Pharyngeal mechanosensory neurons control food swallow in Drosophila melanogaster
- School of Life Sciences, IDG/McGovern Institute for Brain Research, Tsinghua University, China
- Tsinghua-Peking Center for Life Science, China
Figures
Figure 1 with 1 supplement
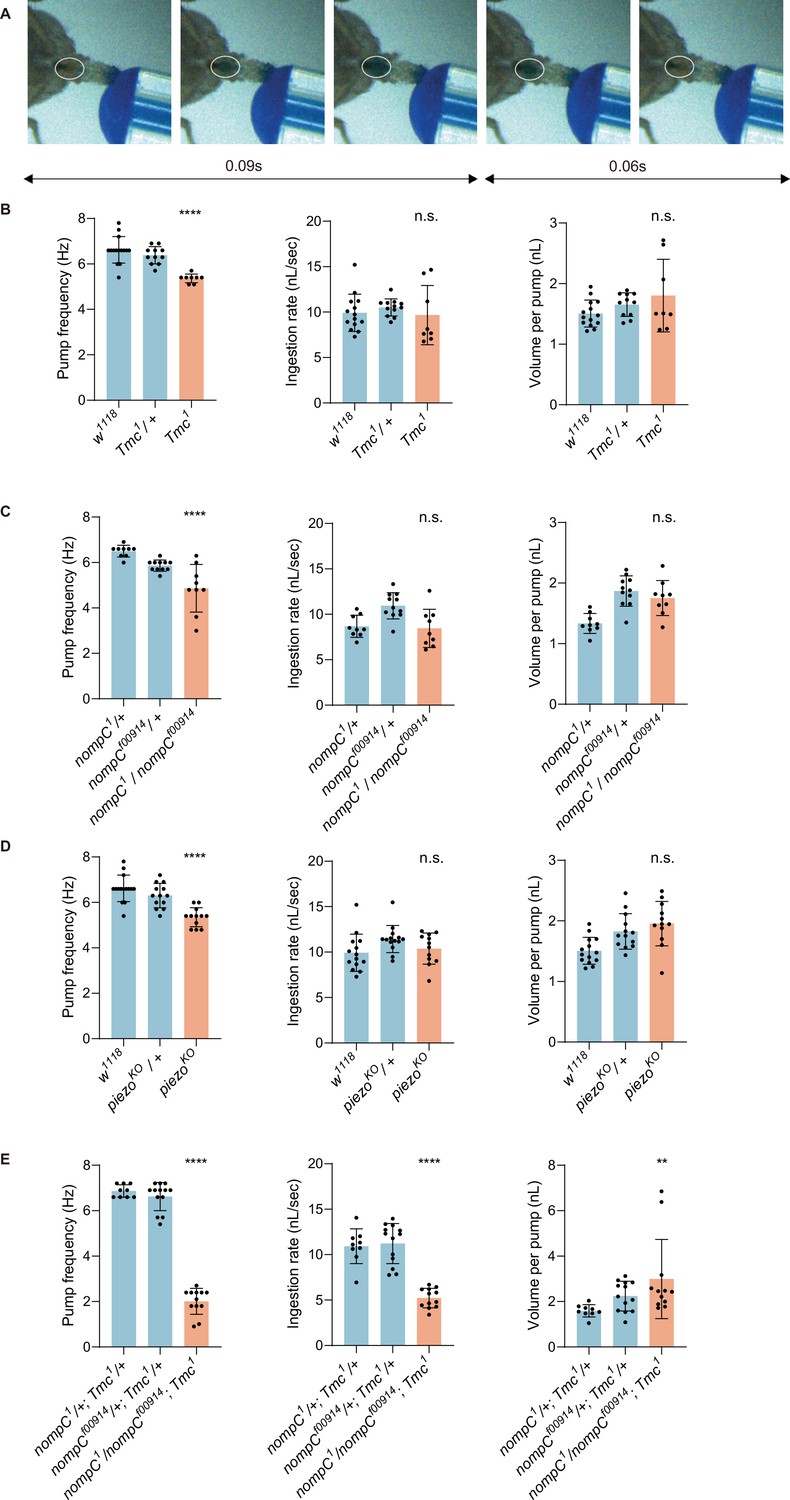
Mechanoreceptor genes are essential for swallow control of Drosophila.
(A) Swallow patterns of liquid food. Filling and emptying process constitutes a cycle. (B–D) Swallowing behavior of Tmc, nompC, and piezo mutant flies. Pump frequency represents the swallowing speed, ingestion rate indicates the efficiency of food intake, while volume per pump shows how much a fly ingests with one pump. N=8–15 in each group. (E) Double mutans of Tmc and nompC show more severe dysphagia. n=9–13 in each group. In all analyses, one-way analysis of variance (ANOVA) followed by Dunnett’s test for multiple comparisons was used, and statistical differences were represented as follows: *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Data were represented as means ± SEM.
Figure 1—figure supplement 1

Mutation of mechanoreceptors leads to a lower pump frequency caused by longer emptying time.
(A–B) Filling time and emptying of the cibarium in flies of different genotypes. n=6–14. (C) Ratio of filling time dividing emptying time indicates the proportion of two processes in one cycle. n=6–14. (D) Tmc and piezo double mutation showed a similar phenotype with single mutation for either gene in swallowing behavior. n=9–24. (E–H) Proportion of time with incomplete emptying during swallowing when flies were fed with methylcellulose solution. n=4–11. Data were represented as means. In all analyses, one-way analysis of variance (ANOVA) followed by Dunnett’s test for multiple comparisons except nonparametric test in (D) was used, and statistical differences were represented as follows: *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Data were represented as means ± SEM.
Figure 2 with 1 supplement
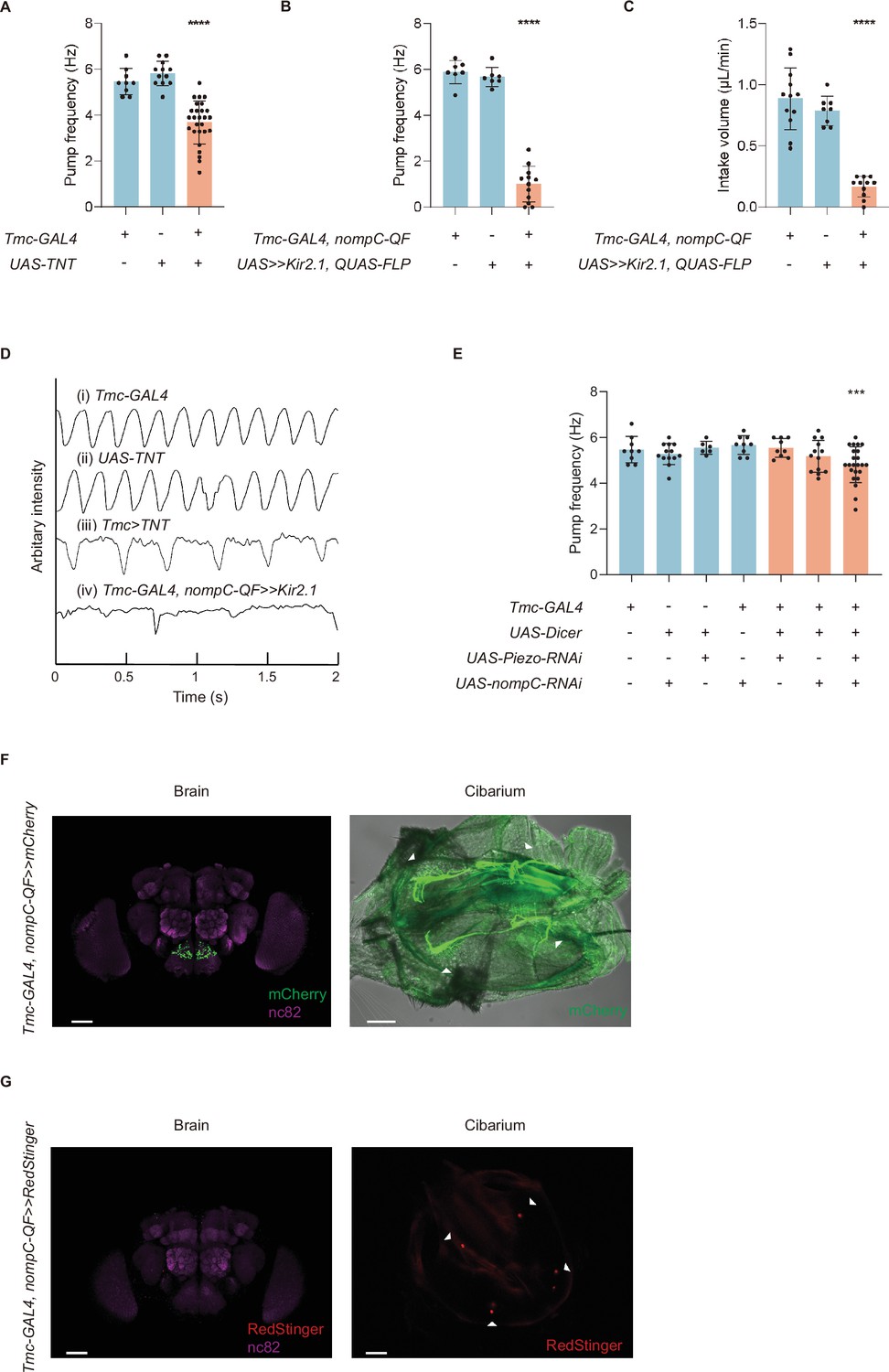
Mechanosensory neurons and mechanoreceptors are essential to swallow.
(A) Blocking synaptic transduction of Tmc positive neurons leads to a lower pump frequency, and the flies display difficulty in cibarium emptying (emptying time>0.3 s). n=9–27 in each group. (B–C) Inhibiting Tmc-GAL4 and nompC-QF double-labeled neurons by Kir2.1 results in a lower pump frequency and intake volume per minute. n=7–12 in each group. (D) Arbitrary intensity of cibarium when flies of different genotype swallow. The ordinate value was graphed using the opposite value of the original arbitrary intensity in the cibarium, so the declining line represents emptying process (the same for Figure 3B). (E) Pump frequency after knocking down the expression level of the mechanoreceptor genes. n=6–23 in each group. (F) Projection pattern of md-C neurons in the brain and cibarium. About two pairs of multi-dendritic neurons are situated along the pharynx. Genotype: Tmc-GAL4; UAS-FRT-mCherry/nompC-QF; QUAS-FLP. Scale bar=50 μm. (G) Somata of md-C neurons situated in the cibarium but not in the cibarium. Genotype: XUAS-RedStinger; Tmc-GAL4, UAS-FRT-GAL80-STOP-FRT/ +; nompC-QF, QUAS-FLP/+. Scale bar=50 μm. In all analyses, one-way analysis of variance (ANOVA) followed by Dunnett’s test for multiple comparisons was used, and statistical differences were represented as follows: *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Data were represented as means ± SEM.
Figure 2—figure supplement 1

Expression pattern of (A) Tmc, (B) nompC, (C) piezo, (D) Tmc & piezo, and (E) nompC & piezo in the brain and cibarium.
Scale bar=50 μm.
Figure 3 with 1 supplement
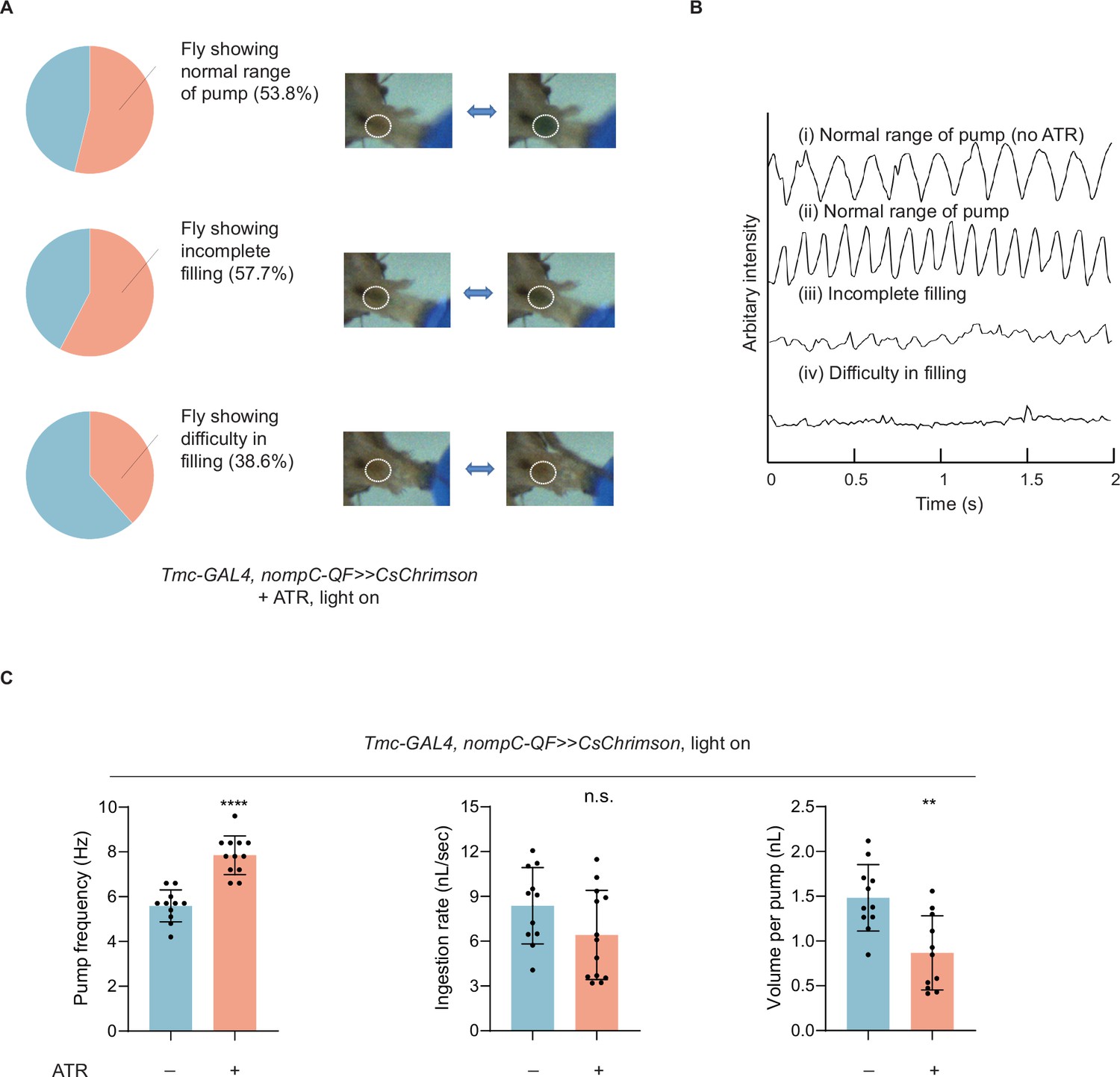
Activation of md-C neurons led to dysphagia.
(A) Optogenetic activation of md-C neurons with CsChrimson during food intake could induce accelerated swallowing, incomplete filling (the expansion range of cibarium decreased to about half the maximum), and difficulty in filling. Distinct states driven by CsChrimson light stimulation of md-C neurons were displayed, with the proportions of flies exhibiting each state. n=26. (B) Arbitrary intensity of fly cibarium when md-C neurons were stimulated as (A) or not optogenetically stimulated (no ATR). (C) Swallowing behavior of flies with or without md-C neurons stimulated. n=11–13 in each group. Pump frequency was calculated only when fly consistently pumped at a normal range (observed dye boundary in the cibarium is wider than 90% width of mouthpart). ATR, all trans retinal. In all analyses, two-tailed unpaired t-tests were used, and statistical differences were represented as follows: **p<0.01 and ****p<0.0001. Data were represented as means ± SEM.
Figure 3—figure supplement 1

md-C neurons, but not md-L neurons, are essential for sensorimotor control of swallow rhythm.
(A) In the brain, the projecting pattern of Tmc-GAL4 & nompC-QF>>GFP exhibited no significant changes post labellum ablation. Following the ablation of the labellum for 36 hr, the GFP signals of md-L in the mouth were scarcely observable when GFP expression was driven by Tmc-GAL4 & nompC-QF. (B) Flies expressing ReaChR in md-C neurons display dysphagia when stimulated by light. n=18. (C) Flies with labellum ablated for 36 hr still showed response to light stimulation of md-C, indicating that md-L neurons are not necessary for controlling swallow rhythm. n=11.
Figure 4 with 1 supplement
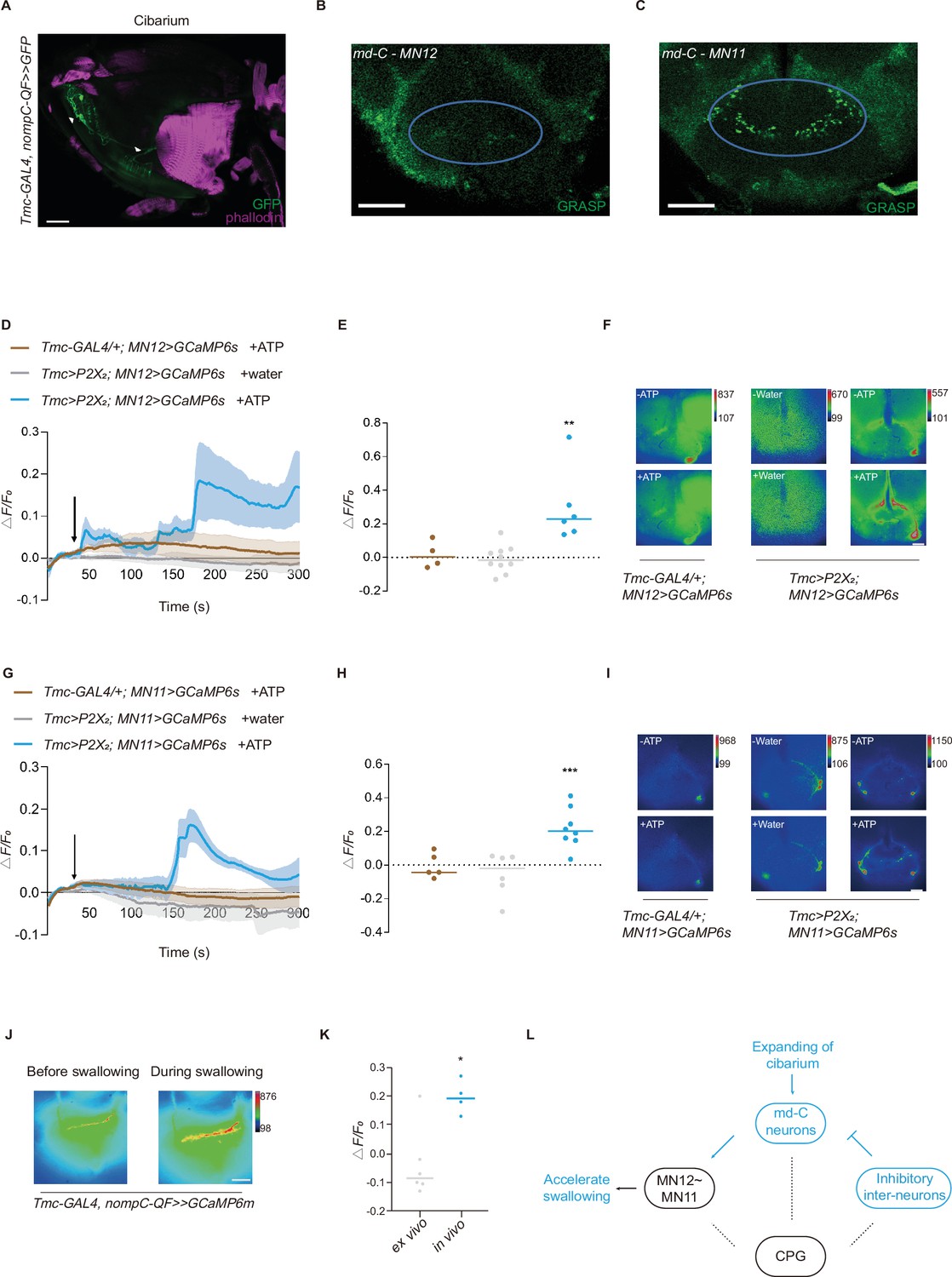
Interaction between motor neurons (MNs) and md-C neurons revealed working pattern of swallow.
(A) md-C neurons labeled by GFP antibody (green) and muscles labeled by phalloidin (magenta) showed that they are in close proximity around cibarium. Genotype: Tmc-GAL4/UAS-FRT-mCD8-GFP; nompC-QF/QUAS-FLP. Scale bar=50 μm. (B–C) GRASP (GFP reconstitution across synaptic partners) signals between md-C neurons and MNs could be observed in the subesophageal zone (SEZ) area. Genotype: Tmc-GAL4, UAS-FRT-GAL80-STOP-FRT/UAS-nSyb-spGFP1-10, lexAop-CD4-spGFP11; nompC-QF, QUAS-FLP/MN-LexA. Scale bar=50 μm. (D–F and G–I) Activation of Tmc+ neurons via P2X2 increased MN activity. Fluorescence changes (ΔF/F0) of GCaMP6s in MNs indicate calcium level changes. Water or ATP solution was added when it came to 30 s after record begins, as black arrow indicates. n=4–10, **p<0.01, ***p<0.001, one-way analysis of variance (ANOVA) followed by Dunnett’s test for multiple comparisons was used, error bars indicate mean ± SEM. Scale bar=50 μm. (J) Significant signal changes could be detected of the md-C neurons’ termini at fly’s SEZ area after feeding fly with 0.1 M sucrose solution when GCaMP6m is expressed in md-C neurons. Scale bar=50 μm. (K) are traces of fluorescence changes. n=4–6, *p<0.05, two-tailed unpaired t-test was used, error bars indicate mean ± SEM. (L) Working model for md-C-MN-CPG controlling swallow of Drosophila. CPG, central pattern generators.
Figure 4—figure supplement 1

Regulation of md-C neurons requires the inhibitory inter-neurons.
(A) Knocking down the expression of GABAA-R in md-C neurons caused a lower pump frequency. n=16–25, one-way analysis of variance (ANOVA) followed by Dunnett’s test for multiple comparisons was used in (B) and (C) while nonparametric test was used in (A), and statistical differences were represented as follows: *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. Data were represented as means ± SEM.
Videos
Video 1
Feeding behavior of wild-type flies and mechanoreceptor mutant flies.
Video 2
Tmc1 and piezoKO flies displayed incomplete emptying when fed with 1% methylcellulose (MC) water.
Video 3
Flies with md-C neurons inhibited displayed difficulty in cibarium emptying.
Video 4
Optogenetic activation of md-C neurons caused accelerated swallowing, incomplete cibarium filling, and difficulty in filling.
Video 5
Optogenetic activation of md-C neurons could still trigger dysphagia in flies without labellum.
Tables
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (D. melanogaster) | Tmc-Gal4 | Bloomington Drosophila Stock Center | BDSC:66557 | |
| Genetic reagent (D. melanogaster) | QUAS-FLP | Bloomington Drosophila Stock Center | BDSC:30127 | |
| Genetic reagent (D. melanogaster) | UAS-FRT-GAL80-STOP-FRT | Bloomington Drosophila Stock Center | BDSC:38880 | |
| Genetic reagent (D. melanogaster) | UAS-FRT-STOP-Kir2.1-FRT | Bloomington Drosophila Stock Center | BDSC:67686 | |
| Genetic reagent (D. melanogaster) | UAS(FRT.mCherry)ReaChR | Bloomington Drosophila Stock Center | BDSC:53743 | |
| Genetic reagent (D. melanogaster) | UAS-TNT | Bloomington Drosophila Stock Center | BDSC:28997 | |
| Genetic reagent (D. melanogaster) | LexAop-GCaMP6s | Bloomington Drosophila Stock Center | BDSC:64413 | |
| Genetic reagent (D. melanogaster) | Tmc1 | Bloomington Drosophila Stock Center | BDSC:66556 | |
| Genetic reagent (D. melanogaster) | w[∗]; P{w[+Mc] = UAS-nSyb-spGFP1-10}2, P{w[+Mc] = lexAop-CD4-spGFP11}2/CyO | Bloomington Drosophila Stock Center | BDSC:64314 | |
| Genetic reagent (D. melanogaster) | UAS-RedStinger | Bloomington Drosophila Stock Center | BDSC:8547 | |
| Genetic reagent (D. melanogaster) | Piezo-GAL4 | Bloomington Drosophila Stock Center | BDSC:59266 | |
| Genetic reagent (D. melanogaster) | UAS-Piezo-RNAi | Vienna Drosophila RNAi Center | VDRC:105132 | |
| Genetic reagent (D. melanogaster) | UAS-FRT-STOP-FRT-GCaMP6m | other | From Laboratory of Chuan Zhou, Institute of Zoology, Chinese Academy of Sciences | |
| Genetic reagent (D. melanogaster) | nompC-QF | Yang et al., 2021 | From Laboratory of Yuh Nung Jan, UCSF | |
| Genetic reagent (D. melanogaster) | nompC1/CyO | Yang et al., 2021 | From Laboratory of Yuh Nung Jan, UCSF | |
| Genetic reagent (D. melanogaster) | nompCf00914/CyO | Yang et al., 2021 | From Laboratory of Yuh Nung Jan, UCSF | |
| Genetic reagent (D. melanogaster) | piezoKO | Zhang et al., 2013 | From Laboratory of Yuh Nung Jan, UCSF | |
| Genetic reagent (D. melanogaster) | UAS-nompC-RNAi; UAS-Dicer2 | Yang et al., 2021 | From Laboratory of Yuh Nung Jan, UCSF | |
| Genetic reagent (D. melanogaster) | UAS-rdl-RNAi | Yang et al., 2021 | From Laboratory of Xin Liang, THU | |
| Genetic reagent (D. melanogaster) | UAS-P2X2 | Yang et al., 2021 | From Laboratory of Yufeng Pan, School of Life Science and Technology, Southeast University | |
| Genetic reagent (D. melanogaster) | UAS(FRT.stop)CsChrimson-mVenus | Wu et al., 2019a | From Laboratory of Yufeng Pan, School of Life Science and Technology, Southeast University | |
| Genetic reagent (D. melanogaster) | MN12-GAL4 | Manzo et al., 2012 | From Laboratory of Kristin Scott, UCB | |
| Genetic reagent (D. melanogaster) | MN11-GAL4 | Manzo et al., 2012 | From Laboratory of Kristin Scott, UCB | |
| Genetic reagent (D. melanogaster) | MN12-LexA | This paper | Hacked from MN12-GAL4 | |
| Genetic reagent (D. melanogaster) | MN11-LexA | This paper | Hacked from MN11-GAL4 | |
| Genetic reagent (D. melanogaster) | Tmc-GAL4, UAS(FRT.stop)CsChrimson-mVenus/CyO | This paper | Recombined from BDSC:66557 and UAS(FRT.stop)CsChrimson-mVenus | |
| Genetic reagent (D. melanogaster) | nompC-QF, QUAS-Flp/TM6B | This paper | Recombined from BDSC:30127 and nompC-QF | |
| Genetic reagent (D. melanogaster) | Tmc-GAL4, UAS-FRT-GAL80-STOP-FRT/CyO | This paper | Recombined from BDSC:66557&38880 | |
| Genetic reagent (D. melanogaster) | Piezo-GAL4AD | This paper | Hacked from BDSC:59266 | |
| Genetic reagent (D. melanogaster) | Tmc-GAL4DBD | This paper | Hacked from BDSC:66557 | |
| Antibody | anti-Rabbit Alexa 488(Goat polyclonal) | Invitrogen | Cat#A11008; RRID: AB_143165 | 1:200 |
| Antibody | anti-Rabbit Alexa 555(Goat polyclonal) | Invitrogen | Cat#A21428; RRID: AB_2535849 | 1:200 |
| Antibody | anti-Mouse Alexa 488(Goat polyclonal) | Invitrogen | Cat#A11001; RRID: AB_2534069 | 1:200 |
| Antibody | anti-Mouse Alexa 647(Goat polyclonal) | Invitrogen | Cat#A21235; RRID: AB_2535804 | 1:200 |
| Antibody | anti-Brp(Mouse monoclonal) | Developmental Studies Hybridoma Bank | Cat# nc82; RRID: AB_2314866 | 1:500 |
| Antibody | anti-GFP(Rabbit polyclonal) | Invitrogen | Cat#A11122; RRID: AB_221569 | 1:500 |
| Antibody | anti-RFP(Rabbit polyclonal) | Rockland | Cat#600-401-379S; RRID: AB_11182807 | 1:500 |
| Antibody | anti-GFP(Mouse monoclonal) | Sigma-Aldrich | Cat# G6539; RRID: AB_259941 | 1:200 |
| Chemical compound, drug | 4% PFA | Dingguo Biotech | Cat#AR-0211;CAS:30525-89-4 | |
| Chemical compound, drug | Brilliant Blue | Shi-tou | Cat#GB 7655.1 | |
| Chemical compound, drug | Methylcellulose | Sigma-Aldrich | M7140-100G | |
| Chemical compound, drug | All trans retinal | Sigma-Aldrich | R2500-500MG | |
| Chemical compound, drug | Phalloidin | AAT bioquest | Cat#23127 | |
| Chemical compound, drug | Adenosine-5 | aladdin | Lot#I2118087 | |
| Software, algorithm | ImageJ and Fiji | NIH; Schindelin et al., 2012 | https://imagej.nih.gov/ij/ http://fiji.sc/ | |
| Software, algorithm | GraphPad Prism 8 | GraphPad Software | https://www.graphpad.com/scientific-software/prism/ | |
| Software, algorithm | Adobe Illustrator | Adobe Systems | https://www.adobe.com/products/illustrator.html |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Pharyngeal mechanosensory neurons control food swallow in Drosophila melanogaster
eLife 12:RP88614.
https://doi.org/10.7554/eLife.88614.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}