Genetic and karyotype divergence between parents affect clonality and sterility in hybrids
- Laboratory of Non-Mendelian Evolution, Institute of Animal Physiology and Genetics of the CAS, Czech Republic
- Military Health Institute, Military Medical Agency, Czech Republic
- Laboratory of Cell Regeneration and Plasticity, Institute of Animal Physiology and Genetics of the CAS, Czech Republic
- Faculty of Fisheries and Protection of Waters, South Bohemian Research Center of Aquaculture and Biodiversity of Hydrocenoses, University of South Bohemia in Ceske Budejovice, Czech Republic
- Laboratory of Fish genetics, Institute of Animal Physiology and Genetics of the CAS, Czech Republic
- Department of Biology and Ecology, Faculty of Science, University of Ostrava, Czech Republic
- Leibniz-Institute of Freshwater Ecology and Inland Fisheries - IGB, Germany
Figures
Figure 1 with 3 supplements
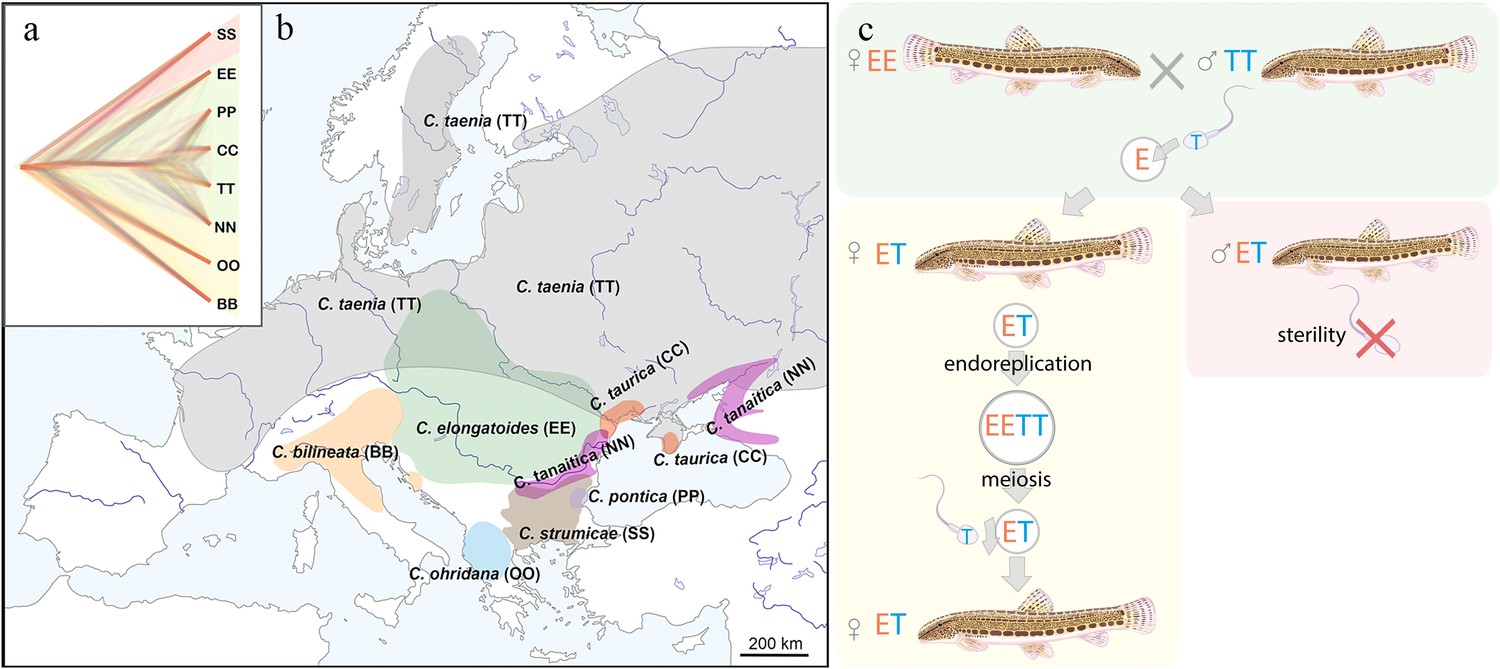
Distribution of the spined loaches used in this study, and schematic representation of the reproduction in hybrids.
(a) Phylogenetic tree based on exome-wide SNP data of the species used in the current crossing experiments. Red color indicates Bicanestrini group; green color indicates Cobitis sensu stricto group; yellow color indicates Adriatic group. (b) Distribution map of the spined loaches (Cobitis) included in this study. While Cobitis elongatoides, C. taenia, C. tanaitica, are known to act as parental species in hybridization events and emergence of clonal lineages, the other species (C. strumicae, C. ohridana, C. taurica, C. pontica) are known to be involved in secondary hybridization events. Abbreviations for all species in brackets. (c) Reproduction scheme of the clonal hybrids in the C. taenia complex, where E represents a haploid genome of C. elongatoides (orange), and T – of C. taenia (blue). Conventionally, hybridization between EE female and TT male lead to ET sterile males and clonal ET females that reproduce via gynogenesis. Gynogenetic females are pseudogamous, that is females produce diploid eggs via premeiotic genome endoreplication. Such eggs require sperm from a parental species (e.g. T sperm) to activate the embryonic development. The sperm genome is not incorporated in the hybrid’s genome but is instead eliminated after the egg’s activation, skipping karyogamy. All hybrid males are sterile because they do not produce spermatozoa or their extremely rare spermatozoa are aberrant and unable to fertilize eggs.
Figure 1—figure supplement 1
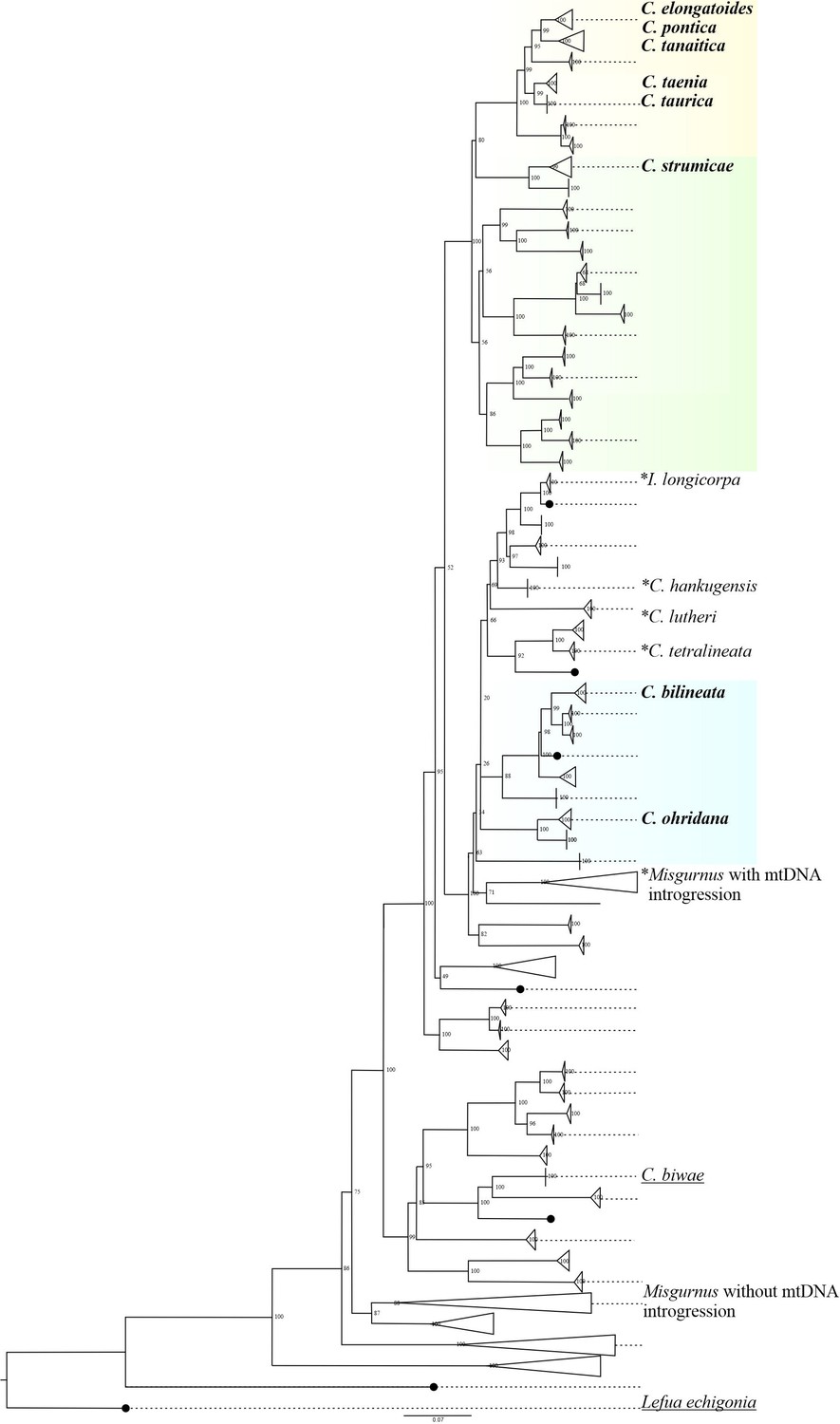
Phylogenetic relationships of the Cobitidae family based on the cytochrome b dataset (modified version of Perdices and co-authors (2016) Perdices et al., 2016).
The species used in the current work for artificial crosses were bold labeled. These species belongs to three major lineages which colonized independently European fresh waters (Perdices et al., 2016). Cobitis sensu stricto lineage (yellow) include all the known species of the C. taenia complex, which known to hybridize and produce asexual diploid and polyploid hybrids. Adriatic lineage (blue) is represented mainly by allopatric species, in this study is represented by C. ohridana and C. bilineata. Bicanestrini group (green) used in this study is represented by C. strumicae. Species used in the current studz are bolded. Additionally, species which were previously reported to interbreed are marked with asterisk; species which were artificially crossed in previous works are underlined, all species contain the following abbreviations: C. biwae = BW; C. lutheri = Lut; C. tetralineata = Tet; M. anguillicaudatus lineage with Cobitis mitochondrial introgression = MAmtC; M.
Figure 1—figure supplement 2
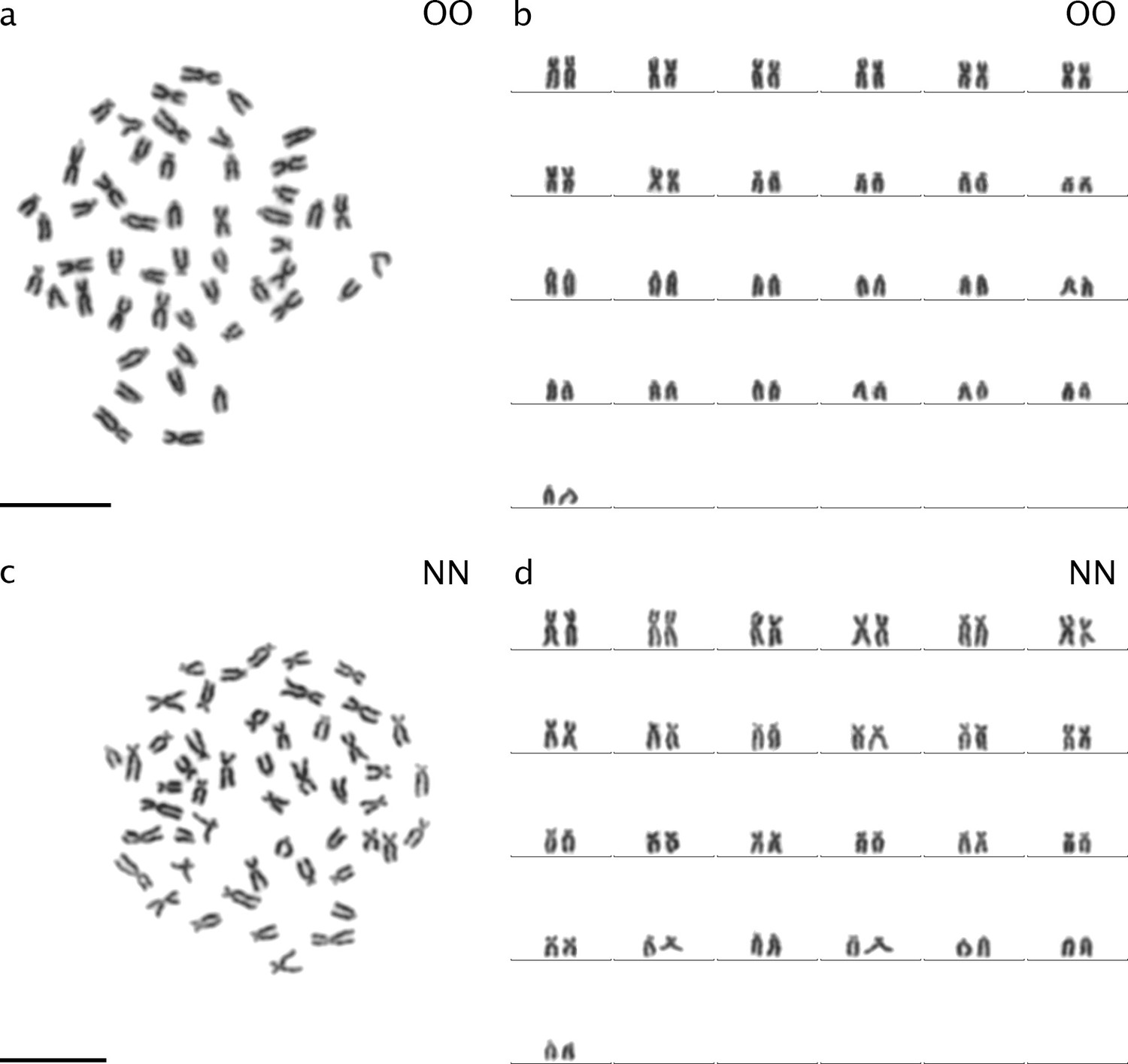
Karyotypes and karyograms of C. ohridana (a, b) and C. tanaitica (c, d).
Both species have 2n=50 chromosomes. C. ohridana has 20 meta-/submetacentric chromosomes and 30 subtelo-/ acrocentric chromosomes. C. tanaitica used in this study has 40 meta-/submetacentric chromosomes and 10 subtelo-/ acrocentric chromosomes. C. ohridana is abbreviated as OO; C. tanaitica is abbreviated as NN.
Figure 1—figure supplement 3

Plot of correlation between Castigla’s AKD index (x-axis) and exome-wide genetic distance (SNP; y-axis).
Figure 2
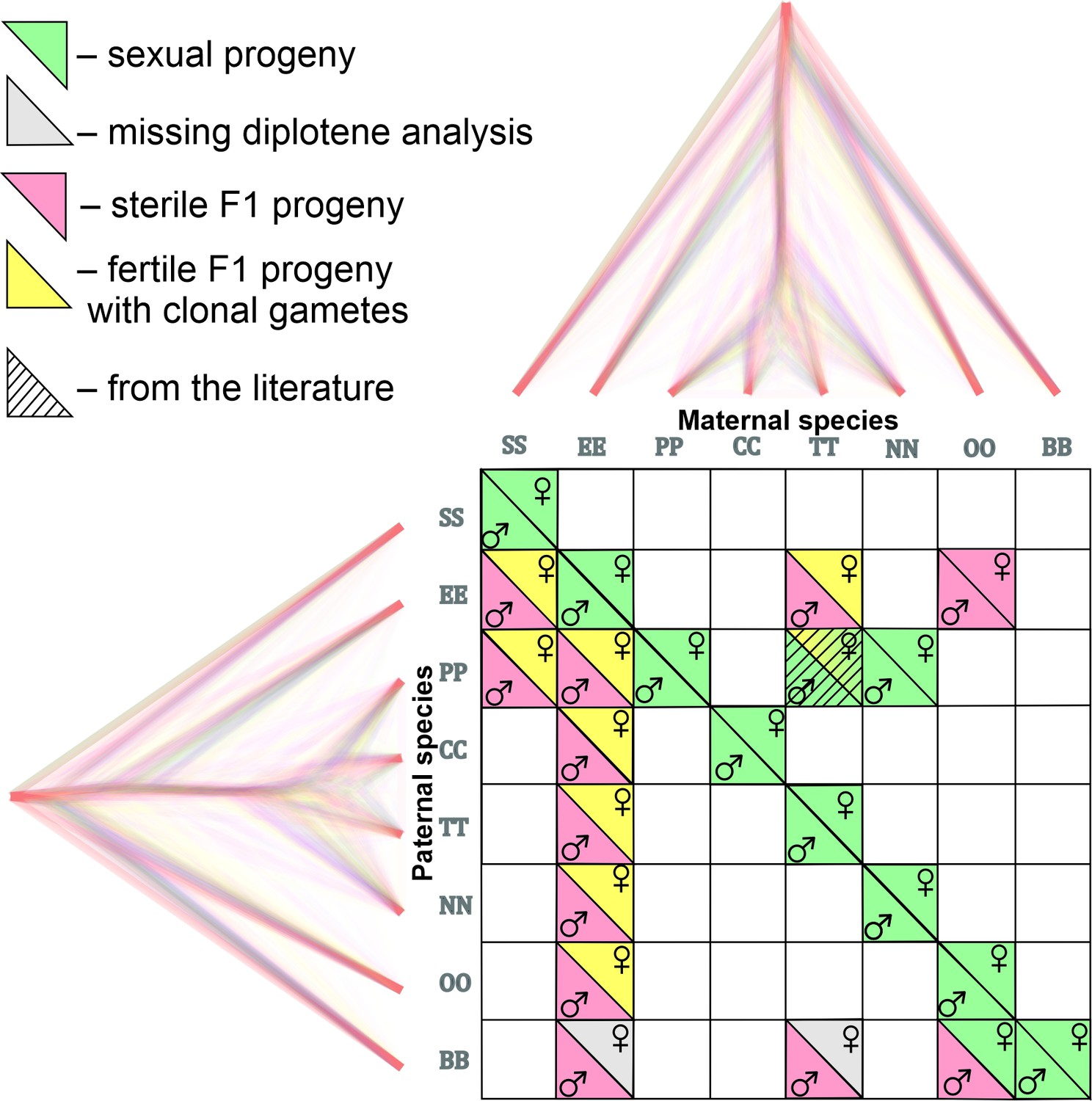
Matrix with successfully established and analyzed crosses between eight species of Cobitis.
Each cell represents a particular cross between parental species including the information about maternal and paternal species (i.e. the direction of the cross). Phylogenomic tree plotted along margins indicates the relationships among crossed species based on exome-wide SNP data. Each color represents a particular reproductive output in F1 hybrids; green labeling indicates production of haploid gametes via normal meiosis; yellow color indicates the F1 progeny which produce unreduced gametes and present fully developed gonads; Red color denotes sterile progeny, predominantly referring to hybrid males. Grey labels potentially sterile hybrids for which we did not obtain diplotenic cells to fully confirm their reproductive output. The striped cell represents the F1 hybrid combination which was previously described Janko et al., 2018 and involves the species used in this study.
Figure 3 with 4 supplements
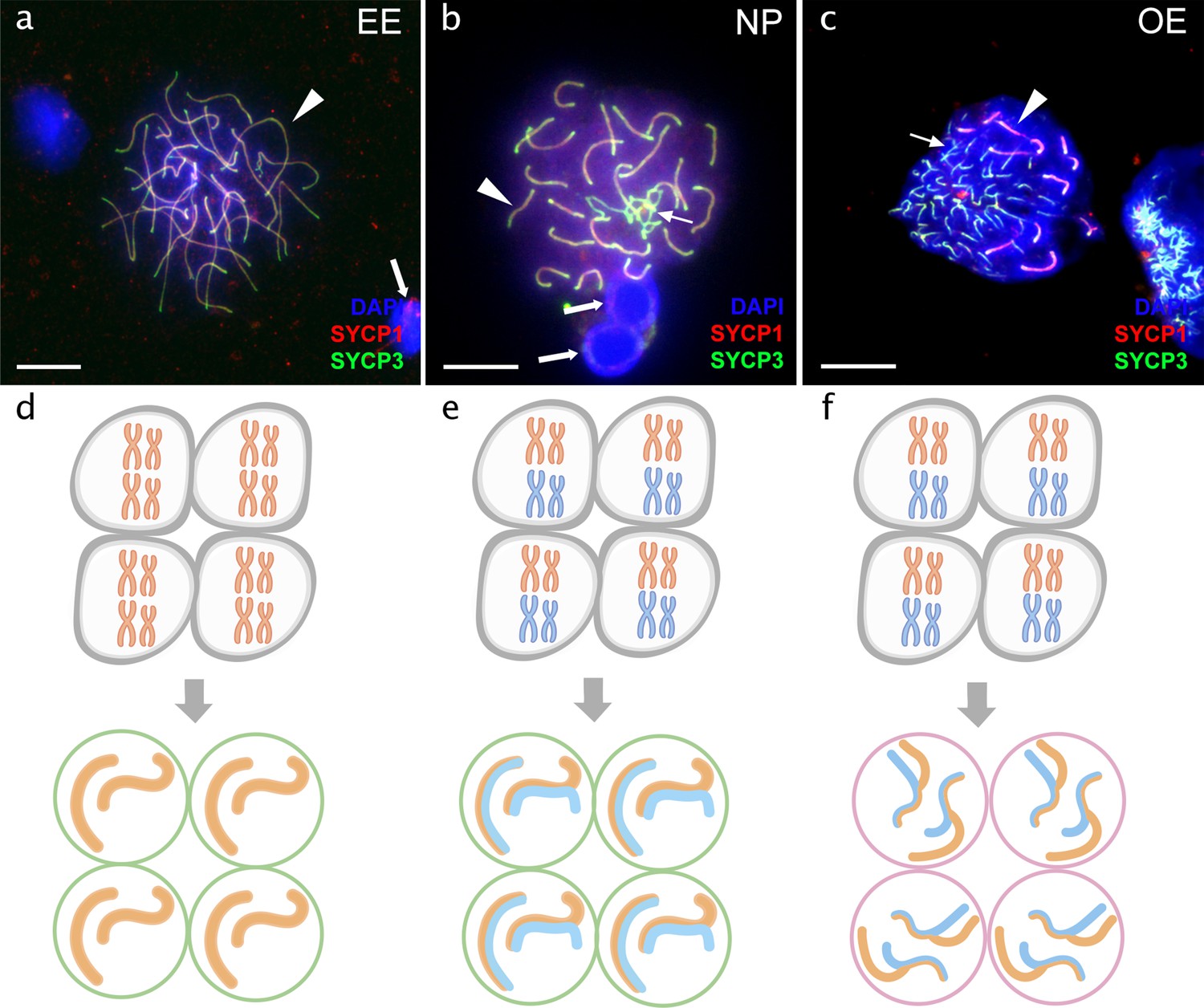
Pachytene spermatocytes in males.
Comparison of pachytene spermatocytes between male of sexual diploid species (a), hybrids’ genotypes (b, c), and corresponding gametogenic pathways (d-f). The spread of pachytene spermatocytes of C. elongatoides with 25 completely paired bivalents (a), and diploid NP hybrid with almost paired chromosomes (b) and OE hybrid with aberrant pairing, including bivalents and univalents (c). Thick arrows indicate bivalents; thin arrows indicate univalents. Scale bar = 10 µm. Schematic representation of the gametogenic pathway including presumptive karyotype composition in gonocytes and pachytene cells in males of sexual species (d) and NP hybrids (e) that are able to complete pairing and form gametes meiotically, and OE hybrids (f) which exhibit abnormal pairing leading to sterility.
Figure 3—figure supplement 1

Visualization of gonial and meiotic cells in male gonads.
Identification of germ cells and meiotic cells in the gonads of TB (a, b), ET (c, d) and OE (e, f) hybrids. Germ cells identified using anti-vasa antibodies (indicated by arrows) were present in all hybrids (a, c, e); meiotic cells during pachytene (indicated by arrowheads) identified using anti SYCP3 and SYCP1 antibodies were not detected in TB hybrids while present in ET and OE hybrids (b, d, f). Scale bars = 10 µm.
Figure 3—figure supplement 2

Pachytene spermatocytes of hybrid males.
Chromosomal spreads of pachytene spermatocytes of diploid EN (a), EB (b), ET (c), OB (d), EC (e), PE (f) and ES (g) males. Synaptonemal complexes are visualized by immunolabeling with antibodies against SYCP3 protein (green) and SYCP1 (red) stained with DAPI (blue). Both SYCP3 and SYCP1 proteins are localized on bivalents (indicated by arrowhead) while only SYCP3 protein is localized on univalent (indicated by thin arrows). Pachytene spreads of diploid hybrid males have incomplete pairing with the number of bivalents and univalents. Scale bar = 10 µm.
Figure 3—figure supplement 3

Relative DNA content of cell nuclei from testes measured by flow cytometry.
Flow cytometry results of the analysis of cell suspension from testis of C. elongatoides (a), and diploid hybrid males (b–h). (a) Sexual male has clear 1 C cells corresponding to haploid sperm; diploid (2 C) cell populations corresponding to spermatogonia; somatic cells and double-diploid (4 C) cell populations corresponding to primary spermatocytes. (b–h) All hybrid males lack the peak of 1 C cells but exhibit diploid (2 C) and high amount of double-diploid (4 C) cell populations accumulated possibly due to problems during chromosomal pairing during pachytene.
Figure 3—figure supplement 4
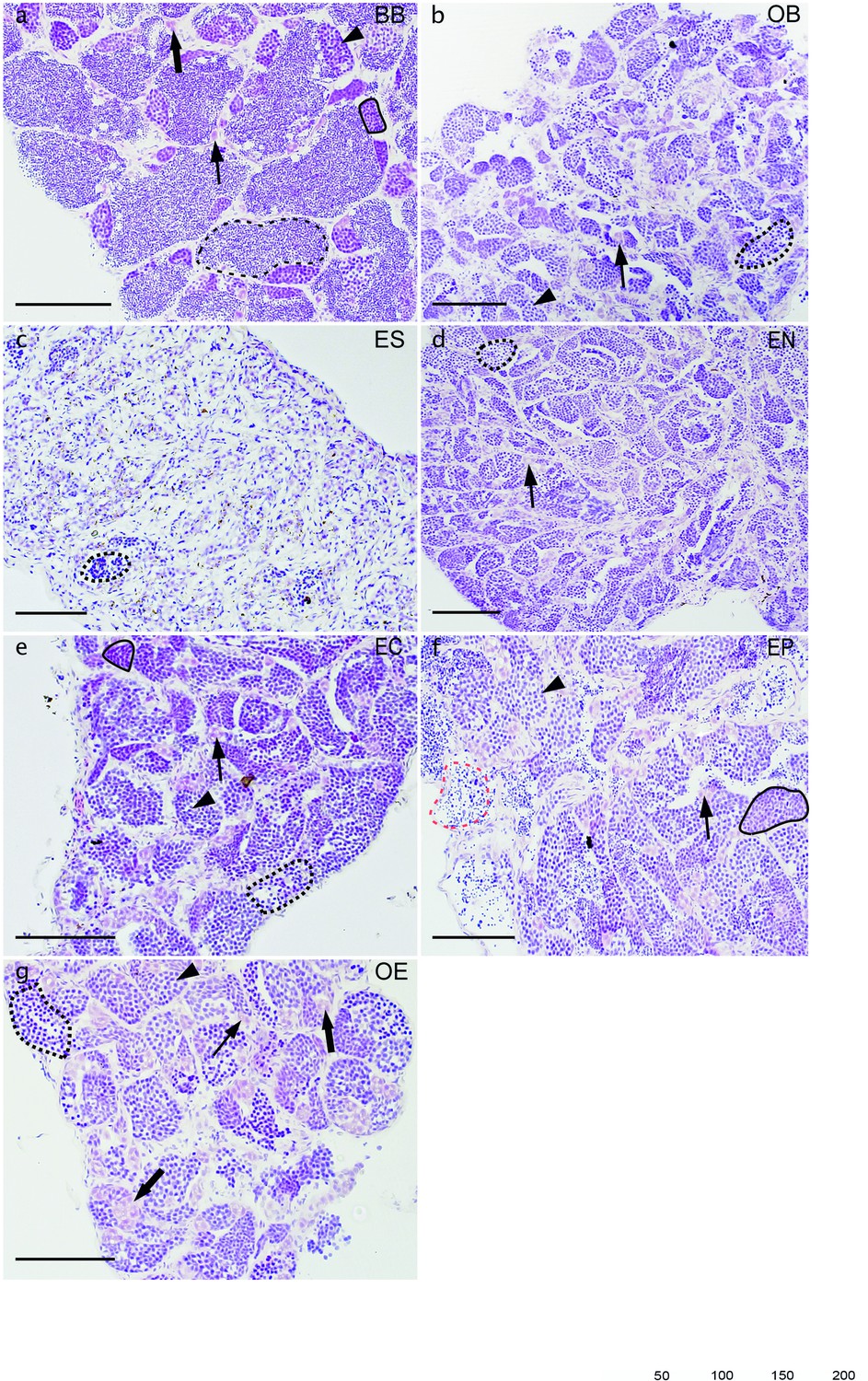
Histological examination of testes.
Comparison of spermatogenesis between male of sexual diploid species C. bilineata (a) and diploid OB (b), ES (c), EN (d), EC (e), EP (f), OE (g) hybrids. Semithin gonadal section of sexual C. bilineata male (a) shows spermatogonia type A (thin arrow), and spermatogonia type B (thick arrow), spermatocytes type I (arrowhead) and spermatocytes type II (black line), as well as spermatozoa (black dash line) in sexual males. Testes of hybrids (b–g) displays defective development with the presence of spermatogonia type A and type B, spermatocytes type I and spermatocytes type II (black line), but only a few spermatids (dotted black line) and/or aberrant spermatozoa (red dash line) germ cells of one cyst at different stages (black dotted line); (b) spermatogonia A with nucleus (N). Scale bar = 100 µm.
Figure 4 with 3 supplements
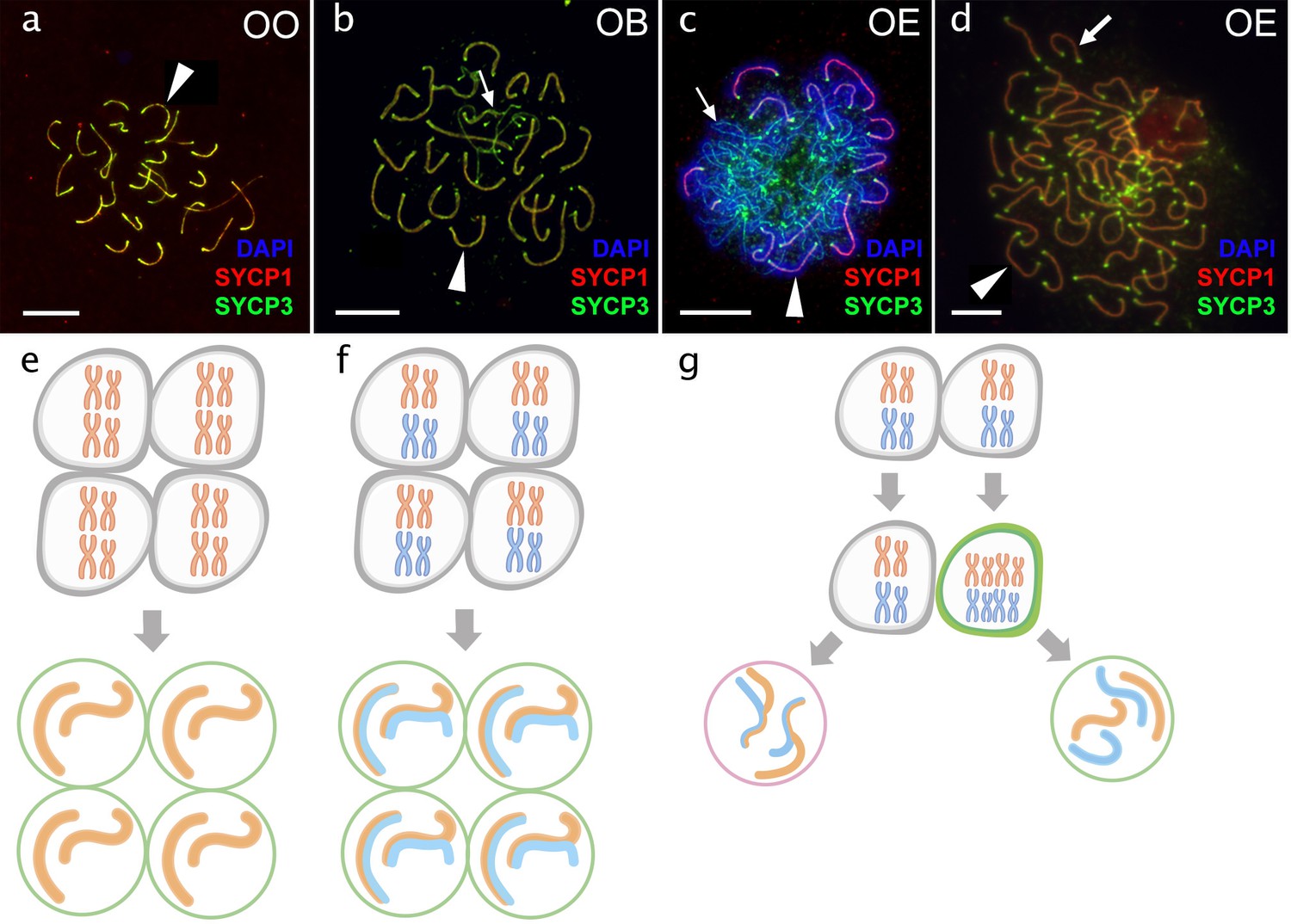
Pachytene oocytes in females.
Comparison of pachytene oocytes between female of sexual diploid species (a) and hybrids genotypes (b, c, d) and corresponding gametogenic pathways (e–h). The SC spreads from pachytene oocyte of C. ohridana with fully paired 25 bivalents (a), diploid OB hybrid with almost paired chromosomes (b) and OE hybrid female which have pachytene cells with aberrant pairing (c) and cells with fully paired chromosomes emerged after premeiotic genome endoreplication (d). Thick arrows indicate bivalents; thin arrows indicate univalents. Scale bar = 10 µm. Schematic representation of gametogenic pathway including presumptive karyotype composition in gonocytes and pachytene cells. Females of sexual species (e) and OB hybrid (f) which are able to fully or partially complete pairing of chromosomes and form gametes meiotically; OE hybrids (g) exhibit two populations of pachytene oocytes: oocytes with unduplicated genomes and oocytes with duplicated genome. Oocytes with unduplicated genome (g, left) have abnormal pairing leading to the inability of proceed beyond pachytene and thus sterility. Oocytes with duplicated genomes (g, right) have normal pairing and thus leading to the formation of unreduced gametes.
Figure 4—figure supplement 1
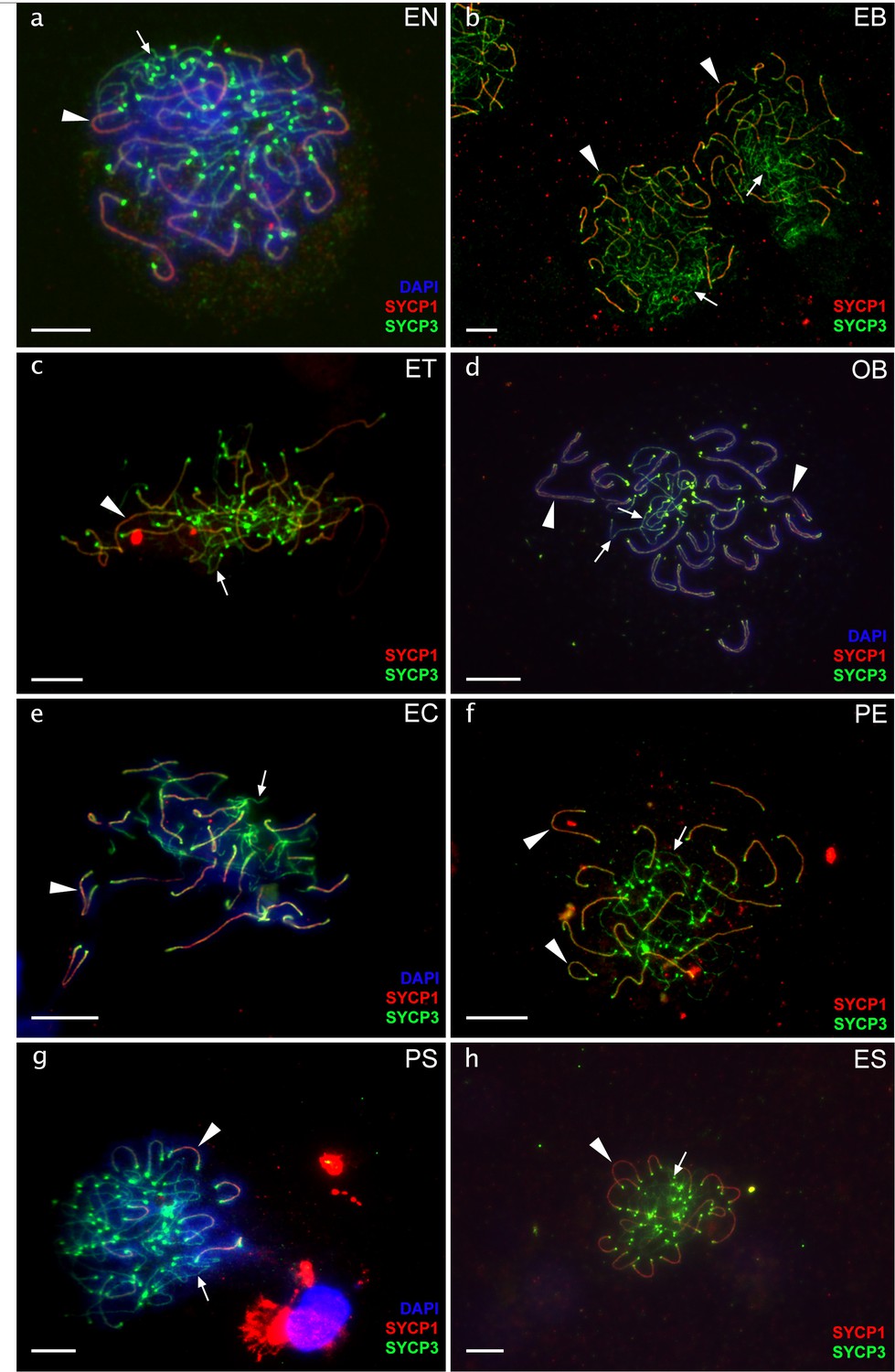
Pachytene oocytes with nonduplicated genomes.
Chromosomal spreads of pachytene oocytes with nonduplicated genomes of diploid EN (a), EB (b), ET (c), OB (d), EC (e), PE (f), PS (g), and ES (h) females. Synaptonemal complexes are visualized by immunolabeling with antibodies against SYCP3 protein (green) and SYCP1 (red) stained with DAPI (blue). Both SYCP3 and SYCP1 proteins are localized on bivalents (indicated by arrows) while only SYCP3 protein is localized on univalent (indicated by arrowheads). Pachytene spreads from oocytes with unduplicated genome in diploid hybrid females exhibit incomplete pairing with the number of bivalents and univalent. Scale bar = 10 µm.
Figure 4—figure supplement 2

Pachytene spreads of sexual species and hybrids.
Chromosomal spreads of pachytene oocytes of sexual species NN (a) and hybrids ET (b), OE (c), OB (d), EC (e) and PE (f) males. Synaptonemal complexes are visualized by immunolabeling with antibodies against SYCP3 protein (green) and recombination loci were detected by antibodies against MLH1 protein (indicated by arrows, red). Chromatin stained with DAPI (blue). Recombination loci were observed only on bivalents, while univalents have no signal. Pachytene spreads from spermatocytes exhibit aberrant pairing. Scale bar = 10 µm.
Figure 4—figure supplement 3

Chromosomal spreads from diplotene oocytes of hybrid females.
Chromosomal spreads from diplotene oocytes of diploid EN (a), ET (b), EC (c), OE (d), PE (e) and OB (f) hybrid females. Full diplotene chromosomal spread from the individual oocyte including 50 (a, c, d, e) and 49 (b) with no univalent or aberrant pairing, suggesting the pairing of homologous chromosome emerged after premeiotic endoreplication. Full diplotene chromosomal spread from OB hybrid is represented by 25 bivalents (f), suggesting the normal pairing of orthologous (O×B) chromosomes. Since the chromosomal spread from individual oocyte was large, several images were taken and merged into one. Arrows show examples of individual bivalents. Scale bar = 50 µm.
Figure 5
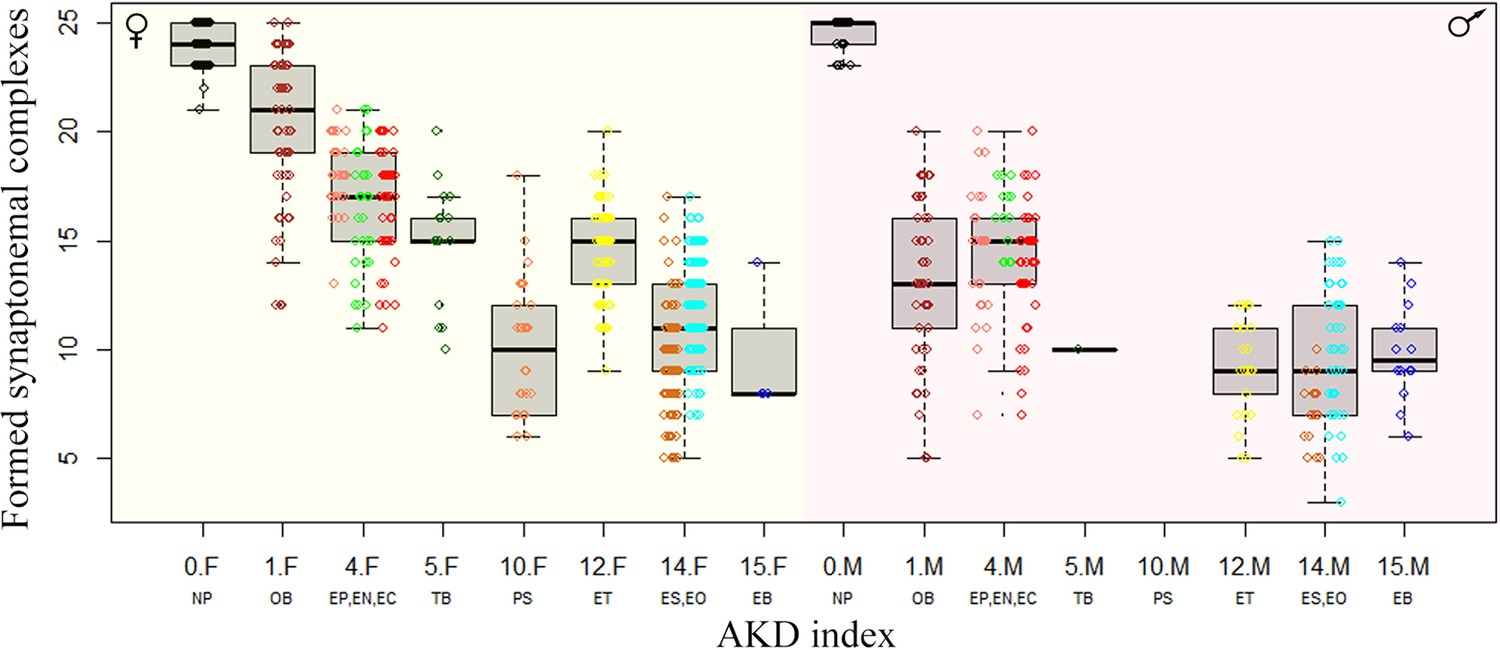
Effect of karyotype dissimilarity on numbers of bivalents in hybrids.
Boxplots showing the number of synaptonemal complexes per cell (y-axis) of the F1 hybrids ranked along x-axis according to the morphological divergence of karyotypes between parental species expressed by AKD index (Castiglia, 2014). Numbers indicate the AKD index values, subscripts F and M indicate females and males, respectively and experimental crosses are indicated by respective letter combinations.
Figure 6
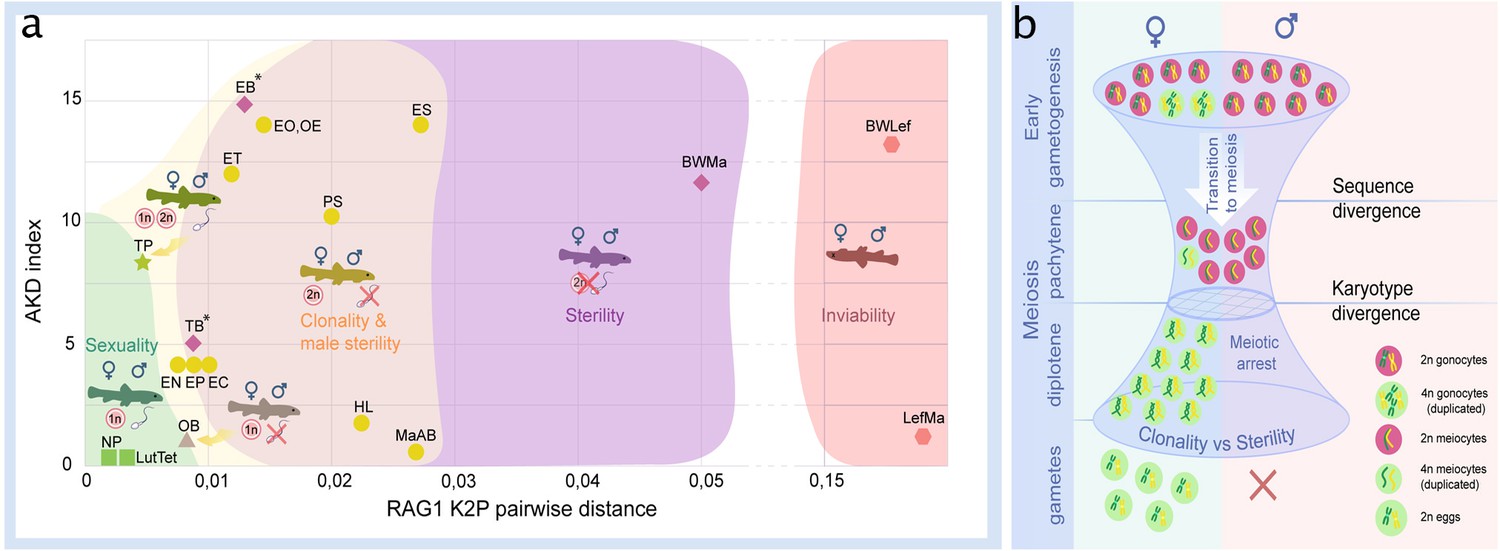
Reproductive outcomes of hybrids from crosses between species from the subfamily Cobitinae.
(a) Plot demonstrating the relationship between reproductive outcome and viability of F1 hybrid loaches and the genetic and karyotype divergences among parental species. Data on F1 hybrids have been obtained in this study as well as from the literature. Karyotype divergence is marked as AKD index and genetic distance is estimated from published RAG1 sequences (K2P distance). Green color shows the ranges of hybrids with sexual reproduction; yellow color shows the ranges of F1 hybrids, which produce unreduced gametes; purple color indicates the ranges of sterility in both sexes; red color shows the ranges of inviable hybrids. To rank reproductive outcomes for every type of cross we labeled them with additional symbols:  – both sexes producing haploid gametes with fully paired chromosomes;
– both sexes producing haploid gametes with fully paired chromosomes;  –both sexes are fertile producing haploid gametes, some females produce unreduced gametes;
–both sexes are fertile producing haploid gametes, some females produce unreduced gametes;  – females produce haploid eggs, males are sterile;
– females produce haploid eggs, males are sterile;  – females produce clonal eggs, males are sterile;
– females produce clonal eggs, males are sterile;  – both sexes are sterile;
– both sexes are sterile;  – both sexes are inviable. Asterisk (*) indicates cross combinations for which diplotene analysis is missing. Each abbreviation represent a subgenome of the parental species: Cobitis elongatoides = EE; C. taenia = TT; C. tanaitica = NN; C. taurica = CC; C. pontica = PP; C. strumicae = SS; C. bilineata = BB; C. ohridana = OO; C. hankugensis = HH; C. biwae = BW; C. lutheri = Lut; C. tetralineata = Tet; Iksookimia longicorpa = LL; M. anguillicaudatus clade A=MaA; M. anguillicaudatus clade B=MaB; Lefua echigonia = Lef. Hypothetical scheme of the selective processes necessary for the emergence of clonality in F1 hybrids (b). First step includes the accumulation of mutations in gametogenic regulatory genes which is required to induce premeiotic genome endoreplication in F1 hybrid females. Second step includes the chromosomal divergence which leads to the aberrant chromosomal pairing followed by the cell arrest of meiocytes with non-duplicated genomes. Premeiotic genome endoreplication is sex specific, while hybrid males are sterile.
– both sexes are inviable. Asterisk (*) indicates cross combinations for which diplotene analysis is missing. Each abbreviation represent a subgenome of the parental species: Cobitis elongatoides = EE; C. taenia = TT; C. tanaitica = NN; C. taurica = CC; C. pontica = PP; C. strumicae = SS; C. bilineata = BB; C. ohridana = OO; C. hankugensis = HH; C. biwae = BW; C. lutheri = Lut; C. tetralineata = Tet; Iksookimia longicorpa = LL; M. anguillicaudatus clade A=MaA; M. anguillicaudatus clade B=MaB; Lefua echigonia = Lef. Hypothetical scheme of the selective processes necessary for the emergence of clonality in F1 hybrids (b). First step includes the accumulation of mutations in gametogenic regulatory genes which is required to induce premeiotic genome endoreplication in F1 hybrid females. Second step includes the chromosomal divergence which leads to the aberrant chromosomal pairing followed by the cell arrest of meiocytes with non-duplicated genomes. Premeiotic genome endoreplication is sex specific, while hybrid males are sterile.
Additional files
-
Supplementary file 1
This study uses a summary of karyotypes and RAG1 sequence of loaches (Cobitidae).
The references are indicated for species karyotypes that were previously described. The Karyotype of C. ohridana is published for the first time in this work.
- https://cdn.elifesciences.org/articles/88366/elife-88366-supp1-v1.xlsx
-
Supplementary file 2
The numbers of analyzed male and female Cobitis samples from each F1 obtained family.
Most specimens were used for the pachytene analysis in order to increase the chance to observe duplicated cells in females, as their incidence per slide is very low in clonal hybrids. Also, we used as many individuals as possible for this approach to have a statistically significant number of SC per individual.
- https://cdn.elifesciences.org/articles/88366/elife-88366-supp2-v1.xlsx
-
Supplementary file 3
Dataset with every SC reported from all analyzed specimens.
All this information was used in order to generate Figure 5.
- https://cdn.elifesciences.org/articles/88366/elife-88366-supp3-v1.xlsx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/88366/elife-88366-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Genetic and karyotype divergence between parents affect clonality and sterility in hybrids
eLife 12:RP88366.
https://doi.org/10.7554/eLife.88366.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}