Reconstitution of kinetochore motility and microtubule dynamics reveals a role for a kinesin-8 in establishing end-on attachments
- Department of Molecular and Cell Biology, University of California, Berkeley, United States
- Biophysics Graduate Group, University of California, Berkeley, United States
Figures
Figure 1 with 2 supplements

Kinetochore reconstitution and dynamics on the microtubule (a) Schematic of a typical kymograph representing a dynamic kinetochore on a microtubule.
Time (min) is on the x-axis and distance (µm) is on the y-axis. Above the kymograph is a cartoon depiction explaining how the kymograph shows three-dimensional data in two-dimensional space. In both the kymograph and cartoon, the yeast microtubule is red, the kinetochore is green, and the porcine microtubule seed is blue. (b) A schematic of the budding yeast kinetochore with its major sub-complexes shown in different colors. These colors correspond to the different kinetochore proteins assayed in (c). The kinetochore is composed of inner kinetochore proteins such as the Cse4 nucleosome and the CBF3 complex, that bind to DNA, and outer kinetochore proteins such as the Ndc80 and Dam1 complex, that bind to the microtubule. (c) Reconstituted kinetochore proteins associated with yeast microtubules. Representative kymographs of kinetochore proteins from each sub-complex are shown. Time is on the x-axis and distance is on the y-axis (scale bars are 5 min and 5 µm, respectively). Kinetochore proteins are green and yeast microtubules are red. The porcine seed is blue. Each of the proteins assayed displays similar dynamics: moving toward the microtubule plus end when bound to the microtubule lateral surface. When the kinetochore proteins reach the plus end of the microtubule, they appear to convert from lateral to end-on attachment, which is often followed by microtubule depolymerization with the kinetochore remaining associated with the plus end. The microtubules were never observed to grow again following the depolymerization events in which a kinetochore protein was attached end-on. (d) Quantification of velocities and dynamics of kinetochores bound to microtubules. Kinetochores bound to the sides of microtubules move toward the microtubule plus end at an average velocity of 0.56 µm/min. There were negligible minus-end directed movements when the kinetochore was side bound. When the kinetochores were associated with the microtubule tips, they tracked the microtubule tips as they depolymerized, moving at an average velocity of 0.4 µm/min. There were also negligible plus-end directed movements when the kinetochore was bound to the tip. Each data point in the graph is the average velocity of one kinetochore. The error bars are the mean and standard error of the mean from four replicate trials of two biological replicates. Statistical analysis was done by a Kruskal-Wallis test where **** is p<0.0001, *** is p=0.0003, and * is p=0.0132. For the different kinetochore proteins: from Spc105 in order in the figure to Ctf3, N=128, 155, 116, 102, 174, 56, 77, 59, 87 proteins tracked, respectively. (e) When the kinetochore is bound to the side of a microtubule, about 30% of its time it is in plus-end directed motion and the remaining 70% of the time it is paused (not moving). Kinetochores associated with plus ends spend 40% of the time moving toward the minus-end and 60% of the time paused.
-
Figure 1—source data 1
Source data supplied.
- https://cdn.elifesciences.org/articles/78450/elife-78450-fig1-data1-v1.zip
Figure 1—figure supplement 1
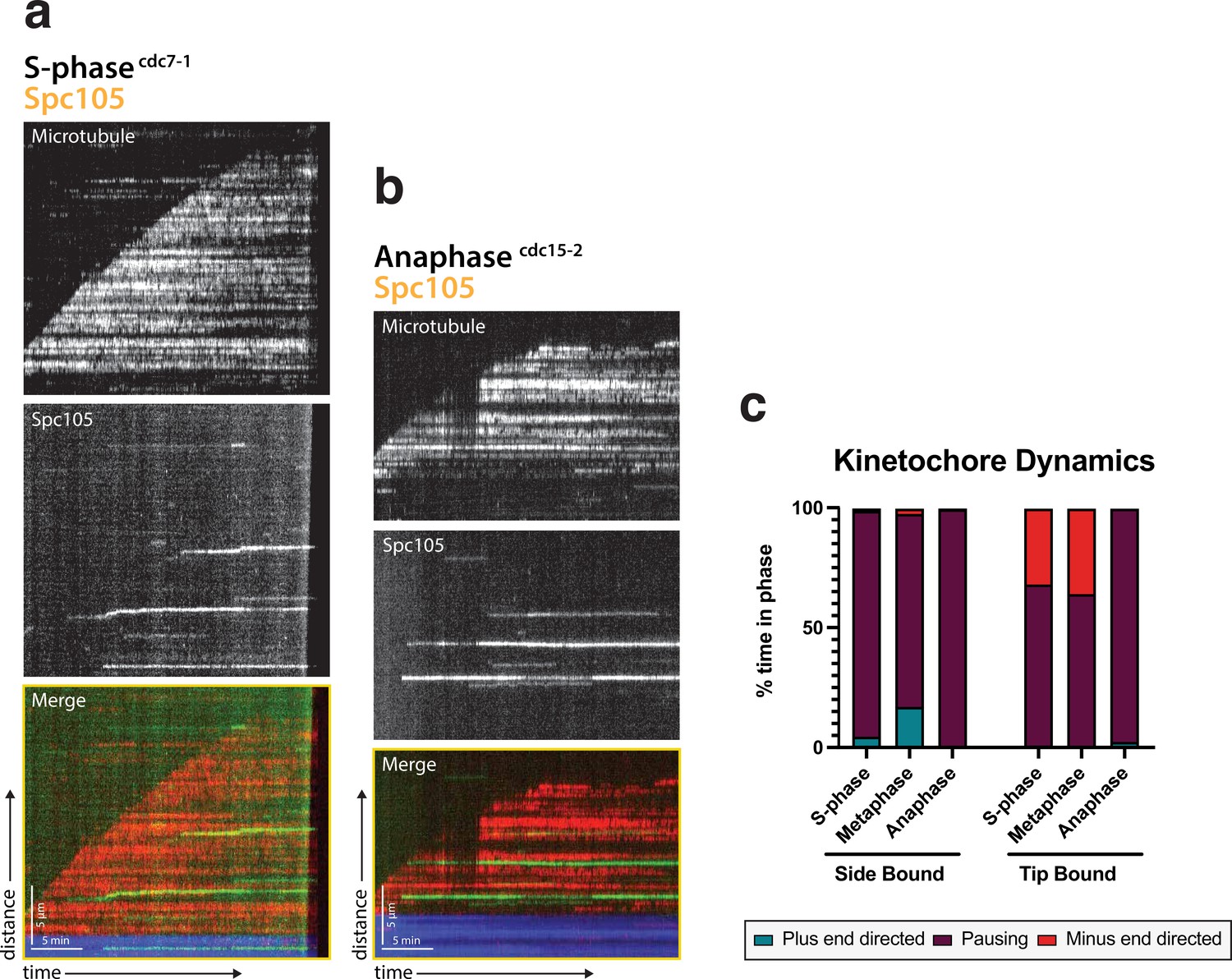
Kinetochore motility is specific to metaphase extracts.
(a) Representative kymograph from a S-phase arrested lysate (cdc7-1) with Spc105 (green) and the microtubule (red). (b) Representative kymograph from an anaphase arrested lysate (cdc15-2) with Spc105 (green) and the microtubule (red). (c) Kinetochore dynamics are shown by percent of time spent in each phase: plus-end directed, paused, or minus-end directed when either side bound or tip bound. S-phase and anaphase arrested kinetochore lysates spend the majority of the time paused on the lattice compared to metaphase.
Figure 1—video 1
Dynamic kinetochores on the microtubule Spc105 (green) moves on the lattice of the microtubule (red) toward the plus-end.
When reaching the tip, it transitions attachment and the microtubule begins to depolymerize, with the kinetochore tip-tracking. Scale bar is 5 µm. Frame rate is 20 frames per second.
Figure 2 with 1 supplement
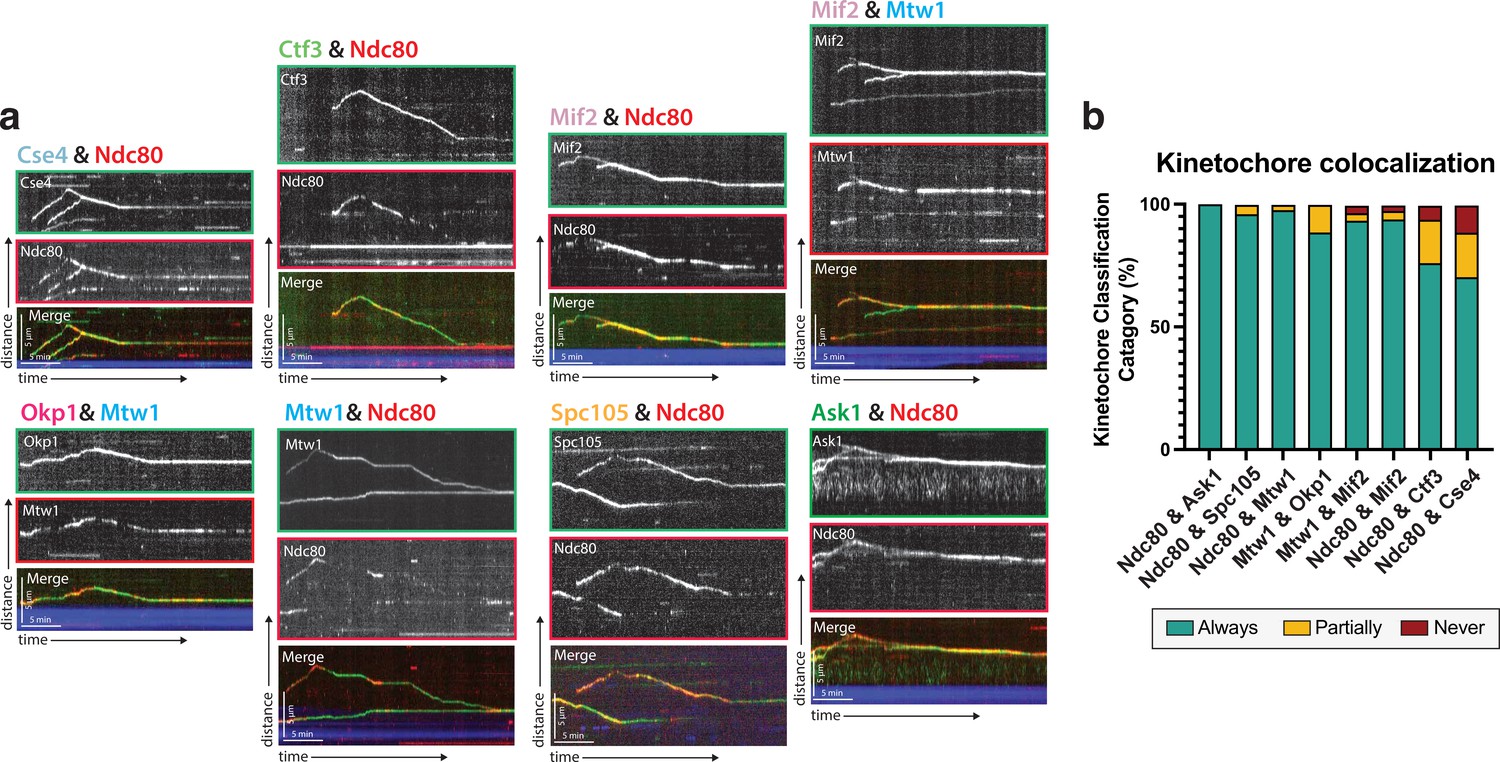
Protein composition of reconstituted kinetochores.
(a) Different pairwise combinations of kinetochore sub-complex proteins colocalize, establishing that the kinetochore spots observed by fluorescence microscopy are multi-protein, multi-subcomplex assemblies. Representative kymographs from each strain expressing a different pair-wise combination of tagged kinetochore proteins are shown. Time is on the x-axis and distance is on the y-axis (scale bars are 5 min and 5 µm, respectively). Ndc80 was used as the common reference marker for multiple pairwise combinations. Additionally, Mtw1 was used in some pairwise combinations to show that multiple sub-complexes are present in the same kinetochore. The tracks of these colocalized proteins follow similar dynamics as in Figure 1, where they move processively toward the microtubule plus end. At a certain point, they reach the plus end and motility becomes minus-end directed concomitant with the onset of microtubule disassembly (note that the microtubules are not visualized due to the inability to measure 4 colors, and the assumption is that this directional switch in kinetochore protein motility corresponds to microtubule depolymerization onset shown in Figure 1). (b) Quantification of kinetochore colocalization for each pair-wise protein combination. Kinetochore tracks were grouped into three distinct pools and counted. These categories were ‘always’ colocalized (blue), ‘partially’ colocalized (yellow), or ‘never’ colocalized (red). For partially colocalized tracks, the mScarlet signal initially appears alone, but a GFP signal appears on it and stays colocalized for the remainder of the movie. For the different kinetochore protein pairs in order in the figure: from Ndc80 and Ask1 in to Ndc80 and Cse4, N=70, 49, 42, 52, 120, 81, 116, 97 tracks categorized, respectively, from two experimental replicates.
Figure 2—video 1
Multiple kinetochore proteins are dynamic on the microtubule together.
Mtw1 (red) colocalizes with Mif2 (green) and they move together in a directional manner. The microtubule is unlabeled due to experimental limitations of four color imaging, but, based on the data acquired when the microtubule could be imaged (Figure 1), kinetochore movement is on the lattice of the microtubule and moving towards the plus-end. When reaching the tip, the kinetochore transitions attachment and the microtubule begins to depolymerize, with the kinetochore tip-tracking. Scale bar is 5 µm. Frame rate is 20 frames per second.
Figure 3 with 2 supplements
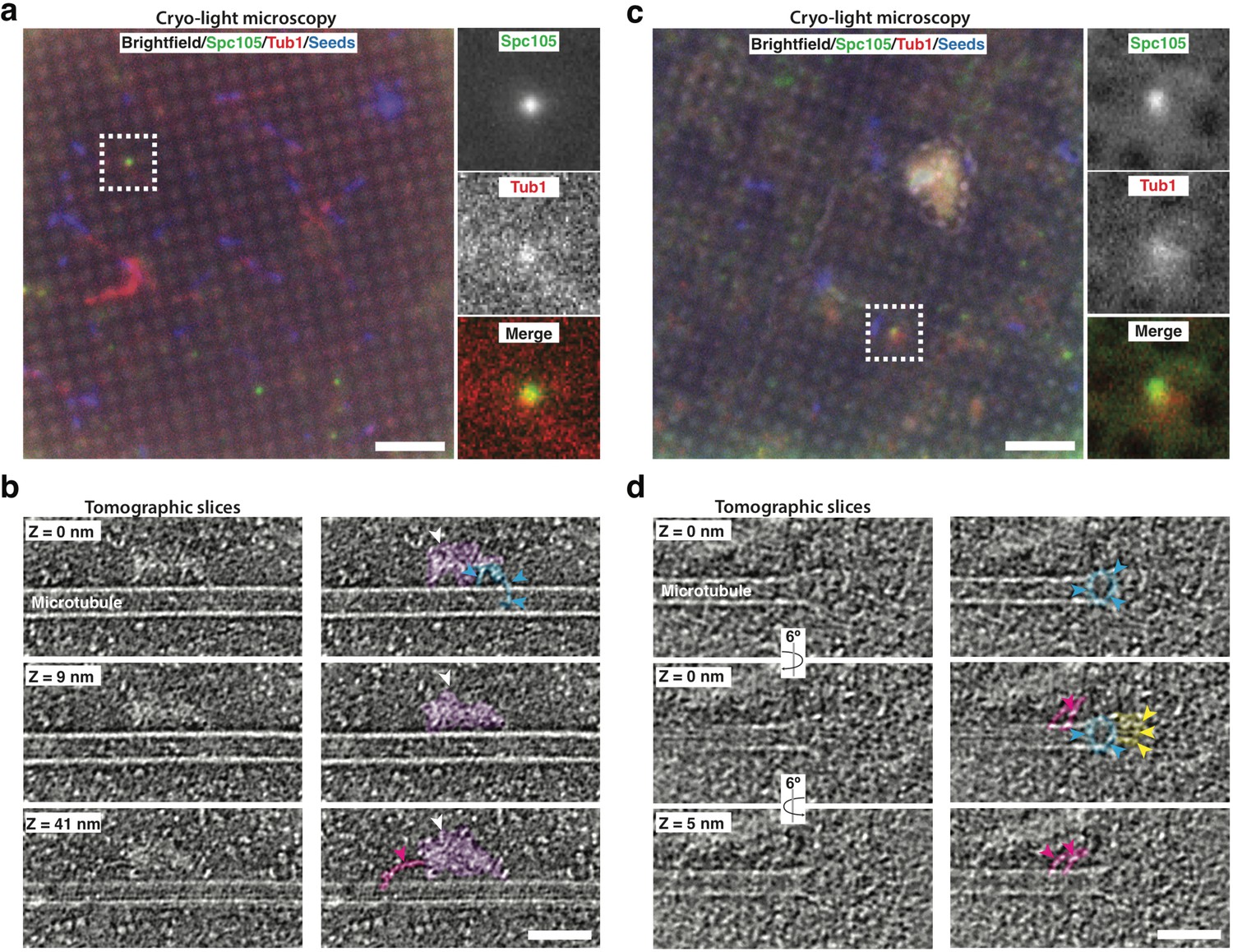
Cryo-correlative light microscopy and electron tomography shows microtubule-associated intact kinetochore complexes.
(a, c) Overlay of brightfield and fluorescent channel cryo-light micrographs showing Spc105-GFP in green, mRuby-Tub1 in red and microtubule seeds in blue. The brightfield channel highlights the holey carbon film pattern. Insets show 2.7x magnified views of boxed regions. (b) Slices of tomogram recorded in the region of the highlighted Spc105 focus in (a) at Z-positions relative to the top slice. Slices show a kinetochore bound to the lateral surface of a microtubule. The largest part of the kinetochore appears as an electron-dense proteinaceous cloud similar to what has been reported for purified yeast kinetochores (white arrowheads; Gonen et al., 2012). A ring-like density (blue arrowheads) and a rod-like density (pink arrowheads), potentially representing Dam1 and Ndc80 sub-complexes, respectively, extend from the cloud-like density. (d) Slices of tomogram recorded in the region of the highlighted Spc105 focus in (c) at Z-positions relative to the top slice, showing a kinetochore complex bound to the end of a microtubule. Arrowheads point at a ring-like (blue) and rod-like (pink) density similar to the ones observed in (b) Additional rod-like extensions (yellow arrowheads) might represent the MIND complex. Right columns in (b,d) show the same images as in the left column, but protein densities are colored to match the arrows. Scale bars, 10 µm in (a, c), 60 nm in (b, d).
Figure 3—video 1
Cryo-electron tomogram of kinetochore bound to the lateral surface of a microtubule.
The largest part of the kinetochore appears as an electron-dense proteinaceous cloud (white arrowheads). A ring-like density (blue arrowheads) and a rod-like density (pink arrowheads), potentially representing Dam1 and Ndc80 sub-complexes, respectively, extend from the cloud-like density. Scale bar, 40 nm. Frame rate 20 frames per second.
Figure 3—video 2
Cryo-electron tomogram of kinetochore bound to the tip of a microtubule.
Blue and pink arrowheads point at a ring-like (blue arrowheads) and rod-like (pink) density similar to the ones observed in Figure 3—video 1. Additional rod-like extensions (white arrowheads) might represent the MIND complex. Scale bar, 40 nm. Frame rate 10 frames per second.
Figure 4 with 1 supplement
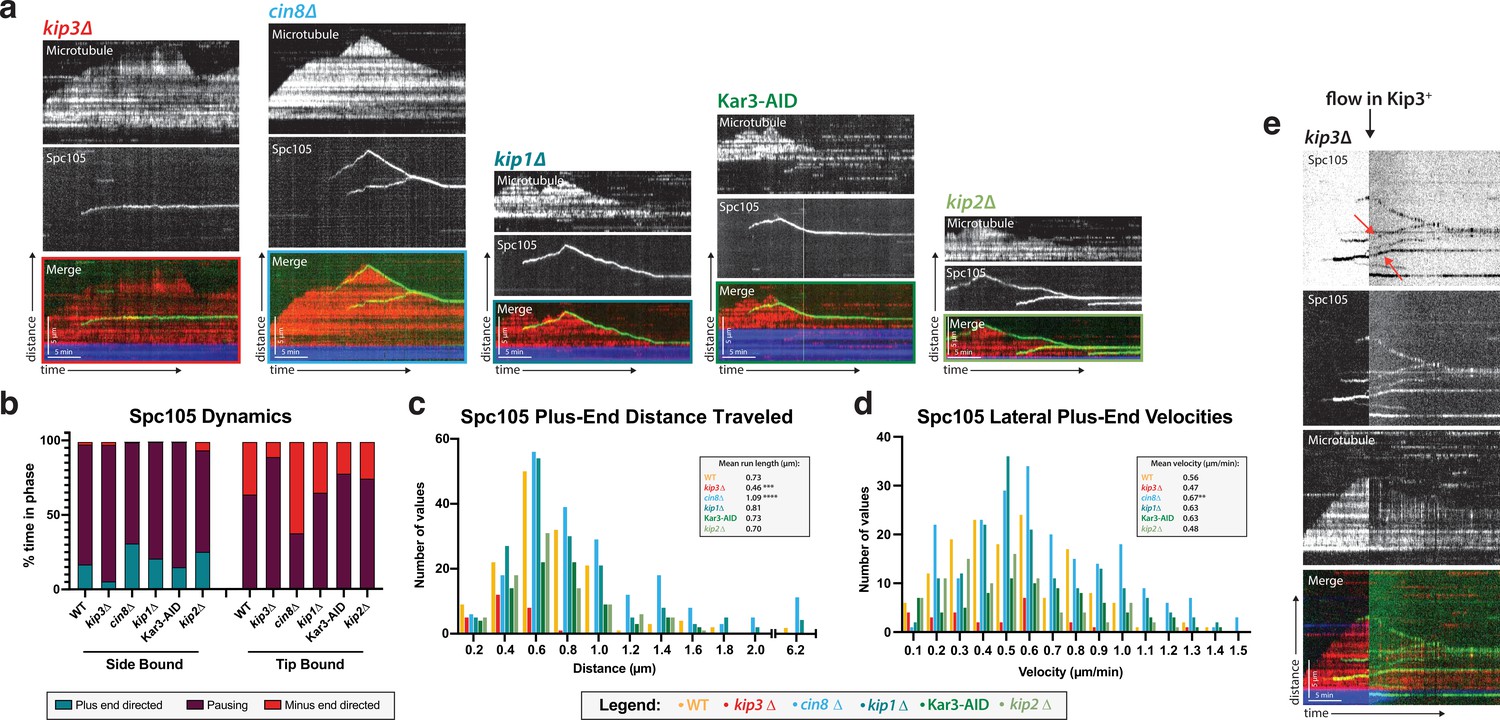
Kip3, a kinesin-8 motor, is required for plus-end directed movement of the kinetochore on the microtubule lattice.
(a) Genes encoding each of the five yeast kinesins were individually deleted (or the proteins degraded using a degron tag) and kinetochore dynamics on microtubules were assayed. Only Kip3, a highly processive kinesin-8, was required for kinetochore movement toward the microtubule plus end. Representative kymographs are shown with time on the x-axis and distance on the y-axis (scale bars are 5 min and 5 µm, respectively). The kinetochore (Spc105) is shown in green and the microtubule is shown in red. The microtubule seed (at the minus-end of the growing red yeast microtubule) is shown in blue. (b) Quantification of dynamics over time shows that kinetochores in the kip3Δ strain spend the majority of the time paused, moving toward the plus end only 5% of the time compared to 30% for wild-type. (c) Run lengths and the number of runs the kinetochore made on the lattice toward the plus end were decreased in the kip3Δ strain. Run lengths were about half of wild-type, 0.46 µm vs 0.73 µm. The number of runs was also decreased to about 25% of the wild-type number of runs. Statistical analysis was done by a Kruskal-Wallis test where *** is p=0.0001 and **** is p<0.0001. (d) Laterally bound kinetochores velocities are unchanged when moving in the plus end direction in a kip3Δ lysate. Statistical analysis was done using a Kruskal-Wallis test where ** is p=0.0047. (b–d) Quantification is from four replicate trials of two biological replicates. For each strain from WT to kip2Δ lysate in the order listed, N=128, 70, 129, 144, 103, 154 Spc105 proteins tracked. (e) Adding back a source of Kip3 to a lysate from a kip3Δ strain restores plus end-directed kinetochore movement. In lysates lacking Kip3, Spc105 (green) was immobile on the microtubule (red) surface. When Kip3 +lysate was added, the same, previously immobile, kinetochores started to translocate toward the microtubule plus ends (marked with red arrows).
-
Figure 4—source data 1
Source data supplied.
- https://cdn.elifesciences.org/articles/78450/elife-78450-fig4-data1-v1.zip
Figure 4—video 1
Kinetochores do not move when the Kip3 gene is deleted.
When Kip3 is deleted from the lysate, Spc105 (green) does not move on the lattice of the microtubule (red). The kinetochore is stationary on the microtubule. Scale bar is 5 µm. Frame rate is 20 frames per second.
Figure 5 with 3 supplements
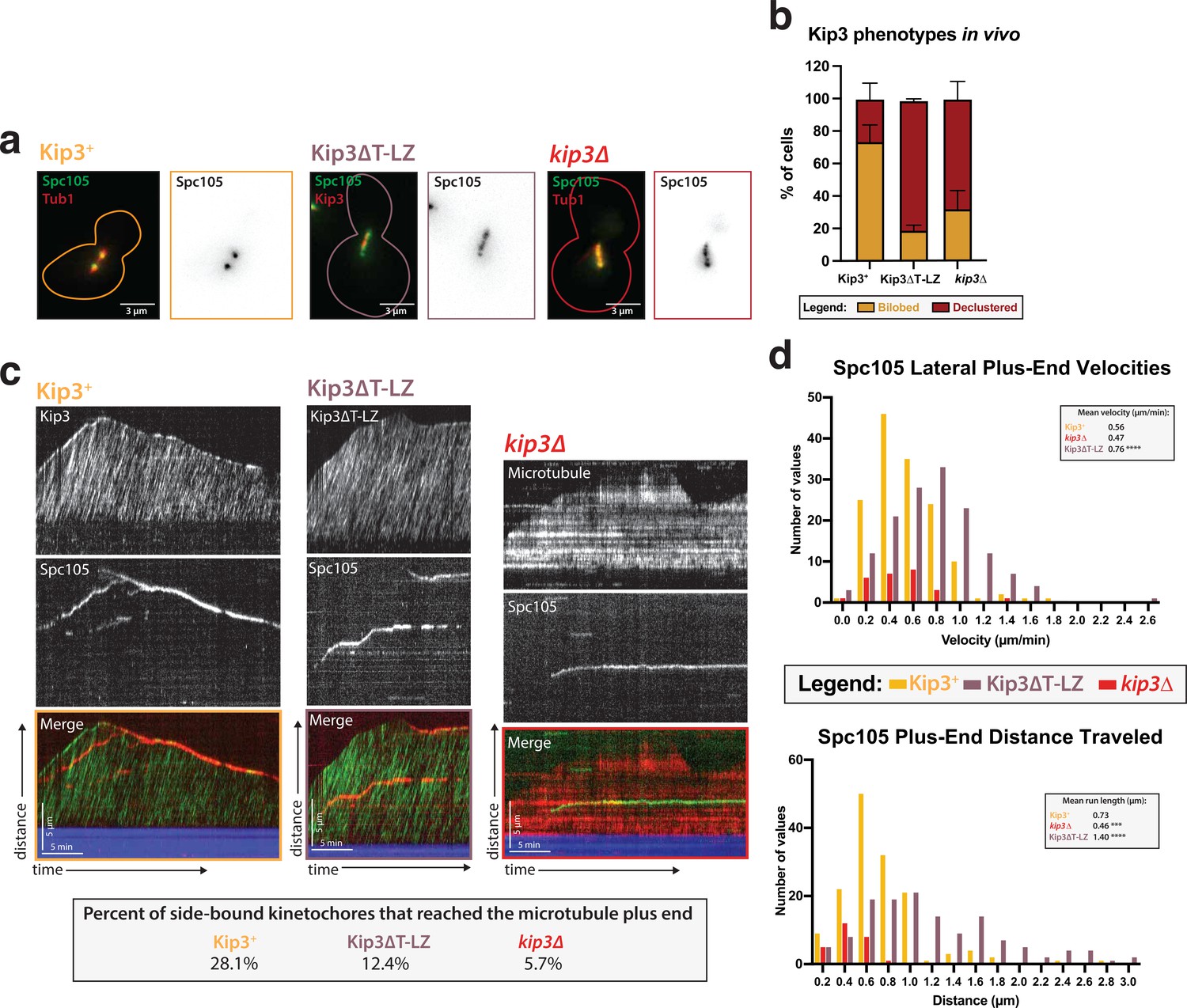
Kip3’s motor activity, but not its depolymerase tail domain, is necessary for processive plus end-directed kinetochore movement.
(a) Metaphase cells with either wild-type Kip3, kip3Δ, or kip3ΔT-LZ alleles imaged in a cdc23-1 strain expressing Spc105-GFP (green), mRuby-Tub1 or Kip3 (red). Note that the cells were not shifted to non-permissive temperature and were not arrested for imaging. Images are a maximum Z-projection of a 5 µm stack of 0.2 µm slices. (b) Quantification of the declustered kinetochore phenotype is shown as a bar graph for spindles that were 2–3 µm in length. Cells were classified as ‘bi-lobed’ by a line scan showing two distinct peaks. The ‘declustered’ phenotype included all line scans that did not show two peaks (line scans not shown). Two replicates were performed, each with n=50 cells. Error bars are standard error of the mean. (c) Representative kymographs of Spc105 motility in wild-type, kip3ΔT-LZ, and kip3Δ lysates. Dual color tagging of wild-type Kip3 and Spc105 showed no clear colocalization between the kinetochore (red) and Kip3 (green). Kip3 is highly processive, moving much faster than the kinetochore. A Kip3 mutant containing a deletion in the tail region (Kip3ΔT-LZ, green) with the kinetochore (red) was still processive along the microtubule. The Kip3ΔT-LZ mutant showed robust processivity but it did not accumulate on microtubule plus ends and depolymerization events were fewer than in Kip3 +lysates, as shown in previously published literature. Lastly, a representative kymograph for kip3Δ lysate shows immobile Spc105 (green) on microtubules (red). (d) Histograms of Spc105 velocities and average run length when laterally bound and moving toward the microtubule plus ends. The number of values is shown on the y-axis and velocity/distance traveled on the x-axis, binned every 0.2 units. The mean velocity of wild-type Kip3 protein and Kip3ΔT-LZ protein is 0.55 µm/min and 0.76 µm/min, respectively. The average run length also increased from 0.73 µm to 1.4 µm. The mean Spc105 velocity in kip3Δ lysates was unchanged compared to wild-type, but the number of runs was reduced to 25% of wild-type. The average length decrease in kip3Δ was reduced from 0.73 µm to 0.45 µm. The mean is from two replicate trials of one biological replicate. Statistical analysis was done using a Kruskal-Wallis test where *** p=0.0006 and **** p<0.0001. For lysates from each strain, as listed from WT to Kip3ΔT-LZ, N=128, 70, 81 kinetochores tracked.
-
Figure 5—source data 1
Source data supplied.
- https://cdn.elifesciences.org/articles/78450/elife-78450-fig5-data1-v1.zip
Figure 5—figure supplement 1
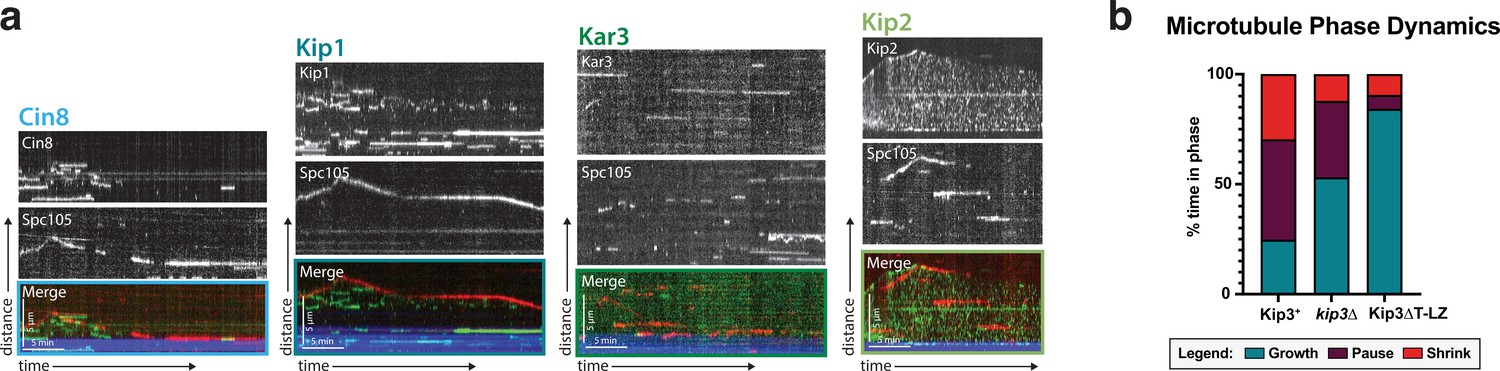
Kinetochores do no colocalize with the other four kinesins and Kip3’s effect on microtubule dynamics.
(a) Dual-color tagging of the kinetochore protein Spc105 and each of the five kinesins shows no clear colocalization. Representative kymographs are shown with the Spc105 in red and the kinesin motor proteins in green. The microtubule seed (minus-end) is in blue. (b) Across each of the three Kip3 conditions: wild-type, kip3Δ, and Kip3ΔT-LZ, the time the microtubule spent growing, shrinking, and paused changes. Wild-type Kip3 is the most dynamic, spending almost equal time in all three phases. The kip3Δ strain has an increased time spent paused and growing, while the Kip3ΔT-LZ strain spends the most time growing, rarely pausing or shrinking.
Figure 5—video 1
Kinetochore and Kip3 dynamics Spc105 (red) and Kip3 (green) move on the lattice of the microtubule towards the plus-end.
Kip3 has short periods where it colocalizes and runs ‘through’ the slow moving kinetochore. Scale bar is 5 µm. Frame rate is 20 frames per second.
Figure 5—video 2
Kinetochore and Kip3ΔT-LZ dynamics Spc105 (red) and Kip3ΔT-LZ (green) move on the lattice of the microtubule towards the plus-end.
Kip3 has short periods where it colocalizes and runs ‘through’ the slow moving kinetochore. Scale bar is 5 µm. Frame rate is 20 frames per second.
Figure 6

Cartoon model of reconstituted kinetochore dynamics on the microtubule Yeast kinetochores reconstituted binding with individual yeast microtubules in a yeast protein extract.
These laterally-bound kinetochores travel in a directional manner to the plus end of the microtubule, powered by the kinesin-8, Kip3. Upon reaching the end of the microtubule, the kinetochore transitions to end-on attachment. Establishment of this end-on attachment coincides with onset of microtubule depolymerization and the end-bound kinetochore preventing the microtubule from elongating again.
Author response image 1
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, MIF2-GFP, mRuby2-TUB1 | This paper | DDY 5806 | MATa, cdc23-1, MIF2-GFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, MIF2-GFP, mRuby2-TUB1 | This paper | DDY 5807 | MATalpha, cdc23-1, MIF2-GFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, MTW1-GFP, mRuby2-TUB1 | This paper | DDY 5808 | MATa, cdc23-1, MTW1-GFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, MTW1-GFP, mRuby2-TUB1 | This paper | DDY 5809 | MATalpha, cdc23-1, MTW1-GFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, CSE4-GFP, mRuby2-TUB1 | This paper | DDY 5810 | MATa, cdc23-1, CSE4-3x flexible linker-yoEGFP::KanMX, mRuby2-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, CSE4-GFP, mRuby2-TUB1 | This paper | DDY 5811 | MATalpha, cdc23-1, CSE4-3x flexible linker-yoEGFP::KanMX, mRuby2-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, OKP1-GFP, mRuby2-TUB1 | This paper | DDY 5812 | MATa, cdc23-1, OKP1-yoEGFP::KanMX, URA3::mRuby2-TUB1, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, OKP1-GFP, mRuby2-TUB1 | This paper | DDY 5813 | MATalpha, cdc23-1, OKP1-yoEGFP::KanMX, URA3::mRuby2-TUB1, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, CTF3-GFP, mRuby2-TUB1 | This paper | DDY 5814 | MATa, cdc23-1, CTF3-yoEGFP::KanMX, URA3::mRuby2-TUB1, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, CTF3-GFP, mRuby2-TUB1 | This paper | DDY 5815 | MATalpha, cdc23-1, CTF3-yoEGFP::KanMX, URA3::mRuby2-TUB1, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc7-1, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5816 | MATa, cdc7-1, SPC105-GFP::KanMX, mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc15-2, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5817 | MATa, cdc15-2, SPC105-GFP::KanMX, mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5818 | MATa, cdc23-1, SPC105-3x flexible linker-yoEGFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5819 | MATalpha, cdc23-1, SPC105-3x flexible linker-yoEGFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-GFP, kip3Δ, mRuby2-TUB1 | This paper | DDY 5820 | MATa, cdc23-1, SPC105-3x flexible linker-yoEGFP::KanMX, kip3Δ::HIS3MX6, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-GFP, kip3Δ, mRuby2-TUB1 | This paper | DDY 5821 | MATalpha, cdc23-1, SPC105-3x flexible linker-yoEGFP::KanMX, kip3Δ::HIS3MX6, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-GFP, cin8Δ, mRuby2-TUB1 | This paper | DDY 5822 | MATa, cdc23-1, SPC105-3x flexible linker-yoEGFP::KanMX, cin8Δ::HIS3MX6, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-GFP, cin8Δ, mRuby2-TUB1 | This paper | DDY 5823 | MATalpha, cdc23-1, SPC105-3x flexible linker-yoEGFP::KanMX, cin8Δ::HIS3MX6, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-mScarlet-I, MTW1-GFP, HaloTag-TUB1 | This paper | DDY 5824 | MATa, cdc23-1, NDC80-mScarlet-I::LEU2, MTW1-GFP::KanMX, ura3-52::PHIS3-HaloTag-flexible linker-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-mScarlet-I, SPC105-GFP, HaloTag-TUB1 | This paper | DDY 5825 | MATa, cdc23-1, NDC80-mScarlet-I::LEU2, SPC105-3x flexible linker-yoEGFP::KanMX, ura3-52::PHIS3-HaloTag-flexible linker-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-mScarlet-I, ASK1-GFP, HaloTag-TUB1 | This paper | DDY 5826 | MATa, cdc23-1, NDC80-mScarlet-I::LEU2, ASK1-yoEGFP::KanMX, ura3-52::PHIS3-HaloTag-flexible linker-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-mScarlet-I, MIF2-GFP, HaloTag-TUB1 | This paper | DDY 5827 | MATa, cdc23-1, NDC80-mScarlet-I::LEU2, MIF2-GFP::KanMX, ura3-52::PHIS3-HaloTag-flexible linker-TUB1::URA3, his3-Δ200, leu2-3,112 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, MTW1-mScarlet-I, OKP1-GFP, HaloTag-TUB1 | This paper | DDY 5828 | MATalpha, cdc23-1, MTW1-mScarlet-I::LEU2, Okp1-yoEGFP::KanMX, ura3::PHis3-HaloTag-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-mScarlet-I, CSE4-GFP, HaloTag-TUB1 | This paper | DDY 5829 | MATa, cdc23-1, NDC80-mScarlet-I::LEU2, CSE4-3x flexible linker-yoEGFP::KanMX, ura3-52::PHIS3-HaloTag-flexible linker-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-mScarlet-I, CTF3-GFP, HaloTag-TUB1 | This paper | DDY 5830 | MATa, cdc23-1, NDC80-mScarlet-I::LEU2, CTF3-yoEGFP::KanMX, ura3-52::PHIS3-HaloTag-flexible linker-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-mScarlet-I, KIP3-GFP, HaloTag-TUB1 | This paper | DDY 5831 | MATa, cdc23-1, SPC105-mScarlet-I::LEU2, KIP3-yoEGFP::KanMX, ura3-52::PHIS3-HaloTag-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-mScarlet-I, CIN8-GFP, HaloTag-TUB1 | This paper | DDY 5832 | MATalpha, cdc23-1, SPC105-mScarlet-I::LEU2, CIN8-GFP::KanMX, ura3-52::PHIS3-HaloTag-TUB1::URA3, his3-Δ200, leu2-3,112 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, SPC105-mScarlet-I, KIP1-GFP, HaloTag-TUB1 | This paper | DDY 5833 | MATa, cdc23-1, SPC105-mScarlet-I::LEU2, KIP1-GFP::KanMX, ura3-52::PHIS3-HaloTag-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, kip3 480 GCN4 coiled-coil-EYFP, SPC105-mScarlet-I, KIP1-GFP, HaloTag-TUB1 | This paper | DDY 5835 | MATalpha, cdc23-1, kip3Δ::KanR::pKIP3-kip3 480 GCN4 coiled-coil-EYFP-tADH::LEU2, SPC105-mScarlet-I::LEU2, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, KIP2-GFP, SPC105-mScarlet-I, HaloTag-TUB1 | This paper | DDY 5836 | MATa, cdc23-1, KIP2-3x flexible linker-yoEGFP::KanMX, SPC105-mScarlet-I::LEU2, ura3-52::PHIS3-HaloTag-TUB1::URA3, his3-Δ200, leu2-3,112, lys2-801(am) |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, kip2Δ, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5837 | MATa, cdc23-1, kip2Δ::CgHIS3, SPC105-3x flex linker-yoEGFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, kip2Δ, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5838 | MATalpha, cdc23-1, kip2Δ::CgHIS3, SPC105-3x flex linker-yoEGFP::KanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, kip1Δ, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5840 | MATalpha, cdc23-1, SPC105-GFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, KIP1Δ::HIS3M × 6, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, kip1Δ, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5841 | MATa, cdc23-1, SPC105-GFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, KIP1Δ::HIS3M × 6, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, ASK1-GFP, mRuby2-TUB1 | This paper | DDY 5842 | MATalpha, his3Δ200, lys2-801(am) am, ura3-52, leu2-3,112, ASK1-EGFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, ASK1-GFP, mRuby2-TUB1 | This paper | DDY 5843 | MATa, his3Δ200, lys2-801(am), ura3-52, leu2-3,112, ASK1-EGFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, KAR3-AID, TIR1, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5844 | MATalpha, lys2, KAR3-AID-9myc::HIS3, TIR1::LEU2, SPC105-GFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, KAR3-AID, TIR1, SPC105-GFP, mRuby2-TUB1 | This paper | DDY 5845 | MATalpha, lys2, KAR3-AID-9myc::HIS3, TIR1::LEU2, SPC105-GFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-GFP, mRuby2-TUB1 | This paper | DDY 5846 | MATalpha, his3Δ200, lys2-801(am), ura3-52, leu2-3,112, NDC80-EGFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, NDC80-GFP, mRuby2-TUB1 | This paper | DDY 5847 | MATa, his3Δ200, lys2-801(am), ura3-52, leu2-3,112, NDC80-EGFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, CNN1-GFP, mRuby2-TUB1 | This paper | DDY 5848 | MATalpha, his3Δ200, lys2-801(am), ura3-52, leu2-3,112, CNN1-EGFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, CNN1-GFP, mRuby2-TUB1 | This paper | DDY 5849 | MATalpha, his3Δ200, lys2-801(am), ura3-52, leu2-3,112, CNN1-EGFP::kanMX, TUB1::PHIS3-mRuby2-TUB1::HPH, cdc23-1 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, GFP-KAR3, SPC105-mScarlet-I | This paper | DDY 5850 | MATa, EGFP-KAR3::KanMX, SPC105-mScarlet-I::LEU2, cdc23-1, his3-Δ200, leu2-3,112, lys2-801(am), ura3-52 |
| Strain, strain background (Saccharomyces cerevisiae) | cdc23-1, MIF2-GFP, MTW1-mScarlet-I | This paper | DDY 5851 | MATalpha, MIF2-GFP::KanMX, MTW1-mScarlet-I::LEU2, cdc23-1, lys2-801(am), ura3-52, leu2-3,112, his3Δ200 |
| Chemical compound, drug | Benzonase, purity >90% | EMD Millipore | Cat. Num: 70746–3 | |
| Chemical compound, drug | Concavalin A | Sigma-Aldrich | Cat. Num: C2010 | |
| Chemical compound, drug | GMPCPP | Jena Bioscience | Cat. Num: NU-405 | |
| Chemical compound, drug | Hellmanex III | Hellma Analytics | Cat. Num: 9-307-011-4-507 | |
| Chemical compound, drug | 3-indole acetic acid, 98% | Sigma-Aldrich | Cat. Num: I3750 | |
| Chemical compound, drug | Janelia Fluor 646 HaloTag ligand | Promega | Cat. Num: GA1120 | |
| Chemical compound, drug | poly-L-lysine grafted with polyethylene glycol | SuSoS AG | PLL(20)-g[3.5]-PEG(2) | |
| Chemical compound, drug | poly-L-lysine grafted with polyethylene glycol-biotin (50%) | SuSoS AG | PLL(20)-g[3.5]-PEG(2)/PEG(3.4)-biotin(50%) | |
| Chemical compound, drug | protease inhibitor cocktail IV | Calbiochem | Cat. Num: 539,136 | |
| Other | BSA gold tracer, 10 nm | Electron Microscopy Science | Cat. Num: 25,486 | Used for electron microscopy |
| Other | holey carbon electron microscopy grid, Quantifoil R 2/1, copper grid | Electron Microscopy Science | Cat. Num: Q210CR1 | Used for electron microscopy |
| Recombinant protein | Tubulin protein (HiLyte Fluor 647) - Porcine brain | Cytoskeleton | Cat. Num: TL670M | |
| Recombinant protein | Tubulin protein (Biotin) - Porcine brain | Cytoskeleton | Cat. Num: T333P | |
| Software, algorithm | Fiji/ImageJ | NIH | https://imagej.net/ | |
| Software, algorithm | IMOD | University of Colorado | https://bio3d.colorado.edu/imod/ | |
| Software, algorithm | Prism | Graphpad |
Author response table 1
Immediate Behavior of Kinetochores that Reach the Microtubule Plus-End.
| Wild-type Kip3 | kip3∆ | Kip3∆T-LZ | |
|---|---|---|---|
| Move toward the seed | 27 (75%) | 4 (57%) | 3 (30%) |
| No net movement | 9 (25%) | 3 (43%) | 7 (70%) |
| Move away from the seed | 0 (0%) | 0 (0%) | 0 (0%) |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Reconstitution of kinetochore motility and microtubule dynamics reveals a role for a kinesin-8 in establishing end-on attachments
eLife 11:e78450.
https://doi.org/10.7554/eLife.78450




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}