Convergence of two global regulators to coordinate expression of essential virulence determinants of Mycobacterium tuberculosis
- CSIR-Institute of Microbial Technology, India
Figures
Figure 1 with 1 supplement

PhoP-depletion impacts expression of a subset of CRP-regulon genes.
(A) Quantitative RT-PCR was carried out to compare the expression level of icl1, umaA, and whiB1 in indicated mycobacterial strains, grown under normal and specific stress conditions. Note that mycobacterial strains include WT-H37Rv (empty bar), ΔphoP-H37Rv (gray bar), and the mutant strain complemented with wild-type phoP (blue bar) or phosphorylation-defective phoPD71N (dark gray bar). The results show average values from biological triplicates, each with two technical repeats (*p≤0.05; **p≤0.01; ***p≤0.001). Fold changes in mRNA levels were determined as described in the Materials and methods. Notably, changes in gene expression levels were insignificant when compared between ΔphoP-H37Rv (gray bar) and the mutant complemented with phosphorylation-defective phoPD71N (dark gray bar). (B) To examine role of PhoP in crp expression, RT-qPCR compared expression levels of crp (Rv3676) in WT-H37Rv (empty bar), ΔphoP-H37Rv (gray bar) and the complemented mutant (black bar) (***p≤0.001). PhoP-dependent aprA expression was shown as a control. (C) In vivo recruitment of PhoP within target promoters was examined by ChIP-qPCR as described in the Materials and methods. espA promoter (espAup), and gapdh-specific enrichments were used as a positive and negative control, respectively. The experiments were performed in biological duplicates, each with two technical repeats (**p≤0.01; ***p≤0.001), and fold enrichment was determined relative to an IP sample without adding antibody (mock control). Non-significant differences are not indicated.
Figure 1—figure supplement 1
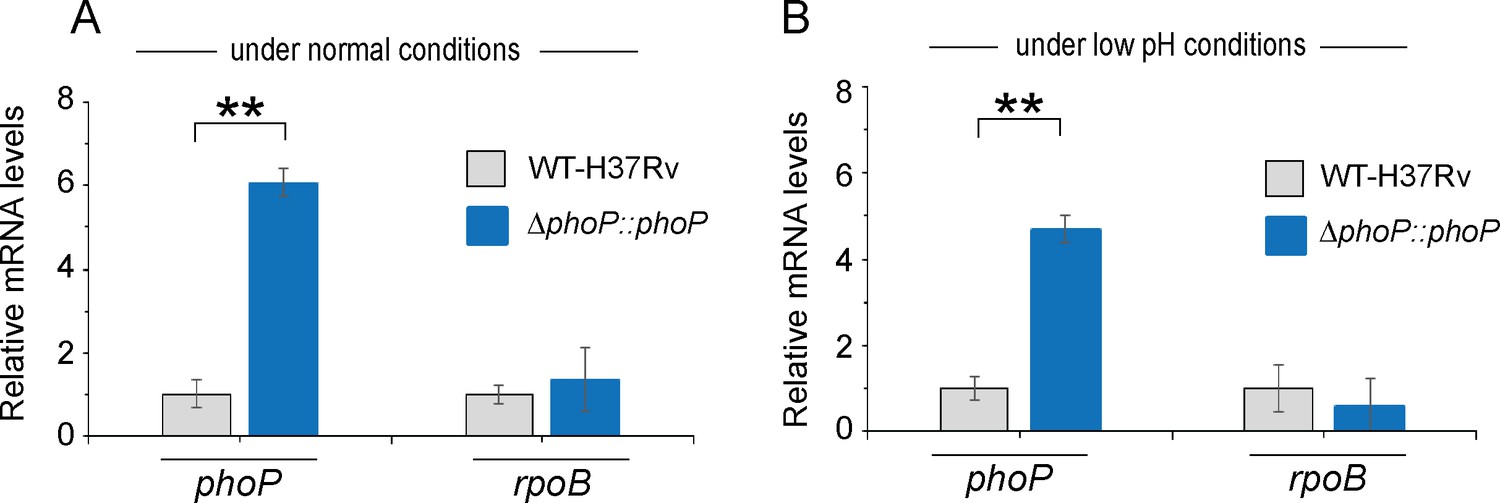
phoP expression in WT-H37Rv and complemented ΔphoP-H37Rv under normal and acidic conditions of growth.
Expression levels of phoP were measured in indicated mycobacterial strains grown under (A) normal pH (pH 7.0) and (B) acidic pH (pH 4.5) using RT-qPCR as described in the Materials and methods. rpoB was used as a control, and the average fold differences were determined from two biological repeats each with two technical repeats (**p≤0.01).
Figure 2 with 1 supplement
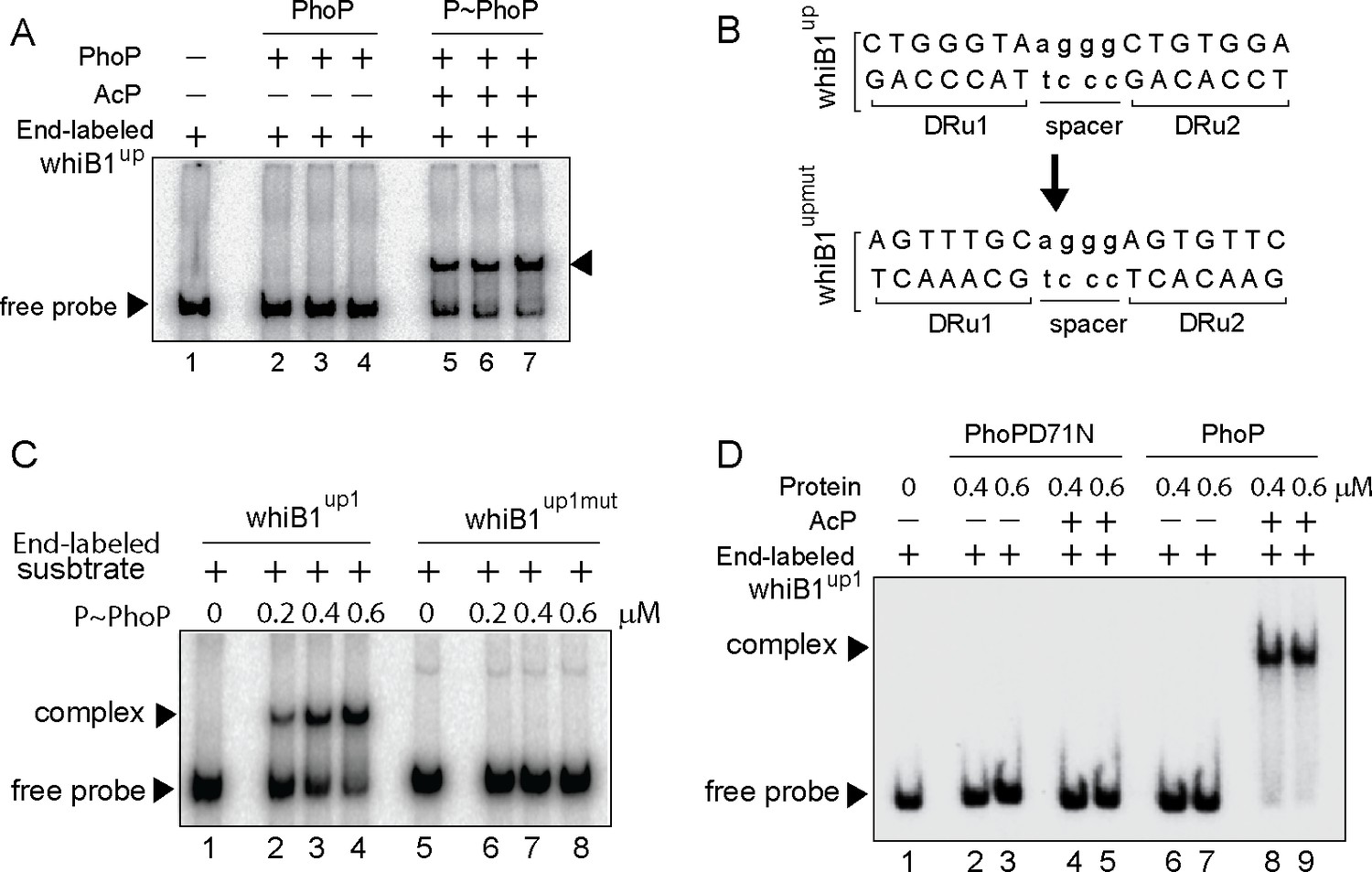
Probing core PhoP binding site within whiB1 promoter region.
(A) EMSA of radio-labeled whiB1up for binding of 0.2, 0.4, and 0.6 µM of PhoP (lanes 2–4) or P~PhoP (lanes 5–7), pre-incubated in a phosphorylation mix with acetyl phosphate (AcP) as the phospho-donor. The arrowheads on the left and right indicate the free probe and a slower moving complex, respectively. (B) PhoP binding motif consists of upstream (DRu1) and downstream (DRu2) repeat units. To construct mutant promoter (whiB1upmut), changes in both the repeat units were introduced by changing As to Cs and Gs to Ts and vice versa, and the orientation of DRu2 was reversed. whiB1upmut represents whiB1up fragment carrying changes only at the PhoP binding site. (C) EMSA experiment of labeled whiB1up (lanes 2–4), and whiB1upmut (lanes 6–8) to increasing concentrations of P~PhoP. The free probe and the slower moving complexes are indicated on the figure. (D) EMSA of radio-labeled whiB1up1 for binding of increasing concentrations of phosphorylation-deficient PhoPD71N (lanes 2–5) or PhoP (lanes 6–9), pre-incubated in phosphorylation mixture with or without AcP, respectively. Lane 1 shows the free probe. The assay conditions, sample analyses, and detection of radio-active samples are described in the Materials and methods.
Figure 2—figure supplement 1
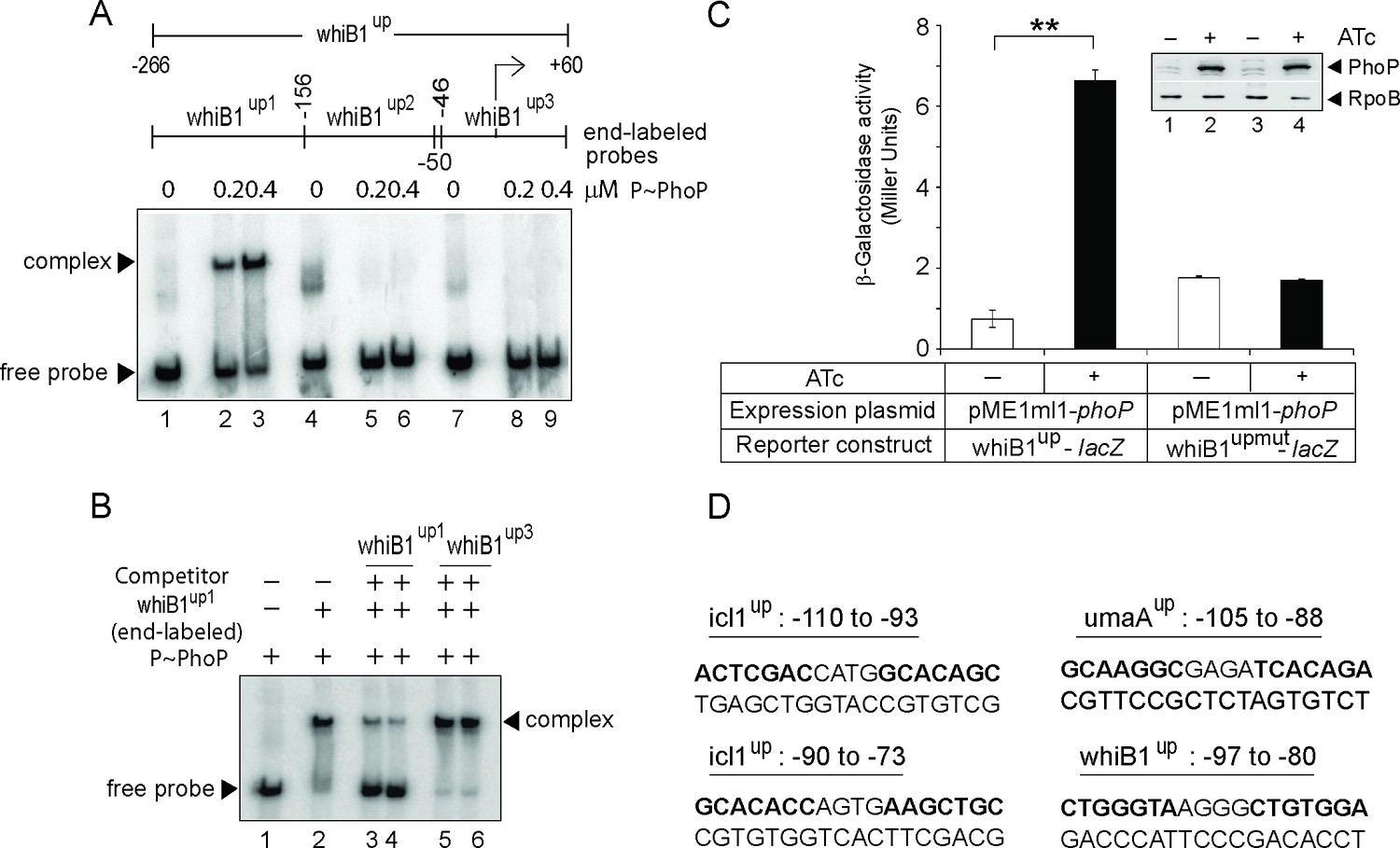
Probing core PhoP binding site within whiB1 promoter region.
(A) EMSA of radio-labeled whiB1up-derived fragments (whiB1up1-whiB1up3, as indicated) were performed with 0.2 and 0.4 µM of P~PhoP (lanes 2–3, 5–6, and 8–9, respectively), pre-incubated in a phosphorylation mix with acetyl phosphate (AcP) as the phospho-donor, to probe core binding site of the regulator. (B) To examine sequence-specific binding, EMSA of whiB1up1 with 0.3 µM of P~PhoP was carried out in absence (lane 2) or in presence of 20-fold, and 40-fold excess of specific competitor (unlabeled whiB1up1, lanes 3–4) or non-specific competitor (unlabeled whiB1up3, lanes 5–6), respectively. The assay conditions, sample analyses, and detection of radio-labeled samples are described in the Materials and methods; free probe and a slower moving complex are indicated on the figure. (C) To examine PhoP-regulated expression of whiB1up-lacZ, and whiB1upmut-lacZ fusions, M. smegmatis mc2155 harboring appropriate fusion constructs were grown, and β-galactosidase activities with or without inducing Mycobacterium tuberculosis PhoP expression were measured at 24 hr as described (Goyal et al., 2011). Note that in whiB1upmut, mutations were introduced only within both the PhoP binding repeat units of whiB1up1 as shown in the figure (Figure 2B). The results show average values with standard deviations from two biological repeats (***p value≤0.001). Inset compares PhoP expression in crude extracts with equal amount of total protein; RpoB was used as a loading control. (D) Nucleotide sequences of likely PhoP binding sites within indicated promoters. The sequences are numbered relative to their corresponding transcription start sites.
Figure 3 with 1 supplement

PhoP promotes CRP recruitment at the whiB1 promoter.
(A) To investigate how the promoter simultaneously accommodates both the regulators, EMSA experiments compared end-labeled whiB1up1 binding to increasing concentrations of purified CRP (lanes 2–3), PhoP (lanes 8–9), and both PhoP and CRP together (lanes 4–7). The assay conditions, sample analyses, and detection are described in the Materials and methods. Positions of the free probe and the complex are indicated on the figure. (B) Western blot analyses of protein fraction extracted from the excised gel fragment representing the complex (lane 5), as indicated by a box, was probed by anti-His antibody in lane 2; lane 1 resolved purified PhoP as a control. (C) Tricine SDS-PAGE analysis was carried out alongside indicating recombinant His-tagged CRP (lane 1) and PhoP (lane 2); lane 3 resolved marker proteins of indicated molecular masses. Protein samples were visualized by Coomassie blue staining. (D, E) To probe protein-protein interaction by M-PFC assays, Mycobacterium smegmatis expressing either (D) M. tuberculosis CRP and PhoP or (E) CRP and PhoPD71N, were grown on 7H10/hyg/kan plates in presence of TRIM, and growth was examined for strains co-expressing indicated fusion constructs. In both cases, empty vectors were included as negative controls, and co-expression of pUAB400-phoP/pUAB300-phoR encoding PhoP and PhoR, respectively, was used as a positive control. All the strains grew well in absence of TRIM.
Figure 3—figure supplement 1

Probing PhoP-CMR interactions.
(A) To probe protein-protein interaction by M-PFC assays, Mycobacterium smegmatis co-expressing M. tuberculosis PhoP and cyclic AMP macrophage regulator (CMR), were grown on 7H10/hyg/kan plates in the absence and presence of TRIM, and growth was examined. In both cases, empty vectors were included as negative controls, and co-expression of pUAB400-phoP/pUAB300-phoR (as a positive control) encoding PhoP and PhoR, respectively, showed M. smegmatis growth in presence of TRIM. All the strains grew well in absence of TRIM. (B) To verify expression of M. tuberculosis CRP and CMR in M. smegmatis, mRNA levels of the regulators were compared by RT-qPCR. Average fold changes in mRNA levels from two biological repeats (each with two technical repeats) are plotted, and non-significant difference is not indicated.
Figure 4
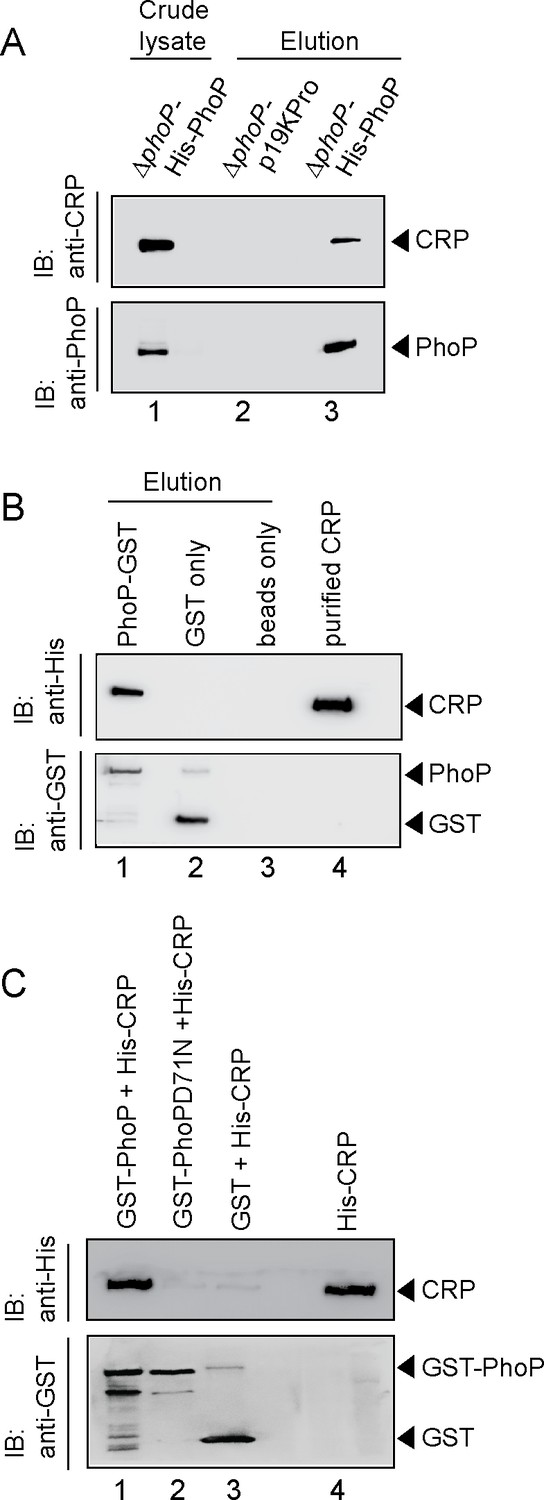
Probing CRP-PhoP interactions.
(A) To examine CRP-PhoP interaction in vivo, DNaseI-treated crude cell lysates of ΔphoP-H37Rv expressing His6-tagged PhoP (p19kpro-phoP; Supplementary file 1b) was incubated with pre-equilibrated Ni-NTA and eluted with 250 mM imidazole; lane 1, input sample; lane 2, control elution from the crude lysate of cells lacking phoP expression; lane 3, co-elution of CRP with PhoP. Blots were probed with anti-PhoP and anti-CRP antibody. (B) To investigate CRP-PhoP interaction in vitro, DNaseI-treated crude extract expressing His6-tagged CRP was incubated with glutathione epharose previously immobilized with GST-PhoP. Bound proteins (lane 1) were analyzed by Western blot using anti-His (upper panel) or anti-GST antibody (lower panel). Lane 1 shows presence of CRP bound to GST-PhoP. Identical experiment used glutathione Sepharose immobilized with GST alone (lane 2), or the resin alone (lane 3); lane 4 resolved recombinant His6-tagged CRP. (C) To examine whether phosphorylation of PhoP impacts CRP-PhoP interaction, crude lysates of cells expressing His6-tagged CRP was incubated with glutathione-Sepharose previously immobilized with GST-tagged PhoP (lane 1) or PhoPD71N (lane 2), carrying a single substitution of Asp-71 to Asn-71 and therefore, remains ineffective for phosphorylation at Asp-71. Analysis of bound fractions (lanes 1–2) was carried out as described in the legend to Figure 5 and control sets include glutathione Sepharose immobilized with GST alone (lane 3), or the resin alone (lane 4); lane 5 resolved recombinant His6-tagged CRP.
Figure 5 with 1 supplement
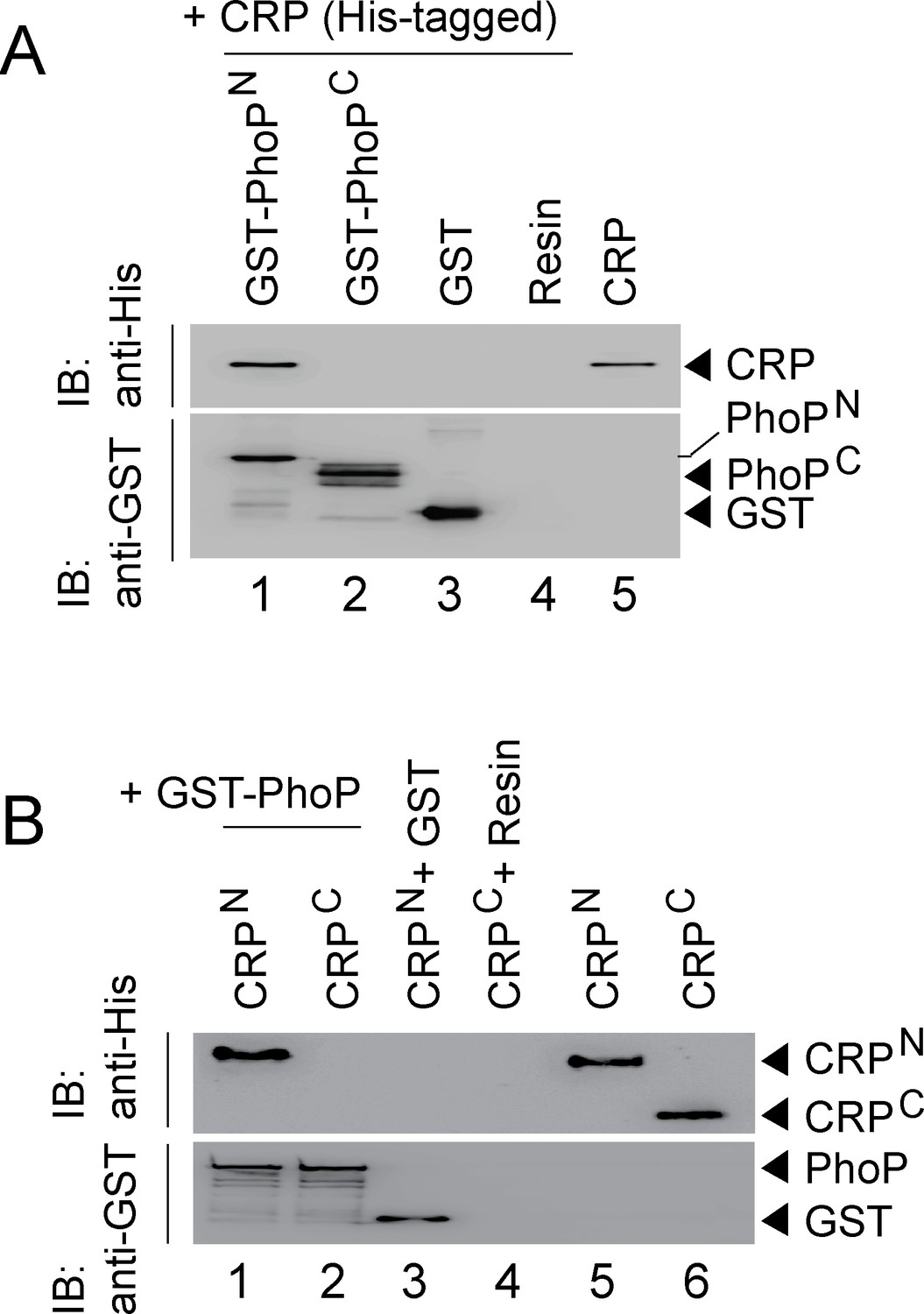
CRP and PhoP interact with each other via their corresponding N-terminal domains.
(A, B) CRP-PhoP interaction was probed by in vitro pull-down assays using either (A) His-tagged CRP and GST-tagged PhoP domains (GST-PhoPN and GST-PhoPC, respectively) or (B) GST-tagged PhoP and His-tagged CRP domains (CRPN, and CRPC, respectively). The domain constructs are listed in Supplementary file 1b. Fractions of bound proteins were analyzed by Western blot using anti-His (upper panel) or anti-GST antibody (lower panel). Control sets include glutathione Sepharose immobilized with GST (lane 3), or the resin (lane 4) alone; lane 5 of (A) resolved purified CRP, while lanes 5, and 6 of (B) resolved purified CRPN, and CRPC, respectively. The experimental procedures, and data analyses are as described in the legend of Figure 4B.
Figure 5—figure supplement 1
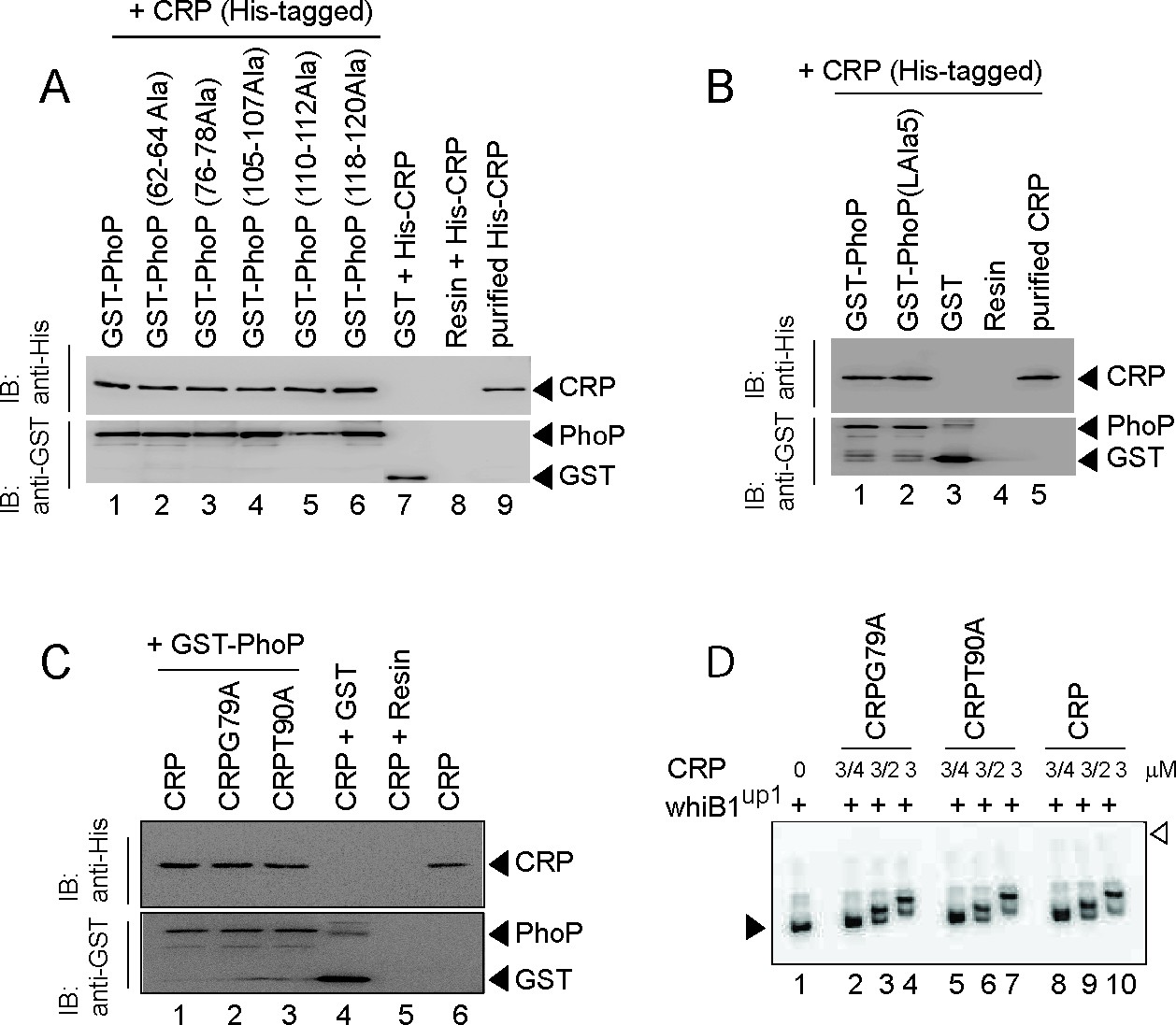
In vitro analysis of CRP-PhoP interactions.
(A) To assess the importance of residues of PhoPN, crude lysates of cells expressing His6-tagged CRP was incubated with glutathione-Sepharose, previously immobilized with GST-tagged WT (lane 1) or mutant PhoP proteins (lanes 2–6), each carrying a substitution of three PhoP residues with alanine (as indicated on the figure). Fractions of bound proteins (lanes 1–6) were analyzed by Western blot using anti-His (upper panel) or anti-GST antibody (lower panel). Control sets include glutathione Sepharose immobilized with GST alone (lane 7), or the resin alone (lane 8); lane 9 resolved recombinant His6-tagged CRP. (B) To examine role of the PhoP linker in CRP-PhoP interactions, crude lysates of cells expressing His6-tagged CRP was incubated with glutathione-Sepharose previously immobilized with GST-tagged PhoP (lane 1) or a PhoP linker mutant (lane 2), carrying a substitution of five linker residues (spanning Gly142 to Pro146 of PhoP) with alanine as described previously (Pathak et al., 2010). Analysis of bound fractions (lanes 1–2) was carried out as above and control sets include glutathione Sepharose immobilized with GST alone (lane 3), or the resin alone (lane 4); lane 5 resolved recombinant CRP. (C) To examine importance of cAMP binding to CRP on CRP-PhoP interaction, in vitro pull-down assays were carried out using His-tagged CRP mutants, deficient for cAMP binding. The experimental procedures, and data analyses are as described in the legend of Figure 4B. (D) To examine functionality of the mutant CRP proteins (CRPG79A and CRPT90A), deficient for cAMP binding, EMSA of radio-labeled whiB1up was carried out using indicated mutants (lanes 2–7) and WT-CRP (as a positive control; lanes 8–10), respectively. The assay conditions, sample analyses, and detection of radio-active samples are described in the Materials and methods. The filled and empty arrowheads on the left and right indicate free probe and the origin of the gel, respectively.
Figure 6
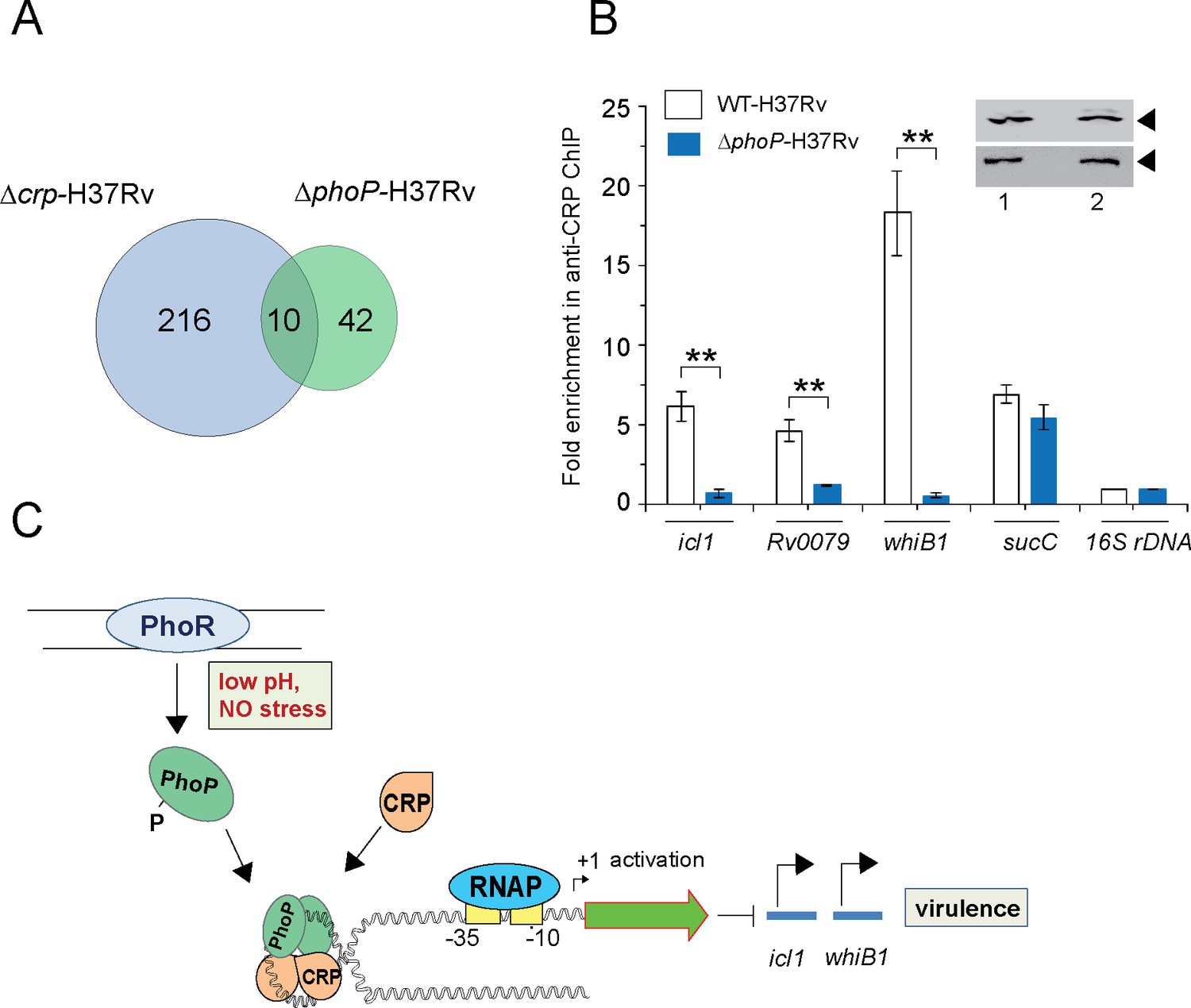
PhoP promotes CRP recruitment to regulate whiB1 expression.
(A) Venn diagram of genes differentially regulated in Δcrp-H37Rv and ΔphoP-H37Rv displays an overlap of ~10 promoters. The results are based on previously reported high throughput ChIP-sequencing data of CRP and PhoP (Kahramanoglou et al., 2014; Solans et al., 2014), respectively. Note that comparisons include genes annotated in Mycobacterium tuberculosis H37Rv genome only. (B) To examine effect of CRP-PhoP interaction, chromatin-immunoprecipitation (ChIP)-qPCR was carried out using anti-CRP antibody to compare in vivo recruitment of CRP in WT-H37Rv and ΔphoP-H37Rv as described in the Methods. Fold PCR enrichment due to CRP binding to indicated promoters was determined by duplicate measurements, each with two technical repeats (**p≤0.01). Inset compares CRP expression in ≈10 μg of indicated crude cell-lysates as probed by anti-CRP antibody; identical extracts were probed with anti-RpoB antibody as a loading control. (C) Schematic model showing newly-proposed molecular mechanism of activation of CRP-regulated promoters by simultaneous binding of CRP and PhoP. We propose that the interacting proteins (CRP and PhoP) remain bound to their cognate sites away from the start sites, and stabilize the transcription initiation complex so that RNA polymerase (RNAP) effectively transcribes these genes. Taken together, these molecular events mitigate stress by controlling expression of numerous genes and perhaps contribute to better survival of the bacilli in cellular and animal models.
Tables
Table 1
CRP and PhoP binding sites within the commonly regulated promoters*.
| Rvnumber | Gene name | PhoP binding sites | Sequence coordinates | CRP binding sites | Sequence coordinates |
|---|---|---|---|---|---|
| Rv0079 | Rv0079 | CCTCAGCTTCTGCGCAGC | –215 to –232 | GGTGACACAGCCCACA | –95 to –110 |
| Rv0116c | Rv0116c | GTACAGCTCGGTCGCAGC | –558 to –576 | TGTGGTCGCGATCACG | +29 to+45 |
| Rv0467 | icl1 | GAAGAGCGCGGAGCAGATC | +20 to +38 | TGTTACAACGCTCACA | –49 to –64 |
| Rv0469 | umaA | GCAAGGCGAGATCACAGA | –88 to –105 | TGTGACAGCCGTTGCG | –326 to –341 |
| Rv1535 | Rv1535 | GTGGTGCCGAAGCTCTGA | –312 to –330 | GTGGTGCCGAAGCTCT | –314 to –330 |
| Rv2329 | nark1 | GCTGTTTTCTTGCTGCGA | –194 to –212 | GGTGCGGCAGCCGGCA | +1460 to +1476 |
| Rv2524 | fas | GTAGAGCGAATTCCCAGC | –370 to –388 | GATTCCGAGCTGATCGAC | +6014 to +6030 |
| Rv2590 | fadD9 | TCACAGCCGATCAGCAGC | –104 to –122 | CCCGTGCCGCATCTCAC | –119 to –135 |
| Rv3219 | whiB1 | CTGGGGTAAGGGCTGTGGA | –191 to –208 | AGTGAGATAGCCCACG | –161 to –176 |
| Rv3616c | espA | TCGCAGCGCAGTTGCAGG | –197 to –215 | CGATCAGCACCTCGCG | +2221 to +2237 |
-
*
ChIP-sequencing data uncover ~10 promoters that belong to CRP regulon and are also regulated by PhoP. Both CRP and PhoP binding sites were identified in selected M. tuberculosis promoters by scanning for respective ChIP-seq data or SELEX-derived consensus sequence motifs (Galagan et al., 2013; He and Wang, 2014; Kahramanoglou et al., 2014; Solans et al., 2014). Nucleotide sequences of indicated binding sites are numbered with respect to corresponding ORF start sites.
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Mycobacterium tuberculosis) | WT-H37Rv | ATCC25618 | Wild-type M. tuberculosis strain | |
| Strain, strain background (M. tuberculosis) | ΔphoP-H37Rv | Walters et al., 2006 | phoPR locus Rv0757-Rv0758 has been inactivated | |
| Strain, strain background (M. tuberculosis) | ΔphoP::phoP | Walters et al., 2006 | ΔphoP-complemented with phoP | |
| Strain, strain background (M. tuberculosis) | ΔphoP:: phoPD71N | This study | ΔphoP-complemented with phoPD71N | |
| Strain, strain background (M. smegmatis) | M. smegmatis mc2155 | Snapper et al., 1990 | ||
| Sequence-based reagent | FPwhiB1up | This study | PCR primers | whiB1up forward primer: AATAATAAGCTTAGATGTGATGGG |
| Sequence-based reagent | RPwhiB1up | This study | PCR primers | whiB1up reverse primer: AATAATGGTACCTACCGGGAAGAA |
| Sequence-based reagent | FPwhiB1upmut | This study | PCR primers | mutagenic forward primer for whiB1upmut: GTCAAACAAGGATCACAAAACGAGATCGCCA |
| Sequence-based reagent | RPwhiB1upmut | This study | PCR primers | mutagenic reverse primer for whiB1upmut: TTGTGATCCTTGTTTGACTGTACTACACTA |
| Sequence-based reagent | FPwhiB1up1 | This study | PCR primers | whiB1up1 forward primer: AATAATAAGCTTAGATGTGATGGG |
| Sequence-based reagent | RPwhiB1up1 | This study | PCR primers | whiB1up1 reverse primer: AATAATGGTACCAGCGCGTGGGCT |
| Sequence-based reagent | FPwhiB1up2 | This study | PCR primers | whiB1up2 forward primer: AATAATGGATCCTACGTAACACTA |
| Sequence-based reagent | RPwhiB1up2 | This study | PCR primers | whiB1up2 reverse primer: AATAATGGTACCAGCGCGTGGGCT |
| Sequence-based reagent | FPwhiB1up3 | This study | PCR primers | whiB1up3 forward primer: AATAATTCCGAAGAAACGCCT |
| Sequence-based reagent | FPphoPstart | This study | PCR primers | His-tagged phoP forward primer: AATAATGGATCCATGCGGAAAGGGGT |
| Sequence-based reagent | RPphoPFLAG | This study | PCR primers | FLAG-tagged phoP reverse primer: AATAAGCTTTCACTTGTCGTCAT CGTCTTTGTAGTCTCGAGGCTCCCGCA |
| Sequence-based reagent | FPmCMR | This study | PCR primers | cmr forward primer: AATAATTGTTCGACTGACG |
| Sequence-based reagent | RPmCMR | This study | PCR primers | cmr reverse primer: AATAATCGGGTTTGTGTTGT |
| Sequence-based reagent | FPmCRP | This study | PCR primers | crp forward primer: AATAATAAGCTTGTGGACGAGATC |
| Sequence-based reagent | RPmCRP | This study | PCR primers | crp reverse primer: AATAATATCGATTTACCTCGCTCG |
| Sequence-based reagent | FPPhoPN | Pathak et al., 2010 | PCR primers | phopN forward primer: CCTGGATCCATGCGGAAAGGGGTT |
| Sequence-based reagent | RPPhoPN | This study | PCR primers | phoPN reverse primer: AATAATCTCGAGGCGTCGCAGGATGA |
| Sequence-based reagent | FPPhoPC | This study | PCR primers | phopC forward primer: AATAATCGGATCCAAGGGCAACAAGGAACCA |
| Sequence-based reagent | RPPhoPC | Pathak et al., 2010 | PCR primers | phoPC reverse primer: GGTCTCGAGTCGAGGCTCCCGCAG |
| Sequence-based reagent | FPCRP | This study | PCR primers | crp forward primer: AATAATCATATGGTGGACGAGATC |
| Sequence-based reagent | FPCRPN | This study | PCR primers | crpN forward primer: ATATATGGATCCCCCGTCGACTTCCCCC |
| Sequence-based reagent | RPCRPN | This study | PCR primers | crpN reverse primer: ATATATAAGCTTTCAACGATCGGCGAT |
| Sequence-based reagent | FPCRPC | This study | PCR primers | crpC forward primer: ATATATCATATGGTGCCCGGTCGGGT |
| Sequence-based reagent | RPCRP | This study | PCR primers | crp reverse primer: AATAATCTCGAGTTACCTCGCTCG |
| Sequence-based reagent | FPCRPG79A | This study | PCR primers | mutagenic crp forward primer: ATGTTCGCGGAGTTGTCGATCT |
| Sequence-based reagent | RPCRPG79A | This study | PCR primers | mutagenic crp reverse primer: CGACAACTCCGCGAACATGTCCGA |
| Sequence-based reagent | FPCRPT90A | This study | PCR primers | mutagenic crp forward primer: GTCCGCGCGCGTCCAGC |
| Sequence-based reagent | RPCRPT90A | This study | PCR primers | mutagenic crp reverse primer: GCTGGACGCGCGCGGACCCGGGT |
| Sequence-based reagent | FPPhoP62-64A | This study | PCR primers | mutagenic phoP forward primer: GCAGCAGCACCGGACGCGGTG |
| Sequence-based reagent | RPPhoP62-64A | This study | PCR primers | mutagenic phoP reverse primer: TGCTGCTGCCCGGGCCCGATC |
| Sequence-based reagent | FPPhoP76-78A | This study | PCR primers | mutagenic phoP forward primer: GCAGCAGCAGGCTTTGGGGTG |
| Sequence-based reagent | RPPhoP76-78A | This study | PCR primers | mutagenic phoP reverse primer: TGCTGCTGCGGGCATCATCAC |
| Sequence-based reagent | FPPhoP105-107A | This study | PCR primers | mutagenic phoP forward primer: GCAGCAGCAATCGCGGGTCTG |
| Sequence-based reagent | RPPhoP105-107A | This study | PCR primers | mutagenic phoP reverse primer: TGCTGCTGCTAGCGAGTCACG |
| Sequence-based reagent | FPPhoP110-112A | This study | PCR primers | mutagenic phoP forward primer: GCAGCAGCACTGGGTGGTGAC |
| Sequence-based reagent | RPPhoP110-112A | This study | PCR primers | mutagenic phoP reverse primer: TGCTGCTGCCGCGATCTTGTC |
| Sequence-based reagent | FPPhoP118-120A | This study | PCR primers | mutagenic phoP forward primer: GCAGCAGCAAAGCCCTTCAGT |
| Sequence-based reagent | RPPhoP118-120A | This study | PCR primers | mutagenic phoP reverse primer: TGCTGCTGCGTCGTCACCACC |
| Sequence-based reagent | FPaprART | Bansal et al., 2017 | RT-qPCR primers | gene specific primer: TTGACCATGACAGCGAGTGT |
| Sequence-based reagent | RPaprART | Bansal et al., 2017 | RT-qPCR primers | gene specific primer: TTGGACAGAAATGCAGGATG |
| Sequence-based reagent | FPcrpRT | This study | RT-qPCR primers | gene specific primer: ATCATCATCTCGGGGAAGGT |
| Sequence-based reagent | RPcrpRT | This study | RT-qPCR primers | gene specific primer: CAGCTGTTCGGAGATTTCG |
| Sequence-based reagent | FPcmrRT | This study | RT-qPCR primers | gene specific primer: ATTGGCCGAAACGTTACAAG |
| Sequence-based reagent | RPcmrRT | This study | RT-qPCR primers | gene specific primer: ACCATCGGCATCTCCAGTAG |
| Sequence-based reagent | FPicl1RT | This study | RT-qPCR primers | gene specific primer: GCTTCTACCGCACCAAGAAC |
| Sequence-based reagent | RPicl1RT | This study | RT-qPCR primers | gene specific primer: TCGAGGTGCTTTTTCCAGTT |
| Sequence-based reagent | FPphoPRT | This study | RT-qPCR primers | gene specific primer: GCCTCAAGTTCCAGGGCTTT |
| Sequence-based reagent | RPphoPRT | This study | RT-qPCR primers | gene specific primer: CCGGGCCCGATCCA |
| Sequence-based reagent | FPumaART | This study | RT-qPCR primers | gene specific primer: CGTTATGCGGCATTCTTTG |
| Sequence-based reagent | RPumaART | This study | RT-qPCR primers | gene specific primer: TGCGCAAATTTGAAGATGTC |
| Sequence-based reagent | FPwhiB1RT | This study | RT-qPCR primers | gene specific primer: CACAAGGCGGTCTGTCGT |
| Sequence-based reagent | RPwhiB1RT | This study | RT-qPCR primers | gene specific primer: GAGTCCTGGCCGGTATTCAG |
| Sequence-based reagent | FPrpoBRT | This study | RT-qPCR primers | gene specific primer: GGAGGCGATCACACCGCAGACGTT |
| Sequence-based reagent | RPrpoBRT | This study | RT-qPCR primers | gene specific primer: CCTCCAGCCCGGCACGCTCACGT |
| Sequence-based reagent | FP16SrDNA RT | This study | RT-qPCR primers | gene specific primer: CTGAGATACGGCCCAGACTC |
| Sequence-based reagent | RP16SrDNA RT | This study | RT-qPCR primers | gene specific primer: CGTCGATGGTGAAAGAGGTT |
| Sequence-based reagent | FPespAup | Anil Kumar et al., 2016 | ChIP-qPCR primers | promoter specific primer: CGTGATCTTGATACGGCTCG |
| Sequence-based reagent | RPespAup | Anil Kumar et al., 2016 | ChIP-qPCR primers | promoter specific primer: GTTGTTGGTACCCTCGGCAAGATCGGC |
| Sequence-based reagent | FPgapdhup | This study | ChIP-qPCR primers | promoter specific primer: GAGTAGGCATCAACGGGTTTG |
| Sequence-based reagent | RPgapdhup | This study | ChIP-qPCR primers | promoter specific primer: GTGCTGTTGTCGGTGATGTC |
| Sequence-based reagent | FPicl1up | This study | ChIP-qPCR primers | promoter specific primer: AATAATAAGCTTACCGGATCCGCA |
| Sequence-based reagent | RPicl1up | This study | ChIP-qPCR primers | promoter specific primer: AATAATGGTACCGTTCGTGTCC |
| Sequence-based reagent | FP16SrDNAup | Singh et al., 2014 | ChIP-qPCR primers | promoter specific primer: CTGAGATACGGCCCAGACTC |
| Sequence-based reagent | RP16SrDNAup | Singh et al., 2014 | ChIP-qPCR primers | promoter specific primer: CGTCGATGGTGAAAGAGGTT |
| Sequence-based reagent | FPsucCup | This study | ChIP-qPCR primers | promoter specific primer: GGCTGTGATTGTGAGTTGGA |
| Sequence-based reagent | RPsucCup | This study | ChIP-qPCR primers | promoter specific primer: GCGAATAACTCCTTGGCTTG |
| Sequence-based reagent | FPumaAup | This study | ChIP-qPCR primers | promoter specific primer: TGTTGCTGCGTATGGTTGAG |
| Sequence-based reagent | RPumaAup | This study | ChIP-qPCR primers | promoter specific primer: AATCGATTGCGACTCTTCGT |
| Sequence-based reagent | FPwhiB1up1 | This study | ChIP-qPCR primers | promoter specific primer: AATAATAAGCTTAGATGTGATGGG |
| Sequence-based reagent | RPwhiB1up1 | This study | ChIP-qPCR primers | promoter specific primer: AATAATGGTACCAGCGCGTGGGCT |
| Recombinant DNA reagent | pET-phoP | Gupta et al., 2009 | Plasmid DNA | His6 tagged-PhoP residues 1–247 cloned in pET15b |
| Recombinant DNA reagent | pGEX-phoP | Gupta et al., 2009 | Plasmid DNA | PhoP residues 1–247 cloned in pGEX-4T-1 |
| Recombinant DNA reagent | pGEX-phoPLAla5 | This study | Plasmid DNA | G142-P146 residues mutated to A in phoP of pGEX-phoP |
| Recombinant DNA reagent | pGEX-phoPN | This study | Plasmid DNA | PhoP residues 1–141 cloned in pGEX-4T-1 |
| Recombinant DNA reagent | pGEX-phoPC | This study | Plasmid DNA | PhoP residues 141–247 cloned in pGEX-4T-1 |
| Recombinant DNA reagent | pGEX-phoP(62-64)Ala | This study | Plasmid DNA | E62-R64 residues mutated to A in phoP of pGEX-phoP |
| Recombinant DNA reagent | pGEX-phoP(76-78)Ala | This study | Plasmid DNA | G76-D78 residues mutated to A in phoP of pGEX-phoP |
| Recombinant DNA reagent | pGEX-phoP(105-107)Ala | This study | Plasmid DNA | Q105-K107 residues mutated to A in phoP of pGEX-phoP |
| Recombinant DNA reagent | pGEX-phoP(110-112)Ala | This study | Plasmid DNA | G110-T112 residues mutated to A in phoP of pGEX-phoP |
| Recombinant DNA reagent | pGEX-phoP(118-120)Ala | This study | Plasmid DNA | Y118-T120 residues mutated to A in phoP of pGEX-phoP |
| Recombinant DNA reagent | pME1mL1-phoPb | Goyal et al., 2011 | Plasmid DNA | PhoP residues 1–247 cloned in pME1mL1 |
| Recombinant DNA reagent | pSM128c | Dussurget et al., 1999 | Plasmid DNA | Integrative promoter probe vector for mycobacteria |
| Recombinant DNA reagent | pSM-whiB1up | This study | Plasmid DNA | whiB1up-lacZ fusion in pSM128 |
| Recombinant DNA reagent | pSM-whiB1upmut | This study | Plasmid DNA | pSM-whiB1up carrying changes in the PhoP binding site |
| Recombinant DNA reagent | pUAB400d | Singh et al., 2006 | Plasmid DNA | Integrative mycobacteria - E. coli shuttle plasmid, Kanr |
| Recombinant DNA reagent | pUAB400-phoP | Singh et al., 2014 | Plasmid DNA | PhoP residues 1–247 cloned in pUAB400 |
| Recombinant DNA reagent | pUAB300b | Singh et al., 2006 | Plasmid DNA | Episomal mycobacteria - E. coli shuttle plasmid, Hygr |
| Recombinant DNA reagent | pUAB300-crp | This study | Plasmid DNA | CRP residues 1–224 cloned in pUAB300 |
| Recombinant DNA reagent | pUAB300-cmr | This study | Plasmid DNA | CMR residues 1–244 cloned in pUAB300 |
| Recombinant DNA reagent | pET-28cd | Novagen | Plasmid DNA | E. coli cloning vector, Kanr |
| Recombinant DNA reagent | pET-crpd | This study | Plasmid DNA | His6 tagged-CRP residues 1–224 cloned in pET-28c |
| Recombinant DNA reagent | pET-crpN | This study | Plasmid DNA | His6 tagged -CRP residues 28–116 cloned in pET-28c |
| Recombinant DNA reagent | pET-crpC | This study | Plasmid DNA | His6 tagged-CRP residues 146–224 cloned in pET-28c |
| Recombinant DNA reagent | p19Kprob | De Smet et al., 1999 | Plasmid DNA | Mycobacteria expression vector |
| Recombinant DNA reagent | p19Kpro-phoP | Anil Kumar et al., 2016 | Plasmid DNA | His6-tagged PhoP residues 1–247 cloned in p19Kpro |
| Recombinant DNA reagent | p19Kpro-phoPFLAG | This study | Plasmid DNA | FLAG-tagged PhoP residues 1–247 cloned in p19Kpro |
| Antibody | Anti-CRP (rabbit polyclonal) | AlphaOmega Sciences (This study) | 1:5000 | |
| Antibody | Anti-FLAG (rabbit polyclonal) | Invitrogen | #PA1-984B | 1:3000 |
| Antibody | Anti-GST (goat polyclonal) | GE Healthcare | # 27457701V | 1:5000 |
| Antibody | Anti-His (mouse monoclonal) | Invitrogen | #MA1-21315 | 1:5000 |
| Antibody | Anti-PhoP (rabbit polyclonal) | AlphaOmegaSciences (This study) | 1:3000 |
Additional files
-
Supplementary file 1
Sequence-based nucleic acid reagents.
(a) Oligonucleotide primers used for amplification and cloning in this study. (b) Plasmids used in this study. (c) Sequence of oligonucleotide primers used in aRT-qPCR and bChIP-qPCR experiments reported in this study.
- https://cdn.elifesciences.org/articles/80965/elife-80965-supp1-v1.docx
-
Source data 1
Source data for figures and figure supplements.
- https://cdn.elifesciences.org/articles/80965/elife-80965-data1-v1.zip
-
MDAR checklist
- https://cdn.elifesciences.org/articles/80965/elife-80965-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Convergence of two global regulators to coordinate expression of essential virulence determinants of Mycobacterium tuberculosis
eLife 11:e80965.
https://doi.org/10.7554/eLife.80965




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}