A system of feed-forward cerebellar circuits that extend and diversify sensory signaling
- Interdisciplinary Graduate Program in Neuroscience, Arizona State University, United States
- School of Life Sciences, Arizona State University, United States
- Barrett Honors College, Arizona State University, United States
Figures
Figure 1
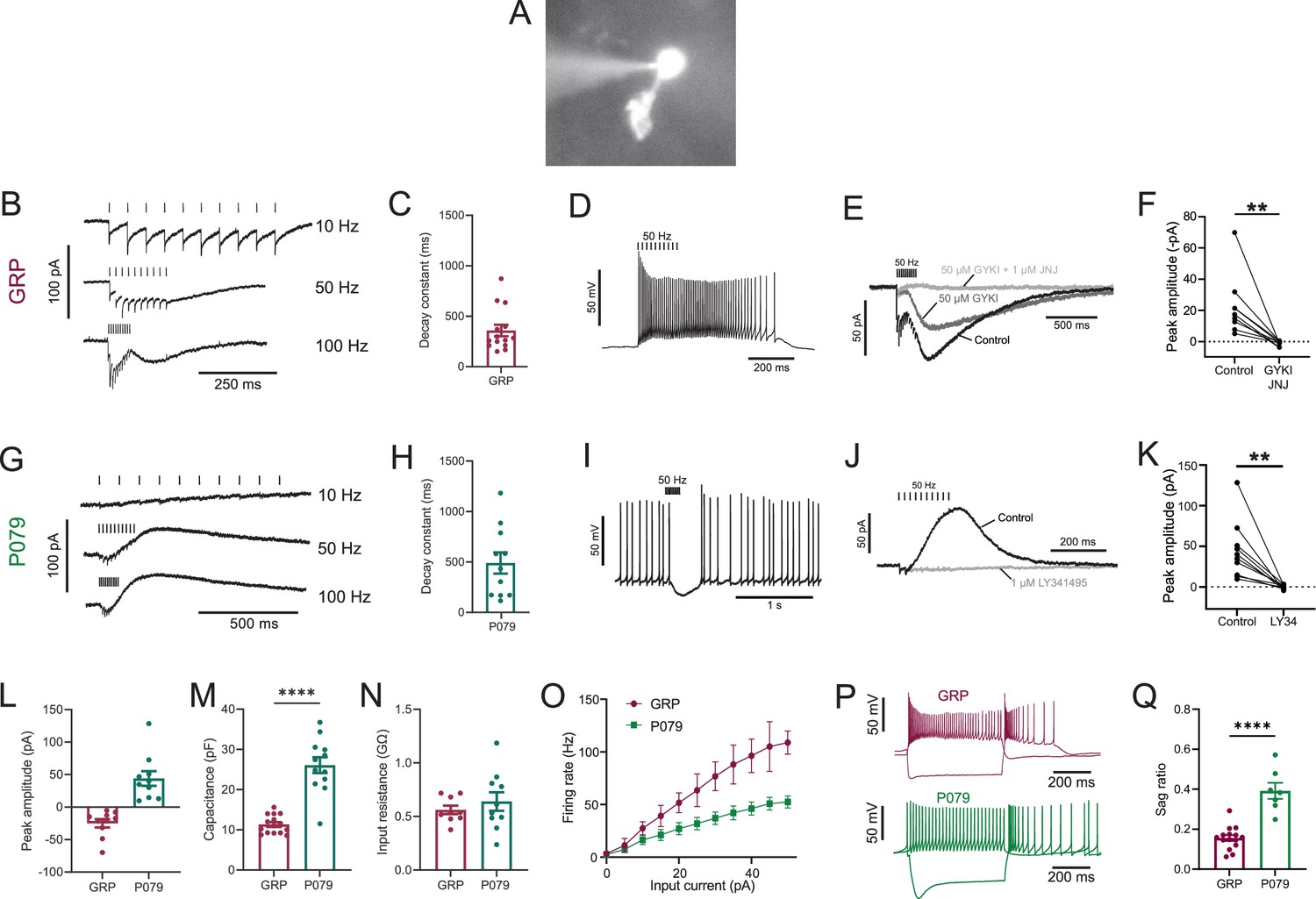
Physiological characterization of GRP and P079 unipolar brush cells (UBCs) in acute brain slices.
(A) Representative image of UBC in whole-cell patch-clamp recording filled with Alexa Fluor 488. (B) In GRP UBCs, mossy fiber stimulation evoked fast excitatory postsynaptic currents (EPSCs) and a slow inward current that became more prominent with faster stimulation rates. (C) Summary of the decay constants of the slow EPSCs generated by a train of 10 stimuli at a 50 Hz rate in GRP UBCs. (D) In current clamp, 50 Hz stimulation produced a burst of spikes that outlasted the stimulus. (E) The fast EPSCs and part of the slow EPSC were blocked by an AMPA receptor antagonist (50 μM GYKI53655). The remaining slow EPSC was blocked by an mGluR1 antagonist (1 μM JNJ16259685). (F) Inward currents in GRP UBCs were consistently blocked by AMPA receptor and mGluR1 receptor antagonists. (G) In P079 UBCs, mossy fiber stimulation evoked slow outward currents. (H) Summary of the decay constants of the slow outward currents generated by a train of 10 stimuli at a 50 Hz rate in P079 UBCs. (I) In current clamp, 50 Hz stimulation generated a pause in spontaneous action potential firing in P079 UBCs. (J) The outward current was blocked by an mGluR2 antagonist (1 μM LY341495). (K) Outward currents in P079 UBCs were entirely blocked by 1 μM LY341495. (L) Peak amplitudes of the slow currents were inward in all recorded GRP UBCs and outward in all recorded P079 UBCs. (M) The capacitance was significantly higher in P079 UBCs compared to GRP UBCs. (N) The input resistance was not different between GRP UBCs and P079 UBCs. (O) Frequency-intensity curves show that GRP UBCs are more excitable and are able to fire at faster rates than P079 UBCs. (P) Example traces showing the response of GRP and P079 UBCs to 40 pA depolarizing and –100 pA hyperpolarizing 500 ms current steps. The GRP UBCs fire at a higher rate than P079 UBCs during a 40 pA depolarizing current step. The P079 UBCs have a more prominent voltage sag in response to the –100 pA hyperpolarizing current step than the GRP UBCs. (Q) The sag ratio (peak-steady state/peak) was larger in P079 UBCs than GRP UBCs. Stimulation artifacts have been removed for clarity. Error bars are SEM.
Figure 2
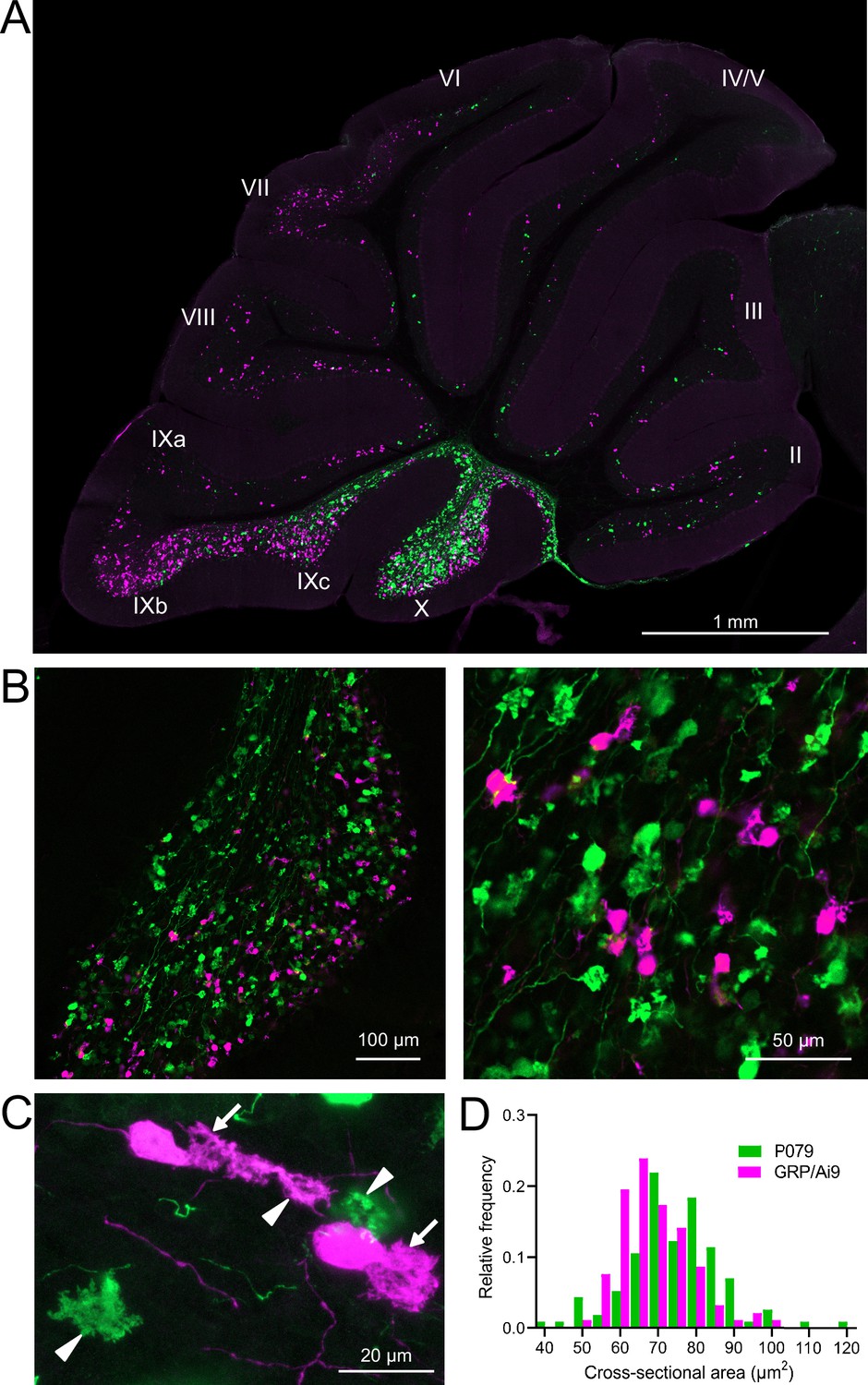
GRP/Ai9/P079 mouse line labels two distinct subsets of unipolar brush cells (UBCs).
(A) Sagittal section of cerebellar vermis. Fluorescence is restricted to UBC cell types. P079 UBCs (mCitrine; green) are present in high density in lobe X and lobe IXc, while GRP UBCs (tdTomato, magenta) are present in those lobes as well as lobe IXb and in a lower density in lobe VI–VIII. Maximum intensity projection. (B) Left: sagittal section showing expression of mCitrine labeling P079 UBCs and tdTomato labeling GRP UBCs in lobe X. More GRP UBCs are present in the ventral leaflet than the dorsal leaflet. Right- UBCs and their axons and terminals are well labeled. Only rarely were UBCs labeled with both colors. Maximum intensity projection. (C) Magnified view of UBCs with their dendritic brushes indicated with arrows. Two P079 terminals and one GRP terminal are indicated with arrowheads. The GRP UBC at the top of the image demonstrates the challenge of differentiating the dendritic brush of a UBC from a terminal that is labeled with the same fluorophore. Maximum intensity projection. (D) In lobe X, P079 UBCs had larger somas than GRP UBCs, although there is a somewhat bimodal distribution in the P079 population.
Figure 3
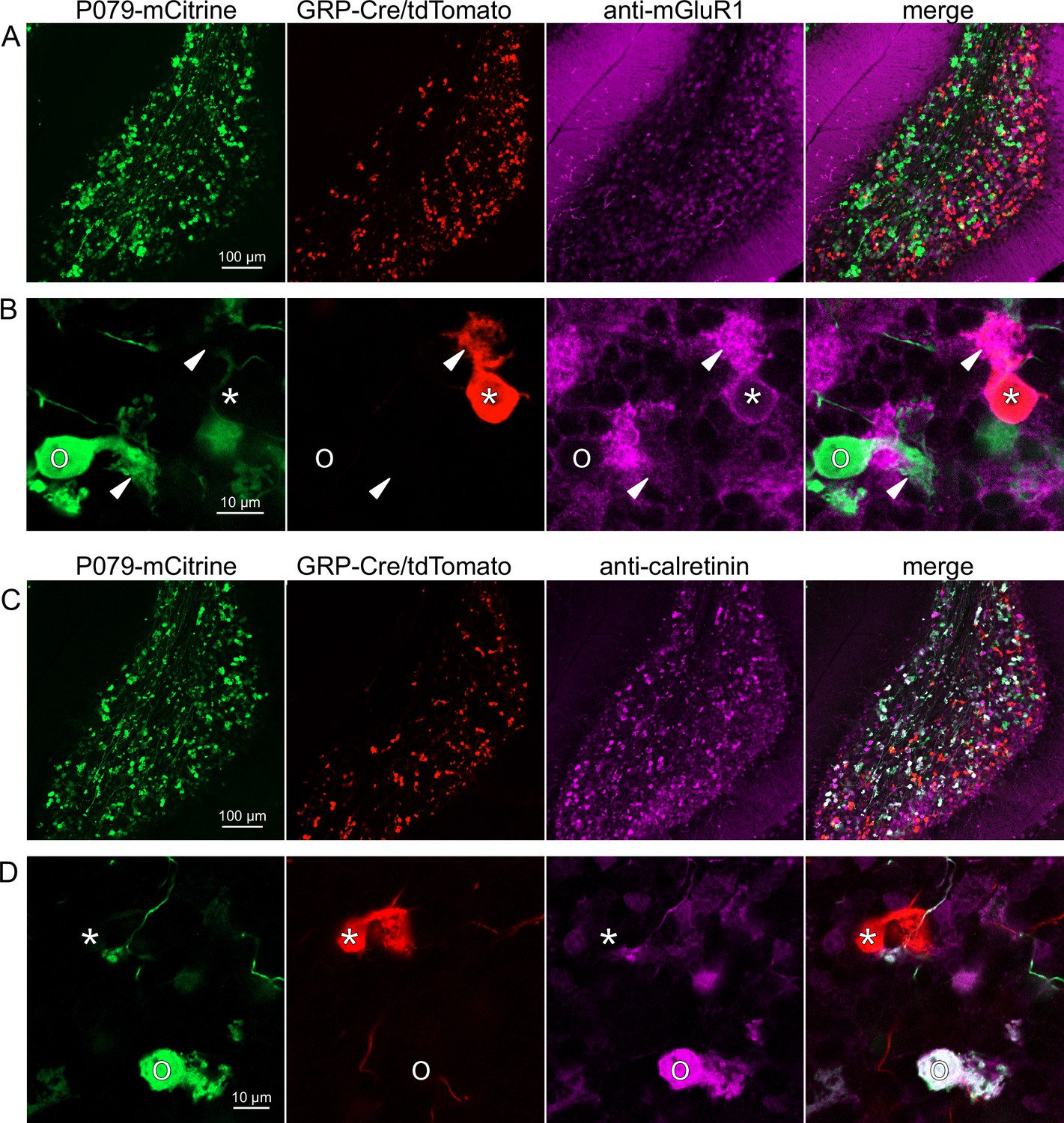
Expression of mGluR1 and calretinin in GRP and P079 unipolar brush cells (UBCs) in the cerebellum.
(A) Sagittal section of lobe X showing genetically expressed mCitrine (green) from the P079 mouse, tdTomato (red) from the GRP-Cre/Ai9 mouse, and immunohistochemical labeling of mGluR1 (magenta). Maximum intensity projections. (B) Example of a P079 UBC (soma indicated with O, brush indicated with arrowhead), GRP UBC (soma indicated with *, brush indicated with arrowhead). The GRP UBCs expresses mGluR1 in the somatic membrane and dendritic brush. Most P079 UBCs do not express mGluR1. Single image planes. (C) Sagittal section of lobe X showing genetically expressed mCitrine (green) from the P079 mouse, tdTomato (red) from the GRP-Cre/Ai9 mouse, and immunohistochemical labeling of calretinin (magenta). Maximum intensity projections. (D) Example of a P079 UBC (soma indicated with O), GRP UBC (soma indicated with *). The P079 UBCs express calretinin in their cytoplasm. GRP UBCs do not express calretinin.
Figure 4
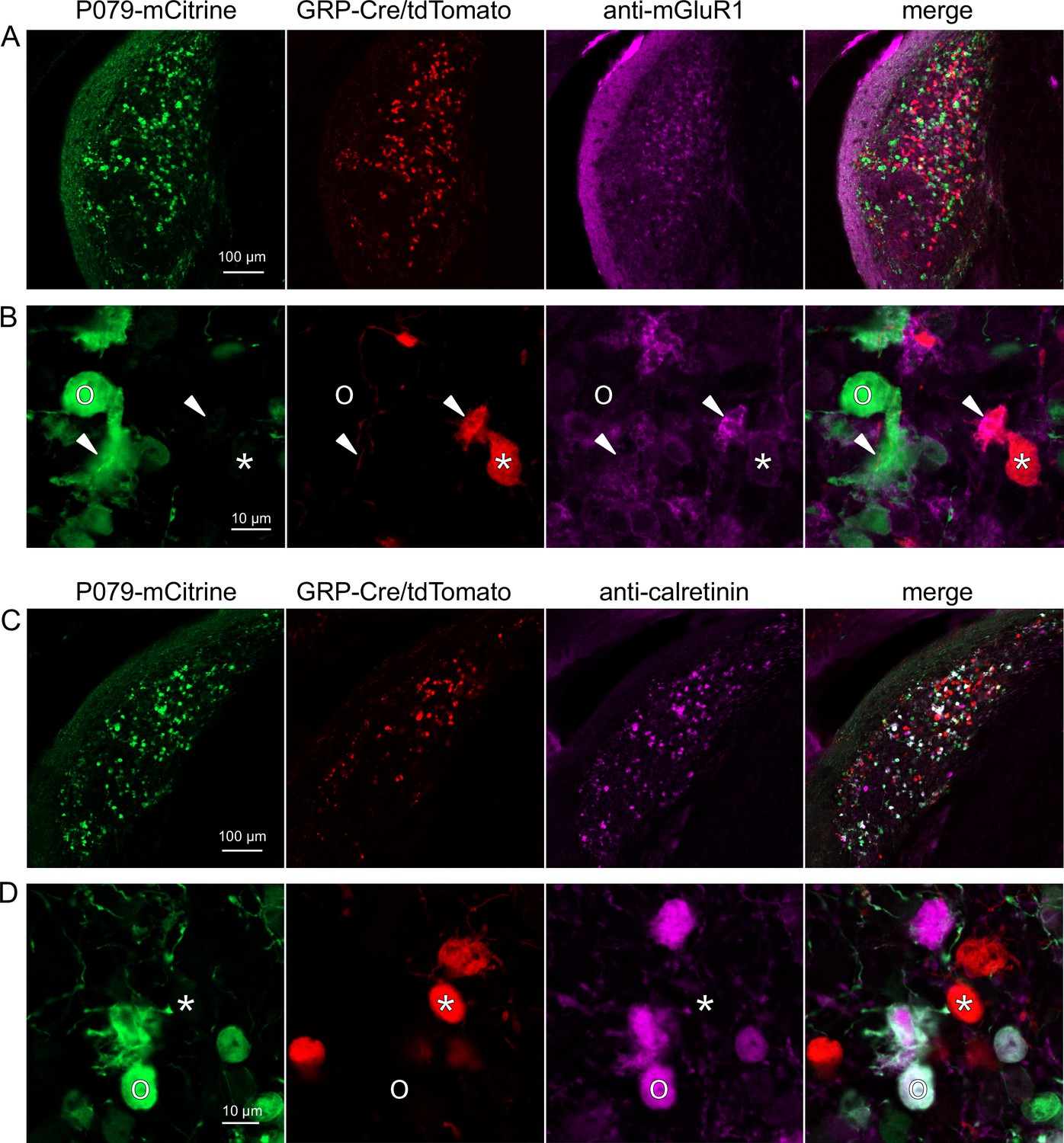
Expression of mGluR1 and calretinin in P079 and GRP unipolar brush cells (UBCs) in the dorsal cochlear nucleus.
(A) Coronal section of the dorsal cochlear nucleus showing genetically expressed mCitrine (green) from the P079 allele, tdTomato (red) from the GRP-Cre/Ai9 alleles, and immunohistochemical labeling of mGluR1 (magenta). Maximum intensity projections. (B) Example of a P079 UBC (soma indicated with O, brush indicated with arrowhead), GRP UBC (soma indicated with *, brush indicated with arrowhead). The GRP UBCs expresses mGluR1 in the somatic membrane and dendritic brush. Most P079 UBCs do not express mGluR1, although some that are weakly labeled do appear to express mGluR1. (C) Coronal section of the dorsal cochlear nucleus showing genetically expressed mCitrine (green) from the P079 allele, tdTomato (red) from the GRP-Cre/Ai9 alleles, and immunohistochemical labeling of calretinin (magenta). Maximum intensity projections. (D) Example of a P079 UBC (soma indicated with O), GRP UBC (soma indicated with *). The P079 UBCs express calretinin in their cytoplasm. GRP UBCs do not express calretinin.
Figure 5
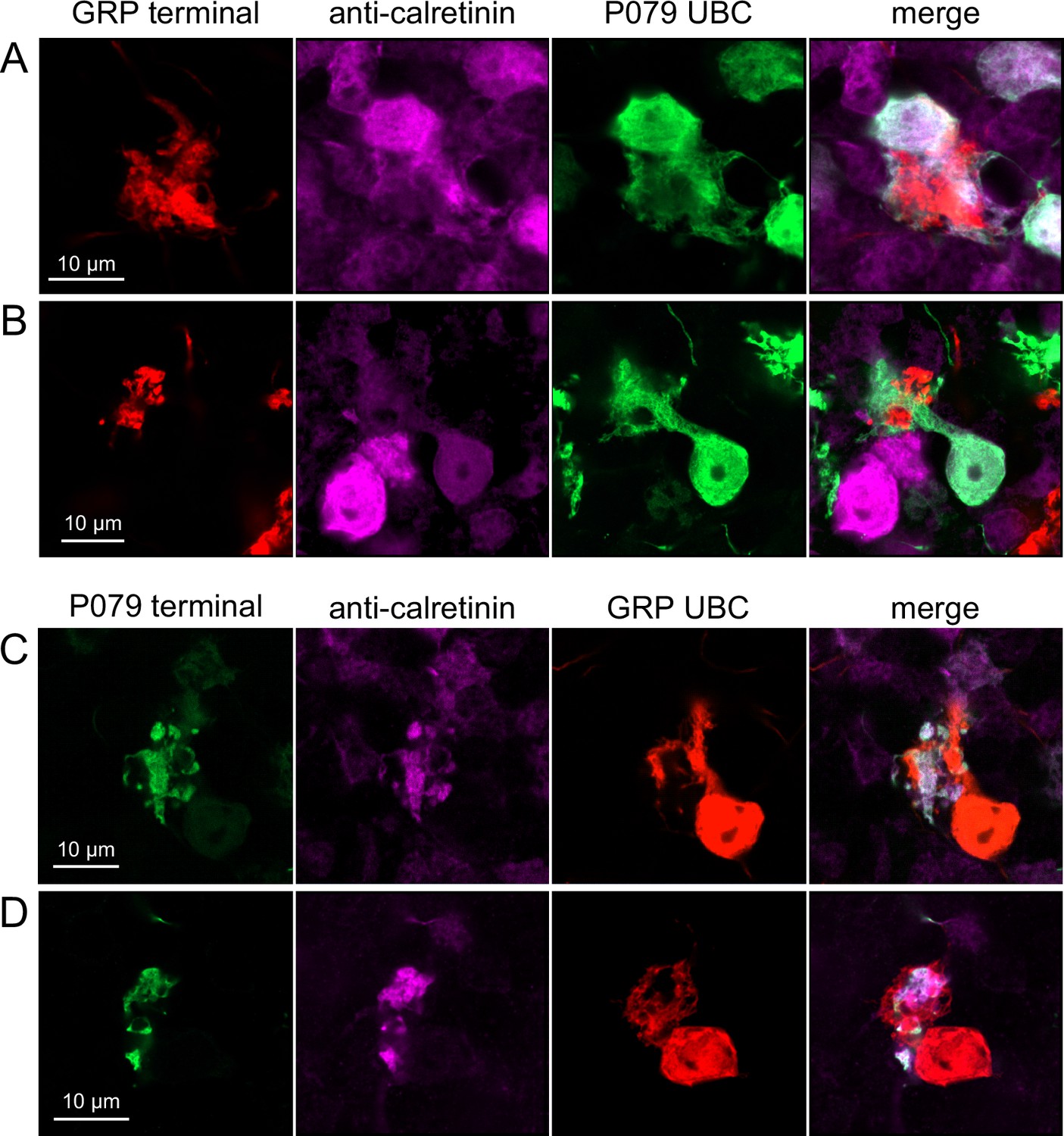
Anatomical evidence showing axonal projections between ON and OFF unipolar brush cell (UBC) subtype.
(A, B) ON UBC axon terminals (GRP, red) that project to OFF UBCs (P079, green), confirmed to express calretinin (magenta). Single image sections. (C, D) OFF UBC axon terminals (P079, green), confirmed to express calretinin (magenta) contacting the dendritic brushes of ON UBCs (GRP, red). Single image sections.
Figure 6
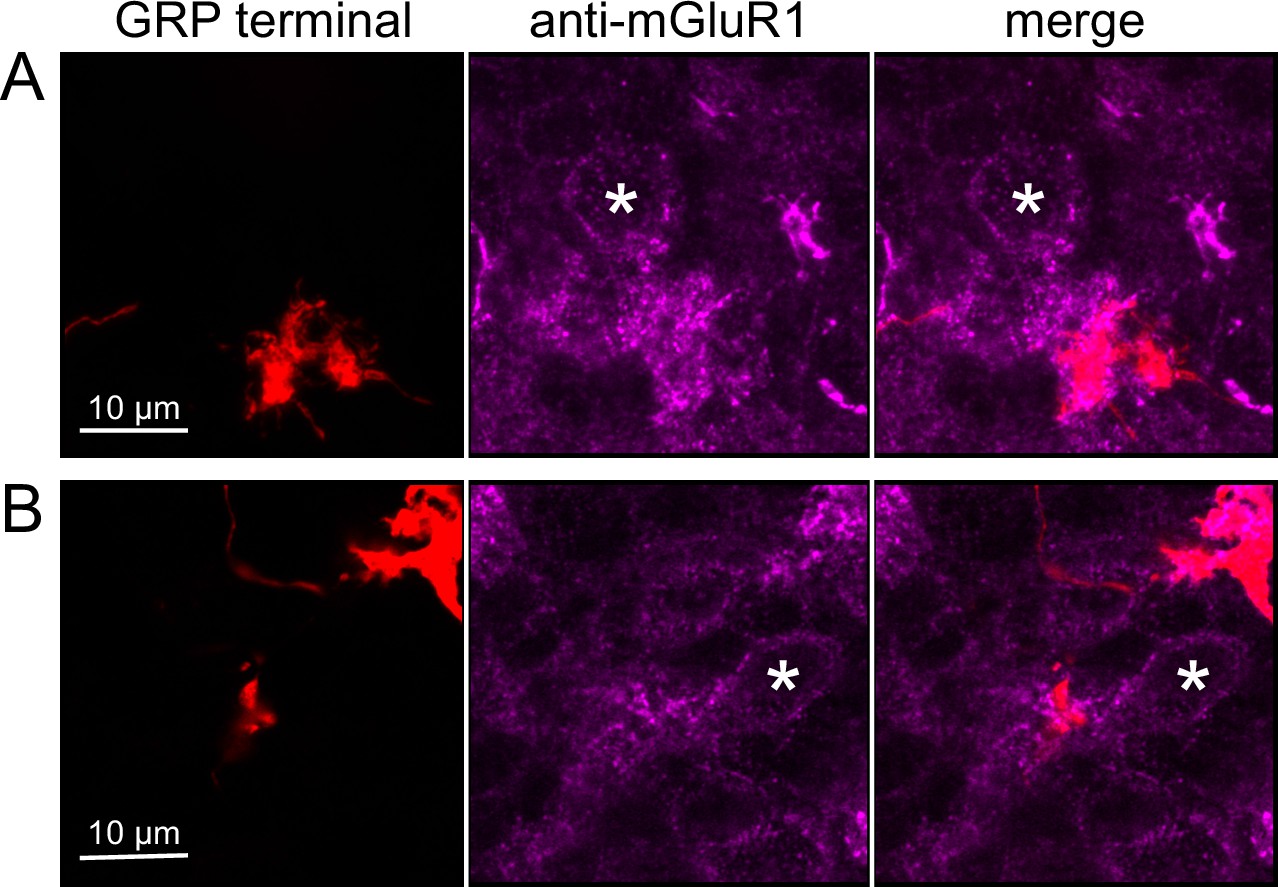
Anatomical evidence for synaptic connections between ON unipolar brush cells (UBCs).
(A, B) GRP ON UBC terminals (red) contacting mGluR1(+) ON UBCs (magenta). Somas indicated with *. Single image sections.
Figure 7
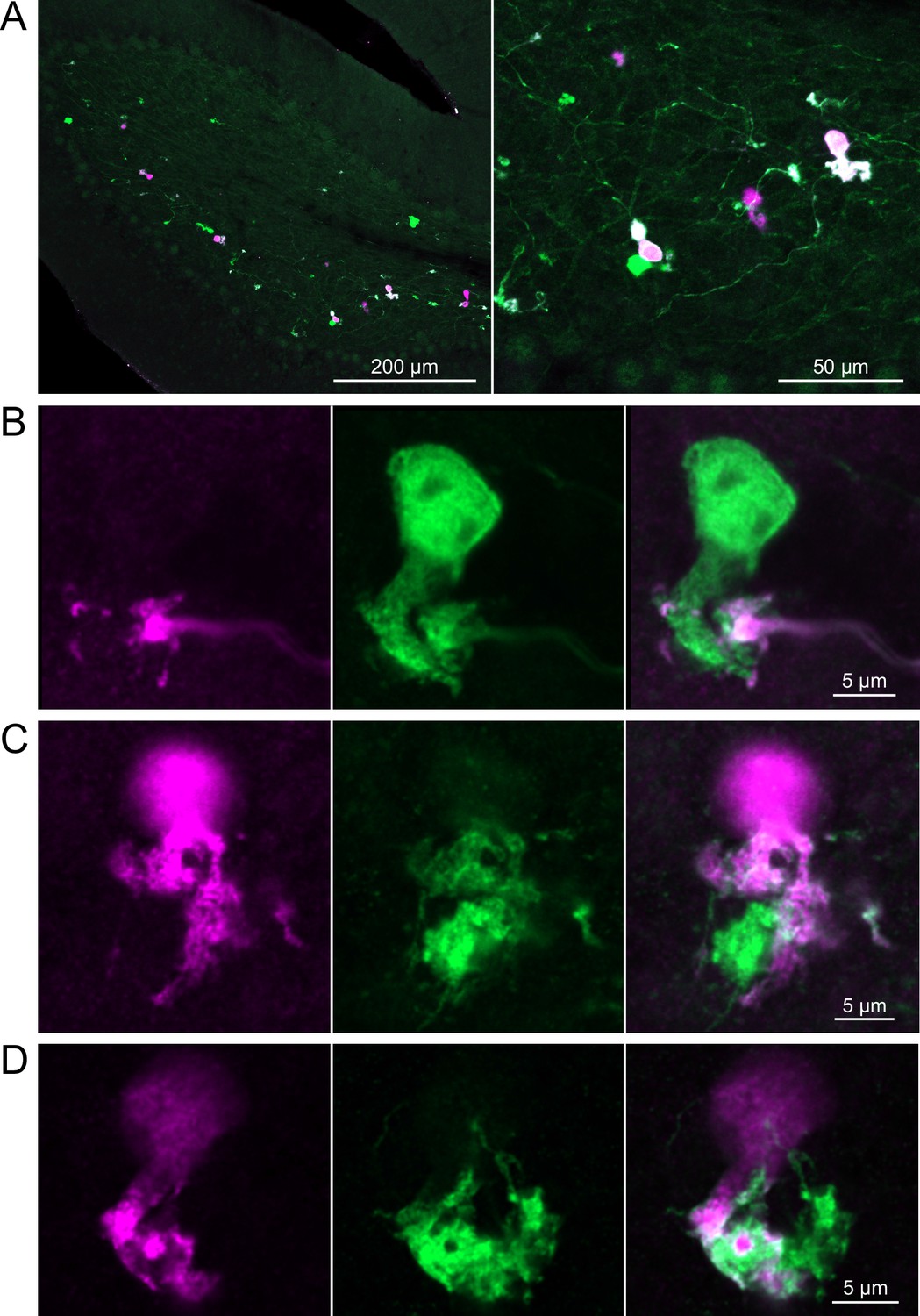
Brainbow reporter labels synaptic connections between GRP ON unipolar brush cells (UBCs).
(A) GRP ON UBCs expressed fluorescent proteins that were either amplified with the anti-GFP antibody or anti-mCherry antibody. GRP UBCs and their axons and terminals are labeled. Maximum intensity projections. (B) Example of a GRP UBC expressing cytoplasmic YFP (green) that appears to be contacted by a presynaptic terminal from the axon of another GRP UBC that expresses both cytoplasmic YFP (green) and cytoplasmic tdimer2(12) (magenta). Single image sections. (C) Example of GRP ON UBC expressing cytoplasmic tdimer2(12) (magenta) and membrane-bound mCerulean (green) that appears to receive a synaptic terminal that expresses cytoplasmic YFP (green). Single image sections. (D) Example of GRP ON UBC that expresses cytoplasmic tdimer2(12) (magenta) receiving input from a GRP UBC axon that expresses cytoplasmic YFP (green). Single image sections.
Figure 8
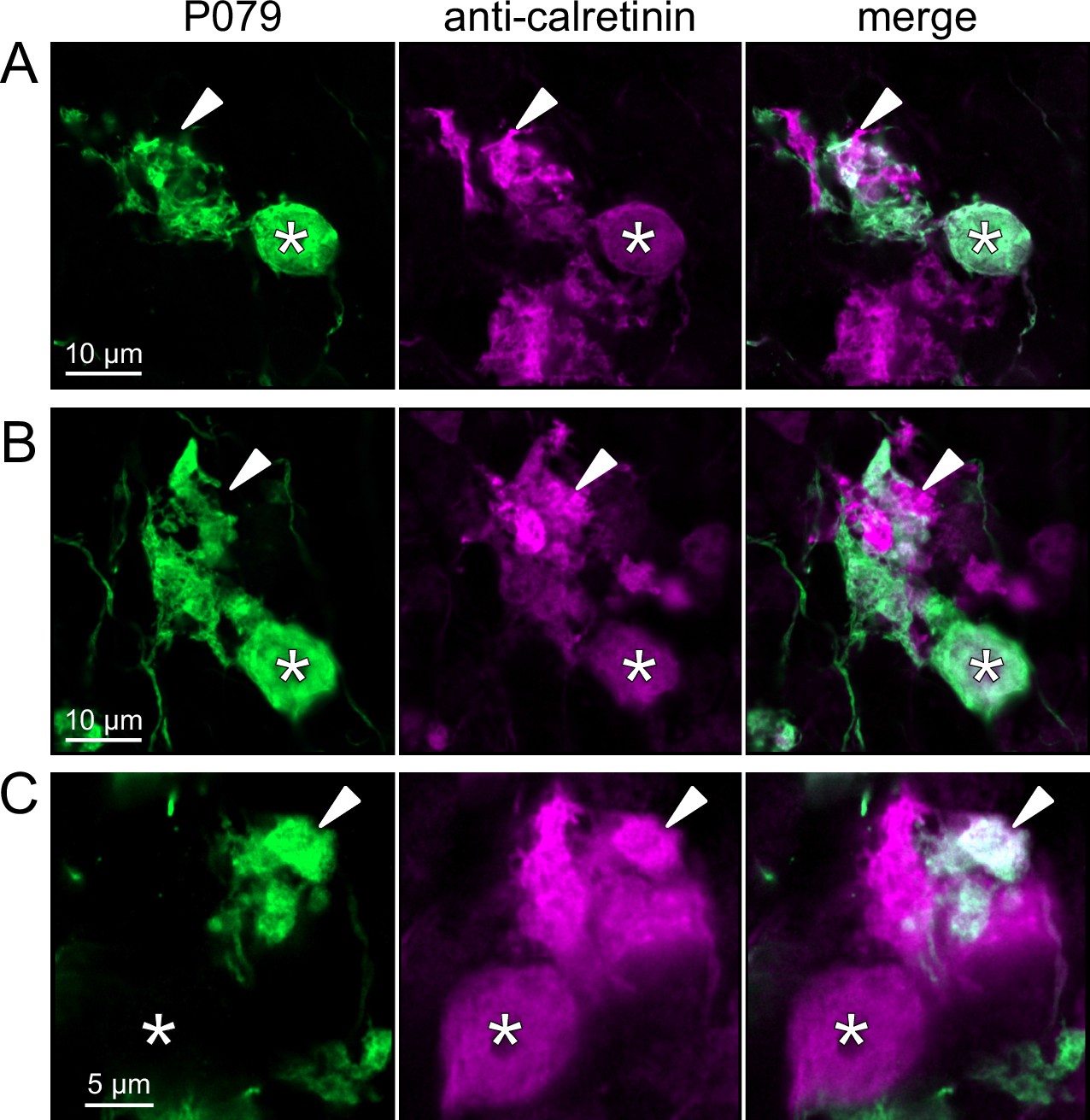
Anatomical evidence for synaptic connections between OFF unipolar brush cells (UBCs).
(A, B) Examples of P079 OFF UBCs (green) that are confirmed to express calretinin (magenta) and are contacted by calretinin(+) terminals that are not labeled in the P079 line. Single image sections. (C) Example of an OFF UBC that expresses calretinin (magenta) that is contacted by a P079 (green) axon terminal that expresses calretinin (magenta). Somas are indicated with *, terminals are indicated with arrowheads. Single image sections.
Figure 9
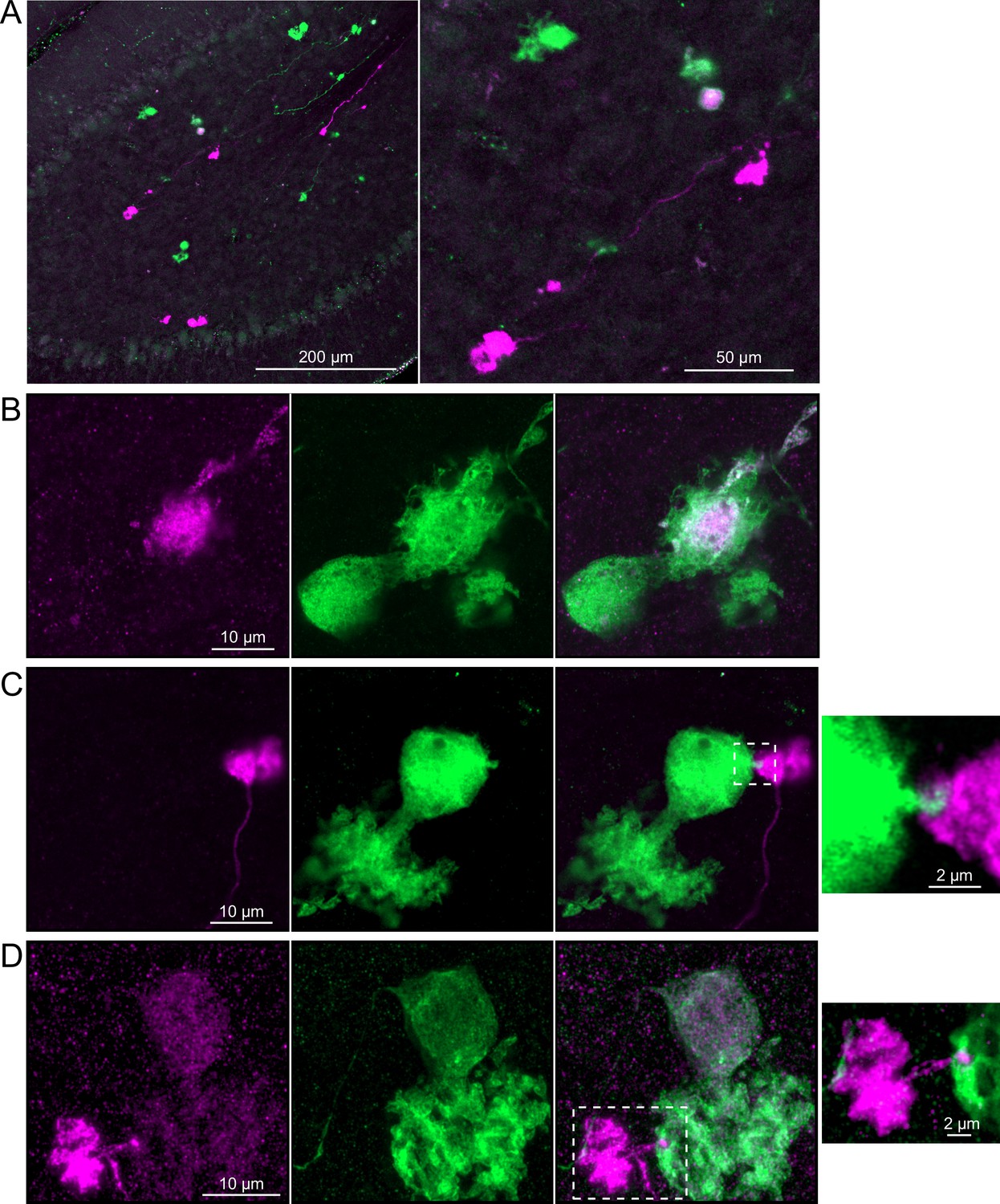
Brainbow reporter labels synaptic connections between OFF unipolar brush cells (UBCs) in calretinin-Cre mouse line.
(A) Calretinin-Cre UBCs expressed fluorescent proteins that were either amplified with the anti-GFP antibody or anti-mCherry antibody, revealing UBCs and their axons and terminals in lobe X. Maximum intensity projections. (B) Example of a labeled calretinin-Cre axon terminal (magenta) that contacted the brush of a calretinin-Cre UBC (green), showing that these OFF UBC project to one another. (C) Example of a calretinin-Cre UBC axon terminal (magenta) that appeared to contact a spine-like extension from the soma of another calretinin-Cre UBC (green). (D) Example of a calretinin-Cre axon terminal (magenta) that projected a small fiber that made a bouton-like synapse onto the brush of another calretinin-Cre UBC (green) that is presumed to have another unlabeled input to the majority of its dendritic brush.
Figure 10
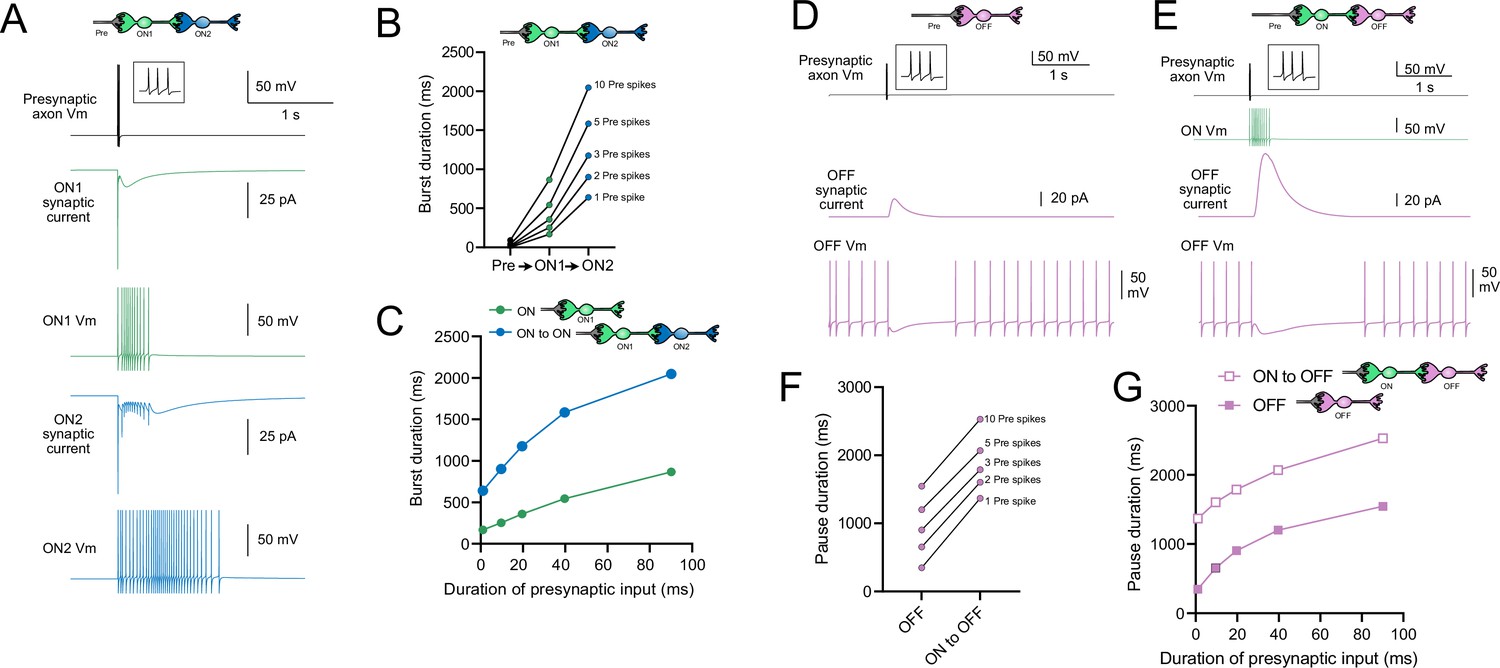
Model predicts that intermediate ON unipolar brush cells (UBCs) extend signal.
(A) Model in which a presynaptic axon fired three action potentials (inset shows presynaptic spikes with expanded time scale) that generated an AMPA receptor-mediated synaptic current and a burst in spikes in the first ON UBC (ON1), which produced an extended AMPA receptor-mediated synaptic current in the postsynaptic UBC (ON2) and an extended burst of spikes. (B) Presynaptic spikes were amplified to longer bursts in each subsequent ON UBC. (C) Burst duration in the intermediate ON UBC (green) and second ON UBC (blue) as a function of presynaptic input duration. (D) Model in which a presynaptic axon fired three action potentials that generated an mGluR2-mediated synaptic current in an OFF UBC, which produced a pause in spontaneous firing lasting ~1 s. (E) An intermediate ON UBC between the presynaptic axon and OFF UBC generated a larger and longer-lasting mGluR2-mediated current that produced an extended pause in spontaneous firing. (F) Pause duration in an OFF UBC was extended by an intermediate ON UBC. (G) Pause duration in an OFF UBC with and without an intermediate ON UBC as a function of presynaptic input duration.
Figure 11
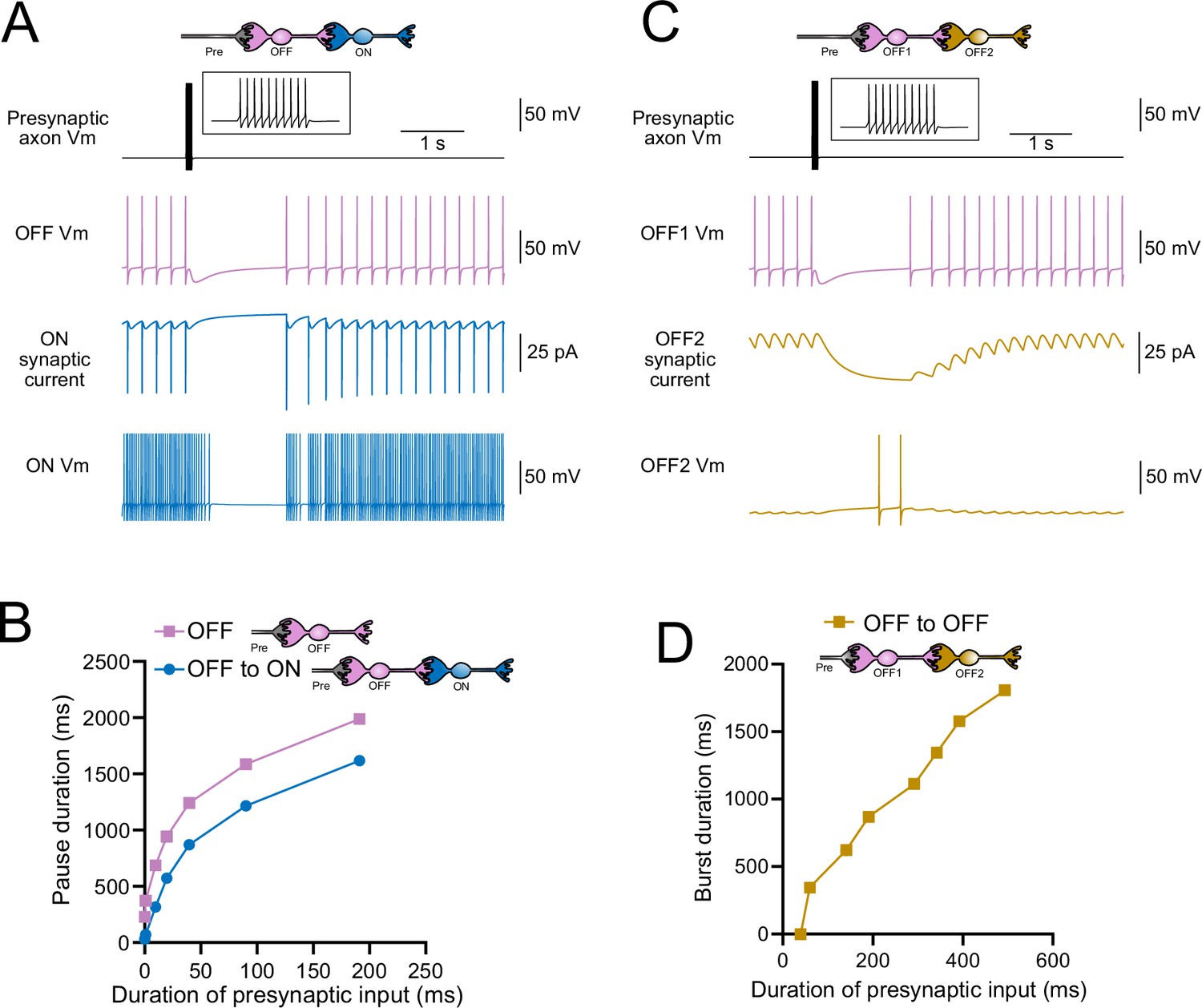
Model predicts that intermediate OFF unipolar brush cells (UBCs) delay signals.
(A) Model showing an intermediate OFF UBC (magenta) that produced a delayed pause in a postsynaptic ON UBC (blue). The spontaneous activity of the intermediate OFF UBC drove irregular spiking in the postsynaptic ON UBC. Presynaptic input (10 spikes shown in inset with expanded time scale) produced a pause in the intermediate OFF UBC, which stopped its release of glutamate onto the ON UBC, and produced a pause after a delay that is due to the slow decay of the AMPA receptor-mediated current. (B) The intermediate OFF UBC (magenta) paused for longer than the postsynaptic ON UBC (blue) because the pause in the ON UBC occurred after a delay and it ended as soon as the intermediate OFF UBC resumed firing. (C) Model showing an intermediate OFF UBC that produced delayed spikes in a postsynaptic OFF UBC. Spontaneous firing of the intermediate OFF UBC (OFF1, magenta) tonically inhibited the postsynaptic OFF UBC (OFF2, gold). Presynaptic input caused a pause in the intermediate OFF UBC, which disinhibited the postsynaptic OFF UBC, allowing it to fire with a delay that depended on the decay of the mGluR2 current. (D) The duration of the burst of action potentials in the postsynaptic UBC increased with longer durations of presynaptic input.
Figure 12
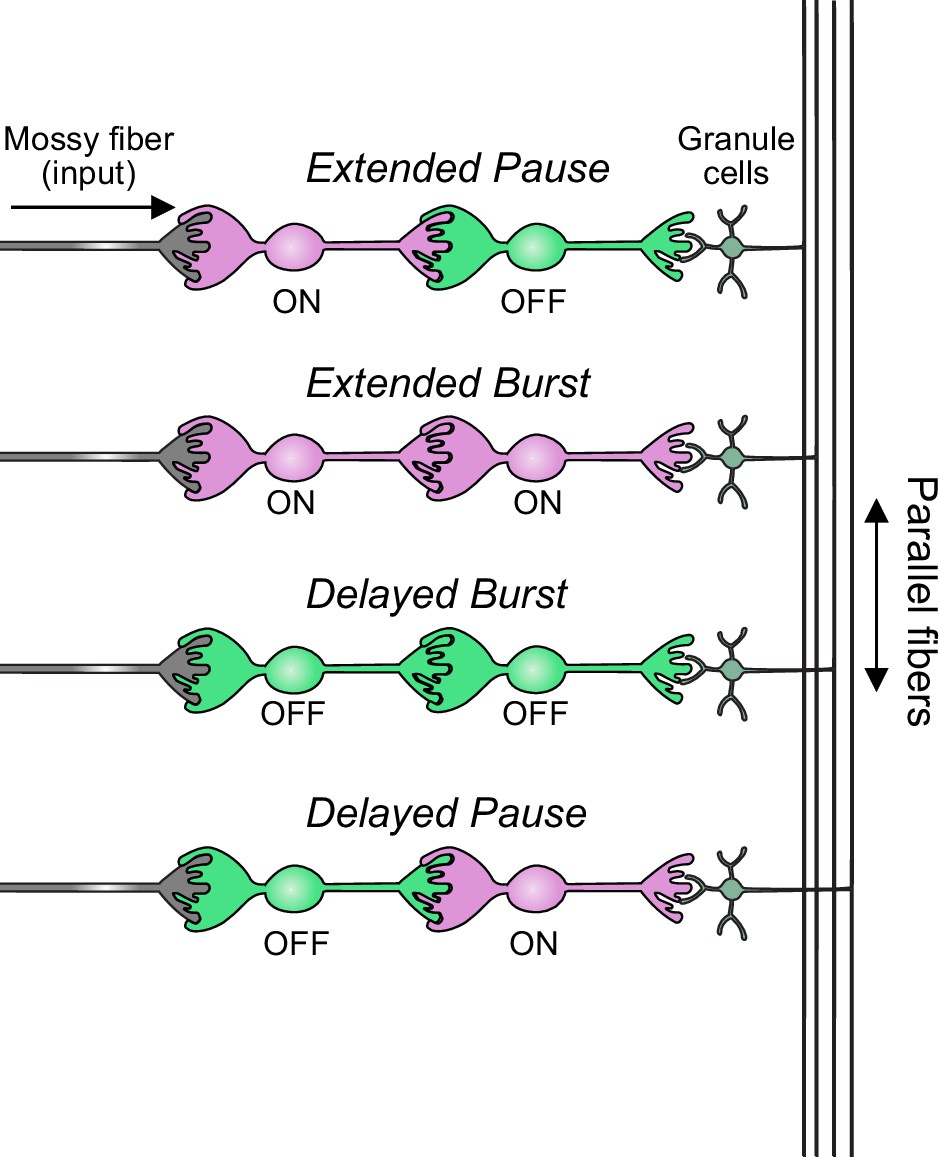
Summary of transformations of spiking patterns in unipolar brush cell (UBC) circuits.
Four patterns of connectivity between UBC subtypes were anatomically defined and computationally modeled. Intermediate ON UBCs extended the usual pause or burst in firing of the postsynaptic OFF or ON UBC. Intermediate OFF UBCs delayed and inverted the response of the postsynaptic UBC, producing either a delayed burst in OFF UBCs or a delayed pause in ON UBCs.
Author response image 1

Example showing small effect of GABAergic and glycinergic inhibition on excitatory currents and burst duration.
(A) Excitatory postsynaptic currents in response to train of 10 presynaptic stimuli at 50 Hz before (black) and after (Grey) blocking GABA and glycine receptors. The slow inward current that occurs at the end of stimulation is little affected. (B) Expanded view of the synaptic currents evoked during the train of stimuli. GABA/glycine receptors mediate the fast outward currents that occur immediately after the first couple stimuli. (C) Three examples of the bursts caused by the 50 Hz stimulation in the same cell without blocking GABA and glycine receptors. (D) Three examples in the same cell after blocking GABA and glycine receptors.
Author response image 2

Example of GRP UBC with disynaptic input.
Three examples of the effect of a single presynaptic stimulus (triangle) in a GRP UBC with presumed disynaptic input. Note the variable latency of the first evoked EPSC, bursts of EPSCs, and spontaneous EPSCs.
Author response image 3

Effect of short-term depression of neurotransmitter release.
(A) The top trace shows the glutamate transient that drives the AMPA receptor model used in our study. No change in release is implemented, although the slow tail of the transient summates during the train. The bottom trace shows the modeled AMPA receptor mediated current. (B) In this model the amount of glutamate released on each stimulus is reduced by 10%. The duration of the slow AMPA current is similar, despite a profound change in the pattern of neurotransmitter exposure.
Author response image 4

Examples of UBCs that receive spontaneous input.
(A) Three ON UBCs that had spontaneous EPSCs, suggesting the presence of an active presynaptic UBC. (B) Two OFF UBCs that had spontaneous outward currents.
Tables
Table 1
ON unipolar brush cell (UBC) conductances.
| ON UBC | ||
|---|---|---|
| Modeled conductance | gmax (S/cm2) | Erev (mV) |
| gNa | 0.1 | 50 |
| gK | 0.03 | –90 |
| gH | 1e-5 | –30 |
| gpas | 2e-4 | –65 |
| gK-slow | 8e-4 | –90 |
| Diameter | 20 µm | |
Table 2
OFF unipolar brush cell (UBC) conductances.
| OFF UBC | ||
|---|---|---|
| Modeled conductance | gmax (S/cm2) | Erev (mV) |
| gNa | 0.1 | 50 |
| gK | 0.03 | –90 |
| gH | 3.1e-4 | –30 |
| gpas | 7e-5 | –62 |
| gK-slow | 8e-4 | –90 |
| Diameter | 27 µm | |
Table 3
Glutamate diffusion at AMPA and mGluR2 receptors.
| Glutamate diffusion parameter | AMPA synapse | mGluR2 synapse |
|---|---|---|
| Molecules released (#) | 3e6 | 3e5 |
| Distance (nm) | 750 | 1750 |
| Diffusion coefficient (cm2/s) | 0.33 | 0.33 |
| Tortuosity | 1.55 | 1.55 |
| Volume fraction | 0.21 | 0.21 |
| Ambient glutamate (mM) | 0.005 | 0 |
Table 4
Synaptic conductances.
| Synaptic receptor | gmax (S/cm2) | Erev (mV) |
|---|---|---|
| ON UBC, AMPA | 4e-9 | 0 |
| OFF UBC, mGluR2 | 1e-8 | –90 |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A system of feed-forward cerebellar circuits that extend and diversify sensory signaling
eLife 12:RP88321.
https://doi.org/10.7554/eLife.88321.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}