Correction: Fgf-signaling is compartmentalized within the mesenchyme and controls proliferation during salamander limb development
Main text
Purushothaman S, Elewa A, Seifert AW. 2019. Fgf-signaling is compartmentalized within the mesenchyme and controls proliferation during salamander limb development. eLife 8:e48507. doi: 10.7554/eLife.48507.
Published 20, September 2019
In the original version of our manuscript, we used a previously published scRNA-seq dataset (Gerber et al., 2018; PMID: 30262634, doi:10.1126/science.aaq0681). After contacting the authors of the published dataset to clarify an ambiguity in their Supplementary Materials, we discovered that we treated the counts in their scRNA-seq dataset as raw counts when in fact they were already normalized and logged (log2(TPM)). Thus, the analysis we published used values that were normalized twice. After re-analysing the data and comparing the outcome to our original data we found that it did not affect any of our conclusions. However, the re-analysed data does slightly affect the visual representation of data presented in Figure 4 and in the two associated Suppl. figures.
The corrected Figure 4 is shown here:
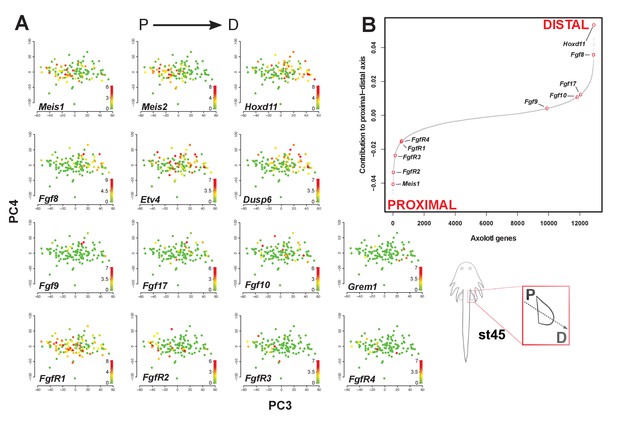
The originally published Figure 4 is shown here for reference:
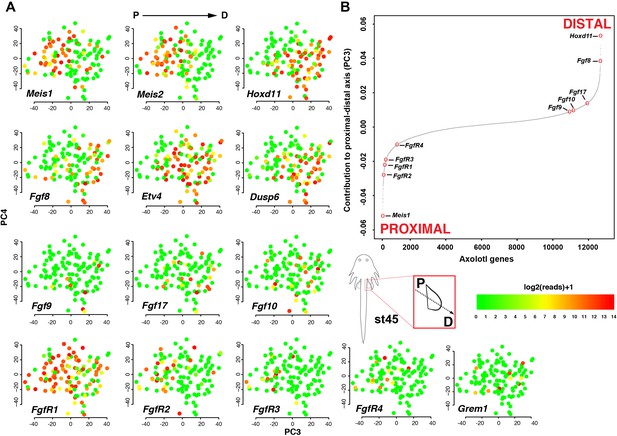
The original figure legend for Figure 4 has changed slightly (changes are underlined).
The sigmoidal curve follows a normal distribution (Shapiro-Wilks normality test, W = 0.95551, p<2.2e-16).
To
The sigmoidal curve follows a normal distribution (Shapiro-Wilks normality test, W = 0.96251, p<2.2e-16). Color scale legends reflect log2(TPM) values.
The corrected Figure 4 – figure supplement 1 is shown here:
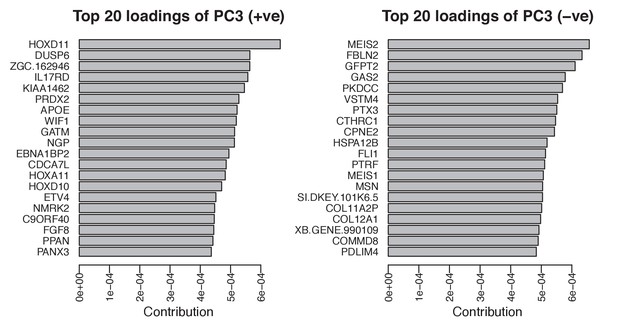
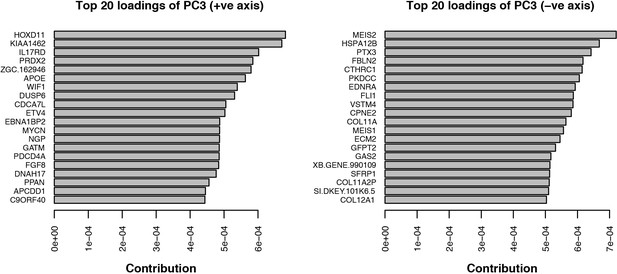
The originally published Figure 4 – figure supplement 1 is shown here for reference:
The corrected Figure 4 – figure supplement 2 is shown here:
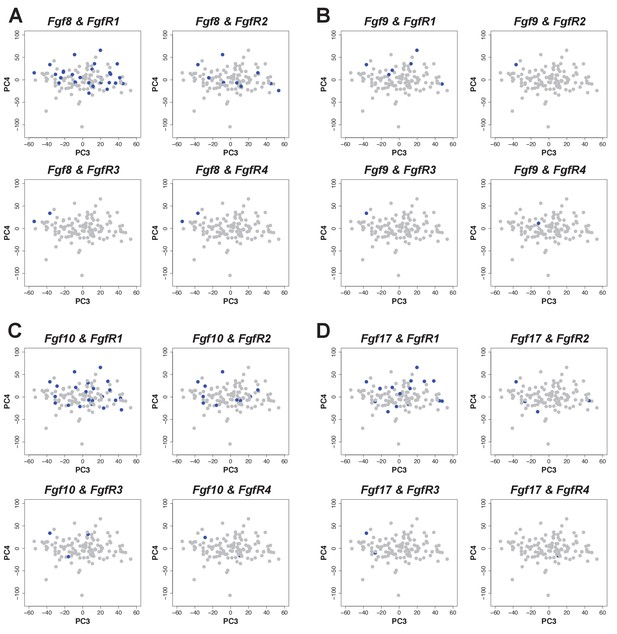
The originally published Figure 4 – figure supplement 2 is shown here for reference:
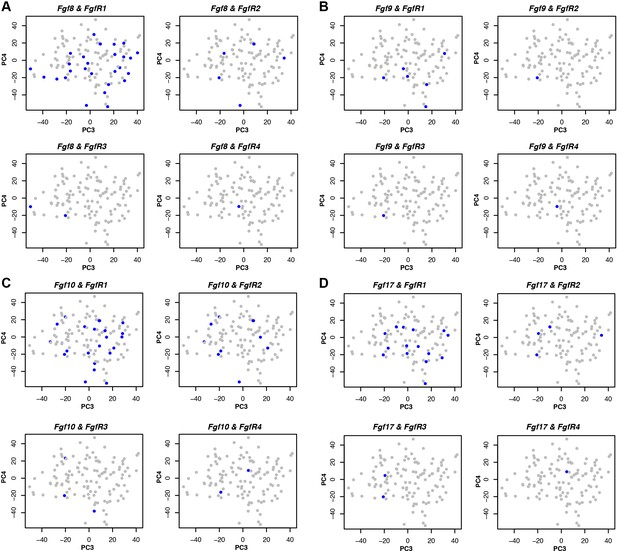
The original figure legend for Figure 4 – figure supplement 2 has changed slightly (changes are underlined).
Expression was defined as one or more RNA-seq reads mapped to the gene in question.
To:
Expression was defined as one or more transcripts per million.
In addition to the figures, a T and p statistic in the Main text for the Welch Two Sample t-test changed from: T = −4.1588, p = 0.0038, TO: T = -3.8104, p = 0.0064
Lastly, we have updated our methods section to reflect the single normalization.
Single-cell RNA-seq data from embryonic limb buds (Gerber et al., 2018, Table S7) were analyzed as follows: read counts from stages 40 and 44 (as reported in Gerber), and which correspond to our stages 44 and 45 (Prayag Murawala, personal communication) were normalized and used as input for principle component analysis. The top twenty loadings of the positive and negative axes of the first five principle components were inspected to identify principle components that segregated developmental axes markers on separate ends. No reliable anterior-posterior, or dorsal-ventral markers segregated in this manner in the current dataset. However, proximal-distal markers (Meis2 and Hoxd11) were segregated on opposite ends of principle component 3 in the PCA of cells from stage 44, but not 40. Due to the low expression levels of most genes of interest, raw read counts were used for visualizing gene expression levels on PCA plots by rescaling the log2 of the read count to span between 0 and 14. To determine co-expression of Fgf ligands and receptors, expression was defined as the presence of one or more reads mapping to the gene in question.
To:
Analysis of single-cell RNA-seq data Single-cell RNA-seq data from embryonic limb buds (Gerber et al., 2018, Table S7) were analyzed as follows: Gene expression (log2(TPM)) from stages 40 and 44 (as reported in Gerber), and which correspond to our stages 44 and 45 (Prayag Murawala, personal communication) were used as input for principal component analysis. The top twenty loadings of the positive and negative axes of the first five principal components were inspected to identify principal components that segregated developmental axes markers on separate ends. No reliable anterior-posterior, or dorsal-ventral markers segregated in this manner in the current dataset. However, proximal-distal markers (Meis2 and Hoxd11) were segregated on opposite ends of principle component three in the PCA of cells from stage 44, but not 40. In order to orient PC3 so that Hoxd11 expression is on the right and Meis2 on the left (i.e., conventional proximal-distal orientation), PC3 values were multiplied by -1. To determine co-expression of Fgf ligands and receptors, expression was defined as the presence of one or more transcript per million.
Also, the methods listed the incorrect concentration of EdU.
Larvae were additionally treated with 0.1 ug/ml of EdU at stage 46 for 24 hr
To:
Larvae were additionally treated with 0.1 mg/ml of EdU at stage 46 for 24 hr
The article has been corrected accordingly.
Article and author information
Author details
Sruthi Purushothaman

Version history
Copyright
© 2021, Purushothaman et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 416
- views
-
- 2
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 2
- citations for umbrella DOI https://doi.org/10.7554/eLife.72022
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Correction: Fgf-signaling is compartmentalized within the mesenchyme and controls proliferation during salamander limb development
eLife 10:e72022.
https://doi.org/10.7554/eLife.72022



