Novel axonemal protein ZMYND12 interacts with TTC29 and DNAH1, and is required for male fertility and flagellum function
- University of Bordeaux, CNRS, France
- Bordeaux INP, Microbiologie Fondamentale et Pathogénicité, France
- CHU Grenoble-Alpes, UM de Génétique Chromosomique, France
- Institute for Advanced Biosciences, INSERM U1209, CNRS UMR 5309, Université Grenoble Alpes, Team Genetics Epigenetics and Therapies of Infertility, France
- Institut Cochin, INSERM U1016, CNRS UMR 8104, Université Paris Cite, France
- Université Bordeaux, Plateforme Protéome, France
- Department of Genetics, Reproductive Biomedicine Research Center, Royan Institute for Reproductive Biomedicine, ACECR, Islamic Republic of Iran
- Department of Andrology, Reproductive Biomedicine Research Center, Royan Institute for Reproductive Biomedicine, ACECR, Islamic Republic of Iran
- Polyclinique les Jasmins, Centre d'Aide Médicale à la Procréation, Centre Urbain Nord, Tunisia
- Human Sperm Bank, West China Second University Hospital of Sichuan University, China
- NHC Key Laboratory of Chronobiology, Sichuan University, China
- Key Laboratory of Birth Defects and Related Diseases of Women and Children (Sichuan University), Ministry of Education, China
- Obstetrics and Gynecology Hospital, Fudan University, China
- Université Grenoble Alpes, CNRS, France
- CHU Grenoble Alpes, Laboratoire de Génétique Moléculaire: Maladies Héréditaires et Oncologie, France
- CHU de Grenoble, UM GI-DPI, France
- Institute for Advanced Biosciences, INSERM U 1209, CNRS UMR 5309, Université Grenoble Alpes, Team Physiology and Pathophysiology of Sperm cells, France
Figures
Figure 1 with 3 supplements
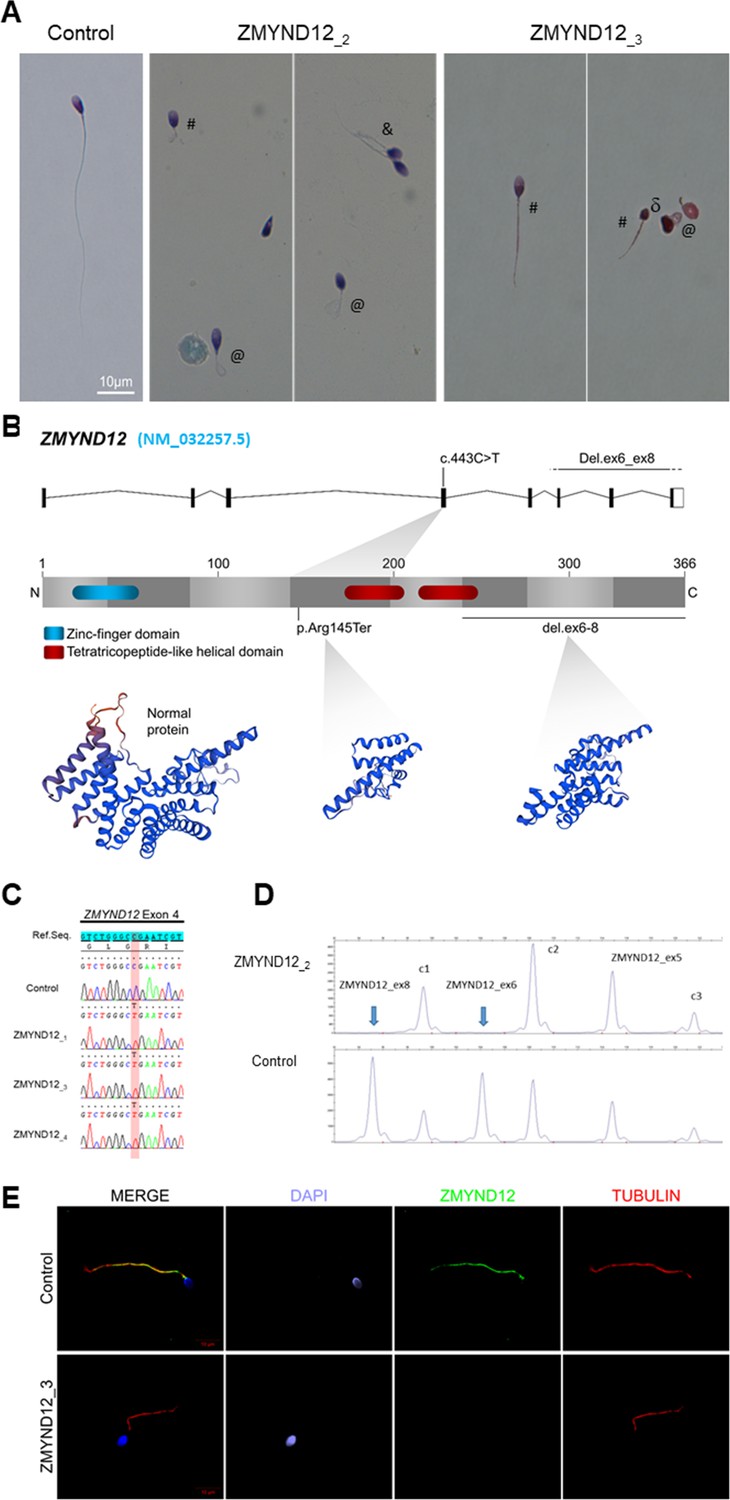
Morphology of normal and ZMYND12 mutant spermatozoa and variants identified in ZMYND12 individuals.
(A) Light microscopy analysis of spermatozoa from fertile control individuals and from individuals ZMYND12_2 and ZMYND12_3. Most spermatozoa from ZMYND12 individuals had short (#), coiled (@), or irregular caliber (&) flagella. Head malformations were also observed (δ). (B) Location of the variants identified within the ZMYND12 gene (NM_032257.5) and protein according to UniprotKB (Q9H0C1). The blue box represents a Zinc Finger domain and the orange boxes represent TPR domains (tetratricopeptide repeat). (C) Sanger sequencing electropherograms indicating the homozygous state of the c.433C>T variant located in exon 4 in the three individuals ZMYND12_1,3 and 4. The substituted nucleotide is highlighted in red. Variants are annotated in line with HGVS recommendations. (D) Confirmation of ZMYND12 exon deletion by MLPA. MLPA profiles obtained for control and ZMYND12_2 individuals. Each set contains three control probes for normalization purposes (c1–c3) and three ZMYND12-specific probes. Blue arrows indicate homozygous deletion of exons 6 and 8 in the ZMYND12 gene. ZMYND12 exon 5 is not included in the deletion. (E) Sperm cells from a fertile control individual and from individual ZMYND12_3 – carrier of the c.433C>T variant – were stained with anti-ZMYND12 (green) and anti-acetylated tubulin (red) antibodies. Sperm nuclear DNA was counterstained with DAPI (4′,6-diamidino-2-phenylindole) (blue). In control sperm, ZMYND12 immunostaining was present along the whole length of the flagellum; no staining was observed in sperm cells from individual ZMYND12_3. Scale bars: 10 μm.
Figure 1—figure supplement 1
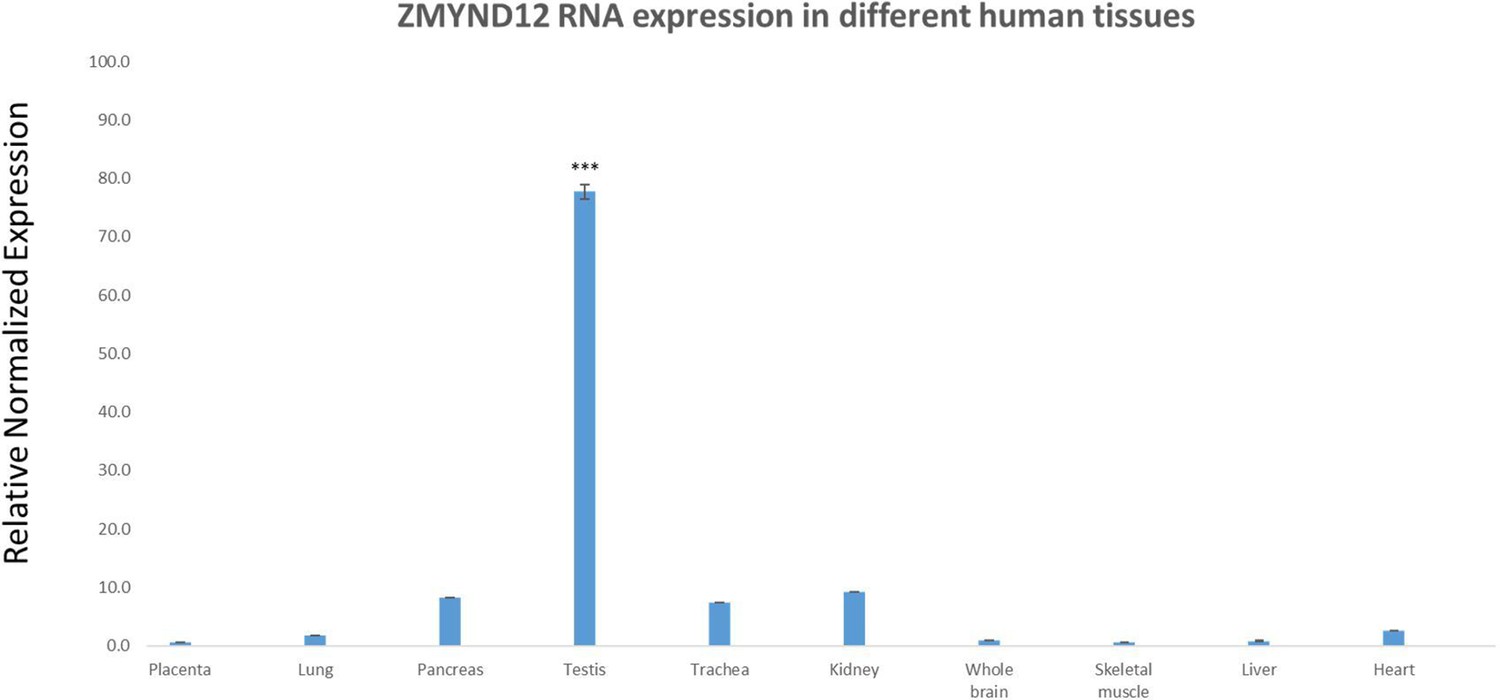
Relative mRNA expression of human ZMYND12 transcripts.
Quantitative real-time RT-PCR (RT-qPCR) was performed with cDNAs from various human tissues purchased from Life Technologies. A panel of 10 organs was used for experiments: placenta, lung, pancreas, testis, trachea, kidney, whole brain, skeletal muscle, liver, and heart. Each sample was assayed in triplicate for each gene on a StepOnePlus (Life Technologies) with Power SYBRGreen PCR Master Mix (Life Technologies). The PCR cycle was as follows: 10 min at 95°C, 1 cycle for enzyme activation; 15 s at 95C, 60 s at 60C with fluorescence acquisition, 40 cycles for the PCR. RT-qPCR data were normalized using the two reference housekeeping genes RPL6 and RPL27 for human samples, applying the −ΔΔCt method (Livak and Schmittgen, 2001). The 2−ΔΔCt value was set to 0 in brain cells, resulting in an arbitrary maximum expression of 1. The efficacy of primers was checked using a standard curve. Melting curve analysis was used to verify the presence of a single PCR product. Statistical significance of differences in expression of ZMYND12 transcripts in several organs was determined by applying a two-tailed t-test using Prism 4.0 software (GraphPad, San Diego, CA). ***p < 0.001.
Figure 1—figure supplement 2

Depth of coverage at the ZMYND12 locus for the ZMYND12_2 individual, carrier of a homozygous deletion of exons 6–8, and four other unrelated Multiple Morphological Abnormalities of the sperm Flagellum (MMAF) individuals.
Compared with the control individuals (blue), affected individual ZMYND12_2 (red) showed a complete absence of sequence coverage for exons 6–8. ZMYND12 is on the reverse genomic strand. The five samples shown were processed by exome-seq in a single batch, using the same Agilent Clinical Research capture kit, Illumina HiSeq 4000 sequencing technology, and bioinformatics pipeline (see Supplemental Methods).
Figure 1—figure supplement 3
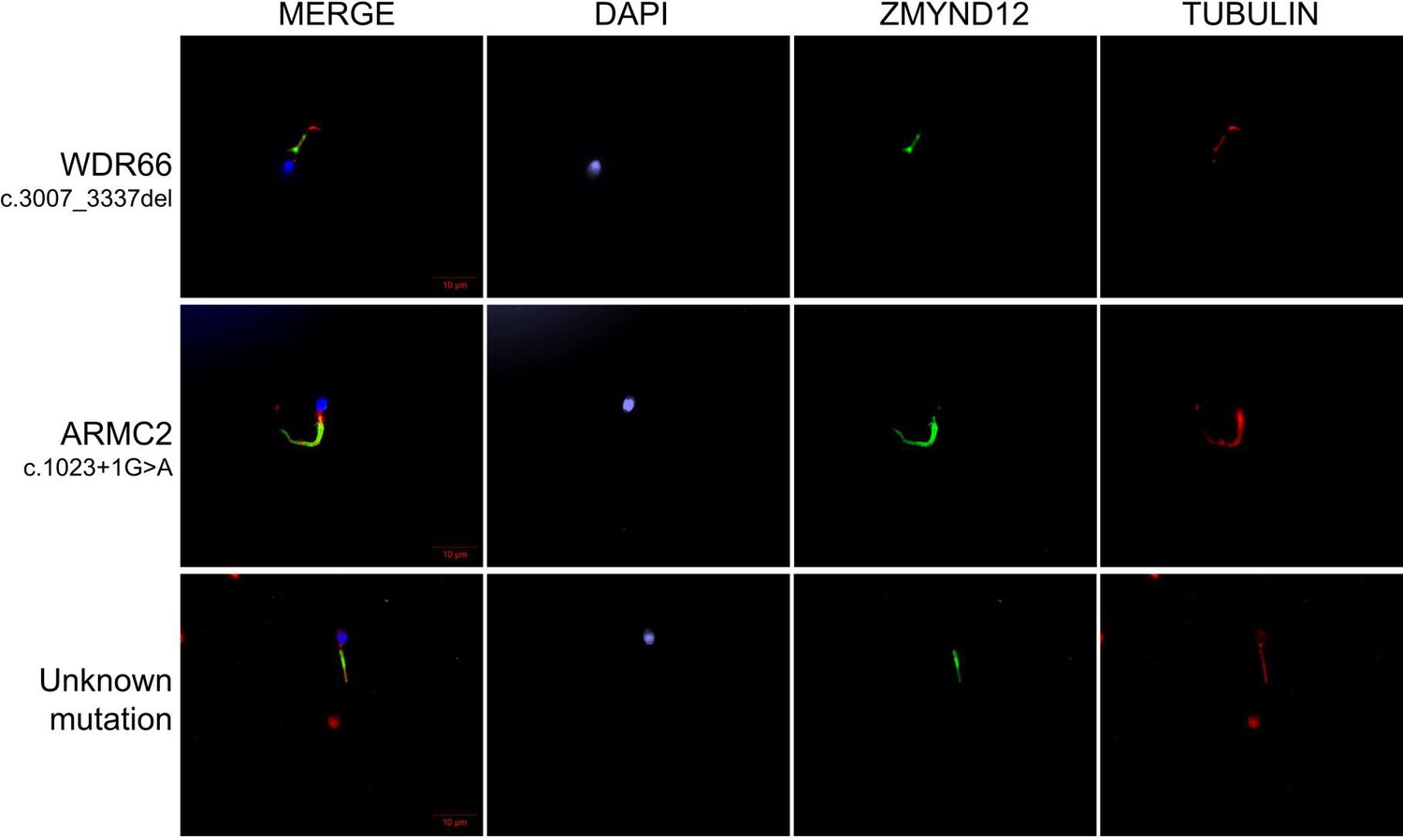
ZMYND12 immunostaining in human spermatozoa from Multiple Morphological Abnormalities of the sperm Flagellum (MMAF) individuals carriers of pathogenic variants in ARMC2 and WDR66 genes, or with unknwon genetic cause.
Sperm cells from a fertile control individual, two MMAF patients, carriers of mutations in ARMC2 and WDR66, and one MMAF patient with unknown genetic cause were stained with anti-ZMYND12 (green) and anti-acetylated tubulin (red) antibodies. Sperm nuclear DNA was counterstained with DAPI (blue). ZMYND12 immunostaining was present throughout the flagellum in all samples, regardless of phenotype. Variants are specified for each patient. Scale bars: 10 μm.
Figure 2 with 4 supplements
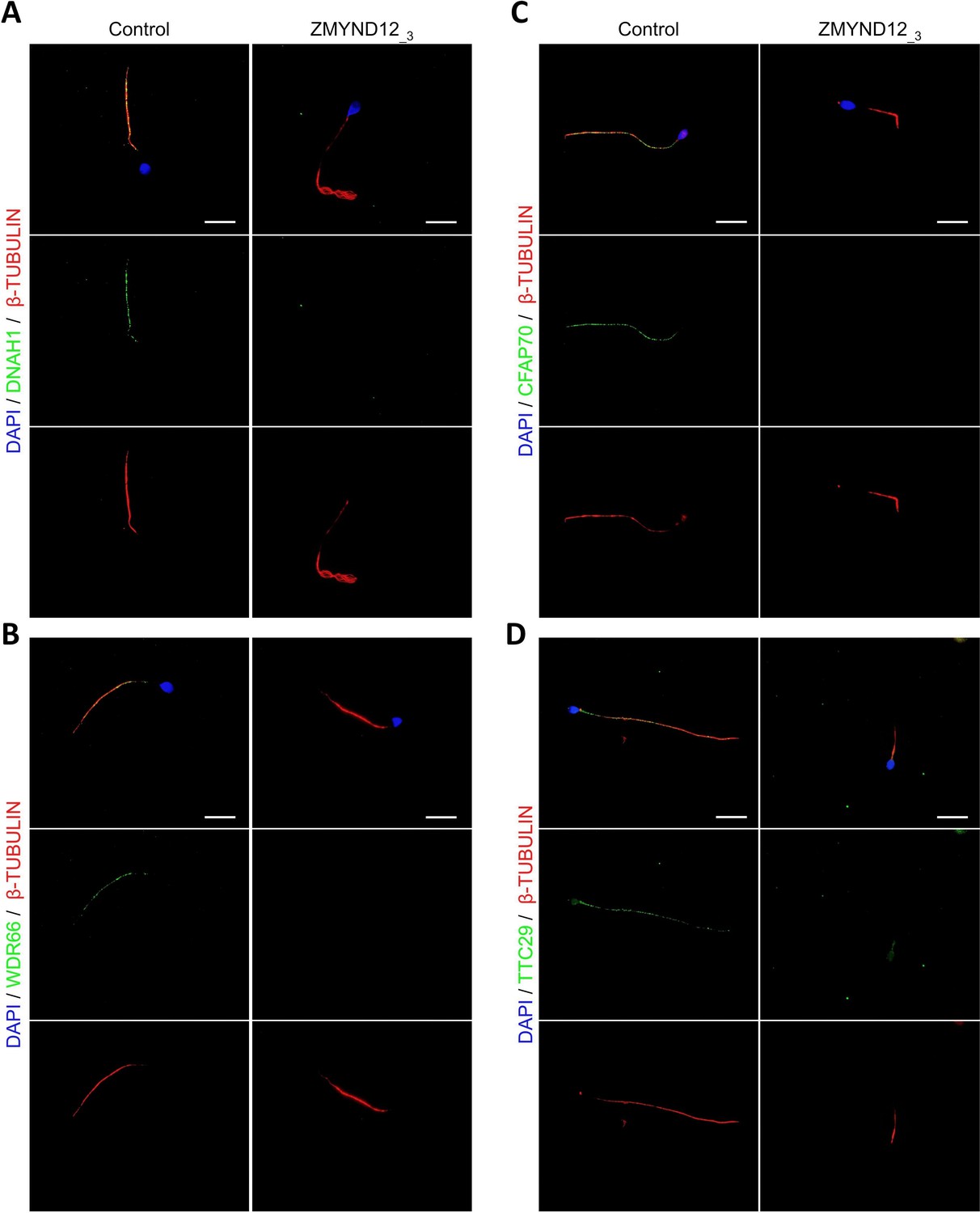
Altered immunostaining for DNAH1, WDR66, CFAP70, and TTC29 in the presence of ZMYND12 variants.
Immunofluorescence experiments were performed using sperm cells from control individuals and from individual ZMYND12_3 carrying the nonsense variant c.433C>T. Although tubulin staining remained detectable, sperm cells from individual ZMYND12_3 showed no immunostaining for (A) DNAH1, (B) WDR66, (C) CFAP70, and (D) TTC29. Scale bars: 10 µm.
Figure 2—figure supplement 1
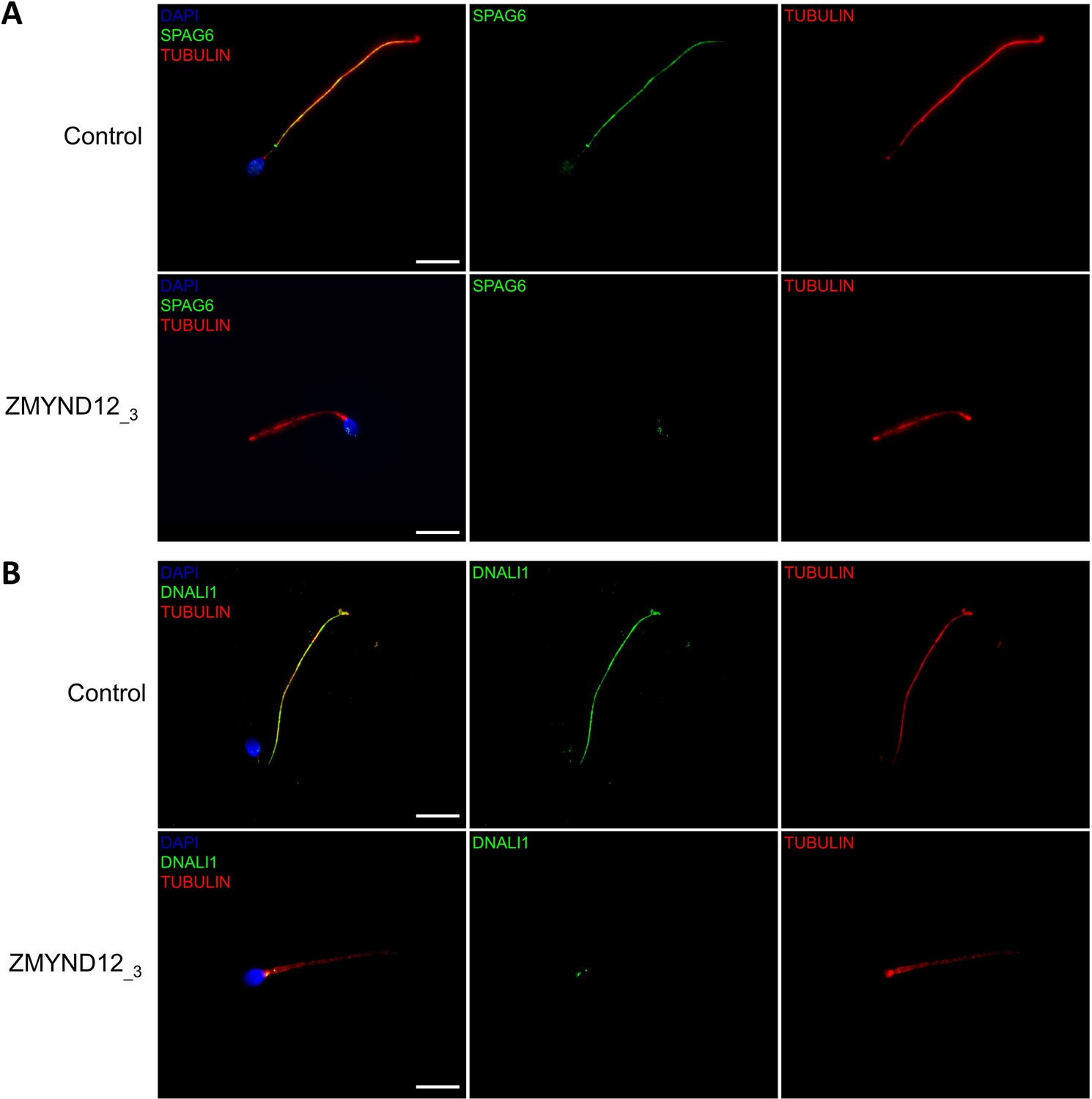
Altered immunostaining for SPAG6 and DNALI1 in the presence of ZMYND12 variants.
Immunofluorescence experiments were performed using sperm cells from control individuals and from individual ZMYND12_3 carrying the nonsense variant c.433C>T. Although tubulin staining remained detectable, sperm cells from individual ZMYND12_3 showed no immunostaining for (A) SPAG6 and (B) DNALI1. The central pair complex (CPC) and the IDAs were therefore strongly disorganized in these cells. Scale bars: 10 µm.
Figure 2—figure supplement 2
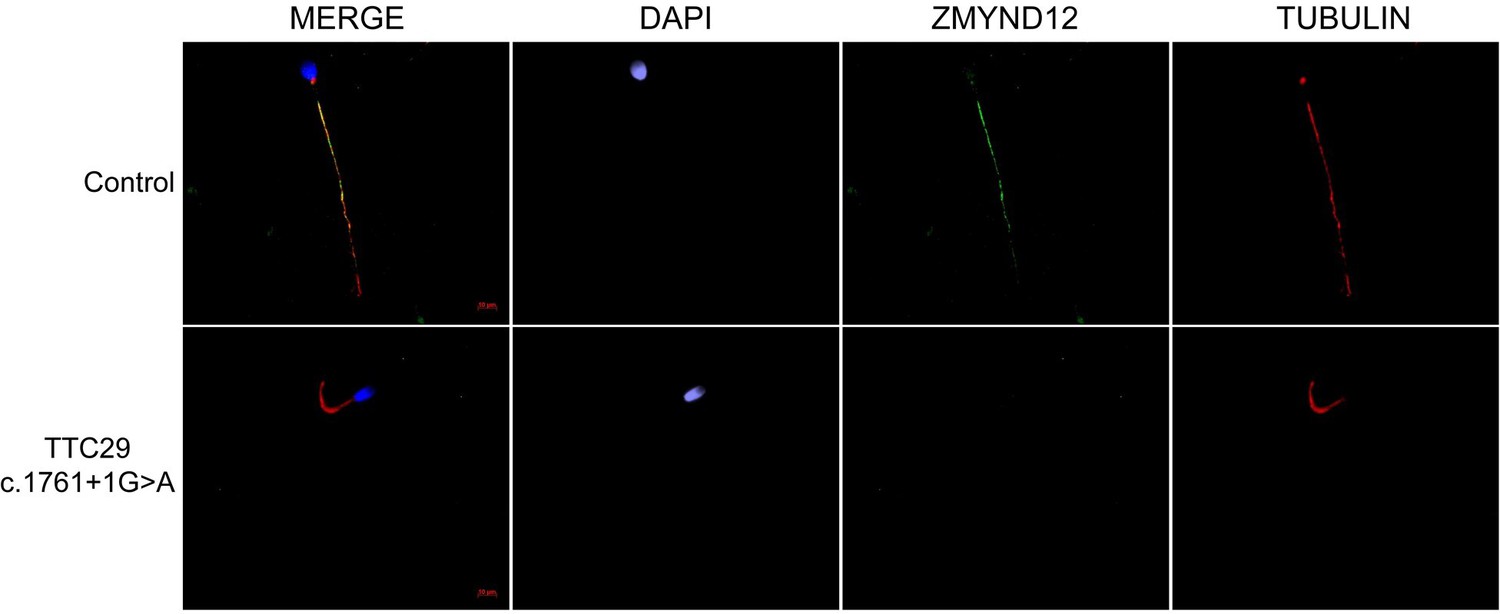
ZMYND12 is absent from sperm cells from TTC29 individuals.
Sperm cells from a fertile control individual and a TTC29 individual carrying the previously reported c.1761+1G>A splice-variant (Lorès et al., 2019) were stained with anti-ZMYND12 (green) and anti-acetylated tubulin (red) antibodies. DAPI counterstaining was used to reveal sperm nuclear DNA (blue). In control sperm, ZMYND12 immunostaining decorated the full length of the flagellum, but in sperm from TTC29 individuals, a total absence of ZMYND12 was recorded. Scale bars: 10 μm.
Figure 2—figure supplement 3
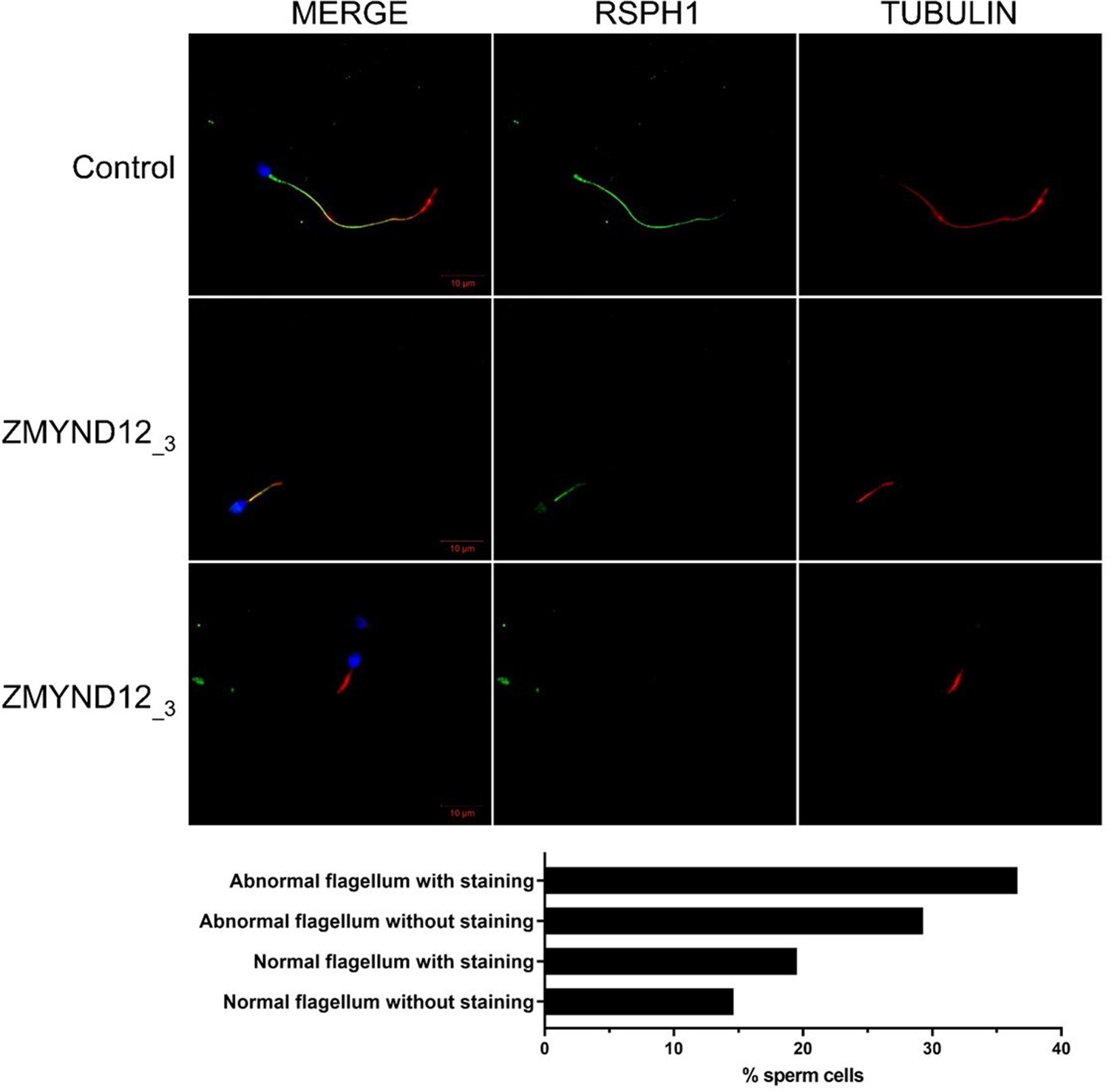
Radial spokes are inconstantly affected in ZMYND12 individuals.
Sperm cells from a fertile control individual and from individual ZMYND12_3 stained with anti-RSPH1 (green) – located in the radial spokes – and anti-acetylated tubulin (red) antibodies. RSPH1 immunostaining is uniformly present along the length of the flagellum in control sperm cells. Sperm cells from individual ZMYND12_3 show heterogeneous staining regardless of flagellar morphology, suggesting that the radial spokes are inconsistently present in sperm cells from this individual. Scale bars: 10 µm. Lower panel: Histogram showing the percentage of analyzed spermatozoa of various morphologies displaying or lacking RSPH1 immunostaining (number of sperm cells analyzed = 200).
Figure 2—figure supplement 4

Immunostaining for AKAP4, DNAH2, DNAH8, DNAH17, DNAI1, and GAS8 is not affected by ZMYND12 mutation.
Immunofluorescence (IF) experiments were performed using sperm cells from control individuals and from individual ZMYND12_3, carrier of the nonsense variant c.433C>T. Immunostaining patterns for AKAP4, DNAH2, DNAH8, DNAH17, DNAI1, and GAS8 in sperm from individual ZMYND12_3 were comparable to those observed in control sperm cells, demonstrating that the fibrous sheath, outer dynein arms (ODAs) and the N-DRC were not directly affected by ZMYND12 variants. Scale bars: 10 µm.
Figure 3 with 2 supplements
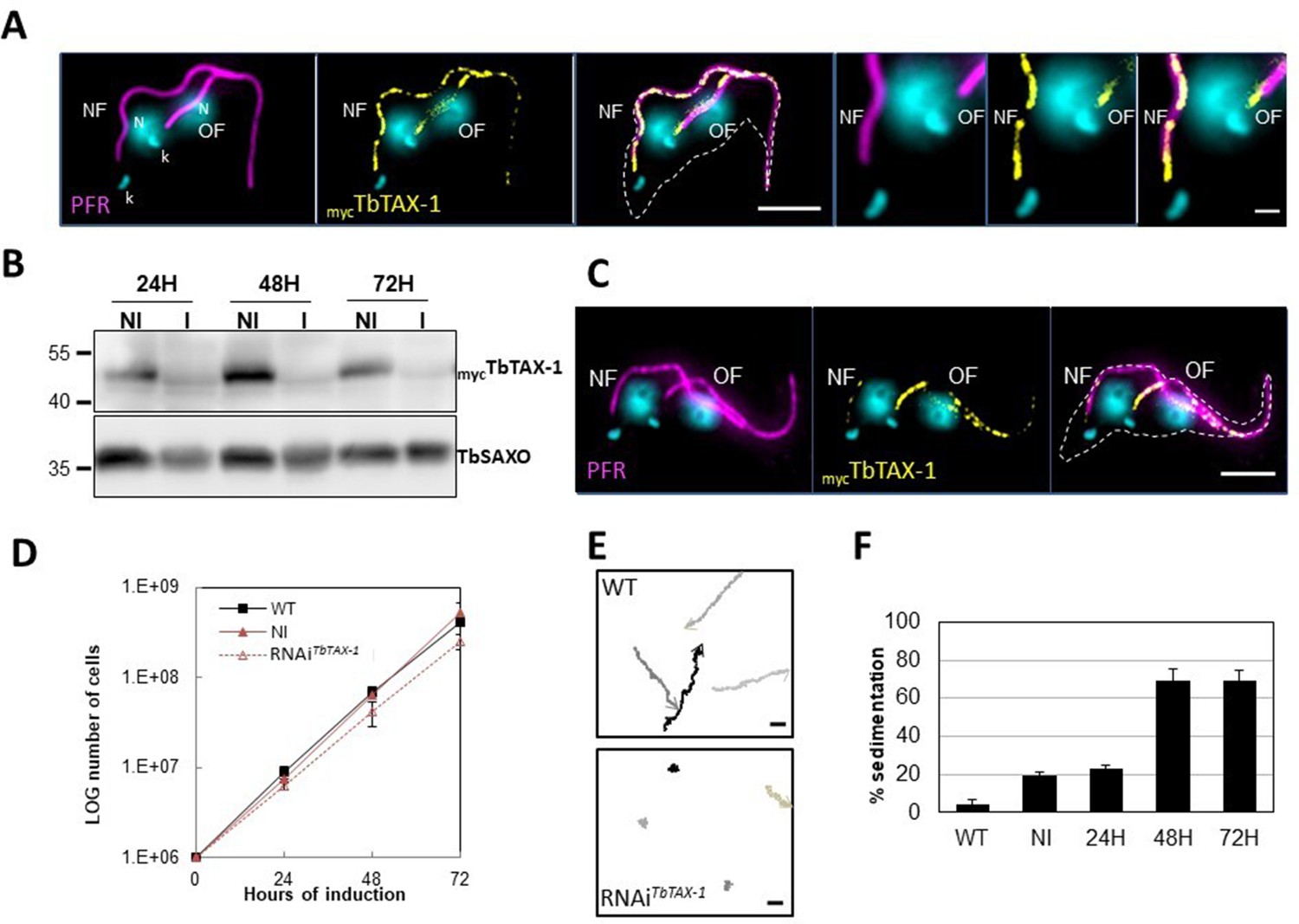
TbTAX-1 is an axoneme-associated protein; its knock-down leads to flagellar motility defects.
(A) Representative image of a post-mitotic detergent-extracted cell immunolabelled with anti-PFR2 (magenta) staining the paraflagellar rod (a para-axonemal structure) and anti-myc (yellow) to reveal mycTbTAX-1. The protein was detected on the axoneme in both the old flagellum (OF) and the new flagellum (NF). Immunolabeling indicated that the axonemal localization of mycTbTAX-1 extended distally along the flagellum beyond the region labelled by PFR2 (see zoom). The mitochondrial genome (kinetoplast, k) and nuclei (N) were stained with DAPI. The outline of the cell body is indicated by the dashed white line. Scale bars: 5 μm in whole-cell images; 1 μm in zoomed images. (B) Anti-myc western blot analysis of RNAiTbTAX-1 knock-down 24-, 48-, or 72-hr post-induction. mycTbTAX-1 protein levels were reduced in induced cells (I) compared to non-induced cells (NI); TbSAXO, a flagellum-specific protein, was used as loading control. (C) Representative image of a detergent-extracted post-mitotic cell after RNAiTbTAX-1 induction, showing a strong decrease in mycTbTAX-1 labeling in the NF compared to the OF. (D) Comparative T. brucei growth curves for wild-type (WT), non-induced (NI), and tetracycline-induced RNAiTbTAX-1 cell lines at 0-, 24-, 48-, and 72-hr post-induction. (E) Mobility tracking from video microscopy recordings of live cells: WT and RNAiTbTAX-1 after induction for 72 hr. The positions of individual cells are plotted at 0.28 s intervals; circles indicate start positions, arrowheads indicate end positions. A dramatic loss of progressive mobility was observed after TbTAX1 protein knock-down. Scale bars: 20 μm. (F) Sedimentation assays for T. brucei WT, non-induced (NI), and tetracycline-induced RNAiTbTAX-1 at 24-, 48-, and 72-hr post-induction. The percentage of cells sedimenting increased from 20 to 70% after depletion of the TbTAX-1 protein.
-
Figure 3—source data 1
Annotated and uncropped western blots and raw images for Figure 3B.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig3-data1-v1.zip
Figure 3—figure supplement 1

TbTAX-1 is the T. brucei ortholog of ZMYND12.
(A) Clustal Omega alignment of human ZYMND12 and TbTAX-1. (B) Schematic representation of wild-type and truncated forms of ZYMND12 and TbTAX-1.
Figure 3—figure supplement 2

TbTAX-1 RNAi knock-down.
Micrograph of wild-type (WT) cells and TbTAX-1 RNAi-induced cells (48 hr) in thin section. The characteristic 9+2 structure of the axoneme and the paraflagellar structure (PFR) are visible, and no obvious structural defects are observed in RNAi-induced cells.
Figure 4 with 1 supplement
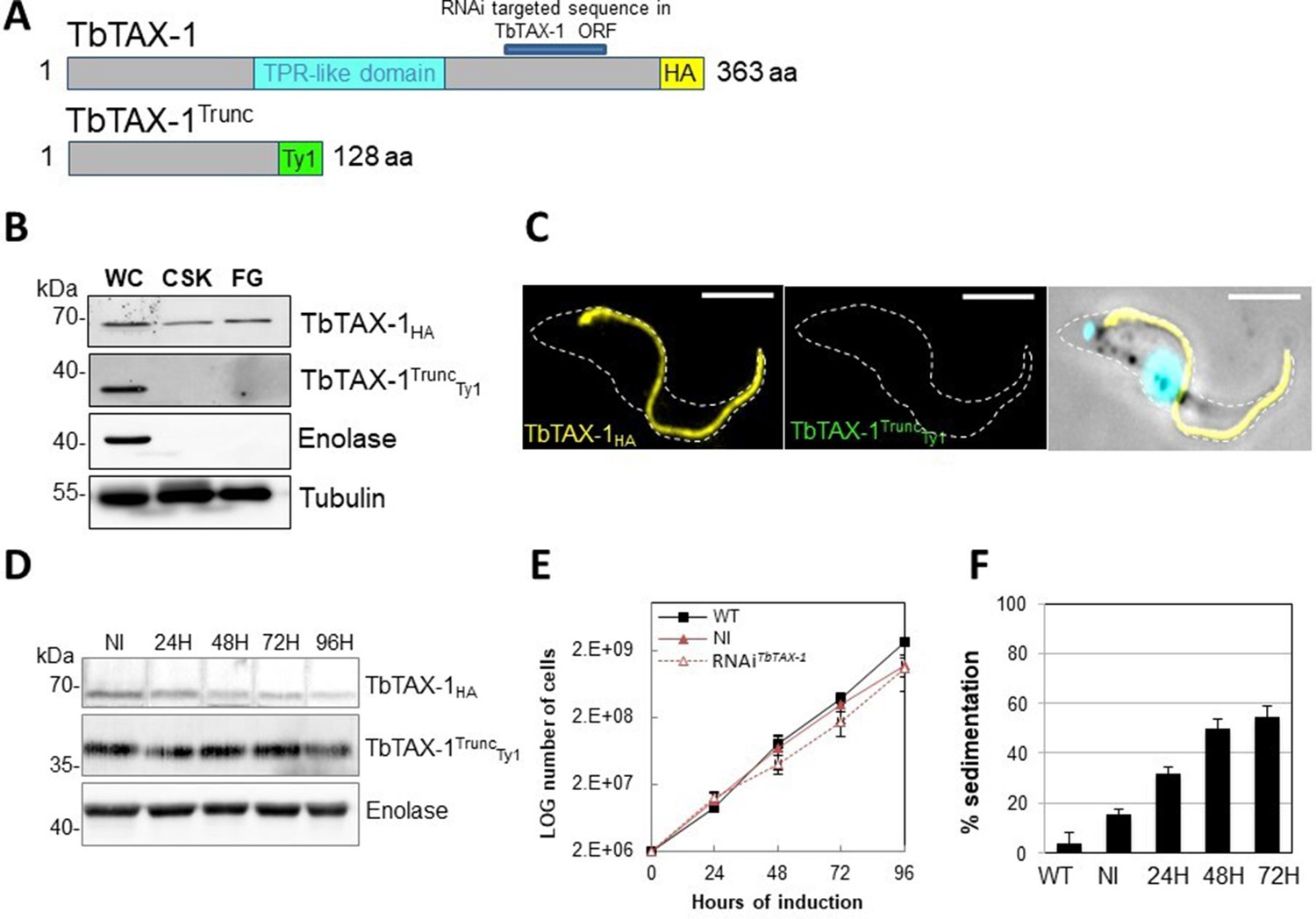
The truncated form of TbTAX-1 is not functional.
(A) Schematic representation of full-length TbTAX-1 and its truncated form. The TPR-like domain is illustrated in light blue, and the targeted RNA interference sequence in dark blue. (B) Western blot analysis of subcellular fractions to determine localization of TbTAX-1HA and its truncated form TbTAX-1TruncTy1. Enolase (cytoplasm) and tubulin (cytoskeleton) were used as loading controls. (C) Immunofluorescence labeling of detergent-extracted cytoskeletons to detect TbTAX-1HA (yellow) and TbTAX-1TruncTy1 (stained green, but not visible as the protein was eliminated during detergent extraction). Scale bars: 5 μm. (D) Western blot analysis of the impact of RNAiTbTAX-1 expression on TbTAX-1TruncTy1 levels. Enolase was used as a loading control. (E) Growth curves of wild-type (WT), non-induced (NI), and RNAiTbTAX-1-induced cells expressing TbTAX-1HA and TbTAX-1TruncTy1. (F) Sedimentation assays for T. brucei WT, non-induced (NI), and RNAiTbTAX-1-induced cells at 24, 48, and 72 hr.
-
Figure 4—source data 1
Annotated and uncropped western blots and raw images for Figure 4B.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig4-data1-v1.zip
-
Figure 4—source data 2
Annotated and uncropped western blots and raw images for Figure 4D.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig4-data2-v1.zip
Figure 4—figure supplement 1
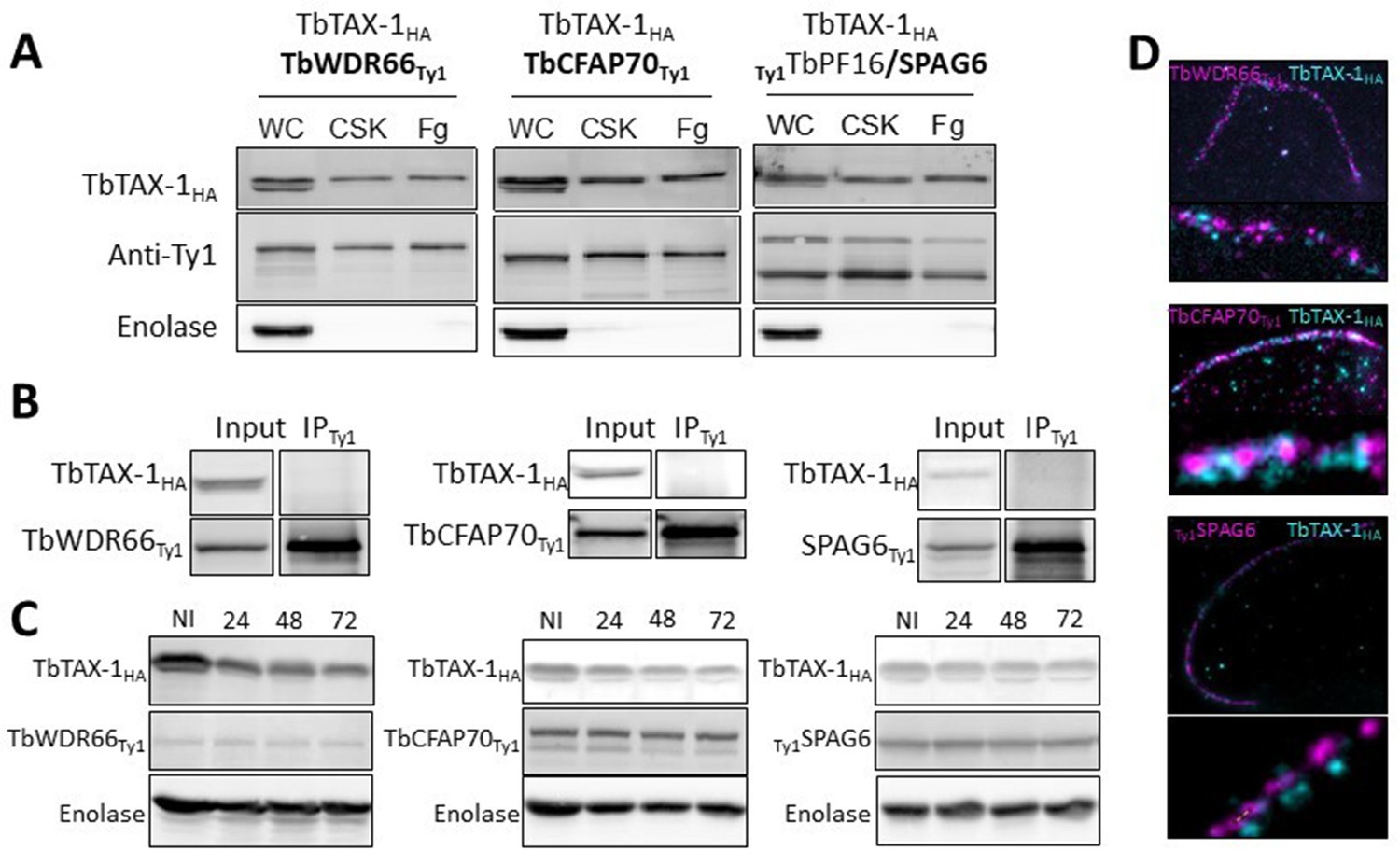
TbTAX-1 knock-down does not affect TbWDR66, TbCFAP70, or TbSPAG6.
(A) TbWDR66, TbCFAP70, and TbSPAG6 are axoneme-associated proteins according to western blot analysis of whole-cell (WC) lysate, detergent-extracted cytoskeletons (CSK), salt-extracted flagella (Fg). Enolase was used as extraction control for cytoskeleton and flagella samples. (B) Co-immunoprecipitation assays using anti-Ty1 (IPTy1) in cell lines expressing TbTAX-1HA alongside TbWDR66Ty1 or TbCFAP70 or TbSPAG6Ty1. Proteins were detected simultaneously by western blotting using anti-Ty1 and anti-HA antibodies and their corresponding fluorescent secondary antibodies. (C) Western blot analysis of the fate of TbWDR66Ty1, TbCFAP70Ty1, or TbPF16/SPAG6Ty1 in TbTAX-1 RNAi cells; non-induced (NI) cells, and cells induced for 24, 48, or 72 hr. Enolase served as loading control. (D) Ultrastructure expansion microscopy (U-ExM) co-immunolabeling of TbTAX-1HA (cyan) and TbWDR66Ty1, TbCFAP70Ty1, or TbSPAG6Ty1 (magenta).
-
Figure 4—figure supplement 1—source data 1
Annotated and uncropped western blots and raw images for Figure 4—figure supplement 1A.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig4-figsupp1-data1-v1.zip
-
Figure 4—figure supplement 1—source data 2
Annotated and uncropped western blots and raw images for Figure 4—figure supplement 1B.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig4-figsupp1-data2-v1.zip
-
Figure 4—figure supplement 1—source data 3
Annotated and uncropped western blots and raw images for Figure 4—figure supplement 1C.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig4-figsupp1-data3-v1.zip
Figure 5
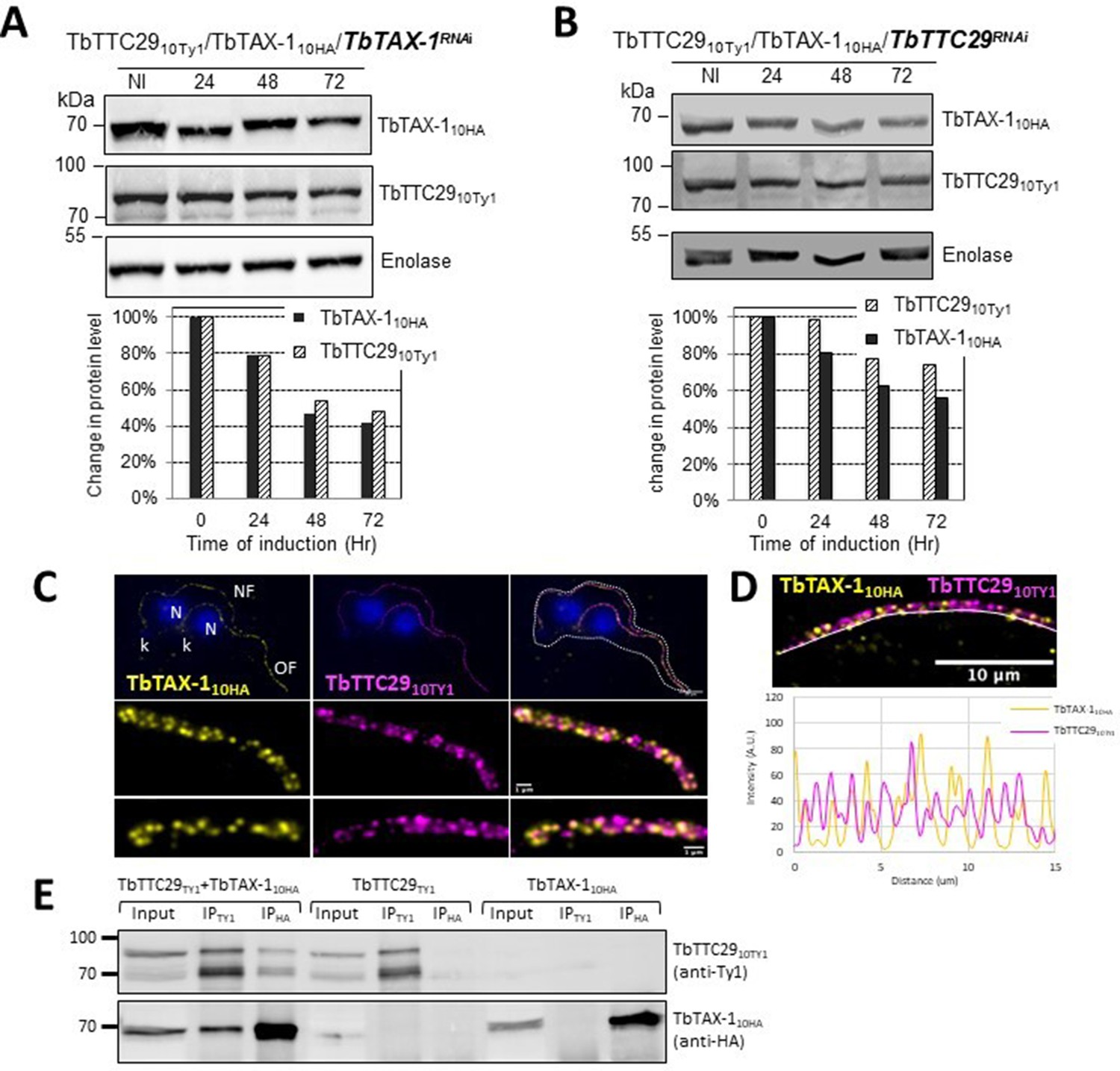
TbTAX-1 and TbTTC29 are part of the same protein complex.
(A) Analysis of the impact of TbTAX-1 RNAi on TbTTC29Ty1 protein levels. Western blot (upper panel) and quantification (lower panel). (B) Analysis of the impact of TbTTC29 RNAi induction on TbTAX-1HA protein levels. Western blot (upper panel) and quantification (lower panel). (C) Ultrastructure expansion microscopy (U-ExM) analysis of TbTAX-1HA (yellow) and TbTTC29TY1 (purple) colocalization in a dividing cell with an old flagellum (OF) and a new flagellum (NF). Kinetoplast (k) and nuclei (N) were counterstained with Hoechst (blue). (D) Confocal image of U-ExM labeling of TbTAX-1HA (yellow) and TbTTC29TY1 (magenta) (upper panel). Lower panel: profile plot along the white line showing overlapping intensity peaks for TbTAX-1 and TbTTC29, confirming true colocalization. (E) Anti-Ty1 (IPTy1) or anti-HA (IPHA) co-immunoprecipitation assays in cell lines expressing both TbTAX-1HA and TbTTC29Ty1, or TbTTC29Ty1 alone or TbTAX-1HA alone to verify IP specificity. Proteins were detected simultaneously by western blotting using anti-Ty1 and anti-HA antibodies and fluorescent anti-mouse (detecting mouse anti-Ty1) and anti-rabbit (detecting rabbit anti-HA) secondary antibodies. IP assays on cell lines expressing only one or the other tagged protein did not pull down the other tagged protein.
-
Figure 5—source data 1
Annotated and uncropped western blots and raw images for Figure 5A.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig5-data1-v1.zip
-
Figure 5—source data 2
Annotated and uncropped western blots and raw images for Figure 5B.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig5-data2-v1.zip
-
Figure 5—source data 3
Annotated and uncropped western blots and raw images for Figure 5E.
- https://cdn.elifesciences.org/articles/87698/elife-87698-fig5-data3-v1.zip
Videos
Video 1
Video microscopy of wild-type (WT) cells.
Video 2
Video microscopy of RNAiTbTAX-1 cells 7 days post-induction.
Tables
Table 1
Detailed semen parameters in the four Multiple Morphological Abnormalities of the sperm Flagellum (MMAF) individuals harboring a ZMYND12 variant.
| ZMYND12 mutated individuals | Semen parameters | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Individuals | Homozygous ZMYND12 variants | Sperm volume (ml) | Sperm concentration (106/ml) | Total motility 1 hr | Vitality | Normal spermatozoa | Absent flagella | Short flagella | Coiled flagella | Bent flagella | Flagella of irregular caliber | Tapered head | Thin head | Micro-cephalic | Macro-cephalic | Multiple heads | Abnormal base | Abnormal acrosomal region |
| ZMYND12_1 | c.433C>T | 2 | 6.5 | 4 | 70 | 0 | 35 | 48 | 10 | 0 | 34 | 22 | 2 | 2 | 4 | 6 | 20 | 80 |
| ZMYND12_2 | Del ex6-8 | 2.5 | 4.25 | 2 | NA | 0 | NA | 72 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| ZMYND12_3 | c.433C>T | 3 | 34 | 2 | 50 | 0 | 20 | 20 | 28 | 0 | 46 | 22 | 6 | 8 | 2 | 2 | 24 | 88 |
| ZMYND12_4 | c.433C>T | 2.6 | 0.1 | 0 | NA | NA | NA | 70 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Reference limits* | 1.5 (1.4–1.7) | 15 (12–16) | 40 (38–42) | 58 (55–63) | 23 (20–26) | 5 (4–6) | 1 (0–2) | 17 (15–19) | 13 (11–15) | 2 (1–3) | 3 (2–4) | 14 (12–16) | 7 (5–9) | 1 (0–2) | 2 (1–3) | 42 (39–45) | 60 (57–63) | |
-
Values are percentages unless specified otherwise. NA: not available.
-
*
Reference limits (5th centiles and their 95% confidence intervals) according to World Health Organization (WHO) standards (Cooper et al., 2010) and the distribution range of morphologically normal spermatozoa observed in 926 fertile individuals (Auger et al., 2016).
Additional files
-
Supplementary file 1
Proteins co-immunoprecipitated with TbTAX-1 and TbTTC29 identified by mass spectrometry analyses.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp1-v1.docx
-
Supplementary file 2
Proteins identification by tandem mass spectrometry (MS/MS) analysis of proteins co-immunoprecipitated with TTC29 from wild-type and Ttc29−/− L5 and L7 mouse testes.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp2-v1.xlsx
-
Supplementary file 3
Primer sequences for Sanger sequencing verification of ZMYND12 variants.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp3-v1.docx
-
Supplementary file 4
ZMYND12 MLPA probes used in this work.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp4-v1.docx
-
Supplementary file 5
Primers used for quantitative real-time RT-PCR (RT-qPCR) detection of ZMYND12 in human tissue extracts.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp5-v1.docx
-
Supplementary file 6
Primary antibodies used in immunofluorescence experiments with human samples.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp6-v1.docx
-
Supplementary file 7
Primary antibodies used in immunofluorescence experiments with Trypanosoma cells.
- https://cdn.elifesciences.org/articles/87698/elife-87698-supp7-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/87698/elife-87698-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Novel axonemal protein ZMYND12 interacts with TTC29 and DNAH1, and is required for male fertility and flagellum function
eLife 12:RP87698.
https://doi.org/10.7554/eLife.87698.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}