E3 ubiquitin ligase Deltex facilitates the expansion of Wingless gradient and antagonizes Wingless signaling through a conserved mechanism of transcriptional effector Armadillo/β-catenin degradation
- Department of Molecular and Human Genetics, Institute of Science, Banaras Hindu University, India
- Department of Integrative Biology and Physiology, University of California Los Angeles, United States
- Department of Cell Biology, NYU Langone Medical Center, United States
Figures
Figure 1
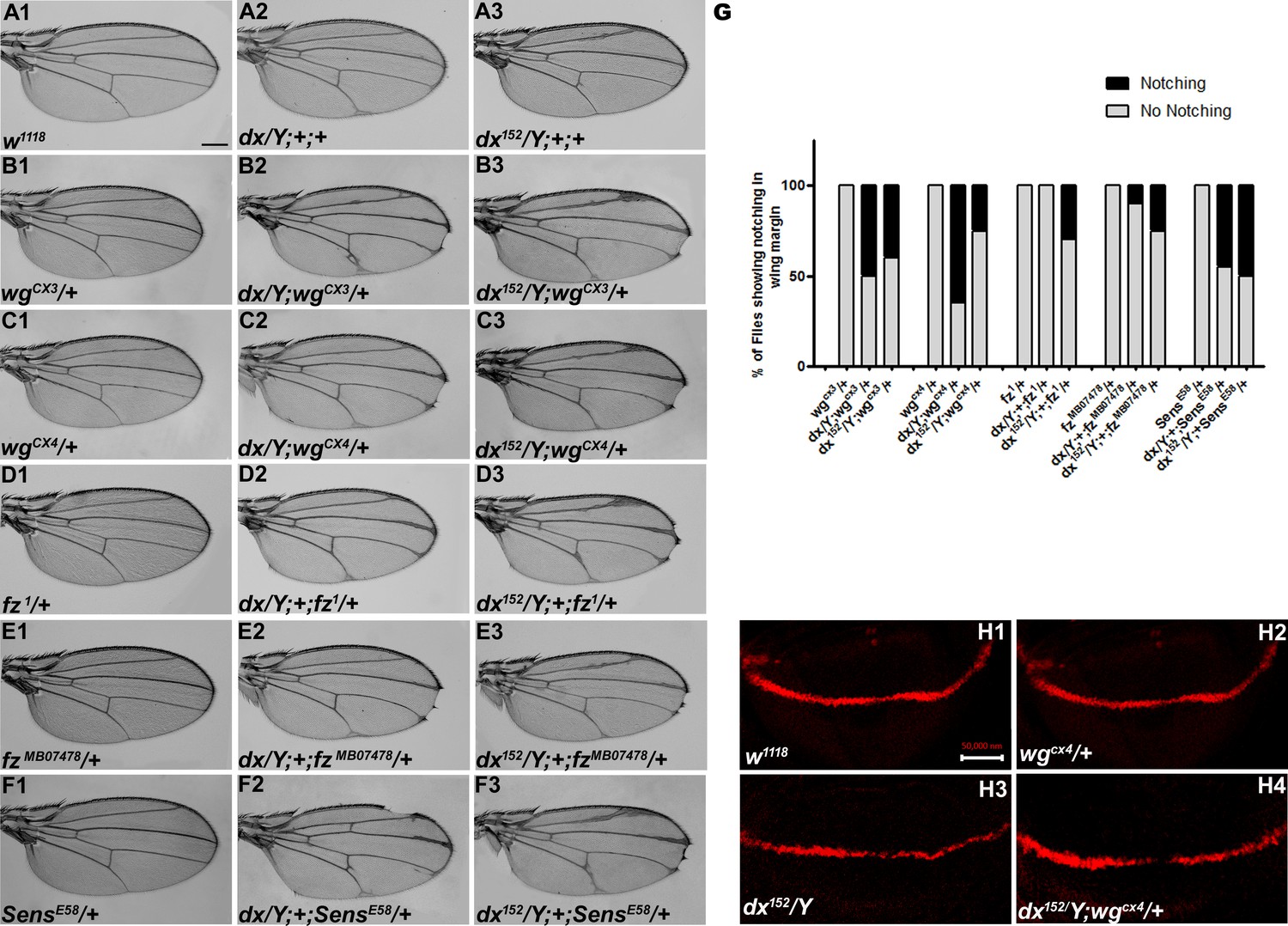
dx genetically interacts with Wg pathway components.
(A1–F3) Representative wings from males with the indicated genotypes. dx (A2) and dx152 (A3) hemizygote show extra vein material at the distal end of the wing compared to wild-type flies (w1118) (A1). (B2, B3, C2, C3) Both the dx allele show an enhancement of wing vein thickening along with wing notching in hemizygous combination with different alleles of wg (wgCX3 and wgCX4) heterozygotes. (D2, D3, E2, E3) Different alleles of fz (fz1 and fzMB07478) in trans-heterozygous conditions show enhanced vein thickening and wing nicking phenotype with dx hemizygotes. (F2, F3) dx alleles show strong genetic interaction with the Wg target gene sens, where a loss-of-function allele of sens (sensE58) shows wing nicking phenotype in flies that are homozygous for dx alleles. (G) Graph showing the frequency of wing notching phenotypes observed in indicated genetic combinations (n = 100). (H1–H4) Representative image of Cut in wing imaginal disc in different genetic combinations. (H2) The expression of Cut in wgCX4 heterozygote is similar to the wild-type Cut expression (H1). (H3) dx152 hemizygotes show a reduction in Cut expression in the dorsal–ventral (D/V) boundary of the wing disc. (H4) Wing disc from dx152/Y; wgCX4/+ genotype shows a further reduction in Cut expression when compared to dx152/Y wing discs. Images in H1–H4 are representatives of three independent experiments (n = 6). Scale bar: A1–F3: 200 µm. H1–H4: 50 µm.
Figure 2
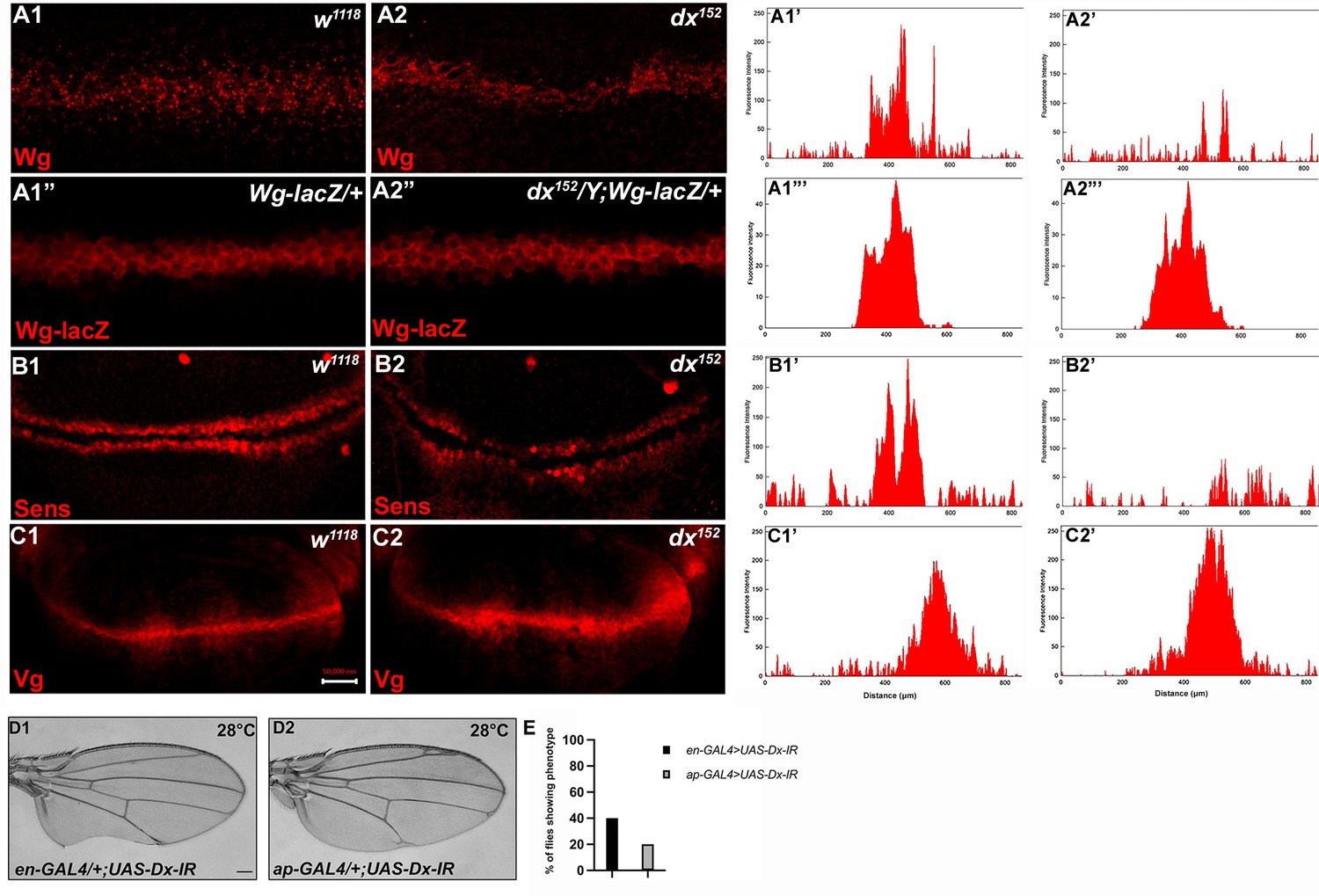
Loss of Dx reduces Wg signaling gradient and target gene expression.
(A2) dx152 wing disc shows a narrow Wg expression gradient compared to wild-type third instar larval wing discs (A1). (A1”, A2”) show Wg-lacZ staining in the mentioned genotype. (B2) A constricted expression of Sens was observed in dx null discs (dx152) (marked by arrowhead) compared to the control wild-type wing imaginal discs (B1). (C1) The expression of Vg in the wild-type disc. (C2) dx null discs showed a broadened Vg expression, marked by arrowheads. (A1’–C2’) Show the average fluorescence intensity of images in A1–C2. (D1, D2) Expression of the RNAi targeting Dx by en-GAL4 or ap-GAL4 results in morphological aberrations in the expressing regions. (E) Graph showing the percentage of flies showing respective phenotypes in the mentioned genotype (n = 100). Images in A–C are representatives of three independent experiments (n = 6). Scale bar: A1–A2: 10 µm. B1–C2: 50 µm. D1, D2: 200 µm.
Figure 3 with 2 supplements
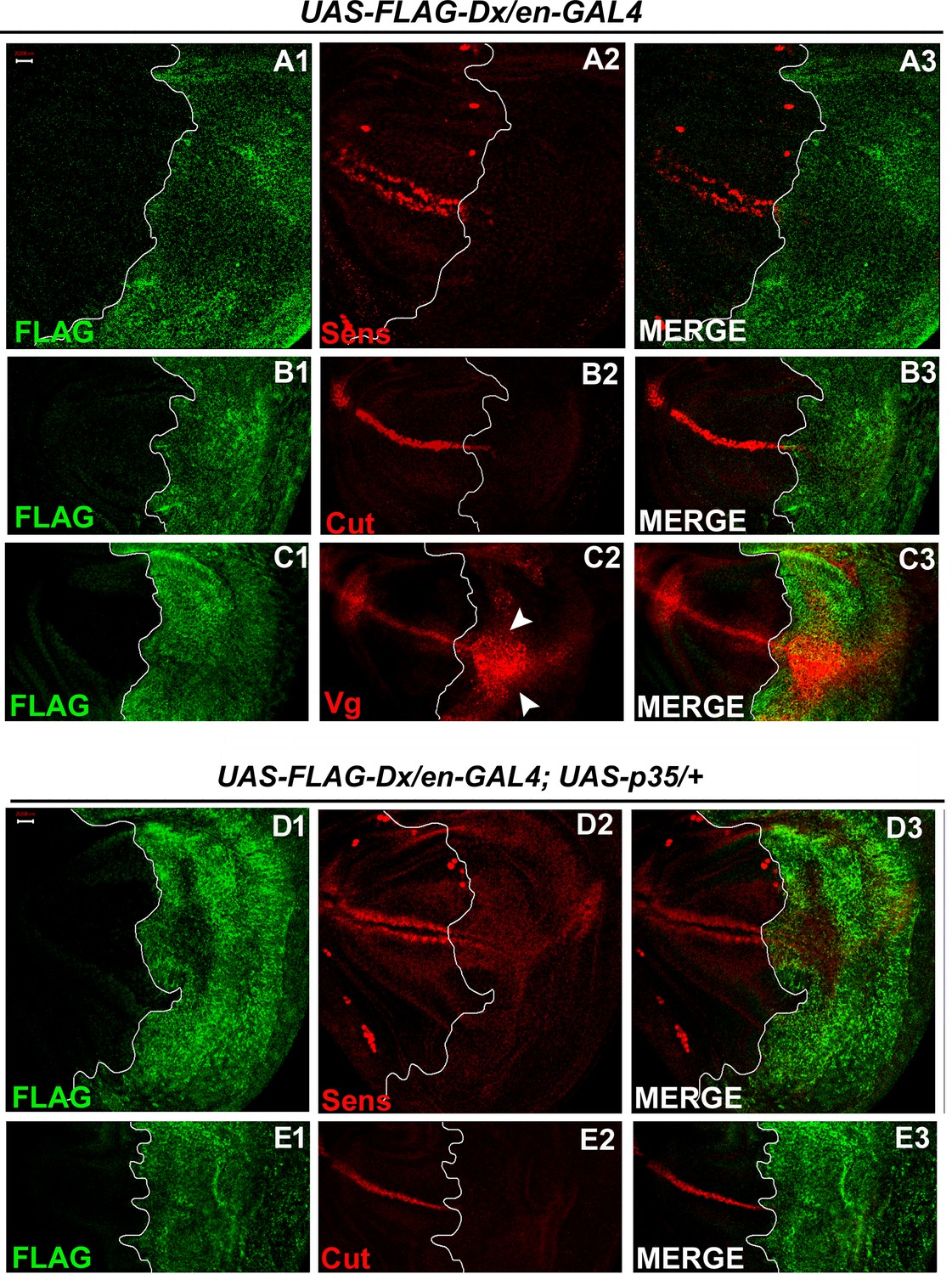
With two supplements: Dx modulates the expression of the Wg target genes.
(A1–A3) Over-expression of Dx using posterior domain-specific en-GAL4 results in a complete loss of Senseless (A2). Third instar larval wing discs of the indicated genotypes were immunostained to detect FLAG (green) and Senseless (red). (B1–B3) A reduction in expression of Cut was observed in the posterior domain of the wing disc upon Dx over-expression with en-GAL4. (C1–C3) Expression of Vestigial (Vg), on the contrary, was found to be expanded in the posterior compartment. Arrowheads indicate the expansion of Vg expression close to the anterior–posterior (A/P) boundary of the wing imaginal disc (C2). (D1–E3) Over-expression of caspase inhibitor p35 together with Dx does not show any rescue in the expression of Senseless (D2) and Cut (E2) suggesting the event is independent of apoptosis. The white line marks the boundary of FLAG expression. Images in A–E are representatives of three independent experiments (n = 6). Scale bar: A1–E3: 50 µm.
Figure 3—figure supplement 1
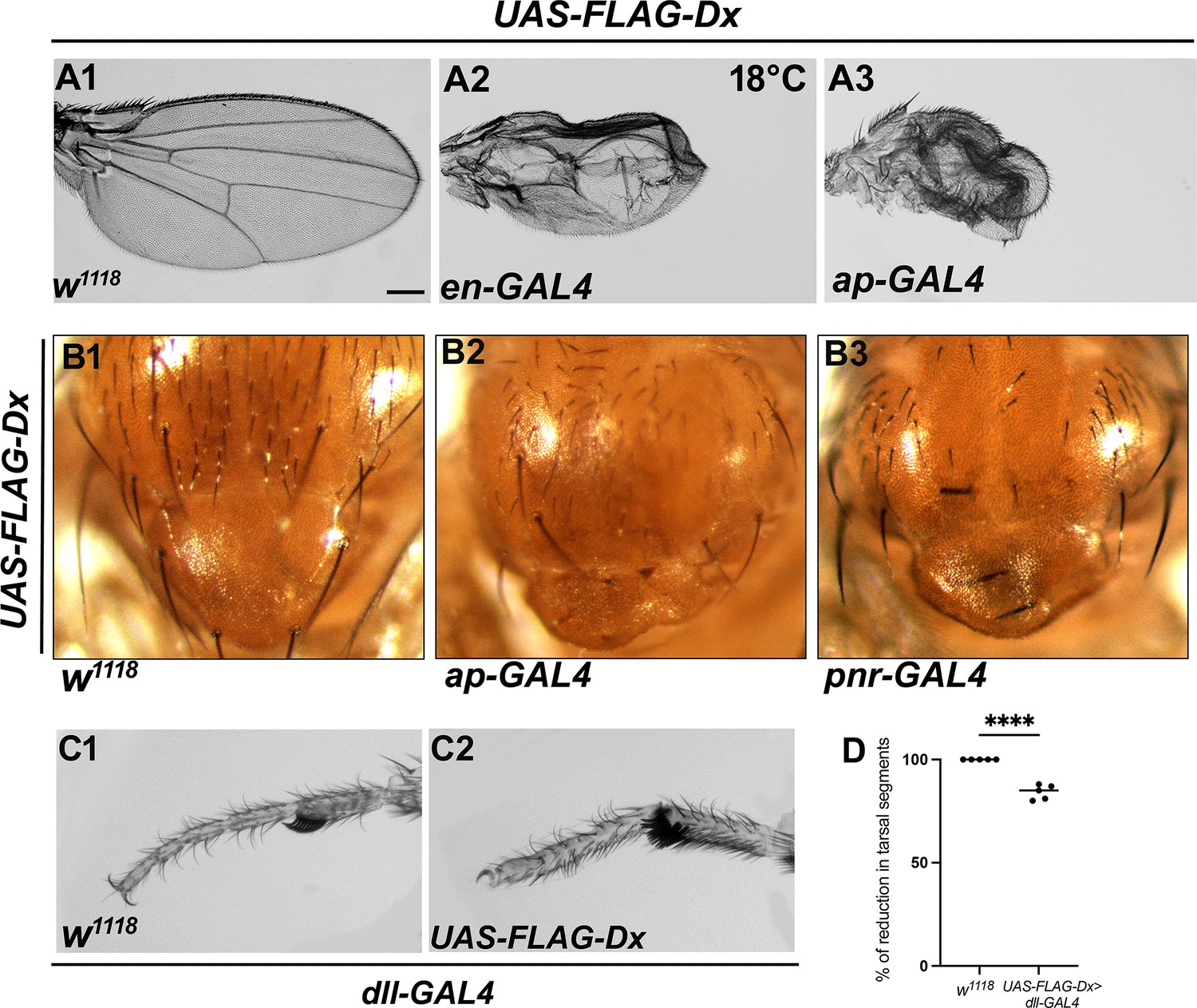
Ectopic Dx mimics Wg loss-of-function phenotype.
(A1) Representative image of wild-type adult wing. Over-expression of Deltex (Dx) in the Drosophila wing by en-GAL4 (A2) or ap-GAL4 (A3) shows a smaller wing with severe morphological defects. (B1) A wild-type notum. (B2) Dx expression with ap-GAL4 shows the loss of macrochaetae on the notum. Loss of scutellar bristles was also noted. (B3) Dx expression with pnr-GAL4 shows similar defects in the notum region with a marked reduction in the macrochaetae and scutellar bristles. (C2) A reduced tarsal segment and dorsal–ventral shift of the sex combs were observed upon Dx over-expression with distal-less GAL4. (D) Graph in D represents the percentage of reduction in tarsal segments in Dx over-expressed flies with dll-GAL4 compared to wild-type flies. ****p < 0.0001; unpaired t-test. Scale bar: A1–C2: 200 µm.
Figure 3—figure supplement 2
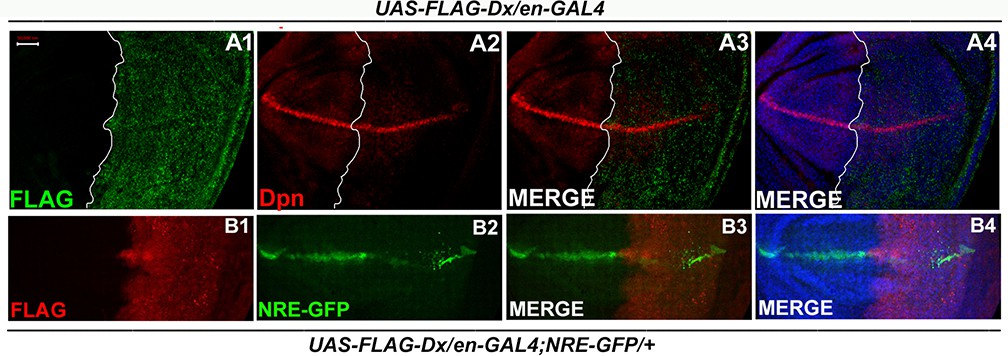
Dx-mediated Wg regulation is independent of Notch.
(A1–A4) Notch target Deadpan (Dpn) remains unaltered upon Dx over-expression. Over-expression of Dx by posterior domain-specific en-GAL4 does not cause any appreciable change in the expression of Notch target Deadpan. (B1–B4) en-GAL4 driven expression of UAS-FLAG-Dx results in a punctate Notch response element (NRE) expression, marked by GFP. Images in A and B are representatives of three independent experiments (n = 6).Scale bar: A1–B4: 50 µm.
Figure 4
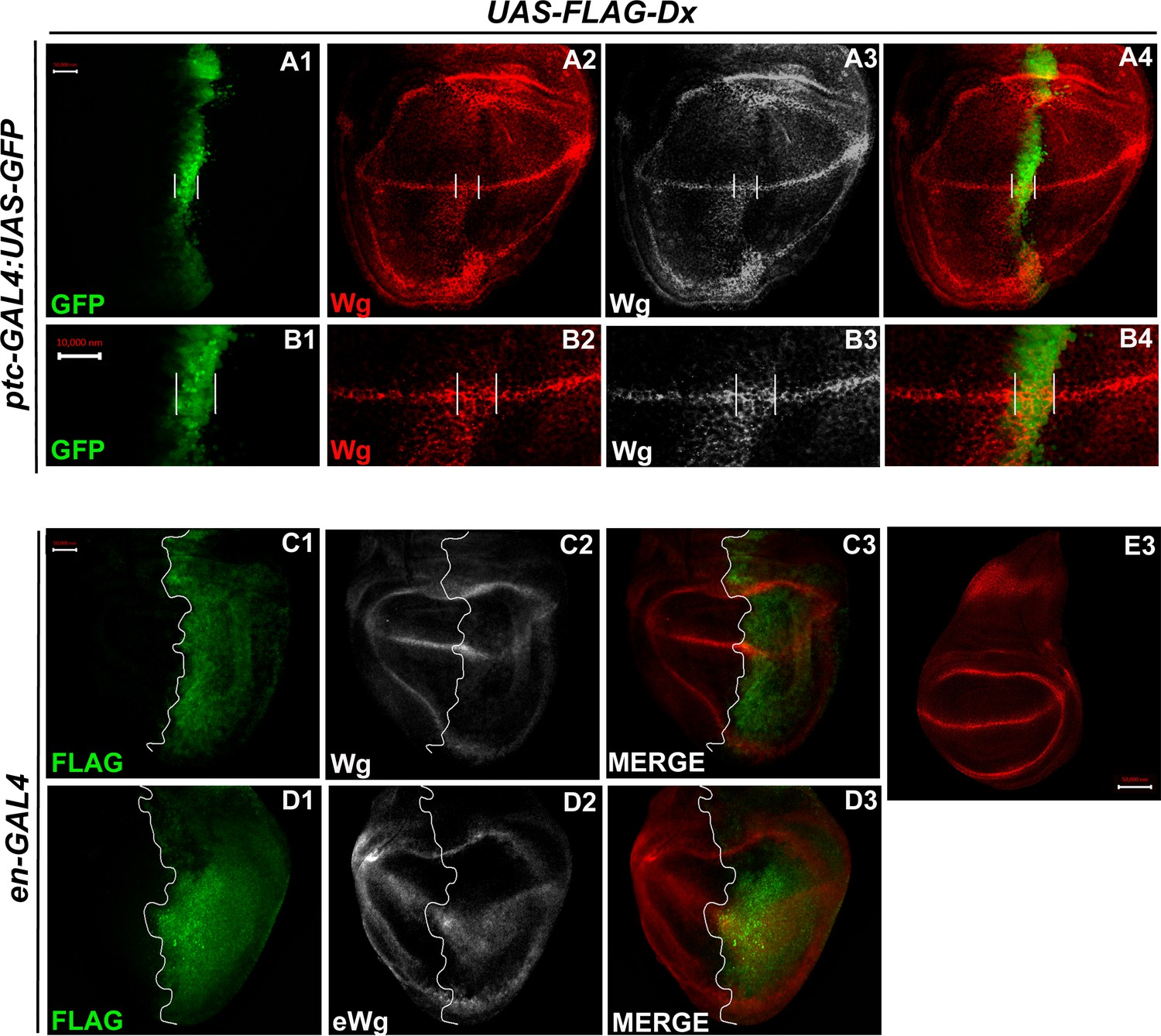
Over-expression of Dx expands the Wg diffusion gradient.
(A1–B4) Over-expression of Dx by anterior–posterior (A/P) domain-specific ptc-GAL4 results in the broadening of the Wg diffusion domain (A2). Grayscale images are for better contrast (A3). (B2) High-magnification picture shows increased Wg puncta at the A/P–dorsal–ventral (D/V) junction with more significant expression in the ventral portion of the disc (marked by arrowhead) (B2, B3). The white line marks the Ptc domain, respectively. (C1–C3) Dx over-expression with en-GAL4 results in a diffused Wg expression in the posterior domain of the wing disc. (D1–D3) A marked increase in extracellular Wg (eWg) was observed upon Dx over-expression with en-GAL4. (E3) Representative image of wild-type Wg expression. Images in A–D are representatives of three independent experiments (n = 6). Scale bar: A1–A4, C1–D3, and E: 50 µm. B1–B4: 10 µm.
Figure 5
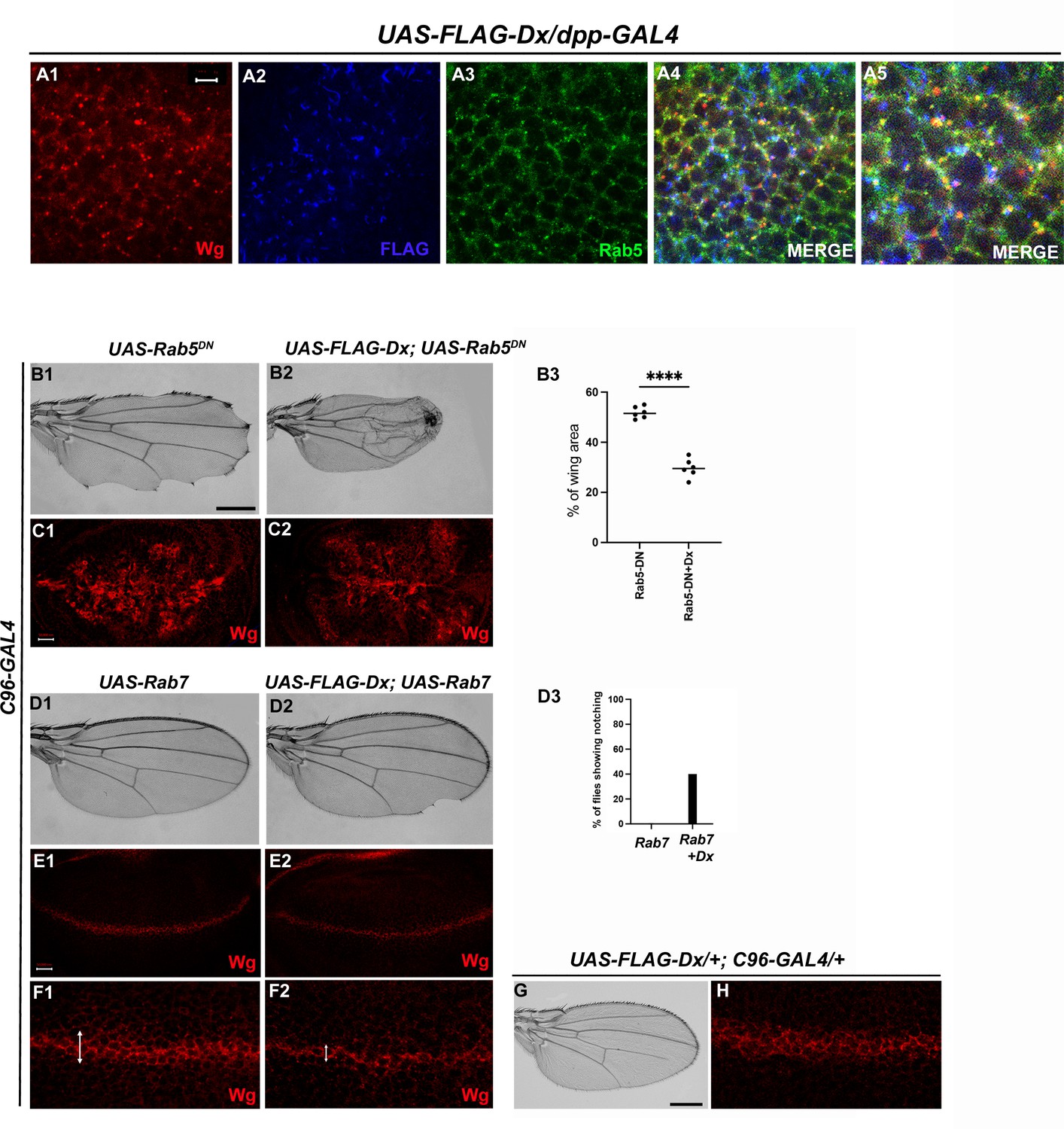
Dx-mediated Wg signaling is regulated by endocytosis.
(A1–A5) Dx (blue) co-localizes with Rab5 (green) and Wg (red) in the same subcellular compartment. The arrows mark the co-localized spots. (A5) High-magnification image of (A4). (C1) Over-expression of a Dominant Negative Rab5 with C96-Gal4 inhibits the endosomal fusion at the dorsal–ventral (D/V) boundary, enhancing the Wg diffusion gradient. (C2) Co-expression of Dx with Rab5DN shows a further enhancement of the Rab5DN phenotype in the Wg diffusion gradient. (B1, B2) Representative adult wing image of the mentioned genotype (n = 50). (B1) Over-expression of Rab5DN results in wing tissue loss consistent with decreased Wg signaling. (B2) A further reduction in wing size is observed upon co-expressing Dx with Rab5DN. (B3) The graph shows the wing area percentage of the mentioned genotypes (n = 6). ****p < 0.0001; unpaired t-test. (E1, F1) Over-expression of Rab7 with C96-GAL4 renders no significant change in Wg expression. (E2, F2) Rab7 when co-expressed with Dx enhances the Wg spreading. (D1, D2) Representative adult wing of defined genotype (n = 50). Note the notching phenotype in the wing upon over-expression of FLAG-Dx together with UAS-Rab7 (D2). (D3) The graph shows the percentage of flies showing notching phenotype (n = 100). (G, H) Representative adult wing image of UAS-FLAG-Dx over-expressed with C96-GAL4. (H) Wg expression in UAS-FLAG-Dx>C96 GAL4 discs. Images in C and E are representatives of three independent experiments (n = 6). Scale bar: A1–A4: 5 µm. C1, C2 and E1, E2: 50 µm. F1, F2: 20 µm. B1, B2 and D1, D2: 200 µm.
Figure 6 with 2 supplements
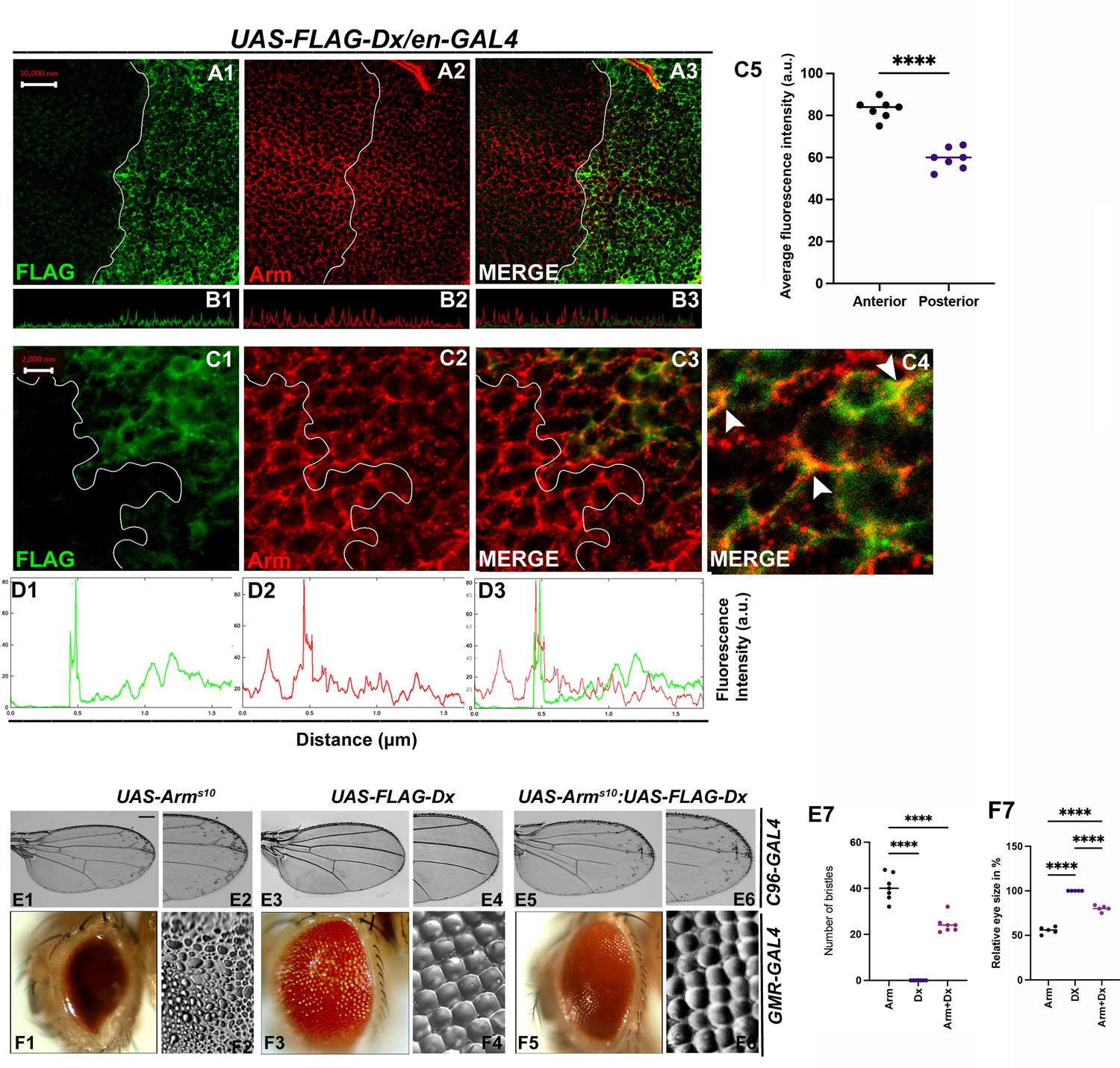
With 2 supplements: Dx modulates Wg Signal transduction.
(A1–C4) Dx over-expression with en-GAL4 in the wing imaginal disc results in a reduction of endogenous Armadillo throughout the disc with a more pronounced reduction at the dorsal–ventral (D/V) boundary. (B1–B3) Confocal Z stack Intensity profiling shows a reduced Arm level in the posterior compartment of the disc. (C1–C4) A higher magnification picture shows a significant overlap between FLAG-tagged Dx and endogenous Arm (marked by arrowhead). Note the reduction of Arm in the posterior compartment of the disc. (C5) Graph shows the average Arm fluorescence intensity (a.u., arbitrary units) (n = 7). ****p < 0.0001; unpaired t-test. (D1–D3) Show the average FLAG and Arm fluorescence intensity profiles for the representative wing disc. (E1–E6) Representative adult wing of defined genotype (n = 50). (E1, E2) Over-expression of the activated form of Arm (ArmS10) with C96-GAL4 shows ectopic hairs more significantly along the wing margin of the adult wing. (E3, E4) Dx over-expression in the wing shows no ectopic hairs. (E5, E6) Co-expression of Dx with ArmS10 results in the suppression of Arm gain-of-function effect and a relative reduction in the number of ectopic bristles was observed. (E7) Graph shows the number of bristles in each combination. ****p < 0.0001; one-way analysis of variance (ANOVA)/Turkey’s multiple comparisons test. (F1–F6) Representative adult eye and eye imprint of defined genotype (n = 10). (F1, F2) GMR-specific expression of an activated form of Arm produces a severely reduced eye with diffused ommatidia. (F3, F4) Dx over-expression renders no significant phenotype. (F5, F6) Arm over-expression phenotype gets dramatically suppressed when Dx was co-expressed in the same background. (F7) Graph represents the phenotypic score of the eye size (n = 6). ****p ,0.0001; one-way analysis of variance (ANOVA)/Turkey’s multiple comparisontest. Images in A–C are representatives of three independent experiments (n = 6). Scale bar: A1–A3: 10 µm. C1–C3: 2 µm. E1–E6 and F1–F6: 200 µm.
Figure 6—figure supplement 1
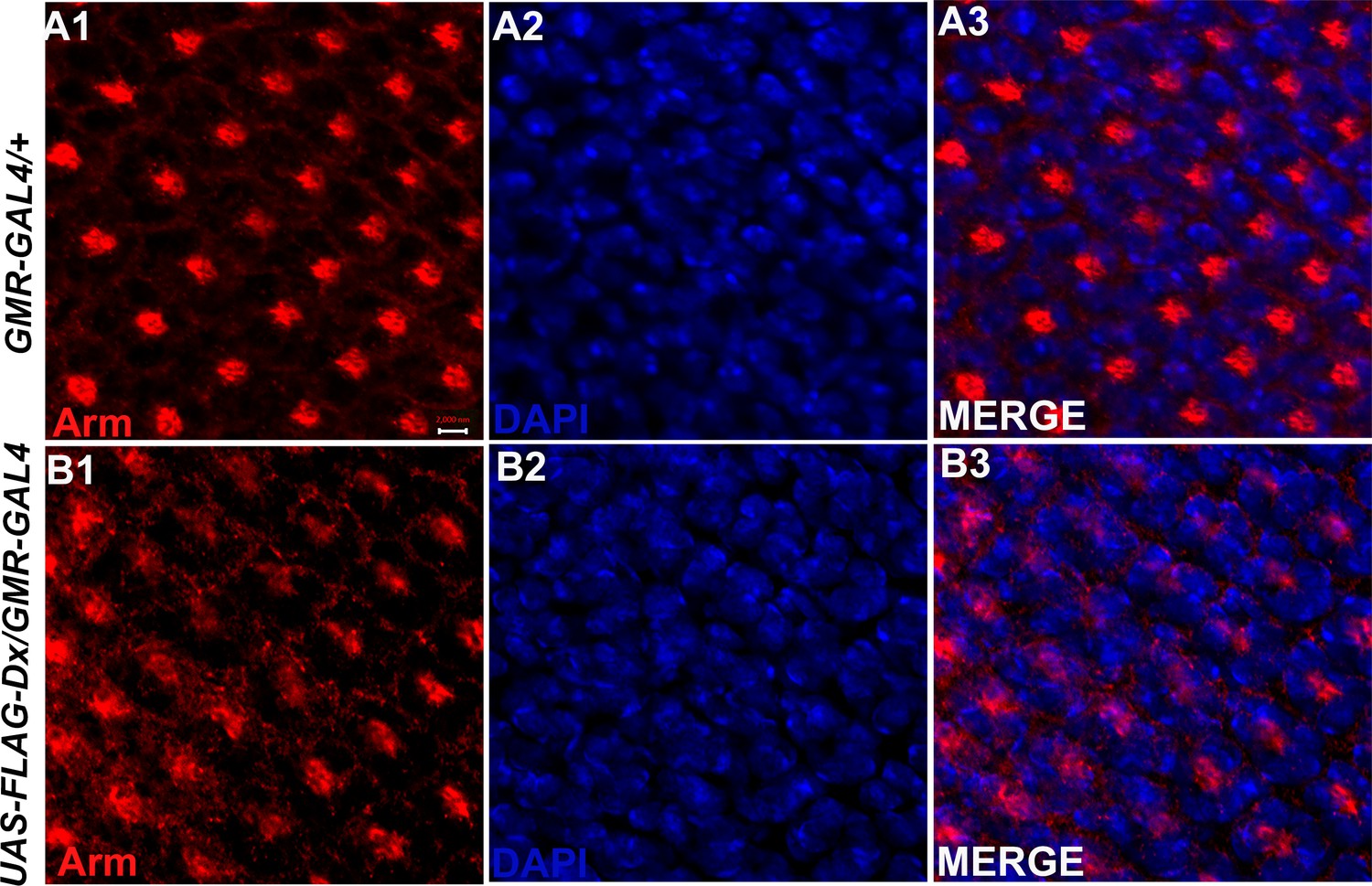
Dx modulates Arm expression in the eye tissue.
A diffused expression of Arm was observed in the eye disc upon Dx over-expression in the GMR domain (B1-B3) in contrast to a wild-type Arm expression (A1-A3). Images in A and B are representatives of three independent experiments (n = 6). Scale bar: A1–B3: 2 µm.
Figure 6—figure supplement 2
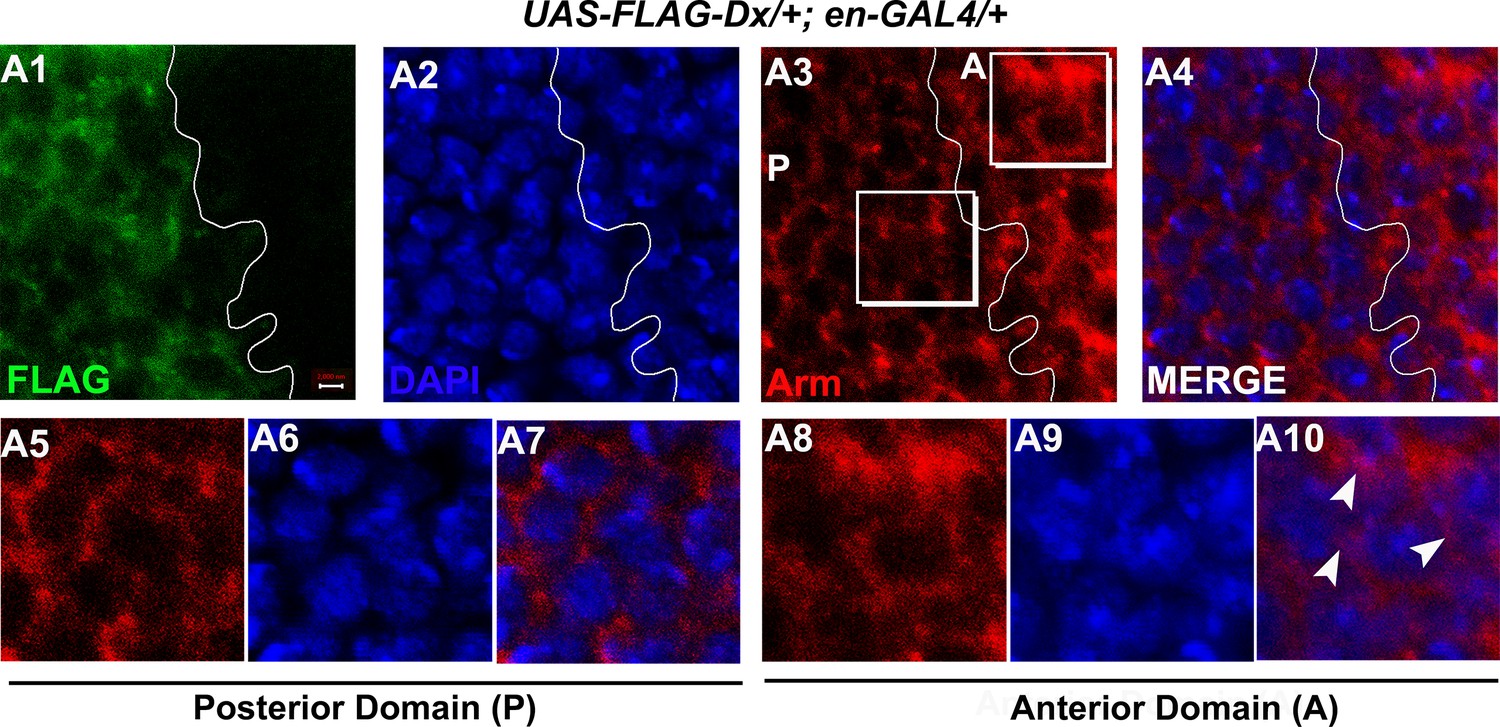
Dx might facilitate the cytoplasmic retention of Arm.
(A1–A10) High-magnification picture of Arm in the anterior (A) and posterior (P) domain of the wing imaginal disc. Note (A8–A10) showed a significant co-localization of Arm with nucleus stained with DAPI [4’,6-diamidino-2-phenylindole (blue)], (A5–A7) however showed no such co-localization, suggesting that Dx might facilitate the nuclear retention of Arm. Images are representatives of three independent experiments (n = 6). Scale bar: A1–A4: 2 µm.
Figure 7
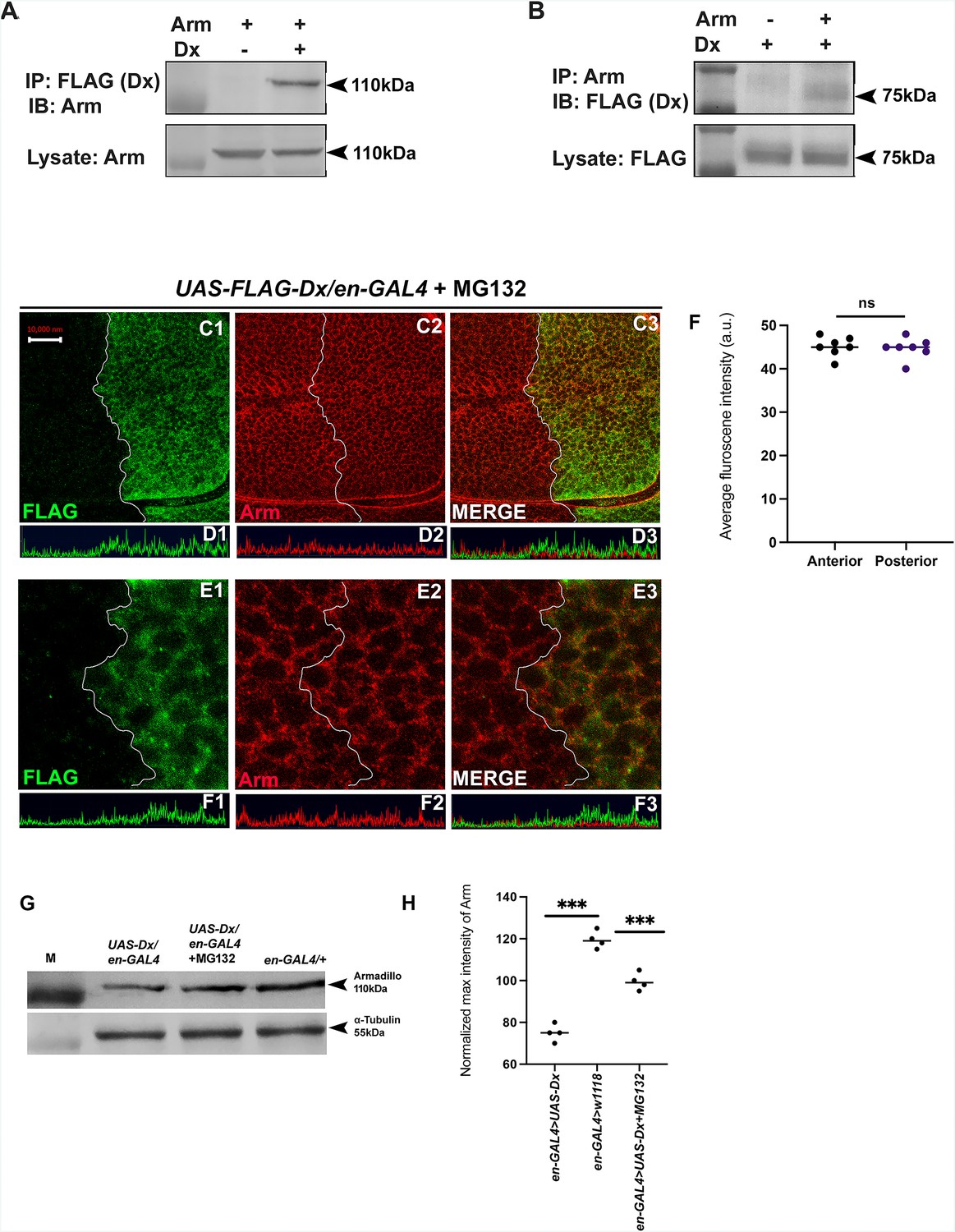
Dx degrades Arm by a proteasome-mediated mechanism.
(A, B) Co-immunoprecipitation of FLAG-Dx and Arm. Co-immunoprecipitation was carried out with lysate over-expressing Arm and FLAG-Dx using GMAR-GAL4. + symbol indicates the presence of lysate and the − symbol shows the absence of lysate. FLAG-Dx immunoprecipitated Arm was detected by anti-Arm antibody (A). Arm immunoprecipitated FLAG-Dx was detected by FLAG antibody. (C1–C3) Dx over-expressed discs treated with MG132 for 3 hr showed a rescue in Arm expression and a more intense Arm staining in the posterior compartment was found after proteasome inhibitor treatment. (D1–D3) Confocal Z stack Intensity profiling shows a comparable Arm level in the posterior compartment of the wing imaginal disc. (E1–E3) A higher magnification picture shows no significant change in the expression of Arm in the posterior compartment of the disc suggesting the rescue in Arm degradation. (F) Graph shows the average Arm fluorescence intensity (a.u., arbitrary units) (n = 7). ns; unpaired t-test. (G) Western blot analysis confirms the immunocytochemical studies where proteasome inhibitor MG132 increases Arm protein levels in Dx over-expressed tissue samples. (H) Graph G represents the intensity profiling of the Western blot. ***p < 0.001; unpaired t-test. Images in A–F are representatives of three independent experiments (n = 6). Scale bar: A1–A4: 10 µm. C1–C4: 2 µm.
-
Figure 7—source data 1
Original file for the co-immunoprecipitation analysis of Figure 7A (anti-Armadillo).
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig7-data1-v1.pptx
-
Figure 7—source data 2
Original scans of the relevant co-immunoprecipitation analysis (anti-Armadillo) with highlighted bands and sample labels for Figure 7A.
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig7-data2-v1.pptx
-
Figure 7—source data 3
Original file for the co-immunoprecipitation analysis of Figure 7B (anti-FLAG).
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig7-data3-v1.pptx
-
Figure 7—source data 4
Original scans of the relevant co-immunoprecipitation analysis anti-FLAG with highlighted bands and sample labels for Figure 7B.
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig7-data4-v1.pptx
-
Figure 7—source data 5
Original file for the Western blot analysis in Figure 7G (anti-Armadillo and anti-tubulin).
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig7-data5-v1.pptx
-
Figure 7—source data 6
Original scans of the relevant Western blot analysis (anti-Armadillo and anti-tubulin) with highlighted bands and sample labels for Figure 7G.
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig7-data6-v1.pptx
Figure 8
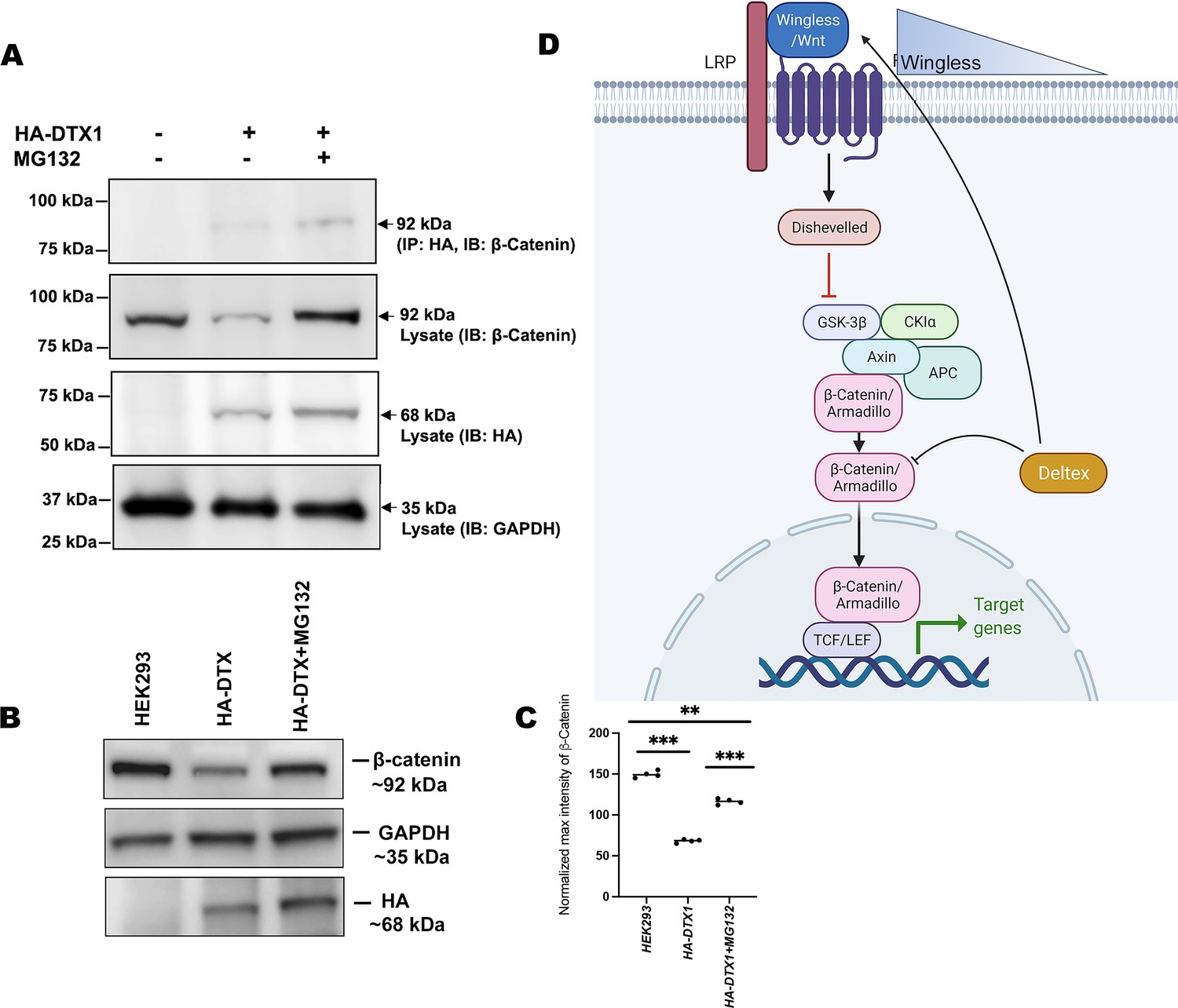
DTX1 physically interacts with β-catenin and facilitates its degradation.
(A) HA-tagged human DTX1 transfected in HEK-293 cell line was pulled down with an anti-HA antibody. Immunoblot was done with β-catenin. A prominent band at 92 kDa corresponding to β-catenin post-MG132 treatment was observed. GAPDH was used as an internal control. (B) A rescue in β-catenin level post-MG132 treatment was observed. GAPDH serves as an internal control. (C) Graph C represents the intensity profiling of the Western blot in C. **p < 0.01, ***p < 0.001; one-way analysis of variance (ANOVA)/Turkey’s multiple comparisons test. Images in A–F are representatives of three independent experiments. (D) The cartoon explains the two plausible mechanisms of Wg regulation through Dx. Dx facilitates Wg gradient formation on the one hand and on the other it targets Arm for its degradation thereby regulating the signaling output.
© 2024, BioRender Inc. Figure 8D was created using BioRender, and is published under a CC BY-NC-ND license. Further reproductions must adhere to the terms of this license.
-
Figure 8—source data 1
Original file for the co-immunoprecipitation analysis in Figure 8A (anti-β catenin, anti-HA, and anti-GAPDH).
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig8-data1-v1.pptx
-
Figure 8—source data 2
Original scans of the relevant co-immunoprecipitation analysis (anti-β catenin, anti-HA, and anti-GAPDH) with highlighted bands and sample labels for Figure 8A.
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig8-data2-v1.pptx
-
Figure 8—source data 3
Original file for the Western blot analysis in Figure 8B (anti-β catenin, anti-HA, and anti-GAPDH).
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig8-data3-v1.pptx
-
Figure 8—source data 4
Original scans of the relevant Western blot analysis (anti-β catenin, anti-HA, and anti-GAPDH) with highlighted bands and sample labels for Figure 8B.
- https://cdn.elifesciences.org/articles/88466/elife-88466-fig8-data4-v1.pptx
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
E3 ubiquitin ligase Deltex facilitates the expansion of Wingless gradient and antagonizes Wingless signaling through a conserved mechanism of transcriptional effector Armadillo/β-catenin degradation
eLife 12:RP88466.
https://doi.org/10.7554/eLife.88466.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}