independent experiments and averaged. The blue and dashed lines represent mono-exponential fits of the averaged data; the gray bars graphed behind the fits represent the standard error of the mean (SEM). (C) As in panel (B), except that Lam4S2 mutants were tested (n = 5–7). The data fits for traces corresponding to spontaneous transport and transport catalyzed by the wild-type protein are taken from panel (B) and shown for comparison. (D) Rate constants (colored symbols) obtained from mono-exponential fits of individual traces from the experiments depicted in panels (B) and (C). The bars show the mean and SEM of the data.
Author response image 1
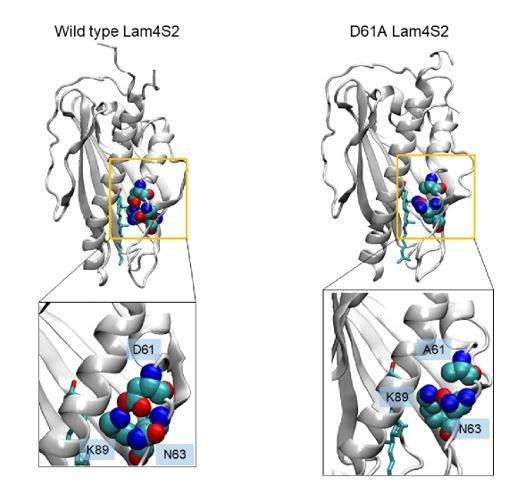
Snapshots of cholesterol-Lam4S2 complex from MD simulations of the wild type protein(left) and the D61A mutant (right), highlighting residue K89 and its interacting partners.
In the wild type system, K89 interacts with D61, whereas in the mutant it interacts with N63.
Videos
Video 1
Molecular dynamics trajectory of cholesterol egress from Lam4S2.
The movie is based on one of the Stage 2 simulations of wild-type Lam4S2. The total length of the trajectory is 350 ns. In the movie, Lam4S2 is shown in white cartoon, the cholesterol molecule is represented in ice-blue colored space-fill, S181, D61, and K89 residues are drawn in space-fill, the oxygen atoms of water molecules in the sterol-binding site are depicted as pink spheres, the membrane leaflet to which Lam4S2 is bound is represented by the nearby lipid phosphate atoms (golden spheres), and lipid molecules within 3 Å of the cholesterol are shown in licorice representation. The rest of the simulation box is omitted. For clarity, the trajectory frames are smoothed for the movie.
Tables
Table 1
Constructs and strains.
| Bacterial plasmids (all constructs are in the pTrcHis6A expression vector and start with the sequence MGGSHHHHHHGMASHHHHHARALEVLFQGPM) | |
|---|---|
| Lam4S21 | Lam4 946–1145 |
| Lam4S2(D61A)3 | Lam4 946–1145 (D1003A) |
| Lam4S2(K89A)3 | Lam4 946–1145 (K1031A) |
| Lam4S2(S181A)3 | Lam4 946–1145 (S1123A) |
| Yeast plasmids | |
| GFP only2 | pRS416 (CEN URA3): GFP + GFP |
| GFP-Lam4S22 | pRS416 (CEN URA3): GFP + Lam4 946–1155 + DV4 |
| GFP-Lam4S2(D61A)3 | pRS416 (CEN URA3): GFP + Lam4 946–1155 (D1003A) + DV4 |
| GFP-Lam4S2(K89A)3 | pRS416 (CEN URA3): GFP + Lam4 946–1155 (K1031A) + DV4 |
| GFP-Lam4S2(S181A)3 | pRS416 (CEN URA3): GFP + Lam4 946–1155 (S1123A) + DV4 |
| Bacterial strain | |
| E. cloni EXPRESS BL21(DE3) | F– ompT hsdSB (rB– mB–) gal dcm lon λ(DE3 [lacI lacUV5-T7 gene one ind1 sam7 nin5]) |
| Yeast strain | |
| lam2Δ (also called ysp2Δ)5 | MATa his3∆1 leu2∆0 met15∆0 ura3∆0 ysp2∆::hphNT1 |
-
1 Described in Jentsch et al. (2018).
2 Described in Gatta et al. (2015).
-
3 Parentheses indicate point mutations, for example K89A, using the Lam4S2 numbering system of Jentsch et al. (2018); residue numbering based on the entire Lam4 sequence is provided in the right-hand column.
4Two amino acids (DV) appended to the end of the Lam4S2 sequence.
-
5Described in Roelants et al. (2018).
Table 2
Primers used for mutagenesis.
| Mutation | Forward primer | Reverse primer |
|---|---|---|
| D61A | CAGAAAGTTATCACTAGAGCTAAGAATAATGTCAATGTGG | CCACATTGACATTATTCTTAGCTCTAGTGATAACTTTCTG |
| K89A | CACTATGAGTACACGGCGAAATTGAACAATTCTATC | GATAGAATTGTTCAATTTCGCCGTGTACTCATAGTG |
| S181A | GAGGGTCAGAAGGTTGCTGTCGATTACATGCTA | TAGCATGTAATCGACAGCAACCTTCTGACCCTC |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Exchange of water for sterol underlies sterol egress from a StARkin domain
eLife 8:e53444.
https://doi.org/10.7554/eLife.53444




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}