Rif2 protects Rap1-depleted telomeres from MRX-mediated degradation in Saccharomyces cerevisiae
- European Research Institute for the Biology of Ageing, University Medical Center Groningen, Netherlands
- Department of Biology, Lund University, Sweden
Figures
Figure 1 with 1 supplement
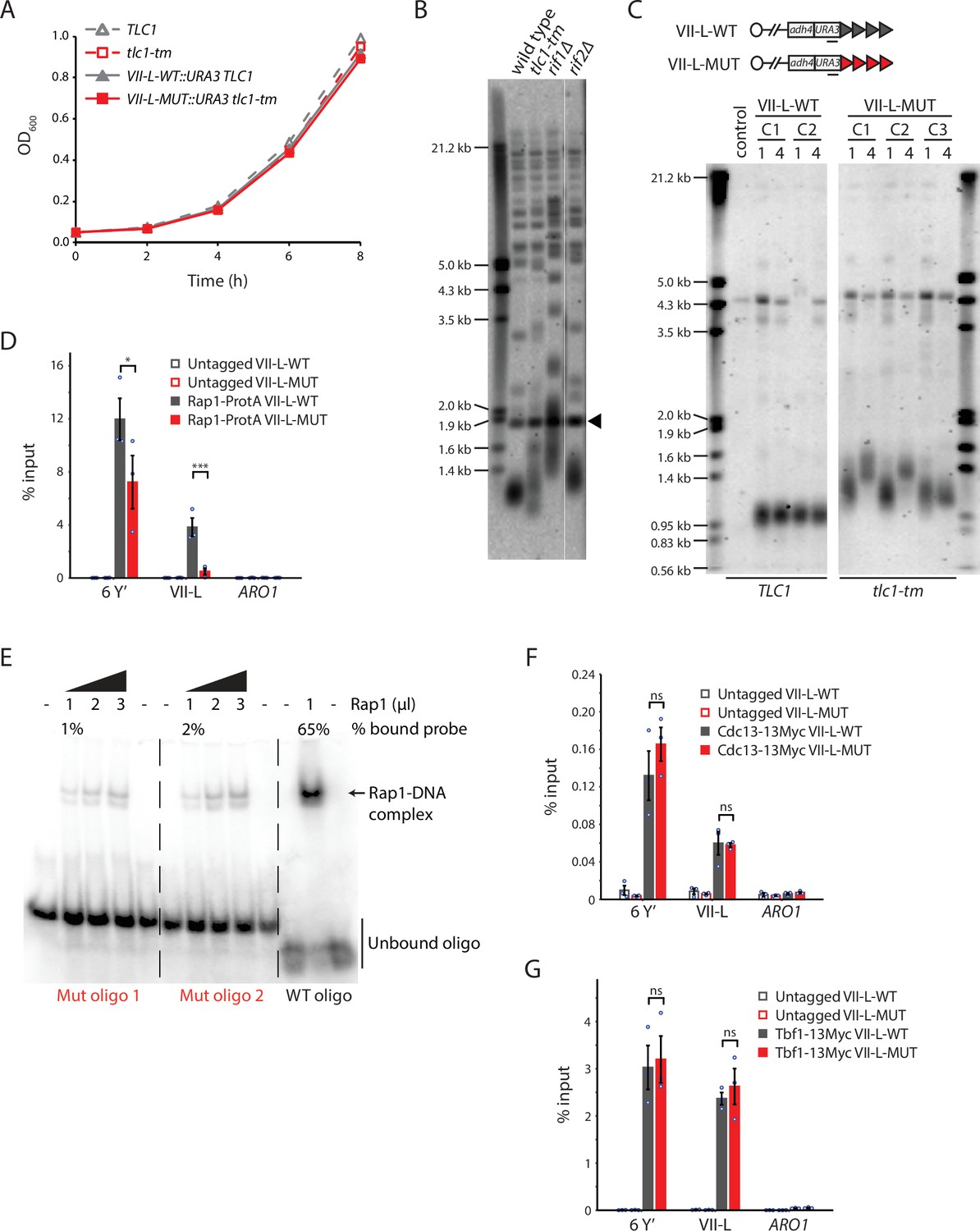
Rap1 binds poorly to tlc1-tm telomere sequences.
(A) Exponentially growing cells of the indicated genotypes were diluted to an OD600 of 0.05 and monitored for 8 hr. Three clones of each genotype were followed, and the average of the measurements at each time point is plotted. Error bars are too small to be visualized. (B) Telomere Southern blot analysis of strains of the indicated genotypes. Black arrowhead indicates a 1.8 kb DNA fragment generated from the BsmAI-digestion of plasmid pYt103 (Shampay et al., 1984). The major terminal restriction fragment is below the 1.8 kb control band. (C) Southern blot analysis of the artificial VII-L telomere, with either wild-type or tlc1-tm mutant sequence, using a probe to the adjacent URA3 gene. Multiple clones were examined, with each clone propagated for 1–4 passages (each passage corresponds to approximately 25 generations). A wild-type strain (lacking the artificial VII-L telomere) was used as a control. (D, F, G) Chromatin immunoprecipitation coupled with quantitative PCR (ChIP-qPCR) analysis of the association of (D) protein A-tagged Rap1, (F) Myc-tagged Cdc13, and (G) Myc-tagged Tbf1 to six Y′ telomeres, the VII-L telomere, or to the non-telomeric ARO1 locus. Untagged wild-type and tlc1-tm strains were used as controls. The mean percentage of input ± SEM is shown (n = 3, *p<0.05, ***p<10–3). Source data are given in Figure 1—source data 1. Tbf1-binding motifs adjacent to the artificial VII-L telomere are shown in Figure 1—figure supplement 1. (E) Electrophoretic mobility shift assay (EMSA) of Rap1 protein incubated with radiolabeled oligonucleotides with either wild-type or tlc1-tm mutant telomeric sequence. The percentage of bound probe was determined by dividing the signal of the shifted band by the total signal (shifted plus unshifted). Uncropped blots for panels (B), (C), and (E) can be found in Figure 1—source data 2.
-
Figure 1—source data 1
Chromatin immunoprecipitation.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig1-data1-v2.xlsx
-
Figure 1—source data 2
Southern blots and EMSA.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig1-data2-v2.zip
Figure 1—figure supplement 1

Tbf1-binding motifs adjacent to the artificial VII-L telomere.
Figure 2
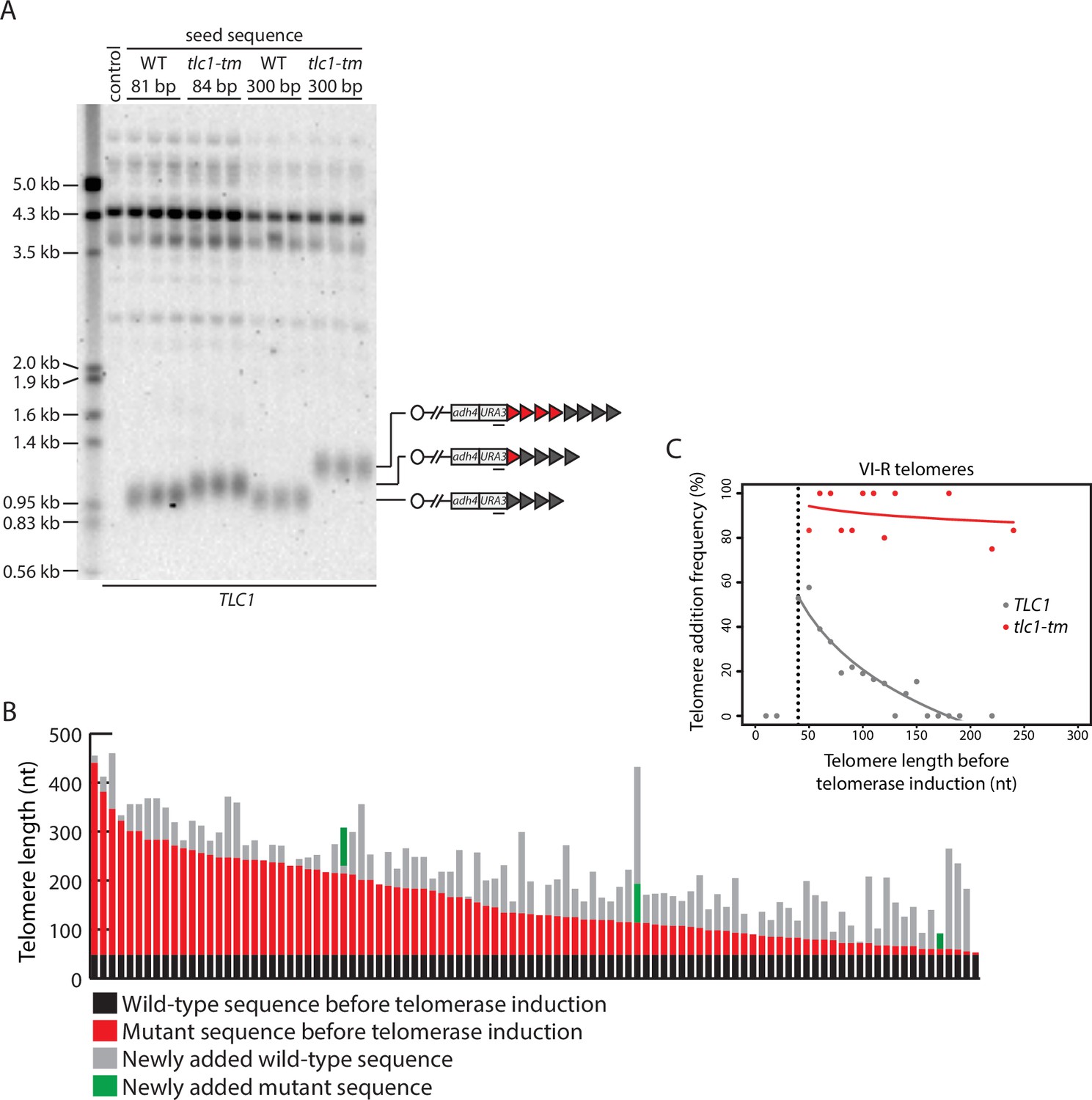
Telomere length regulation is disrupted in tlc1-tm cells.
(A) Southern blot analysis of the artificial VII-L telomere using a probe to the adjacent URA3 gene. The telomere was seeded with either wild-type or tlc1-tm mutant sequence of the indicated lengths in a strain expressing wild-type TLC1. Multiple clones of each strain were examined. A wild-type strain (lacking the artificial VII-L telomere) was used as a control. The uncropped blot can be found in Figure 2—source data 1. (B) In vivo extension of tlc1-tm telomeres was examined using the inducible Single Telomere EXtension (iSTEX) assay. Telomere VI-R was amplified and sequenced after the induction of wild-type telomerase. Each bar represents an individual telomere. The black and red portions of each bar represent wild-type and tlc1-tm sequence, respectively, that is identical in sequence and thus present before telomerase induction. The length of the black/wild-type sequence is 48 bp. Sequence that is divergent from the black and red sequence is shown in gray and green. Gray represents newly added wild-type sequence after the induction of telomerase. Green represents divergent tlc1-tm sequence, most likely a result of homologous recombination. Telomeres are sorted based on the length of the undiverged (black plus red) sequence. (C) Telomere VI-R sequences obtained from the iSTEX analysis in (B) were binned into groups of 10 nt in size according to telomere length before telomerase induction. iSTEX data for the extension of wild-type telomeres were taken from previous studies (Strecker et al., 2017; Stinus et al., 2017) and included for comparison. Groups containing less than four telomeres were excluded from this analysis. Frequency of extension and average telomere length before telomerase induction were calculated and plotted for each group. Logarithmic regression curves for each dataset were also included in the plot. Telomeres shorter than 40 nt before telomerase induction, which are not efficiently recognized and extended by telomerase (Strecker et al., 2017), were removed from the regression analysis. Source data are given in Figure 2—source data 2.
-
Figure 2—source data 1
Southern blots.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig2-data1-v2.zip
-
Figure 2—source data 2
iSTEX assay.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig2-data2-v2.xlsx
Figure 3
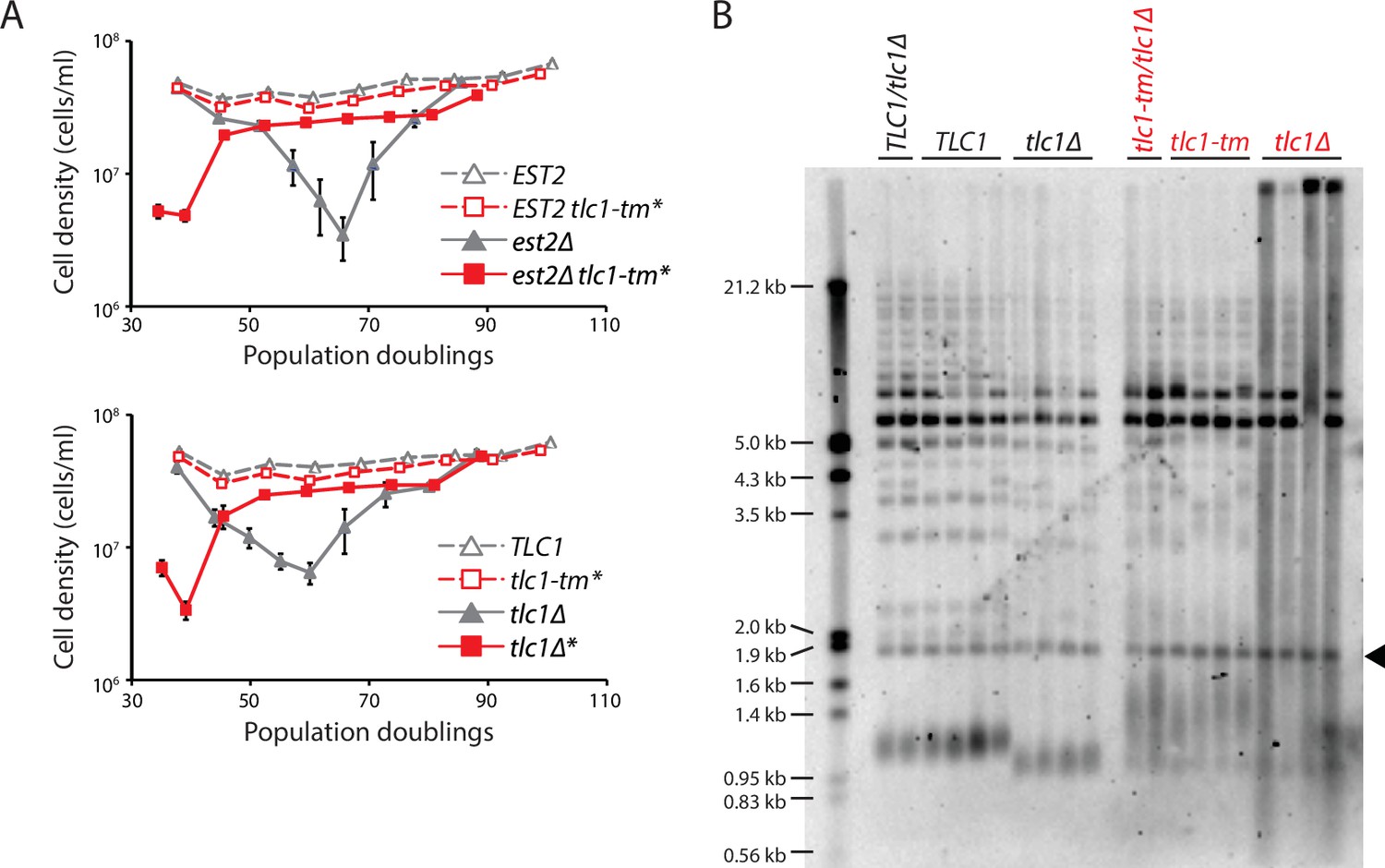
Fast degradation of tlc1-tm telomeres in the absence of telomerase.
(A) Senescence rates were measured by serial passaging of strains of the indicated genotypes, derived from the sporulation of est2Δ/EST2 and est2Δ/EST2 tlc1-tm/tlc1-tm diploids (top panel) or tlc1Δ/TLC1 and tlc1Δ/tlc1-tm diploids (bottom panel). Cell density was measured every 24 hr, followed by dilution to 1 × 105 cells/ml. Mean ± SEM of four independent isolates per genotype is plotted. (B) Telomere Southern blot analysis of samples obtained at the first time point of the senescence assays in (A). Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. The uncropped blot can be found in Figure 3—source data 1.
-
Figure 3—source data 1
Southern blot.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig3-data1-v2.zip
Figure 4 with 1 supplement
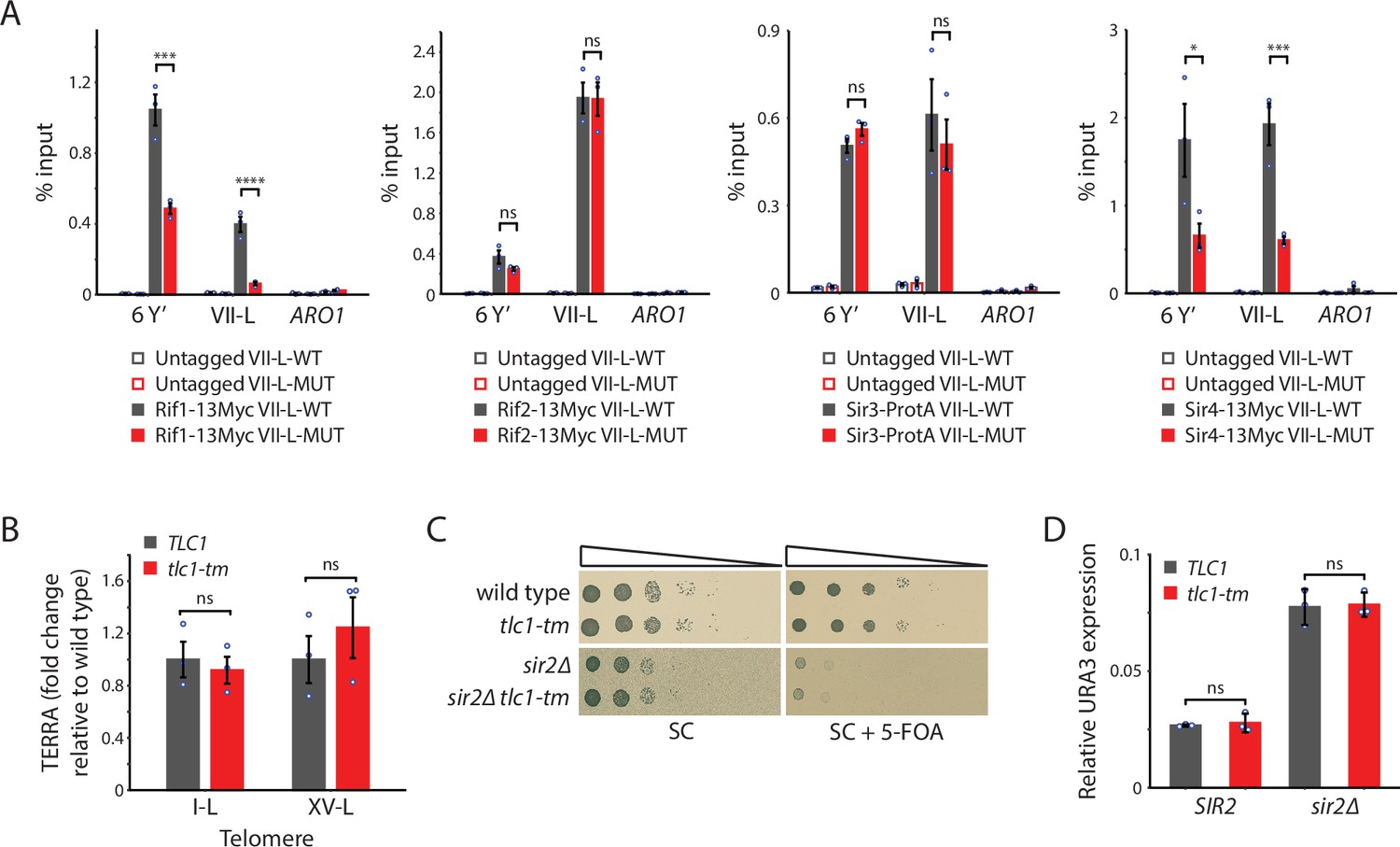
Recruitment of Rif1 and Sir4, but not Rif2 nor Sir3, is significantly reduced at tlc1-tm telomeres.
(A) Chromatin immunoprecipitation coupled with quantitative PCR (ChIP-qPCR) analysis of the association of Myc-tagged Rif1, Rif2, Sir4, and protein A-tagged Sir3 to six Y′ telomeres, the VII-L telomere, or to the non-telomeric ARO1 locus. Untagged wild-type and tlc1-tm strains were used as controls. (B) Total RNA was reverse transcribed and telomeric repeat-containing RNA (TERRA) from specific telomeres (I-L and XV-L) was analyzed by qPCR. TERRA values were normalized to ACT1 levels, and to the respective wild type (TLC1). (C) Tenfold serial dilutions of strains with the indicated genotypes were spotted on SC plates without or with 5-fluoroorotic acid (5-FOA). (D) The expression of the subtelomerically integrated URA3 gene was measured in the indicated yeast strains by RT-qPCR. All data (except C) are shown as mean ± SEM (n = 3, ***p<10–3, ****p<10–4). Source data are given in Figure 4—source data 1.
-
Figure 4—source data 1
Chromatin immunoprecipitation.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig4-data1-v2.xlsx
-
Figure 4—source data 2
Southern blot.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig4-data2-v2.zip
Figure 4—figure supplement 1

Examining the effect of deleting SIR3 in tlc1-tm and tlc1-476A cells.
(A) Telomere Southern blot analysis of strains of the indicated genotypes. Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. (B) Tenfold serial dilutions of strains with the indicated genotypes were spotted on a YPD plate and grown at 30ºC.
Figure 5 with 2 supplements
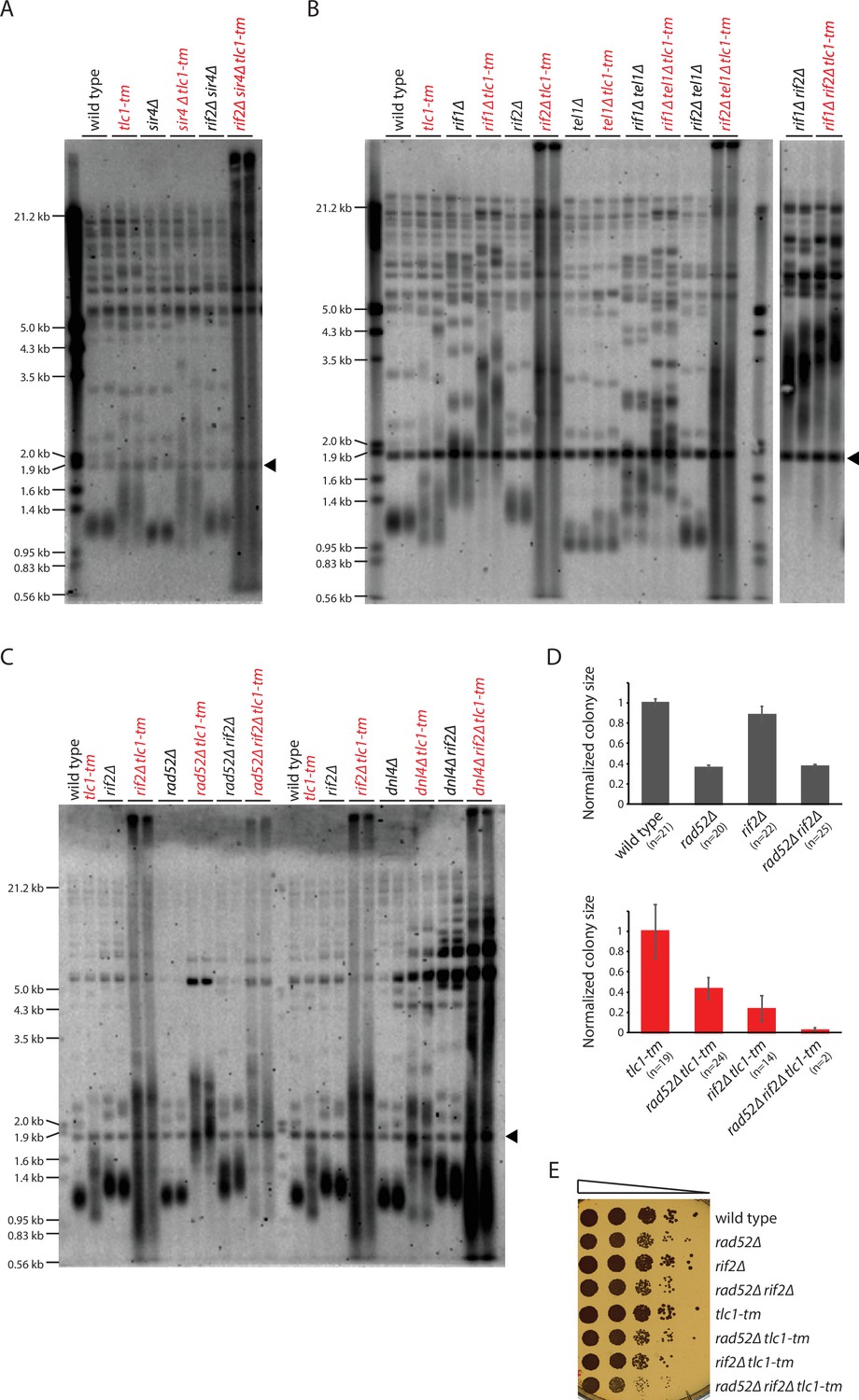
Rif2 prevents degradation of tlc1-tm telomeres.
(A–C) Telomere Southern blot analysis of strains of the indicated genotypes. Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. A Southern blot analysis of the artificial VII-L telomere using a probe to the adjacent URA3 gene in strains from (B) is shown in Figure 5—figure supplement 1. The slight differences in the telomere profile of isogenic tlc1-tm strains are examined further in Figure 5—figure supplement 2. The uncropped blots can be found in Figure 5—source data 1. (D) Colony sizes of haploid meiotic progeny derived from the sporulation of rad52∆/RAD52 rif2∆/RIF2 and rad52∆/RAD52 rif2∆/RIF2 tlc1-tm/tlc1-tm diploid strains were measured and normalized to wild-type (top panel) or tlc1-tm (bottom panel). The number of colonies analyzed is indicated in parenthesis. Error bars show SEM. (E) Tenfold serial dilutions of strains with the indicated genotypes were spotted on a YPD plate and grown at 30°C.
-
Figure 5—source data 1
Southern blot.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig5-data1-v2.zip
Figure 5—figure supplement 1

Rif2 protects tlc1-tm telomeres from degradation.
Southern blot analysis of the artificial VII-L telomere using a probe to the adjacent URA3 gene in strains of the indicated genotypes.
Figure 5—figure supplement 2
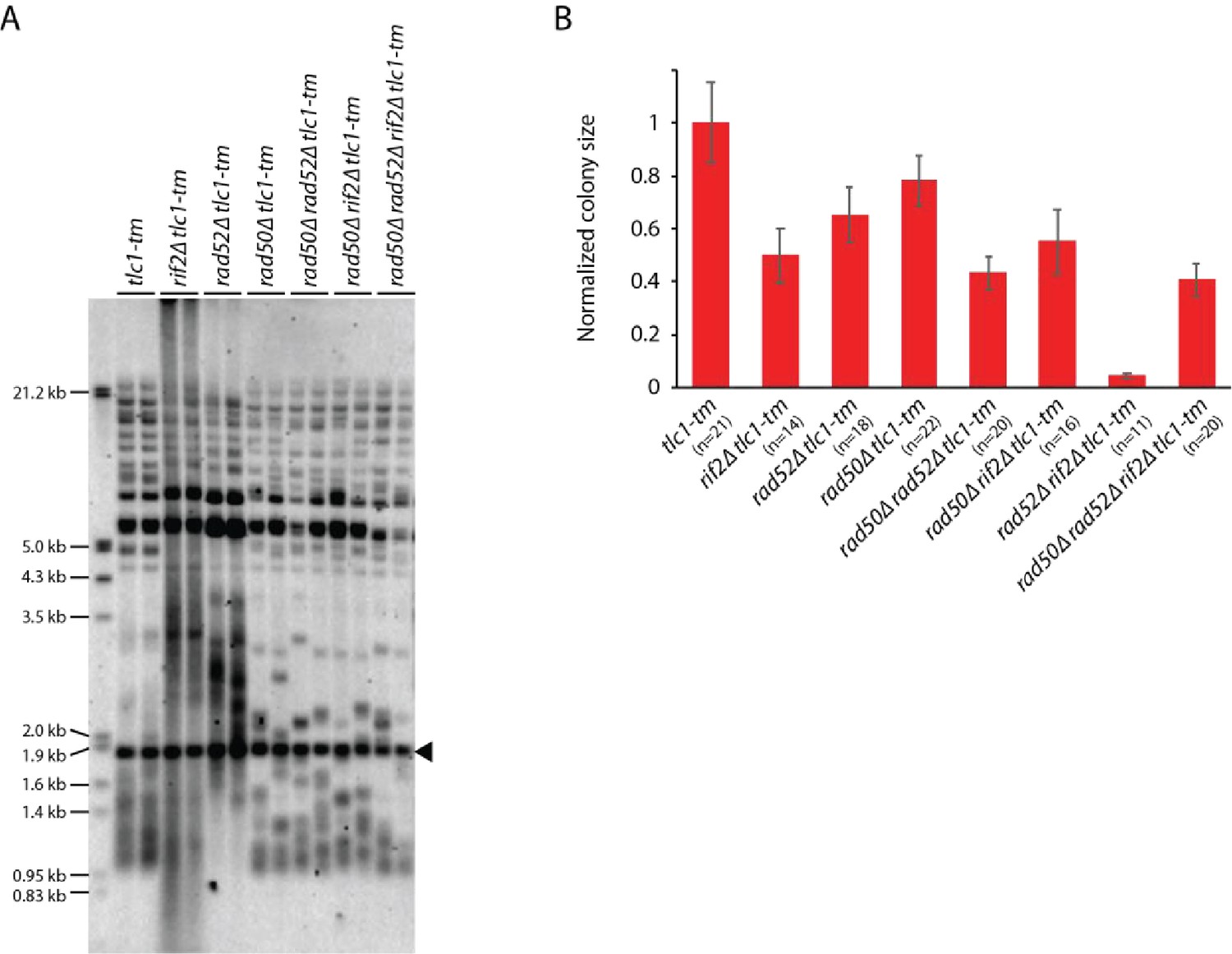
Slight differences in the telomere profile of isogenic tlc1-tm strains are not due to Rad52-dependent homologous recombination.
(A) Telomere Southern blot analysis of strains of the indicated genotypes (duplicate isolates per strain). Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. (B) Colony sizes of haploid meiotic progeny derived from the sporulation of a rad50∆/RAD50 rad52∆/RAD52 rif2∆/RIF2 tlc1-tm/tlc1-tm diploid strain were measured and normalized to tlc1-tm. The number of colonies analyzed is indicated in parenthesis. Error bars show SEM.
Figure 6 with 1 supplement
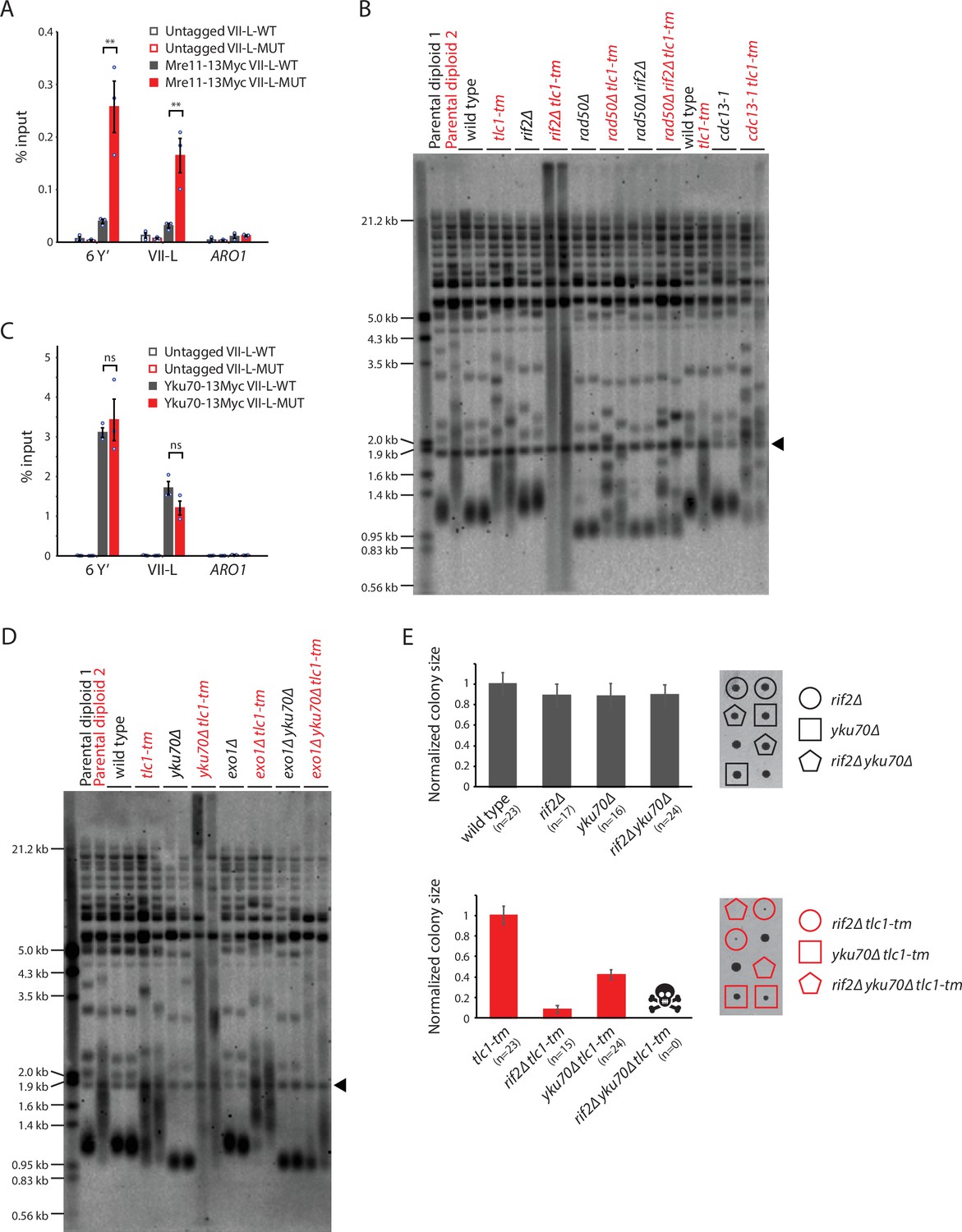
Rif2 protects tlc1-tm telomeres by inhibiting the MRX complex and acts in parallel with the Yku complex.
(A, C) Chromatin immunoprecipitation coupled with quantitative PCR (ChIP-qPCR) analysis of the association of Myc-tagged Mre11 and Yku70 to six Y′ telomeres, the VII-L telomere, or to the non-telomeric ARO1 locus. Untagged wild-type and tlc1-tm strains were used as controls. The mean percentage of input ± SEM is shown (n = 3, **p<0.01). Source data are given in Figure 6—source data 1. (B, D) Telomere Southern blot analysis of strains of the indicated genotypes. Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. (B) Parental diploid 1 is rad50∆/RAD50 rif2∆/RIF2; parental diploid 2 is rad50∆/RAD50 rif2∆/RIF2 tlc1-tm/tlc1-tm. The last six lanes are derived from cells grown at 22°C. (D) Parental diploid 1 is exo1∆/EXO1 yku70∆/YKU70; parental diploid 2 is exo1∆/EXO1 yku70∆/YKU70 tlc1-tm/tlc1-tm. The uncropped blots can be found in Figure 6—source data 2. (E) Colony sizes of haploid meiotic progeny derived from the sporulation of rif2∆/RIF2 yku70∆/YKU70 and rif2∆/RIF2 yku70∆/YKU70 tlc1-tm/tlc1-tm diploid strains were measured and normalized to wild-type (top panel) or tlc1-tm (bottom panel). The number of colonies analyzed is indicated in parenthesis. Error bars show SEM. Representative images of dissected tetrads are shown on the right. Each column of colonies arose from a single tetrad.
-
Figure 6—source data 1
Chromatin immunoprecipitation.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig6-data1-v2.xlsx
-
Figure 6—source data 2
Southern blots.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig6-data2-v2.zip
Figure 6—figure supplement 1

Examining the effect of deleting RAD52 in cdc13-1 tlc1-tm.
(A) Telomere Southern blot analysis of strains of the indicated genotypes. Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. (B) Tenfold serial dilutions of strains with the indicated genotypes were spotted on a YPD plate and grown at 22ºC.
Figure 7 with 1 supplement
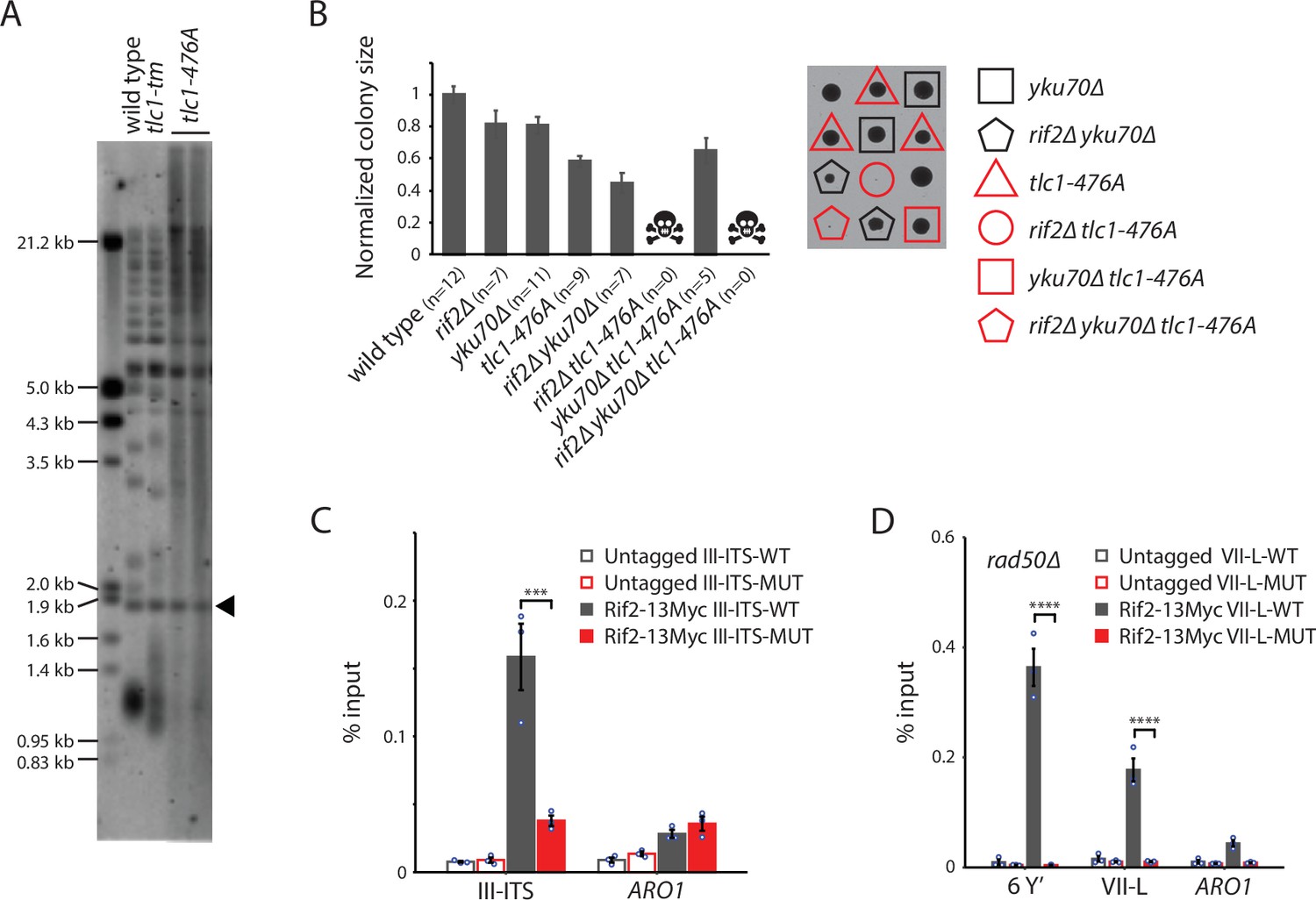
Rif2 association to tlc1-tm telomeres requires the MRX complex.
(A) Telomere Southern blot analysis of strains of the indicated genotypes. Black arrowhead indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B. The uncropped blot can be found in Figure 7—source data 1. (B) Colony sizes of haploid meiotic progeny derived from the sporulation of a rif2∆/RIF2 yku70∆/YKU70 tlc1-476A/TLC1 diploid strain were measured and normalized to wild type. The number of colonies analyzed is indicated in parenthesis. Error bars show SEM. Representative images of dissected tetrads are shown on the right. Each column of colonies arose from a single tetrad. (C) Chromatin immunoprecipitation coupled with quantitative PCR (ChIP-qPCR) analysis of the association of Myc-tagged Rif2 to a 300 bp ITS located on chromosome III containing either wild-type or tlc1-tm sequence. The mean percentage of input ± SEM is shown (n = 3, ***p<10–3). (D) ChIP-qPCR analysis of the association of Myc-tagged Rif2 to six Y′ telomeres, the VII-L telomere, or to the non-telomeric ARO1 locus in a rad50∆ background. Untagged strains were used as controls. The mean percentage of input ± SEM is shown (n = 3, ****p<10–4). Source data for ChIP experiments are given in Figure 7—source data 2.
-
Figure 7—source data 1
Southern blots.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig7-data1-v2.zip
-
Figure 7—source data 2
Chromatin immunoprecipitation.
- https://cdn.elifesciences.org/articles/74090/elife-74090-fig7-data2-v2.xlsx
Figure 7—figure supplement 1

Expression of a Rif2-L44R,V45E,E347R mutant suppresses neither the telomere degradation nor slow growth of rif2∆ tlc1-tm cells.
rif2∆ and rif2∆ tlc1-tm cells were transformed with plasmids pRS315 (vector), pRS315-RIF2, or pRS315-rif2- L44R,V45E,E347R, then passaged for approximately 100 generations before telomere Southern blot analysis (A) and spotting of tenfold serial dilutions on a YPD plate (B). Black arrowhead in A indicates the 1.8 kb telomere sequence-containing fragment loaded as control, as in Figure 1B.
Author response image 1
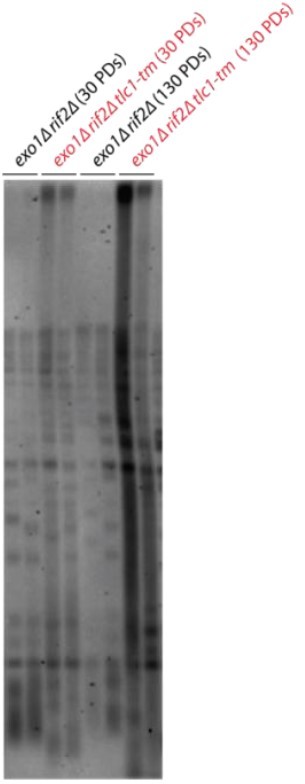
Additional files
-
Supplementary file 1
Supplementary tables.
(A) Yeast strains used in this study. (B) Wild-type and mutant oligo sequences used to replace the VII-L telomere. (C) qPCR primers.
- https://cdn.elifesciences.org/articles/74090/elife-74090-supp1-v2.xlsx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/74090/elife-74090-transrepform1-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Rif2 protects Rap1-depleted telomeres from MRX-mediated degradation in Saccharomyces cerevisiae
eLife 11:e74090.
https://doi.org/10.7554/eLife.74090




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}