Bumblebees retrieve only the ordinal ranking of foraging options when comparing memories obtained in distinct settings
- Department of Psychology, School of Public Health, Southern Medical University, China
- Ecology and Genetics Research Unit, University of Oulu, Finland
- Biological and Experimental Psychology, School of Biological and Behavioural Sciences, Queen Mary University of London, United Kingdom
- Department of Biological Sciences, Macquarie University, Australia
- Department of Psychiatry, Zhujiang Hospital, Southern Medical University, China
- Guangdong-Hong Kong-Macao Greater Bay Area Center for Brain Science and Brain-Inspired Intelligence, Southern Medical University, China
Figures
Figure 1 with 4 supplements

Bumblebees make decisions based on ordinal comparisons.
(A, C, and E) The corresponding sucrose concentration of each stimulus is displayed on a log scale to visually represent their relative differences according to Weber’s Law (Akre and Johnsen, 2014). Training sessions are indicated by separate grey backgrounds, and the test options in each experiment are indicated with dashed lines. The bi-colour squares indicate that the colours for the focal options used were counterbalanced across bees. (B, D, and F) Test results for each experiment. Groups indicate different counterbalanced training sequence and colour-reward contingency (see Figure 1—figure supplement 3 for more details). Each filled circle represents the proportion of choices for option C by an individual bumblebee (10 individuals per group). Dashed horizontal lines indicate chance performance. Vertical lines indicate mean ± SEM. p values were calculated from generalised linear models (Materials and methods); NS: p > 0.05; *: p < 0.01.
Figure 1—figure supplement 1
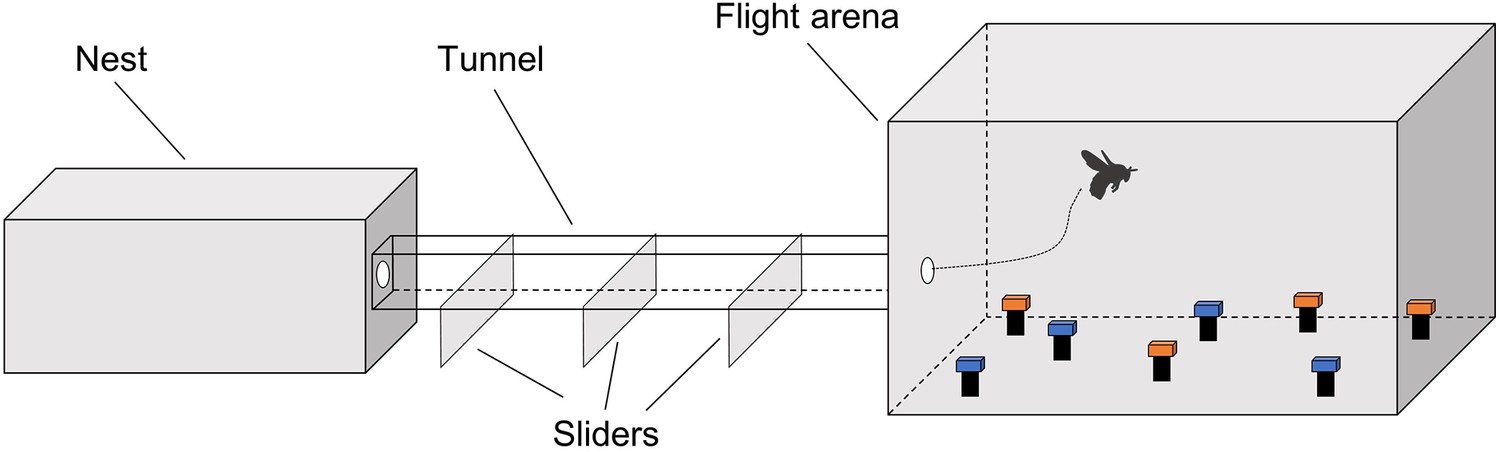
General setup for experiments 1, 2, and 3.
In each experiment, artificial flowers of paired colours were horizontally presented in the training and testing phase.
Figure 1—figure supplement 2
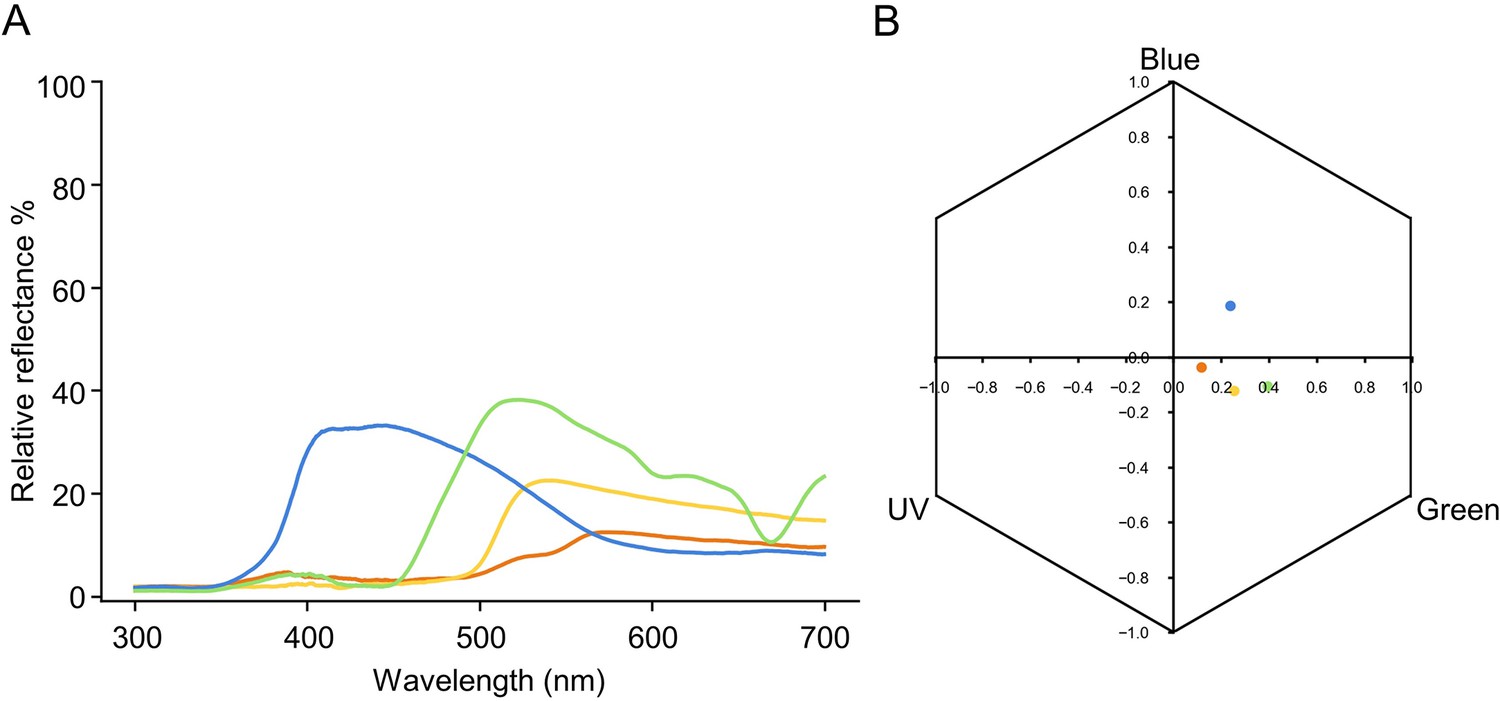
Specifications of the colours used for all the experiments.
(A) Spectral reflectance plot of blue, orange, yellow, and green colours used. (B) Loci of colours in the hexagonal bee colour space, determined by the responses each colour elicits on the bee’s UV, blue, and green photoreceptors (Chittka, 1992).
Figure 1—figure supplement 3
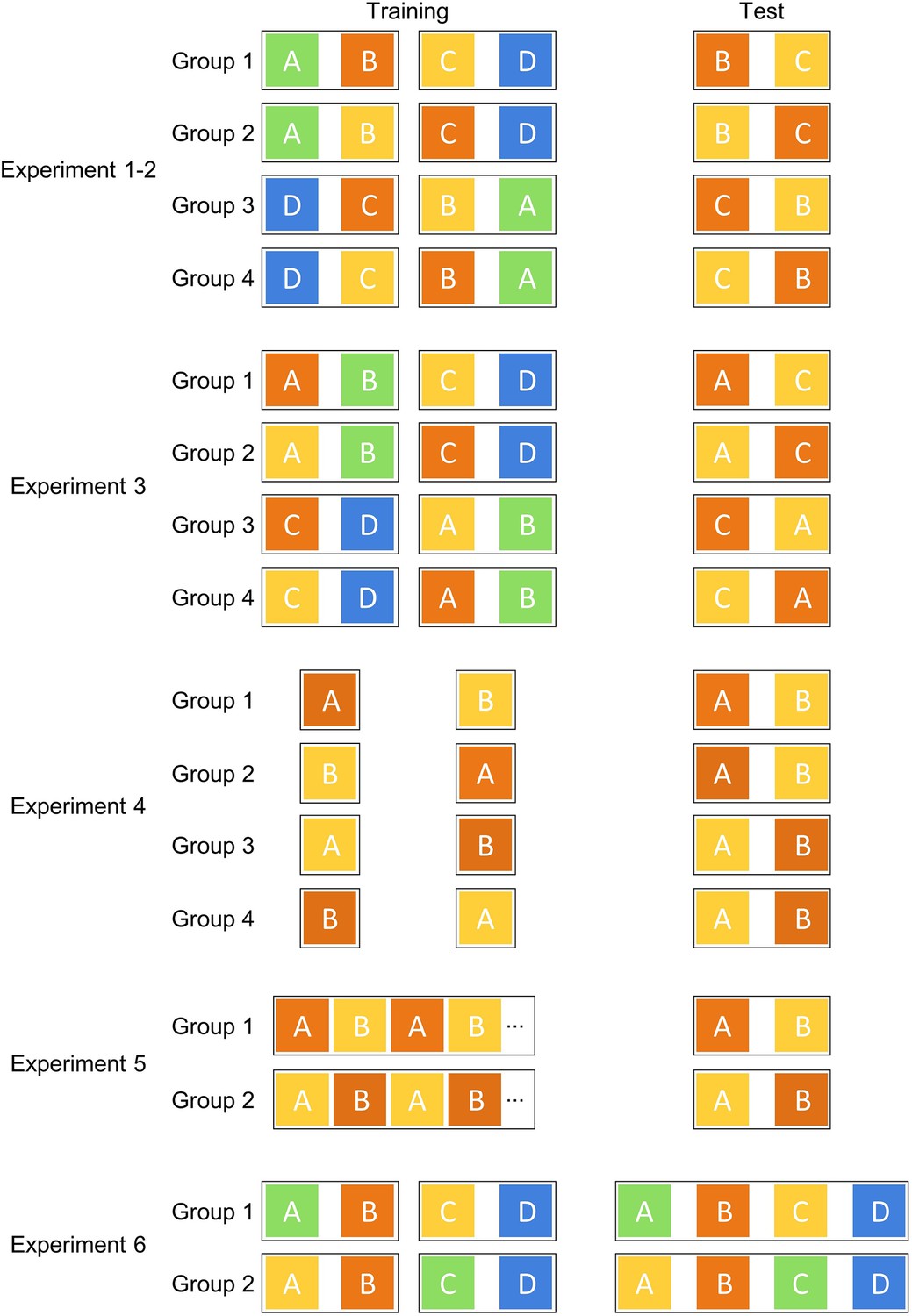
Counterbalanced colour sets used in experiments 1–6.
Groups of bees were trained and tested with counterbalanced colour sets and training sequences in each experiment.
Figure 1—figure supplement 4
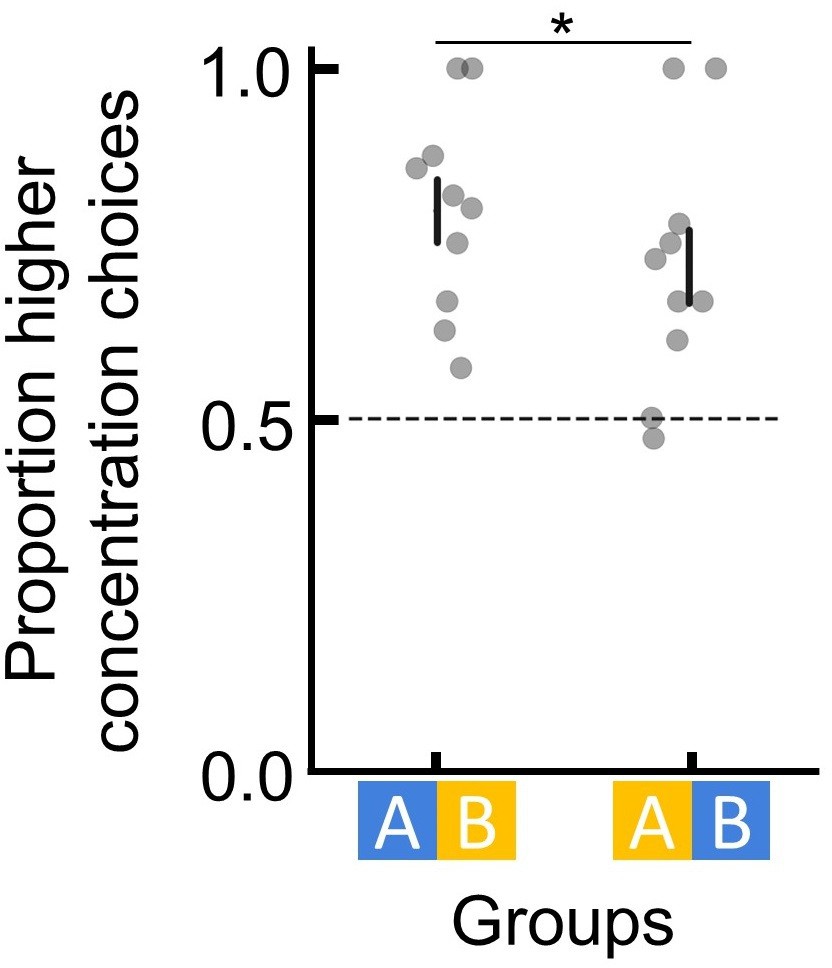
Bees can discriminate flowers of different colours and sucrose concentrations.
To ensure that bees were able to learn to discriminate between two differently coloured flowers in our setup, we examined bees’ preference between two flower types after having been trained individually on these flowers. One group of bees (n=10) learned that blue flowers contained 45% sucrose solution and that yellow flowers contained 30% sucrose solution. Another group of bees (n=10) learned the counterbalanced colour-reward contingency. During a subsequent unrewarded test (all flowers with unrewarding water), bees showed a clear preference for (landed more often on) the flowers that had been associated with the higher reward during training (generalised linear model [GLM]: N=20, 95% CI = [0.64, 1.37], and p=3.39e-5). These results show that in our setup, bumblebees were able to easily learn to discriminate the different flower colours used in our experiments and do so via the different sucrose concentrations associated with each flower type. Groups indicate different counterbalanced colour-reward contingency for bees. Dotted horizontal lines indicate chance performance. Vertical lines indicate mean ± SEM. p values were calculated from generalised linear models (Materials and methods); *: p < 0.01.
Figure 2
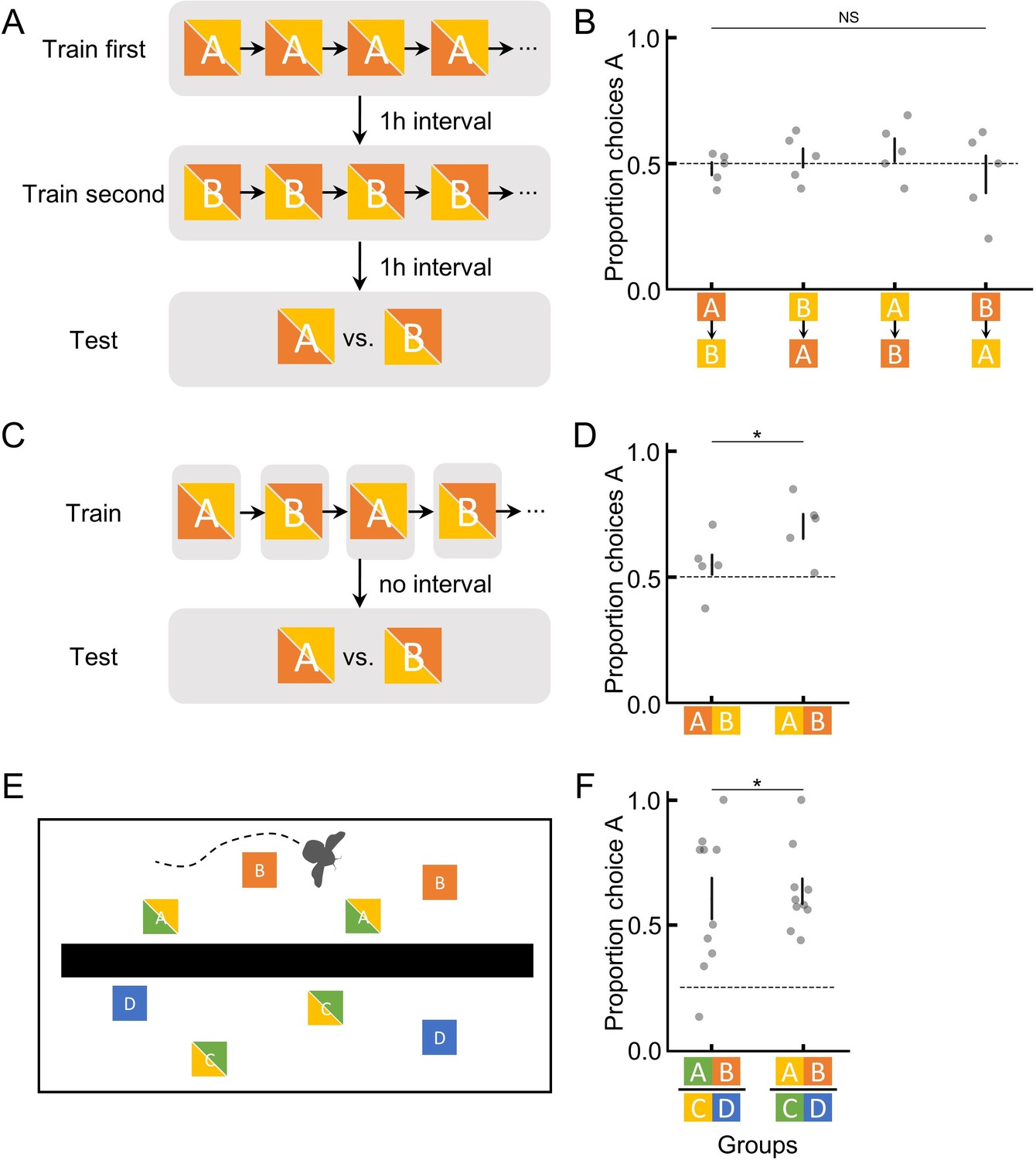
Bumblebees are only able to utilise absolute information for short periods of time in order to encode and recall ranking information in novel contexts.
(A) Illustration of the training and testing procedure in experiment 4, where two options were separately trained and tested, with a 1-hr interval in between to ensure clear spatial and temporal separation (and no ranking information). (C) The training and testing procedure in experiment 5, where two options were trained in alternating bouts (spatially separated but temporally close). (E) Top view of setup for experiment 6 where a wall separated flowers such that the bee could not see both groups of flowers at the same time. (B, D, and F) Results of the unrewarding test in experiments 4–6. Groups indicate different colour-reward contingency for bees (B & D: 5 bees per group; F: 10 bees per group; Figure 1—figure supplement 3). The bi-colour squares indicate that the colours for the focal options used were counterbalanced across bees. Dashed horizontal lines indicate chance performance. Vertical lines indicate mean ± SEM. p values were calculated from generalised linear models (Materials and methods); NS: p > 0.05; *: p < 0.01.
Tables
Table 1
Predictions for various decision strategies for bumblebees’ flower preferences in experiments 1, 2, and 3.
The left column lists the different categories of decision strategies. The middle three columns show the predicted results of each strategy for each experiment. The right column shows whether the predictions of each strategy match the behaviour of bumblebees in all experiments. A lexicographic combination strategy is where choices are determined by a difference in a priority dimension, but if options are equal in that dimension, then choices rely on a secondary dimension. For example, bumblebees might choose between two flower types based on a remembered ranking (as long as flowers differed in ranking during training), no matter what the absolute sugar concentration differences were. However, if the remembered rankings of the flowers are the same (i.e. they cannot use ranking to make a choice), bumblebees might then use any difference in absolute memory. In contrast, a non-lexicographic combination strategy is where differences in either dimension can be used to make a choice. For example, bumblebees might use absolute memories to choose flowers that differ in ranking, even if the ranking and absolute contrasts are distinct (e.g. one ordinal rank difference vs threefold absolute difference). Value by association entails one option acquiring a higher value because it was experienced in a richer environment (Pompilio and Kacelnik, 2010), i.e. bumblebees might assign a higher value to flower B because it was experienced with A (the highest rewarding flower), compared to a lower value assigned to C because it was experienced with D (the lowest rewarding flower). State-dependent valuation learning (SDVL) refers to assigning values to options based on whether subjects were in a better or worse internal state when experiencing those options (McNamara et al., 2012). For example, bumblebees may assign different values to a flower type based on whether they are foraging in a more or less profitable environment, i.e. based on the comparison between each flower type’s sugar concentration and the average sucrose concentration within the bee’s current crop load.
| Strategy | Expected preference | Matches bees’ behaviour | ||||
|---|---|---|---|---|---|---|
| Experiment 1B (30%) vs C (30%) | Experiment 2B (30%) vs C (15%) | Experiment 3A (45%) vs C (30%) | ||||
| Absolute memory | Indifferent | B | A | ✗ | ||
| Remembered ranking | C | C | Indifferent | ✓ | ||
| Lexicographic combination—absolute memory priority | C | B | A | ✗ | ||
| Lexicographic combination—ranking priority | C | C | A | ✗ | ||
| Non-lexicographic combination | C | B/C/indifferent | A | ✗ | ||
| Value by association | B | B | A | ✗ | ||
| State-dependent valuation learning | C | C | C | ✗ | ||
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Bumblebees retrieve only the ordinal ranking of foraging options when comparing memories obtained in distinct settings
eLife 11:e78525.
https://doi.org/10.7554/eLife.78525




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}