The cardiopharyngeal mesoderm contributes to lymphatic vessel development in mouse
- Department of Physiological Chemistry and Metabolism, Graduate School of Medicine, The University of Tokyo, Japan
- Department of Pathology and Matrix Biology, Graduate School of Medicine, Mie University, Japan
- Department of Animal Nursing Science, Yamazaki University of Animal Health Technology, Japan
- Heart Center, Department of Pediatric Cardiology, Tokyo Women's Medical University, Japan
- Laboratory for Cell Asymmetry, RIKEN Center for Biosystems Dynamics Research, Japan
Figures
Figure 1 with 2 supplements
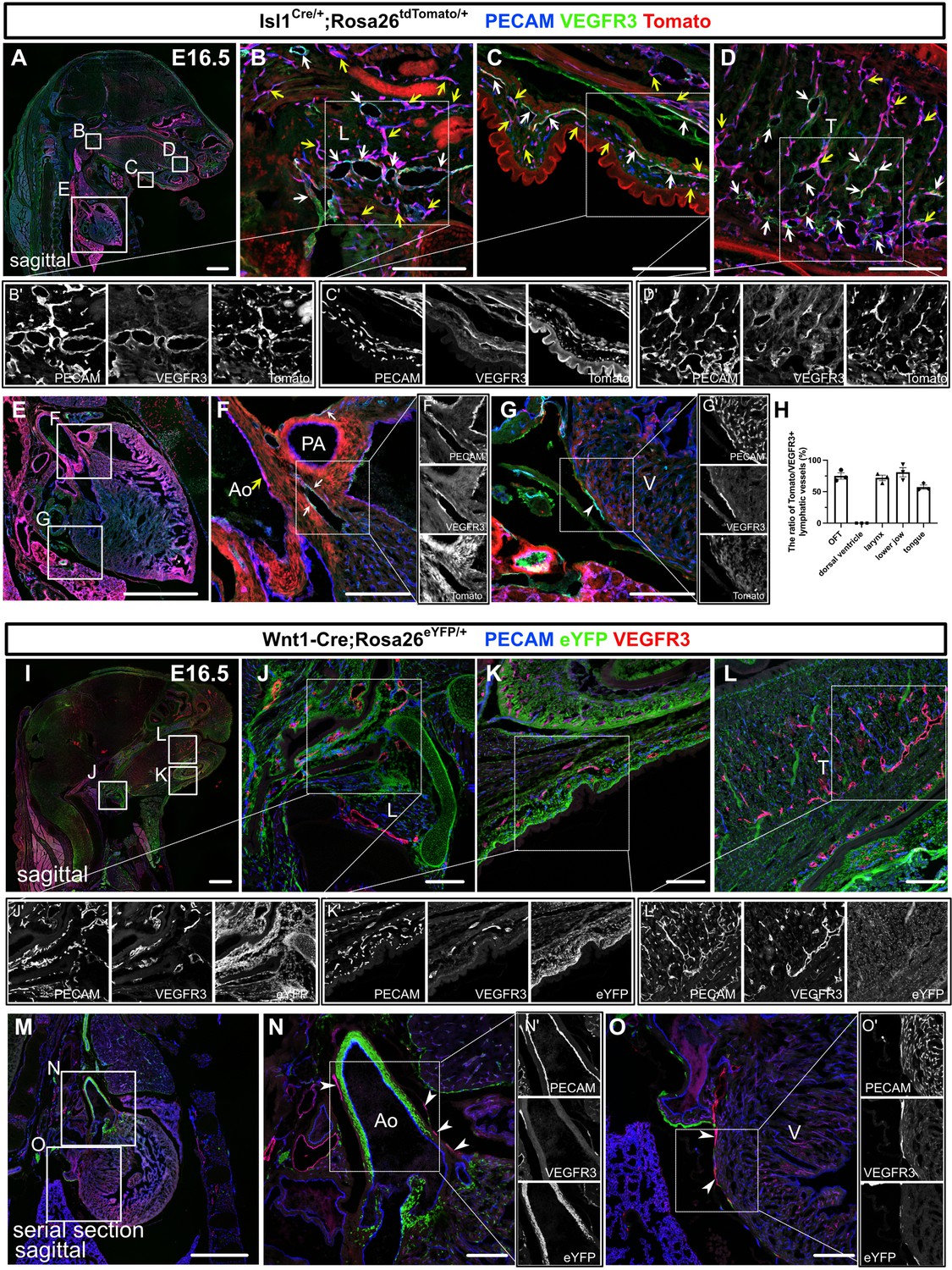
Isl1+ lineages contribute to cranial and cardiac lymphatic vessels.
(A–G’) Sagittal sections of Isl1Cre/+;Rosa26tdTomato/+ embryos, in which platelet endothelial cell adhesion molecule (PECAM), tdTomato, and vascular endothelial growth factor receptor 3 (VEGFR3) were labeled at embryonic day (E) 16.5, are shown. (A–F) tdTomato colocalized with PECAM/VEGFR3 (white arrows) and PECAM (yellow arrows) in and around the larynx (B), the skin of the lower jaw (C), the tongue (D), and the cardiac outflow tracts (F) (n=3). (G) tdTomato did not colocalize with VEGFR3 in the LECs on the dorsal side of the ventricles (white arrowhead, n=3). (H) The results of quantitative analysis of the percentage of the area in tdTomato+/VEGFR3+ lymphatic vessels among all VEGFR3+ lymphatic vessels are shown. (I–O’) Sagittal sections of Wnt1-Cre;Rosa26eYFP/+ embryos, in which PECAM, eYFP, and VEGFR3 were labeled at E16.5, are shown. There were no eYFP+/VEGFR3+ lymphatic vessels in or around the larynx, the skin of the lower jaw, the tongue, or the heart (the white arrowheads indicate cardiac lymphatic vessels around the cardiac outflow tracts and the dorsal side of the ventricles; n=3). Each dot represents a value obtained from one sample. L, larynx; T, tongue; Ao, aorta; PA, pulmonary artery; V, ventricle. Scale bars, 100 μm (B–D, F, G, J–L, N, O), 1 mm (A, E, I, M).
-
Figure 1—source data 1
Quantification of Isl1+ lineages contribution to craniofacial and cardiac lymphatic vessels.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig1-data1-v2.xlsx
Figure 1—figure supplement 1
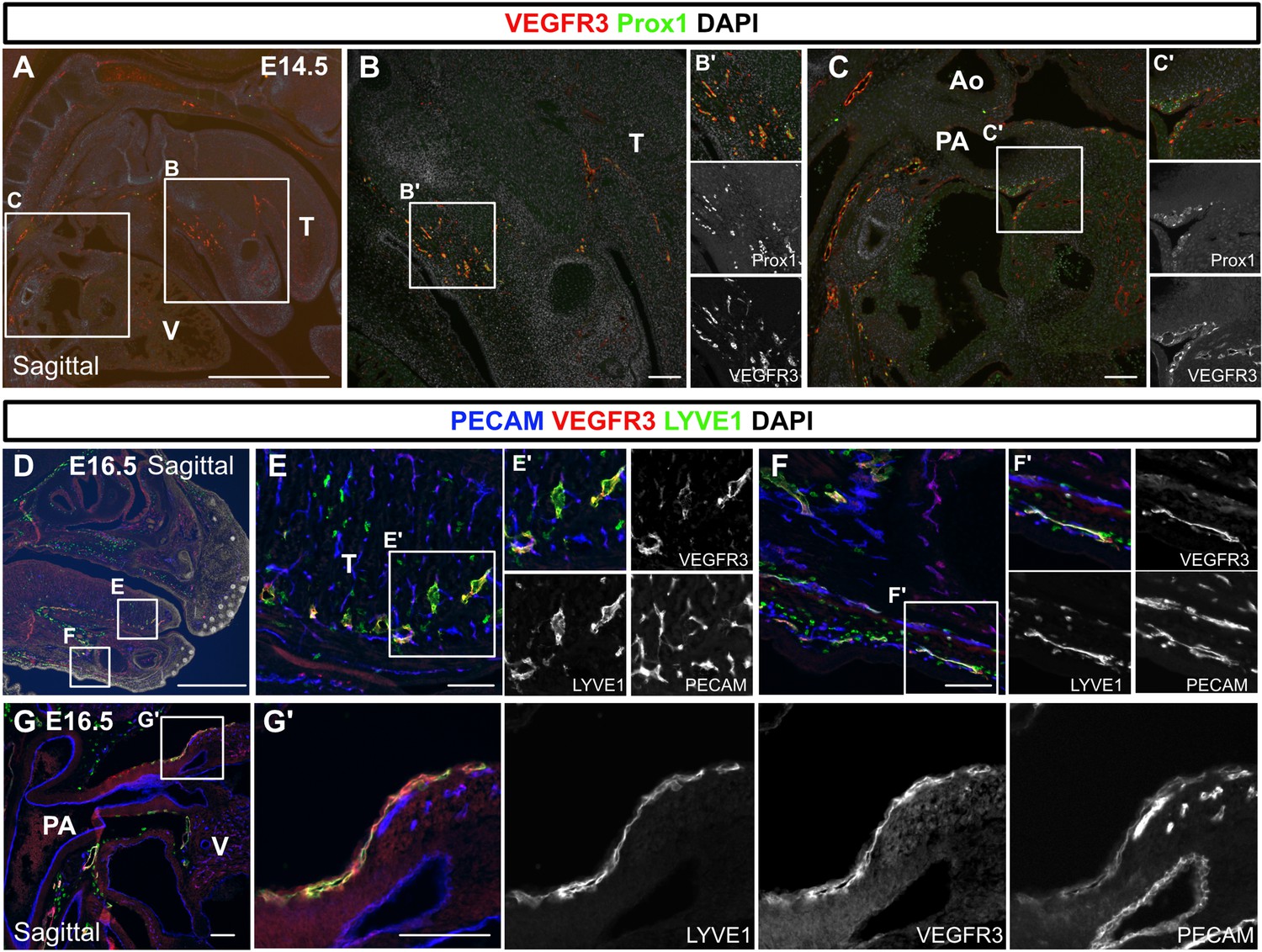
Lymphatic vessel endothelial hyaluronan receptor 1 (LYVE1) and vascular endothelial growth factor receptor 3 (VEGFR3) are co-expressed in lymphatic vessels in the tongue, facial skin, and the outflow tracts.
(A–G’) Sagittal sections were stained for VEGFR3 and Prox1 at embryonic day (E) 14.5 (n=2) (A–C’), with additional platelet endothelial cell adhesion molecule (PECAM) staining at E16.5 (D–G’). At E14.5, VEGFR3 and Prox1 were co-expressed in the lymphatic endothelial cells in the tongue and skin of the lower jaw (B, B’) and around the pulmonary artery (C, C’). At E16.5, VEGFR3 and LYVE1 were co-expressed in the lymphatic endothelial cells in the tongue and skin of the lower jaw (E, E’, F, F’) and around the pulmonary artery (G, G’) (n=2). T, tongue; V, ventricle; Ao, aorta; PA, pulmonary artery; cardiac outflow tract. Scale bars, 100 μm (B, C, E, F, G, G’), 1 mm (A, D).
Figure 1—figure supplement 2
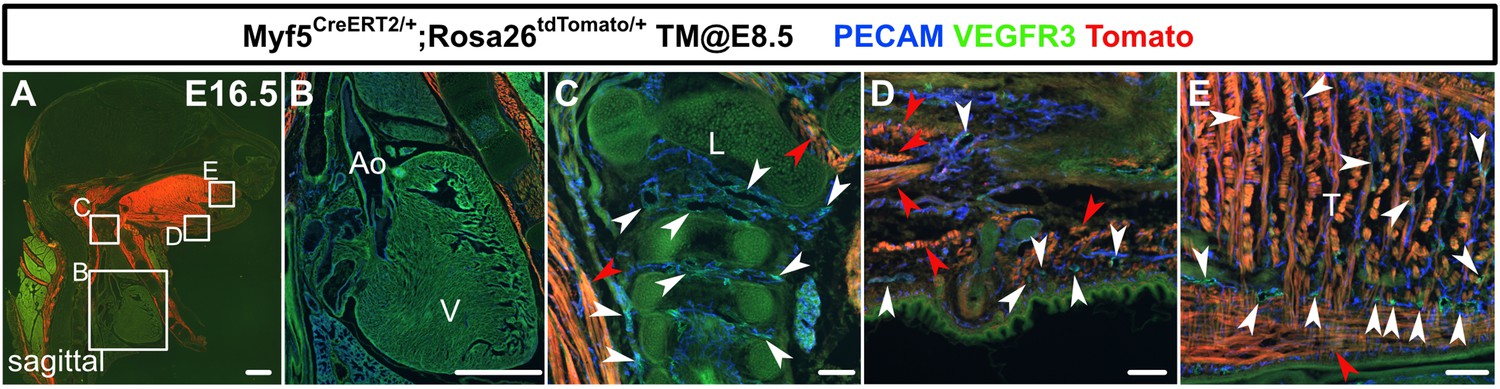
Myf5+ lineages do not generate lymphatic endothelial cells (LECs) in the cranial or cardiac regions.
(A–E) Sagittal sections of Myf5CreERT2/+;Rosa26tdTomato/+ embryos, in which platelet endothelial cell adhesion molecule (PECAM), tdTomato, and vascular endothelial growth factor receptor 3 (VEGFR3) were labeled at embryonic day (E) 16.5, are shown. Tamoxifen was administered at E8.5. There were no tdTomato+/VEGFR3+ lymphatic vessels in or around the heart (B), the larynx (C), the skin of the lower jaw (D), or the tongue (E). (White arrowheads indicate tdTomato-/VEGFR3+ lymphatic vessels. Red arrowheads indicate skeletal muscles.) Scale bars, 100 μm (C–E), 1 mm (A, B).
Figure 2 with 1 supplement
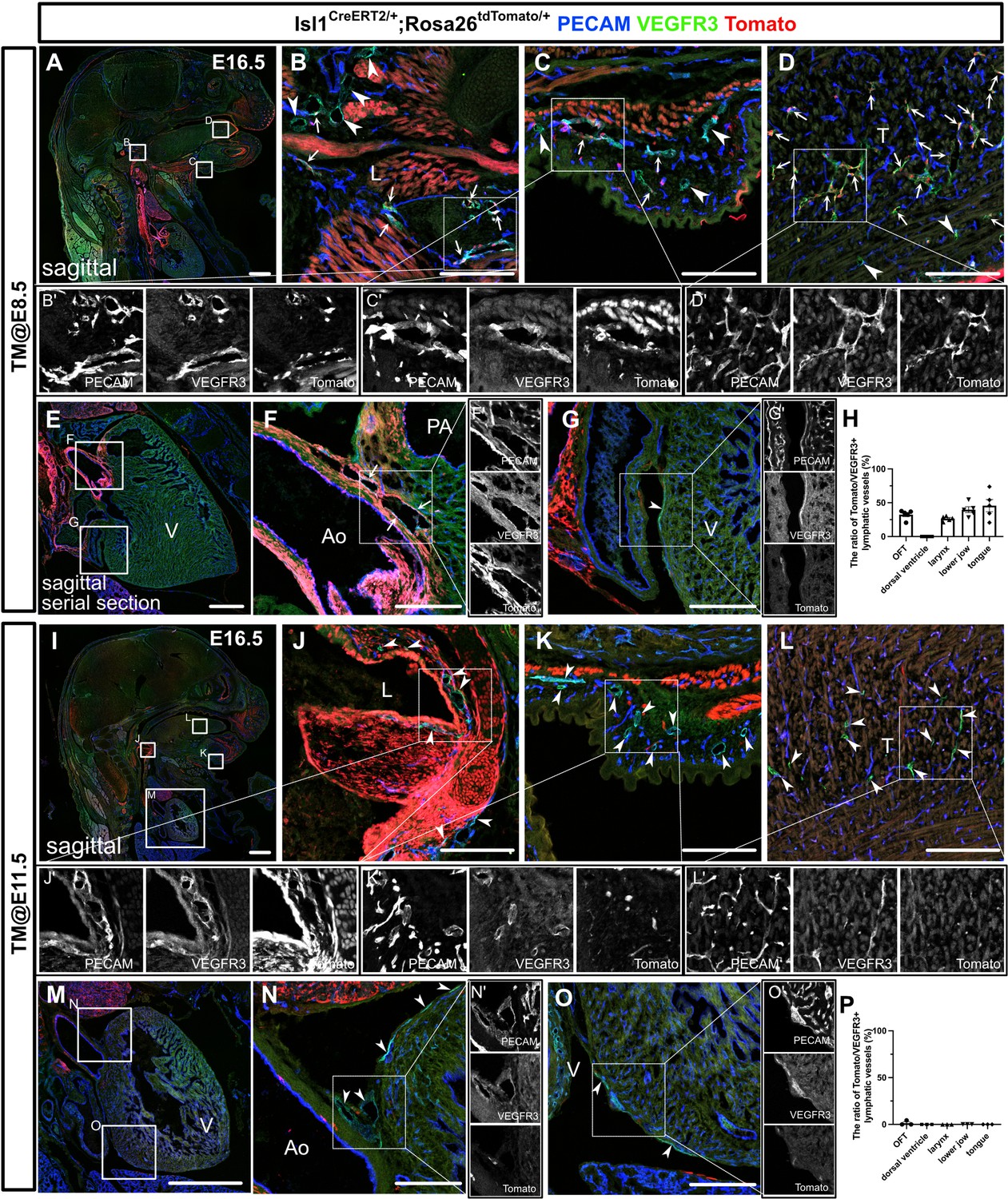
The differentiation of Isl1+ cardiopharyngeal mesoderm (CPM) cells into lymphatic endothelial cells (LECs) occurs during the early embryonic period.
(A–G’, I–O’) Sagittal sections of Isl1CreERT2/+;Rosa26tdTomato/+ embryos, in which platelet endothelial cell adhesion molecule (PECAM), tdTomato, and vascular endothelial growth factor receptor 3 (VEGFR3) were labeled at embryonic day (E) 16.5, are shown. Tamoxifen was administered at E8.5 (A–G’) or E11.5 (I–O’). (A–G’) Both tdTomato+ (white arrows) and tdTomato- (white arrowheads) VEGFR3+ lymphatic vessels were observed in and around the larynx (B), the skin of the lower jaw (C), the tongue (D), and the cardiac outflow tracts (F) (n=5). (G) tdTomato did not colocalize with VEGFR3 in the LECs on the dorsal side of the ventricles (white arrowhead, n=5). (I–O’) Almost all of the VEGFR3+ lymphatic vessels in and around the larynx (J), the skin of the lower jaw (K), the tongue (L), the cardiac outflow tracts (N), and the dorsal side of the ventricles (O) were tdTomato- (white arrows, n=3). (H, P) The results of a quantitative analysis of the percentage of the area of tdTomato+/VEGFR3+ lymphatic vessels among all VEGFR3+ lymphatic vessels are shown. Tamoxifen was administered at E8.5 (H) or E11.5 (P). Each dot represents a value obtained from one sample. L, larynx; T, tongue; Ao, aorta; PA, pulmonary artery; V, ventricle. Scale bars, 100 μm (B–D, F, G, J–L, N, O), 1 mm (A, E, I, M).
-
Figure 2—source data 1
Quantification of Isl1+ lineages contribution to craniofacial and cardiac lymphatic vessels at embryonic day (E) 16.5 with tamoxifen treatment at E8.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig2-data1-v2.xlsx
-
Figure 2—source data 2
Quantification of Isl1+ lineages contribution to craniofacial and cardiac lymphatic vessels at embryonic day (E) 16.5 with tamoxifen treatment at E11.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig2-data2-v2.xlsx
Figure 2—figure supplement 1

The Isl1+ cardiopharyngeal mesoderm (CPM) broadly contributes to cranial and cardiac lymphatic vessels.
(A–H) Sagittal sections of Isl1CreERT2/+;Rosa26tdTomato/+ embryos, in which platelet endothelial cell adhesion molecule (PECAM), tdTomato, and vascular endothelial growth factor receptor 3 (VEGFR3) were labeled at embryonic day (E) 16.5, are shown. Tamoxifen was administered at E6.5. (B, D–F) tdTomato+/VEGFR3+ lymphatic vessels were observed in and around the cardiac outflow tracts (B), the larynx (D), the skin of the lower jaw (E), and the tongue (F) (n=2). (C, G, and H) tdTomato+/VEGFR3+ lymphatic vessels were not observed on the dorsal side of the ventricles (C) or in the back skin (G and H) (white arrowheads) (n=2). (I) The results of a quantitative analysis of the percentage of tdTomato+/VEGFR3+ lymphatic vessels among all VEGFR3+ lymphatic vessels. Each dot represents a value obtained from one sample. Ao, aorta; PA, pulmonary artery; V, ventricle; L, larynx; T, tongue. Scale bars, 100 μm (B–H), 1 mm (A).
-
Figure 2—figure supplement 1—source data 1
Quantification of Isl1+ lineages contribution to craniofacial and cardiac lymphatic vessels at embryonic day (E) 16.5 with tamoxifen treatment at E6.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig2-figsupp1-data1-v2.xlsx
Figure 3 with 1 supplement
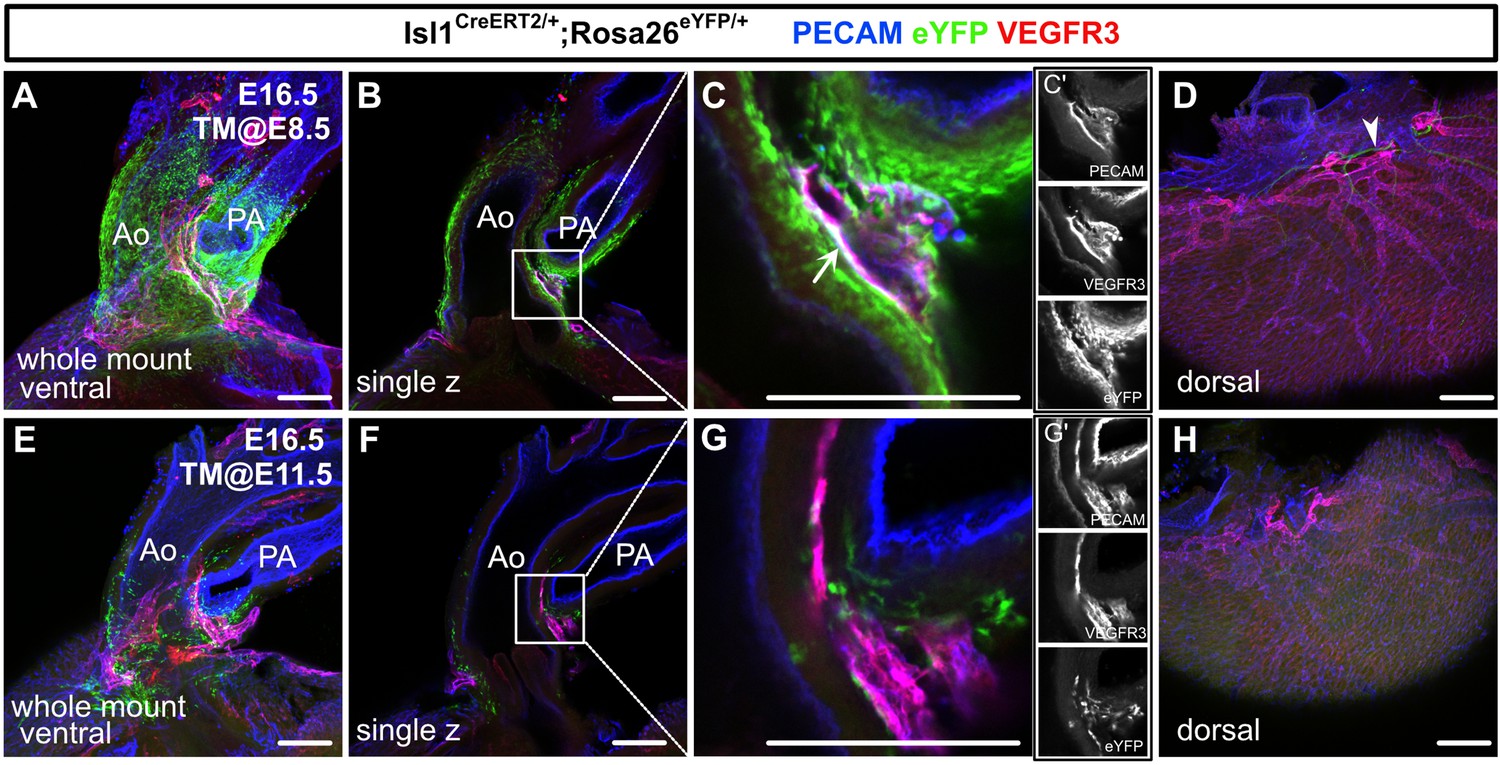
Isl1+ cardiopharyngeal mesoderm (CPM) cells contribute to cardiac lymphatic vessel development.
(A–H) Whole-mount confocal images of Isl1CreERT2/+;Rosa26eYFP/+ hearts, in which platelet endothelial cell adhesion molecule (PECAM), eYFP, and vascular endothelial growth factor receptor 3 (VEGFR3) were labeled at embryonic day (E) 16.5, are shown. Tamoxifen was administered at E8.5 (A–D) or E11.5 (E–H). (A–D) Many eYFP+ cells were observed around the cardiac outflow tracts, and they contributed to lymphatic vessels (white arrow) (n=9/9 hearts [100%]) (A–C’). (D) There were no eYFP+ lymphatic vessels on the dorsal side of the ventricles (n=9/9 hearts [100%]). The cardiac nerves were also positive for eYFP after tamoxifen treatment at E8.5 (white arrowhead) (D). (E–H) Fewer eYFP+ cells were observed around the cardiac outflow tracts (E–G’), and there were no eYFP+ lymphatic vessels around the cardiac outflow tracts (F) or on the dorsal side of the ventricles (n=0/8) (H). Ao, aorta; PA, pulmonary artery. Scale bars, 100 μm (A–C, E–G), 500 μm (D, H).
Figure 3—figure supplement 1
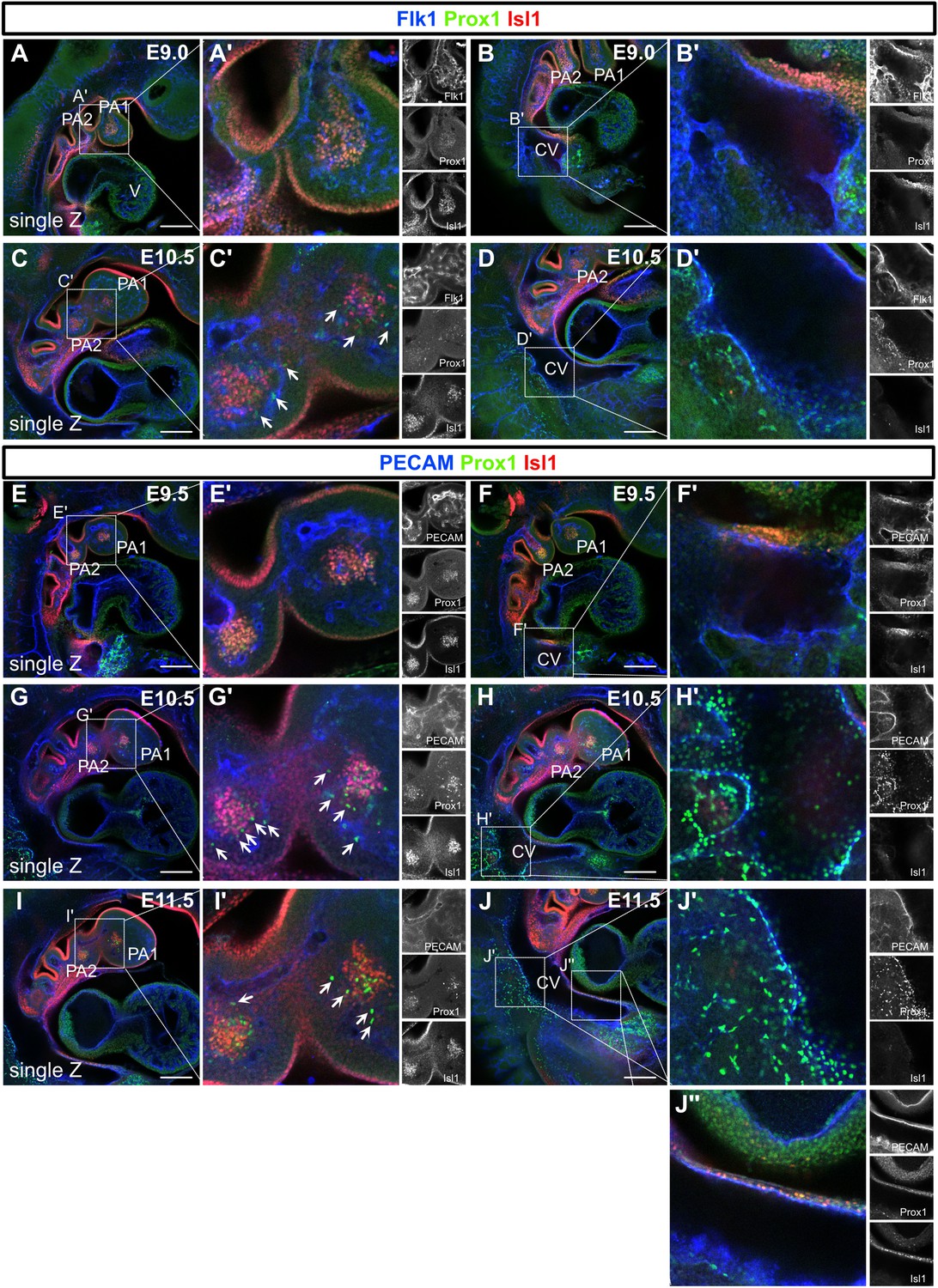
Isl1 is not expressed in the endothelium in the first and second pharyngeal arches and cardinal vein at early embryonic period.
(A–J’’) Whole-mount images of embryos, in which Isl1, Prox1, Flk1, or platelet endothelial cell adhesion molecule (PECAM) were labeled at embryonic day (E) 9.0, E9.5, E10.5, and E11.5 are shown. Isl1 was not expressed in the endothelium in the first and second pharyngeal arches and cardinal vein. (C’, G’, I’) Isl1+/Prox1+ cells were observed around cores of the first and second pharyngeal arches at E9.0 to E11.5. Isl1-/Prox1+ cells began to migrate from the first and second pharyngeal arches (white arrows). V, ventricle; PA1, first pharyngeal arch; PA2, second pharyngeal arch; CV, cardinal vein. Scale bars, 100 μm (A, B, C, D, E, F, G, H, I, J).
Figure 4
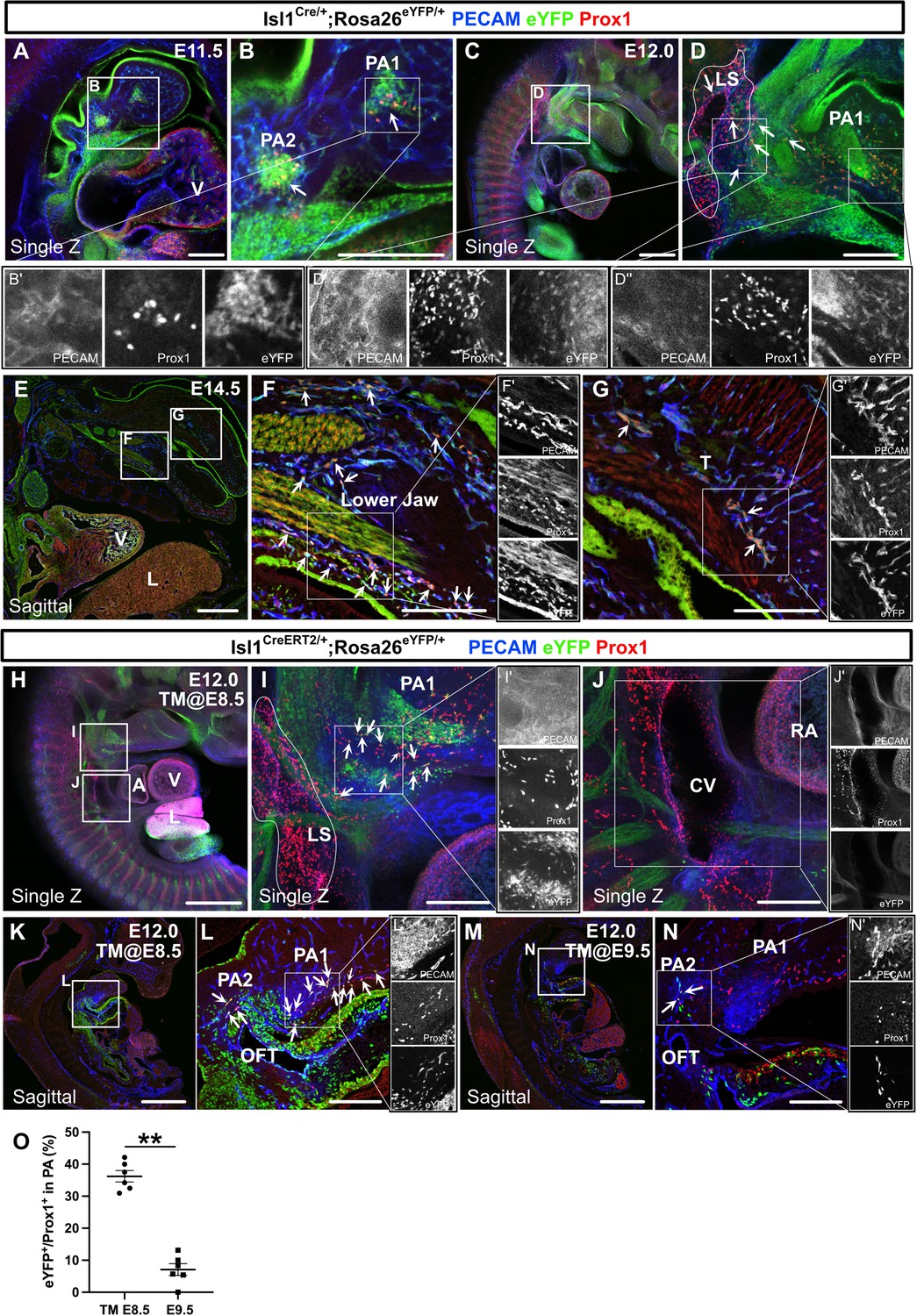
The spatiotemporal development of Isl1+ cardiopharyngeal mesoderm (CPM)-derived lymphatic endothelial cells (LECs).
(A–G’) Whole-mount and sagittal section images of Isl1Cre/+;Rosa26eYFP/+ embryos, in which platelet endothelial cell adhesion molecule (PECAM), eYFP, and Prox1 were labeled at embryonic day (E) 11.5, E12.0, or E14.5, are shown. (A, B) eYFP+/Prox1+ cells were observed around the cores of the first and second pharyngeal arches at E11.5 (white arrowheads). (C–D’’) eYFP+/Prox1+ LECs migrated from the first and second pharyngeal arches to the lymph sac-forming domain (white dotted region) adjacent to the anterior cardinal vein at E12.0 (white arrows). (E–G’) Some of the eYFP+/Prox1+ LECs expressed PECAM and formed lymphatic capillaries in the lower jaw and tongue at E14.5 (white arrows). (H–N’) Whole-mount and sagittal section images of Isl1CreERT2/+;Rosa26eYFP/+ embryos, in which PECAM, eYFP, and Prox1 were labeled at E12.0, are shown. Tamoxifen was administered at E8.5 (H–L’) or E9.5 (M–N’). (I, I’) eYFP+/Prox1+ LECs (white arrows) migrated from the first and second pharyngeal arches to the lymph sac-forming domain (white dotted region) at E12.0. (J) There were no eYFP+/Prox1+ cells in or around the cardinal vein (n=4). (K–N) eYFP+/Prox1+/PECAM+ LECs were seen in the first and second pharyngeal arches of the embryos at E12.0, when tamoxifen was administered at E8.5 or E9.5 (white arrows), although the number of these cells was decreased in the E9.5 group (M–N’). (O) The results of a quantitative analysis of the percentage of eYFP+/Prox1+ cells among Prox1+ cells in the first and second pharyngeal arches at E12.0 after tamoxifen treatment at E8.5 (the number of eYFP+/Prox1+ cells [10.83 (mean)±1.249 (SEM)]: Prox1+ cells [30.83±4.549]) or E9.5 (the number of eYFP+/Prox1+ cells [2.833±1.108]: Prox1+ cells [35.50±5.847]) are shown. **p=0.0022. All of the data are presented as the mean ± SEM, and statistical analyses were performed using the non-parametric Mann-Whitney U-test. V, ventricle; PA1, first pharyngeal arch; PA2, second pharyngeal arch; CV, cardinal vein; LS, lymph sac-forming domain; L, liver; T, tongue; RA, right atrium; OFT, cardiac outflow tract. Scale bars, 100 μm (A, B, D, F, G, I, J, L, N), 500 μm (C, E, H, K, M); **p<0.01.
-
Figure 4—source data 1
Quantification of eYFP+/Prox1+ cells among Prox1+ cells in the first and second pharyngeal arches at embryonic day (E) 12.0 with tamoxifen treatment at E8.5 and E9.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig4-data1-v2.xlsx
Figure 5 with 1 supplement
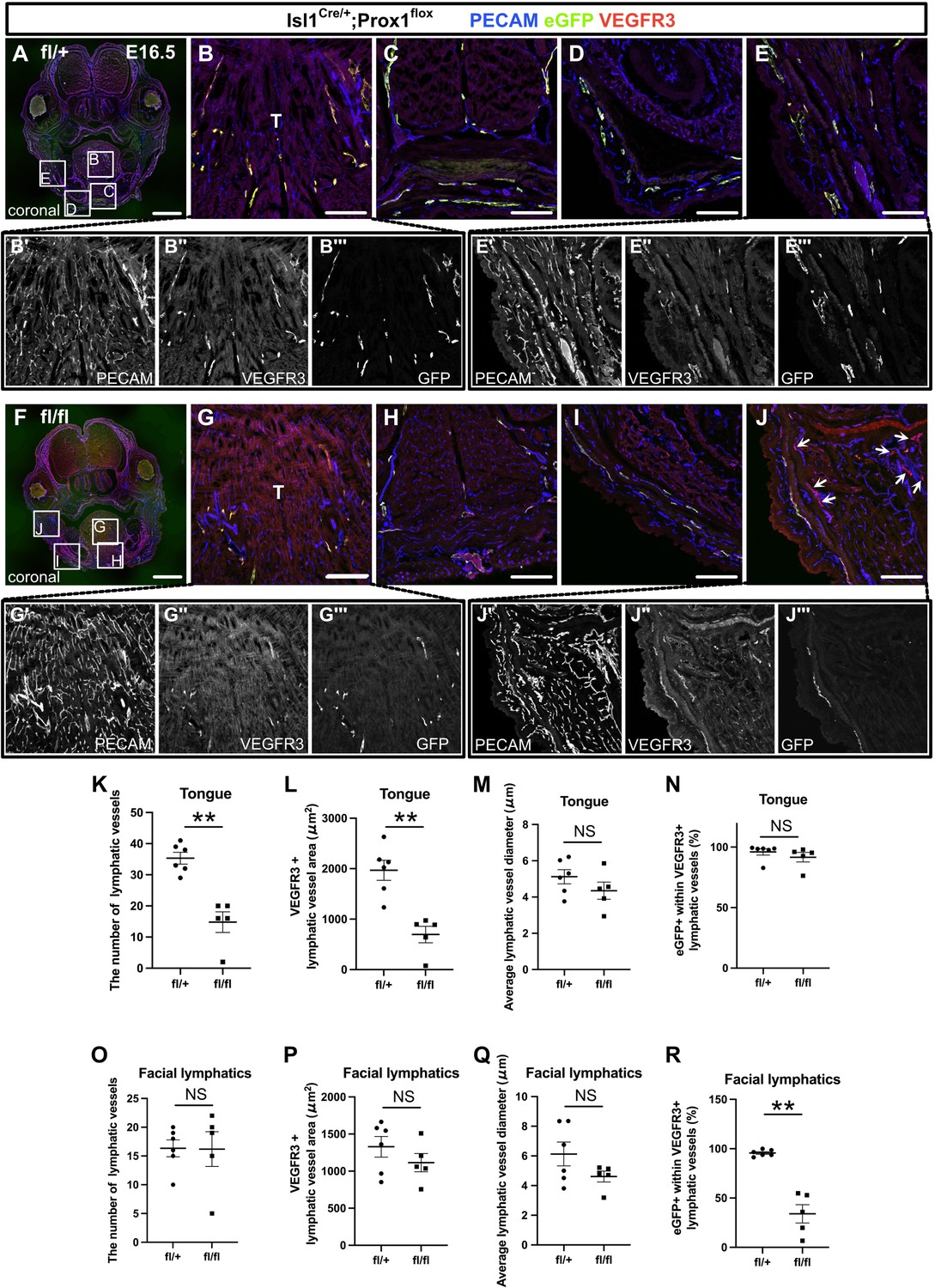
The inactivation of Prox1 in Isl1+ lineages confirmed the contribution of the cardiopharyngeal mesoderm (CPM) to cranial lymphatic vessel development.
(A–J”’) Coronal sections of Isl1Cre/+;Prox1fl/+ and Isl1Cre/+;Prox1fl/fl mouse embryos, in which platelet endothelial cell adhesion molecule (PECAM), eGFP, and vascular endothelial growth factor receptor 3 (VEGFR3) were labeled at embryonic day (E) 16.5, are shown. (J) The number of eYFP-/VEGFR3+ lymphatic vessels in facial skin was increased in the Isl1Cre/+; Prox1fl/fl homozygous mice (white arrows). (K–R) The results of quantitative analysis of lymphatic vessel formation in the tongue (K–N) and facial skin (O–R) are shown. (**p=0.0043 (K, L, R)). All the data are presented as the mean ± SEM, and statistical analyses were performed using the non-parametric Mann-Whitney U-test. Each dot represents a value obtained from one sample. **p<0.01; T, tongue. Scale bars, 100 μm (B–E, G–J), 1 mm (A, F).
-
Figure 5—source data 1
Quantification of lymphatic vessels phenotypes in Isl1Cre/+;Prox1fl/+ and Isl1Cre/+;Prox1fl/fl mouse embryos at embryonic day (E) 16.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig5-data1-v2.xlsx
Figure 5—figure supplement 1
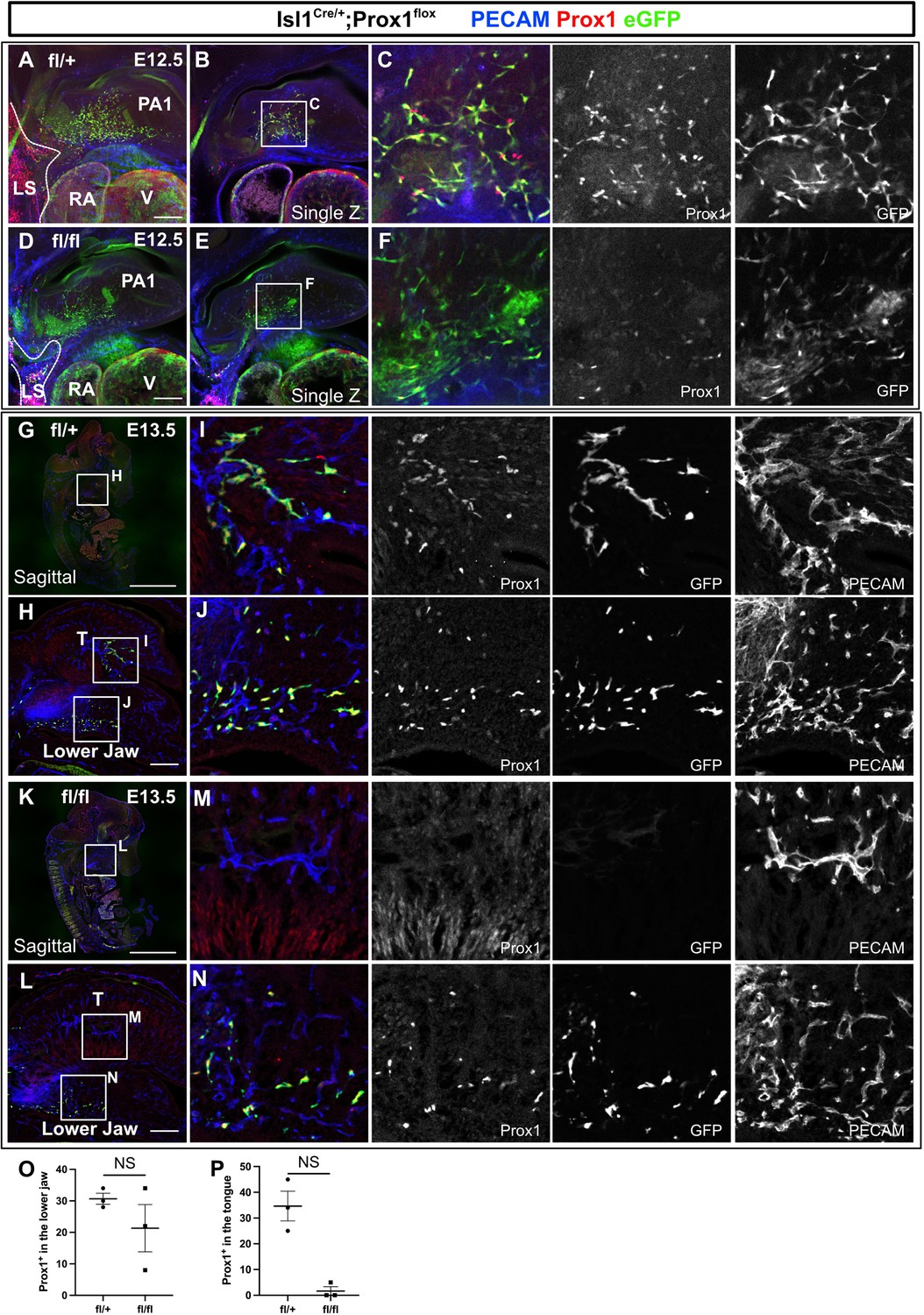
Prox1 expression level was decreased in Isl1+ lymphatic endothelial cells (LECs) in Isl1Cre/+;Prox1fl/fl embryos.
(A–N) Whole-mount and sagittal section images of Isl1Cre/+;Prox1fl/+ and Isl1Cre/+;Prox1fl/fl mouse embryos, in which platelet endothelial cell adhesion molecule (PECAM), eGFP, and Prox1 were labeled at embryonic day (E) 12.5 (A–F), or E13.5 (G–P). (A–C) eGFP+/Prox1+ cells were observed around the cores of the first pharyngeal arch in Isl1Cre/+;Prox1fl/+ embryos at E12.5. (D–F) eGFP+/Prox1- cells were observed around the cores of the first pharyngeal arch in Isl1Cre/+;Prox1fl/fl embryo. The numbers of Prox1+ cells in the first pharyngeal arch were 71.5 (mean)±3.5 (SEM) and 7.0±1.0 in Isl1Cre/+;Prox1fl/+ (n=2) and Isl1Cre/+;Prox1fl/fl (n=2) embryos, respectively (A–F). (G–J) PECAM+/eGFP+/Prox1+ lymphatic capillaries were observed in the tongue (H, I) and the lower jaw (H, J) of Isl1Cre/+;Prox1fl/+ embryos at E13.5. (K–N) Lymphatic capillaries were not observed in the tongue of Isl1Cre/+;Prox1fl/fl embryos at E13.5 (K, M). Decreased PECAM+/eGFP+/Prox1+ lymphatic capillaries were observed in the lower jaw of Isl1Cre/+;Prox1fl/fl embryos at E13.5 (L, N). (O, P) Quantitative analysis of the number of Prox1+ cells in the tongue and the lower jaw. All of the data are presented as mean ± SEM, and statistical analyses were performed using the non-parametric Mann-Whitney U-test. V, ventricle; PA1, first pharyngeal arch; LS, lymph sac-forming domain; T, tongue; RA, right atrium. Scale bars, 100 μm (A, D, H, L), 1 mm (G, K).
-
Figure 5—figure supplement 1—source data 1
Quantification of Prox1+ cells in the first pharyngeal arch in Isl1Cre/+; Prox1fl/+ and Isl1Cre/+;Prox1fl/fl mouse embryos at embryonic day (E) 12.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig5-figsupp1-data1-v2.xlsx
-
Figure 5—figure supplement 1—source data 2
Quantification of the number of Prox1+ cells in the tongue and the lower jaw of Isl1Cre/+;Prox1fl/+ and Isl1Cre/+;Prox1fl/fl mouse embryos at embryonic day (E) 13.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig5-figsupp1-data2-v2.xlsx
Figure 6 with 2 supplements
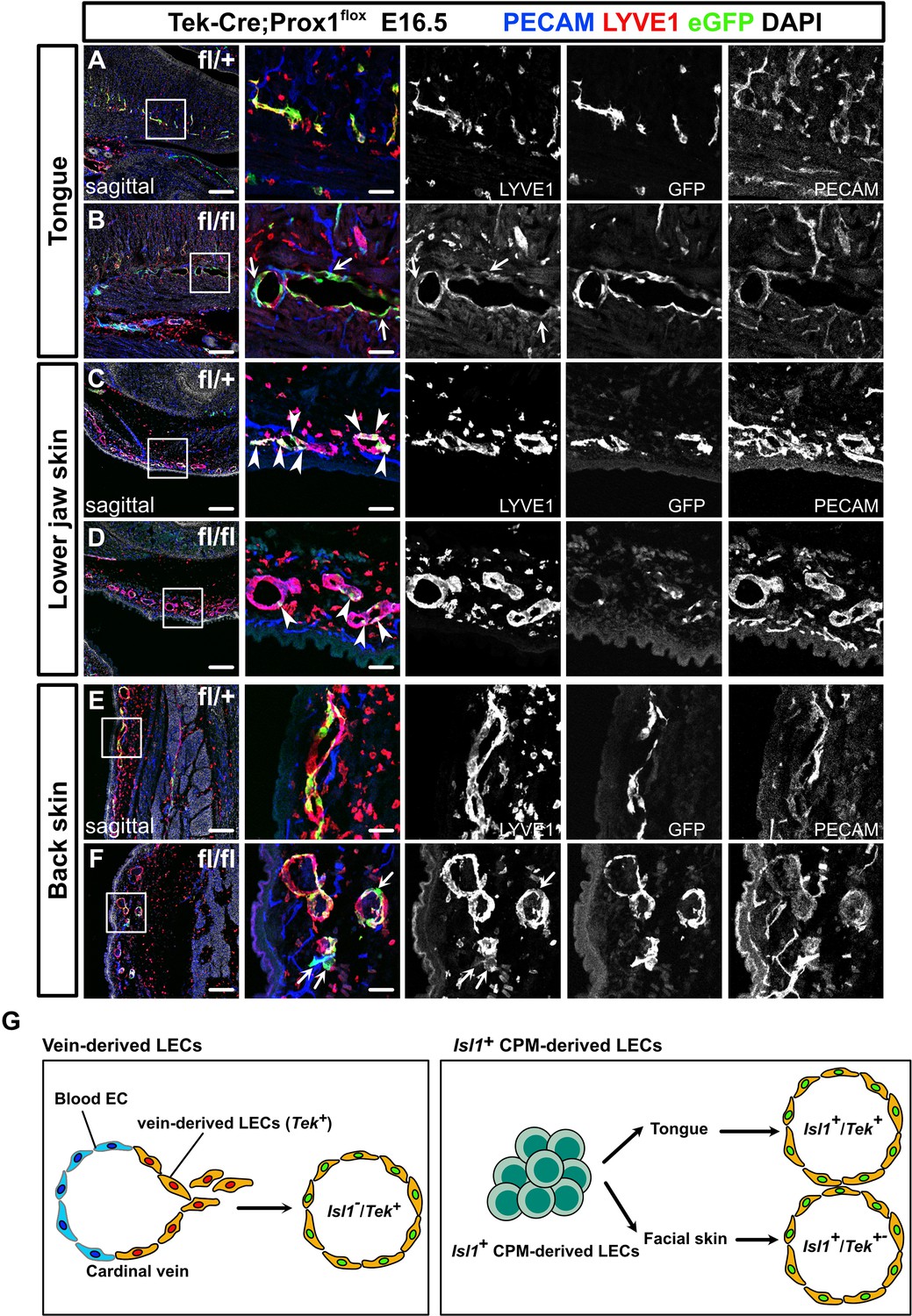
Prox1 knockdown in Tek+ lineages revealed regional differences in lymphatic vessel development.
(A–F) Sagittal sections of Tek-Cre;Prox1fl/+ or Tek-Cre;Prox1fl/fl mouse embryos, in which platelet endothelial cell adhesion molecule (PECAM), eGFP, lymphatic vessel endothelial hyaluronan receptor 1 (LYVE1), and DAPI were labeled at embryonic day (E) 16.5, are shown. (A, B) In the tongue, the number of LYVE1-/eGFP+/PECAM+ cells in lymphatic vessels was increased in the Tek-Cre;Prox1fl/fl embryos (white arrows). (C, D) In the skin of the lower jaw, the contribution of eGFP+ cells to lymphatic vessels was small in both the Tek-Cre; Prox1fl/+ and Tek-Cre;Prox1fl/fl embryos (white arrows). (E, F) LYVE1-/eGFP+/PECAM+ cells were observed in the back skin of the Tek-Cre;Prox1fl/fl embryos (white arrows). (G) Schematic representation of lineage classification in venous-derived LECs and Isl1+ CPM-derived LECs. The LECs in the tongue are derived from the Isl1+/Tek+ lineage, whereas the LECs in facial skin, including the skin on the lower jaw, are derived from the Isl1+/Tek+ and Isl1+/Tek− lineages. Scale bars, 100 μm (A–F).
Figure 6—figure supplement 1
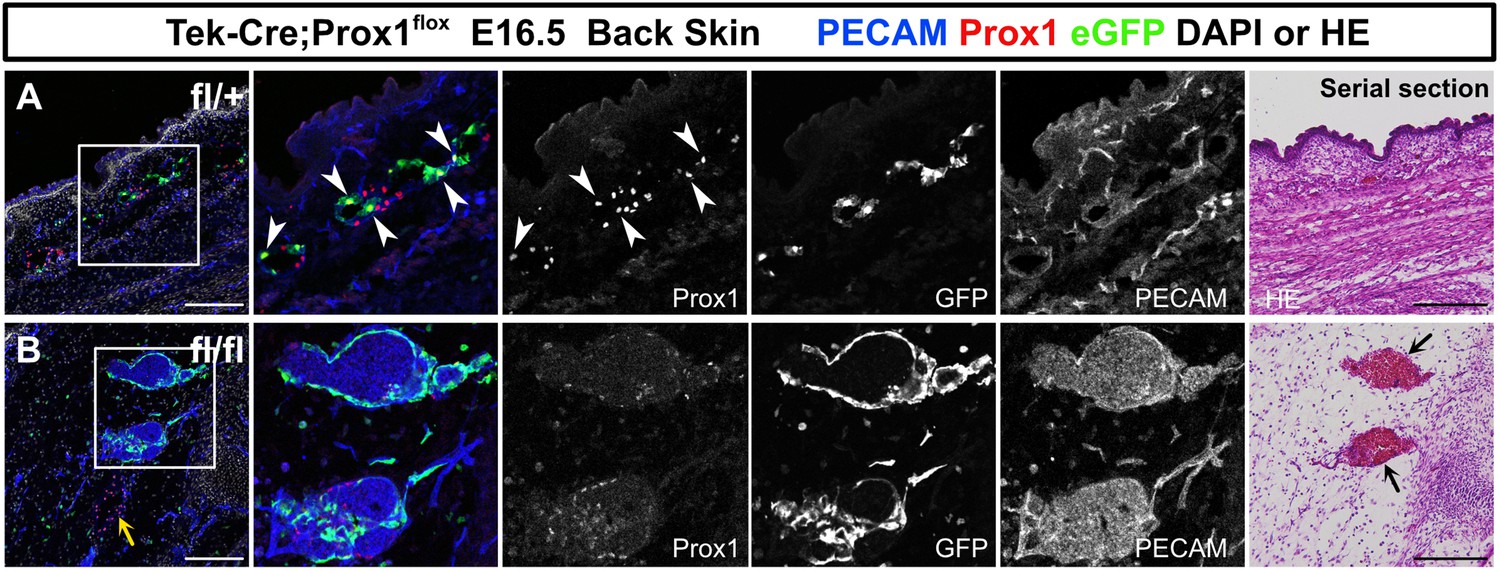
Prox1 expression level was decreased in Tek+ lymphatic endothelial cells (LECs) in Tek-Cre;Prox1fl/fl embryos.
(A, B) Sagittal sections of Tek-Cre;Prox1fl/+ or Tek-Cre;Prox1fl/fl mouse embryos (n=3, respectively), in which platelet endothelial cell adhesion molecule (PECAM), eGFP, Prox1, DAPI, or hematoxylin and eosin (HE) were labeled at embryonic day (E) 16.5. (A) eGFP+/Prox1+ LECs were observed in Tek-Cre;Prox1fl/+ embryos in the back skin (white arrowheads). (B) The Prox1 expression was decreased in eGFP+ cells in Tek-Cre;Prox1fl/fl embryos, whereas the expression was retained in eGFP- LECs (yellow arrow). Blood-filled lymphatic vessels were observed in Tek-Cre;Prox1fl/fl embryos (black arrows). Scale bars, 100 μm (A, B).
Figure 6—figure supplement 2
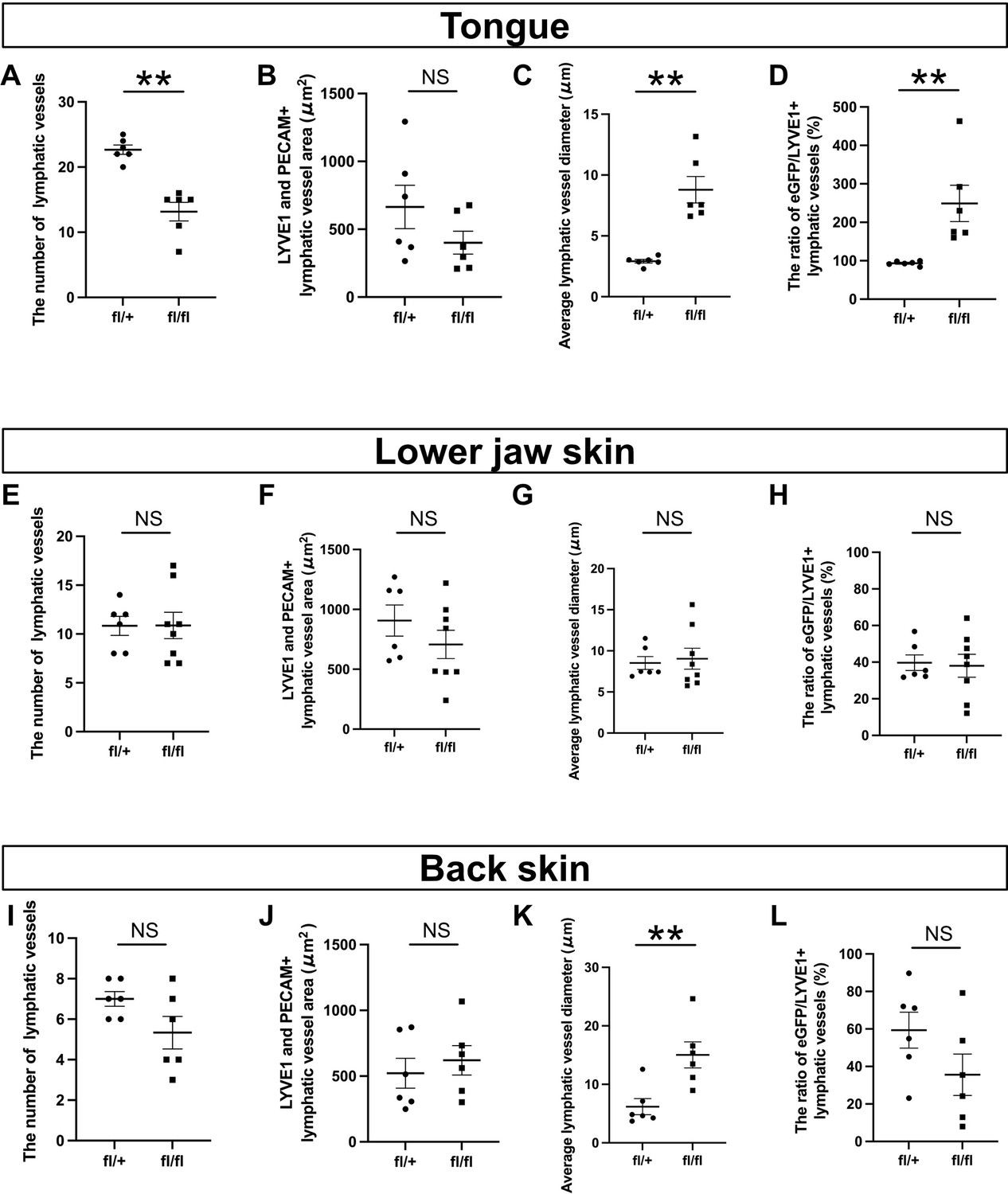
Lymphatic vessel phenotypic differences between Tek-Cre;Prox1fl/+ and Tek-Cre;Prox1fl/fl embryos.
(A–L) The results of quantitative analyses of lymphatic vessel phenotypes in the tongue (A–D), the skin of the lower jaw (E–H), and back skin (I–L) in Tek-Cre;Prox1fl/+ and Tek-Cre;Prox1fl/fl embryos at embryonic day (E) 16.5 are shown. **p=0.0022 (A, C, D) **p=0.0087 (K). All results are expressed as the mean ± SEM, and statistical analyses were performed using the non-parametric Mann-Whitney U-test. Each dot represents a value obtained from one sample. **p<0.01.
-
Figure 6—figure supplement 2—source data 1
Quantification of lymphatic vessels phenotypes in Tek-Cre;Prox1fl/+ and Tek- Cre; Prox1fl/fl embryos at embryonic day (E) 16.5.
- https://cdn.elifesciences.org/articles/81515/elife-81515-fig6-figsupp2-data1-v2.xlsx
Figure 7
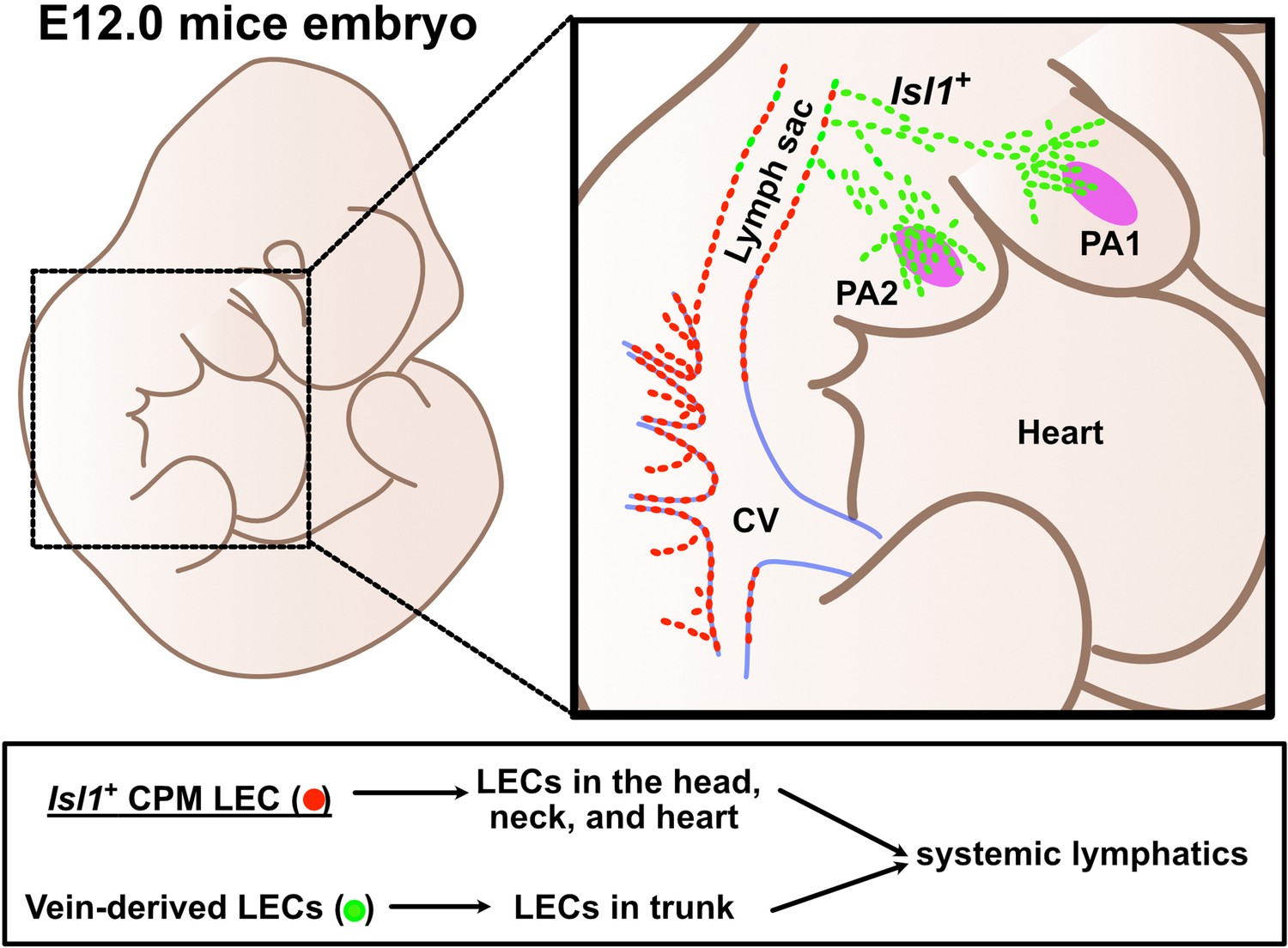
Lymphatic endothelial cells (LECs) are derived from two distinct origins.
Schematic representation of the origins of LECs. LECs are mainly derived from cardinal veins (red dots) and the cardiopharyngeal mesoderm (CPM) (green dots).
Additional files
-
Supplementary file 1
Primers used for genotyping.
- https://cdn.elifesciences.org/articles/81515/elife-81515-supp1-v2.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/81515/elife-81515-mdarchecklist1-v2.pdf
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The cardiopharyngeal mesoderm contributes to lymphatic vessel development in mouse
eLife 11:e81515.
https://doi.org/10.7554/eLife.81515




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}