The pelvic organs receive no parasympathetic innervation
- Institut de Biologie de l’ENS (IBENS), Inserm, CNRS, École normale supérieure, PSL Research University, France
- Faculty of Medicine, Health & Human Sciences, Macquarie University, Macquarie Park, Australia
- Institut Curie, PSL University, ICGex Next-Generation Sequencing Platform, France
- GenomiqueENS, Institut de Biologie de l'ENS (IBENS), Département de biologie, École normale supérieure, CNRS, INSERM, Université PSL, France
- Inserm U955, Mondor Institute for Biomedical Research (IMRB), France
- Institut Curie, Inserm U830, PSL Research University, Diversity and Plasticity of Childhood Tumors Lab, France
- Institute of Clinical Neuroanatomy, Dr. Senckenberg Anatomy, Neuroscience Center, Goethe University, Germany
Figures
Figure 1 with 4 supplements
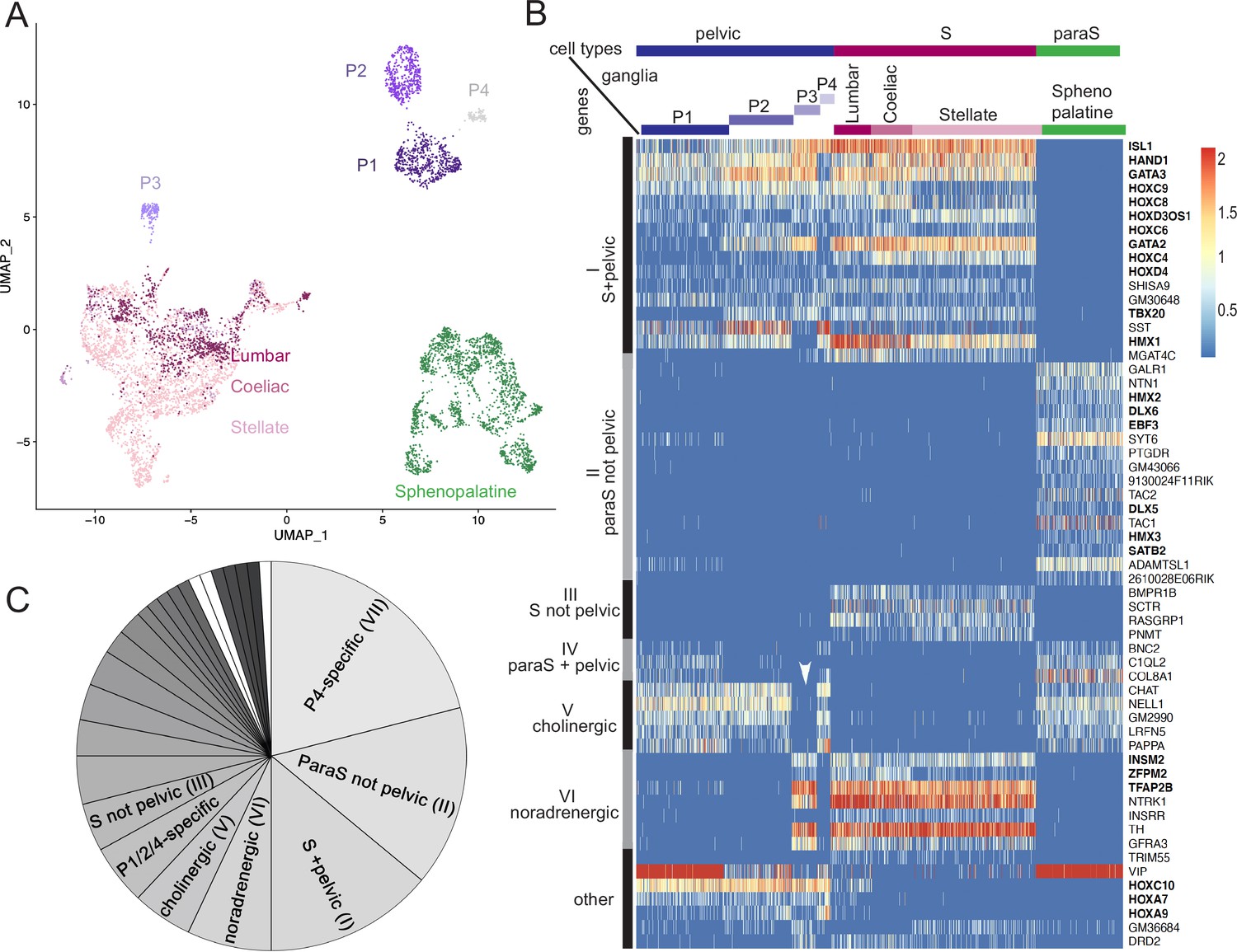
The pelvic ganglion does not contain parasympathetic neurons and is made of sympathetic-like neurons.
(A) Uniform Manifold Approximation and Projection (UMAP) of cells isolated from three sympathetic ganglia (lumbar, stellate, and celiac), a parasympathetic ganglion (sphenopalatine), and the pelvic ganglion dissected from postnatal day 5 mice. The pelvic ganglion is sharply divided into four clusters (P1–4), none of which co-segregates with sympathetic or parasympathetic neurons. (B) Heatmap of the highest scoring 100 genes in an all-versus-all comparison of their dichotomized expression pattern among the four ganglia and four pelvic clusters (see ‘Materials and methods’), excluding genes specific to the pelvic ganglion (shown in Figure 1—figure supplement 2), and keeping only the top-scoring comparison for genes that appear twice. For overall legibility of the figure, the three largest cell groups (lumbar, stellate, and sphenopalatine) are subsampled and genes are ordered by expression pattern (designated on the left), rather than score. ‘Cholinergic’ and ‘noradrenergic’ genes are those that are coregulated with ChAT or Th, regardless of known function. ‘Other’ refers to various groupings that split sympathetic ganglia and are thus not informative about a sympathetic or parasympathetic identity. Transcription factors are indicated in bold face. White arrowhead: pelvic P3 cluster; S, sympathetic; ParaS, parasympathetic. (C) Pie chart of the top 100 genes, counted by expression pattern in the all-versus-all comparison. Genes specific for the P4 cluster dominate (see heatmap in Figure 1—figure supplement 2), followed by those which are ‘parasympathetic-not-pelvic’ and ‘sympathetic-and-pelvic’ (seen in B). The three genes marked in white (which form group IV: Bnc2, C1ql2, Col8a1) are the only ones that are compatible with the current dogma of a mixed sympathetic/parasympathetic pelvic ganglion, by being expressed in the sphenopalatine ganglion and a subset of pelvic clusters (other than the full complement of cholinergic ones, which define group V).
Figure 1—figure supplement 1
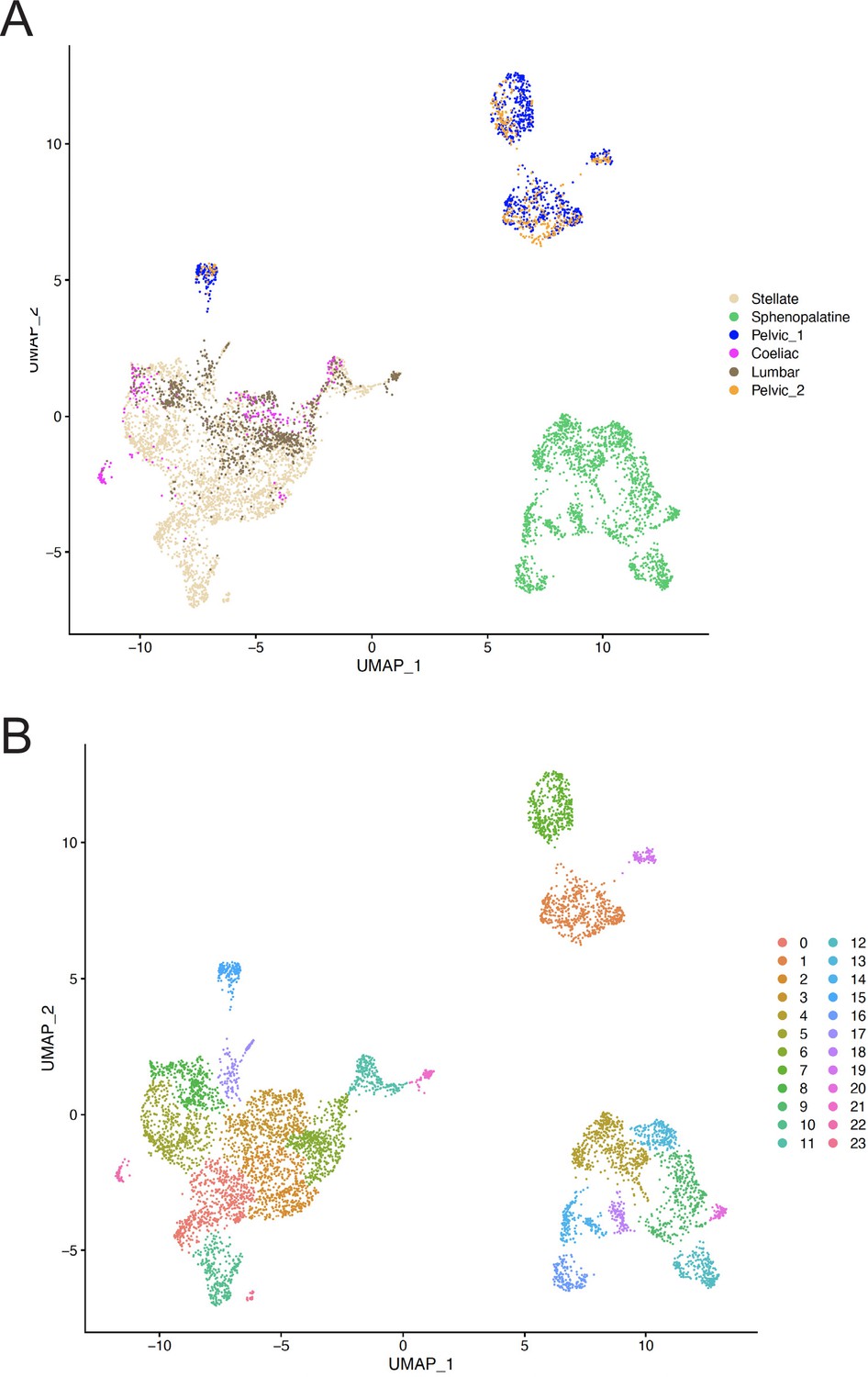
Uniform Manifold Approximation and Projection (UMAP) of all ganglionic neurons.
(A) UMAP of neurons where the sample origin of cells is color-coded to show that both samples of the pelvic ganglion contribute to each of the P1–4 pelvic clusters. (B) UMAP of neurons where the clusters as defined by Seurat are color-coded. Clusters 1, 7, 15, and 19 correspond to ganglion clusters P1, P2, P3, P4 in the text.
Figure 1—figure supplement 2
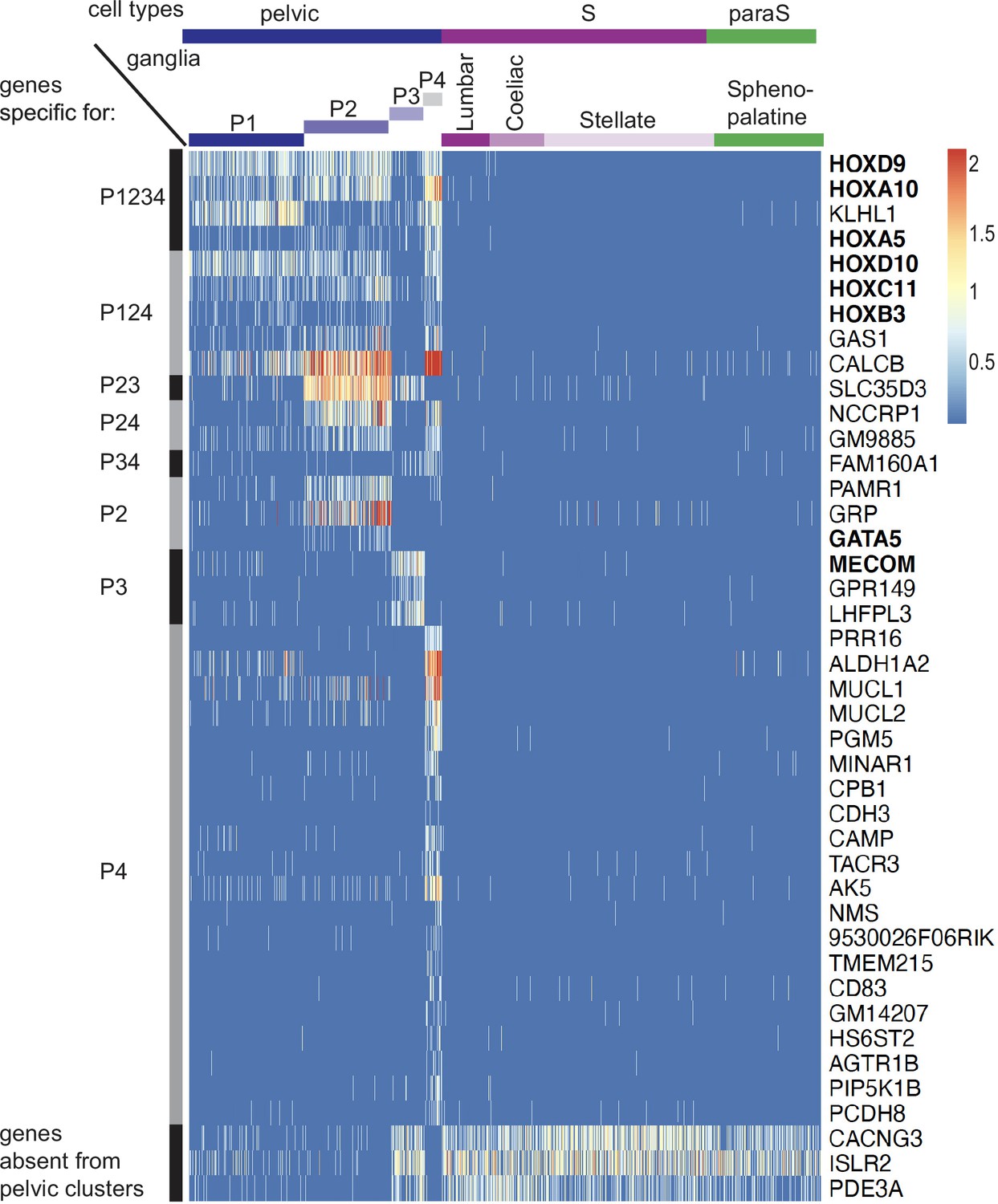
Genes specific for pelvic ganglionic cells among the top 100 genes of an all-versus-all comparison.
These 42 genes correspond to pattern VII of main text. Transcription factors are indicated in bold face.
Figure 1—figure supplement 3
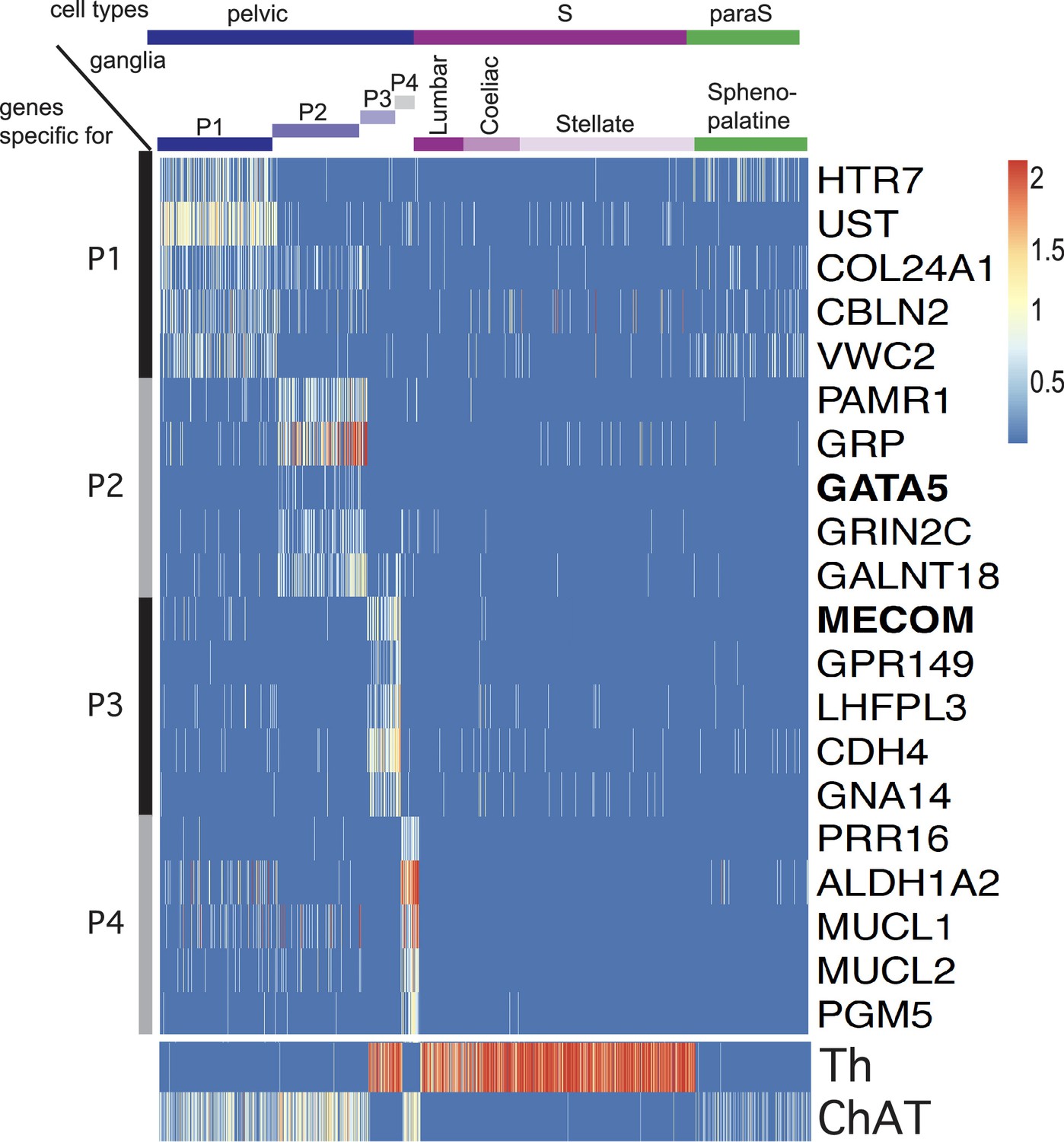
Five top genes for each of the four individual pelvic clusters in an all-versus-all comparison.
P1 appears as the cluster the least sharply defined by specific genes. The only two transcription factors (indicated in bold face) among these top genes are Gata5, expressed in P2 and Mecom expressed in P3. Noradrenergic and cholinergic cells are indicated by Th and ChAT expression in the lower panel.
Figure 1—figure supplement 4
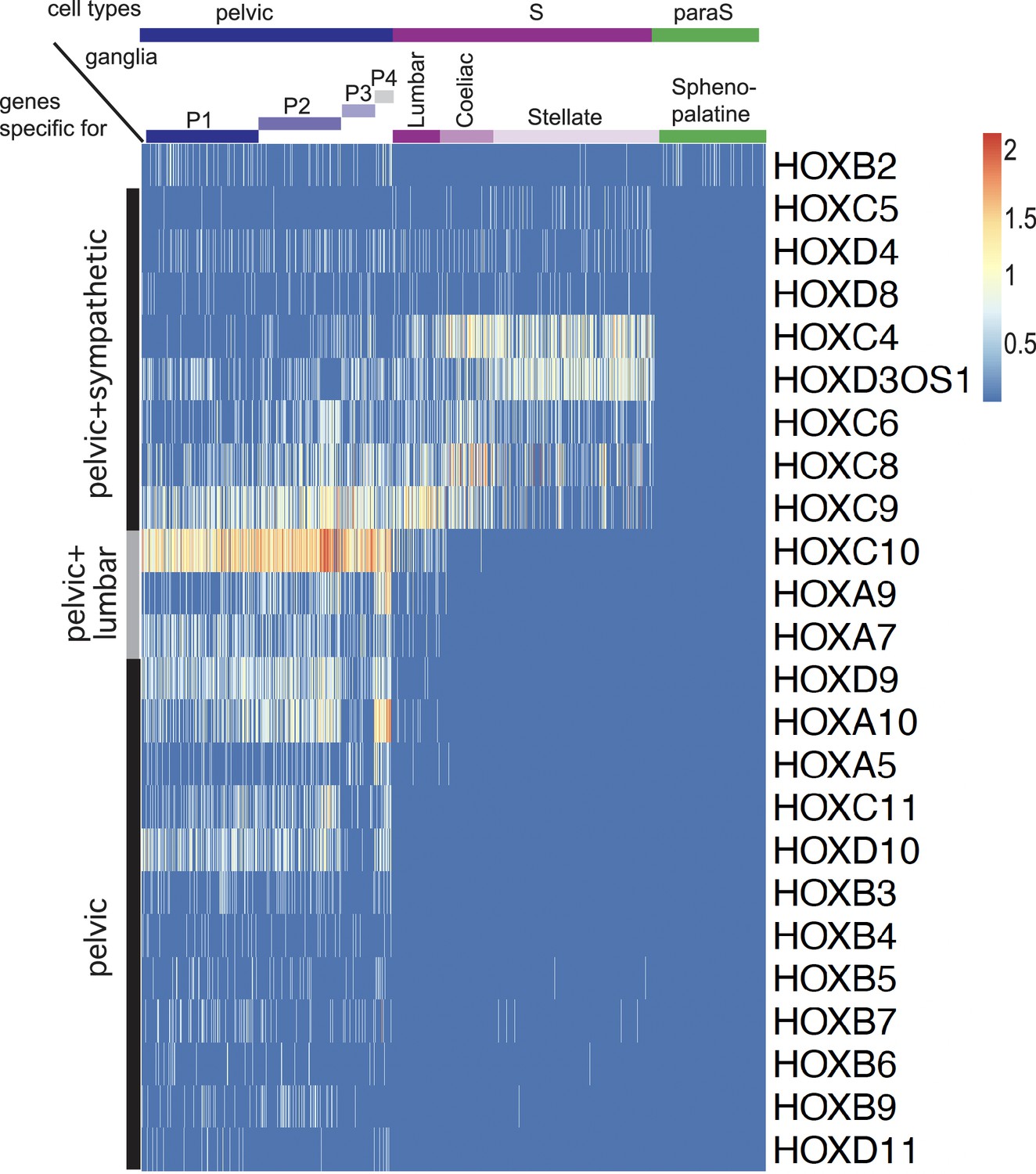
Expression of all Hox genes captured by the single-cell RNA sequencing dataset.
Apart from Hoxb2, all Hox genes are excluded from the sphenopalatine and are expressed either in all sympathetic and pelvic cells (8 genes), caudal sympathetic and pelvic cells (3 genes), or only in pelvic cells (12 genes). Hoxb2 appears in 177th position as {P124/Sphenopalatine} versus {P3/Lumbar/Coeliac/Stellate}, thus as a cholinergic gene in the dichotomized comparison.
Figure 2
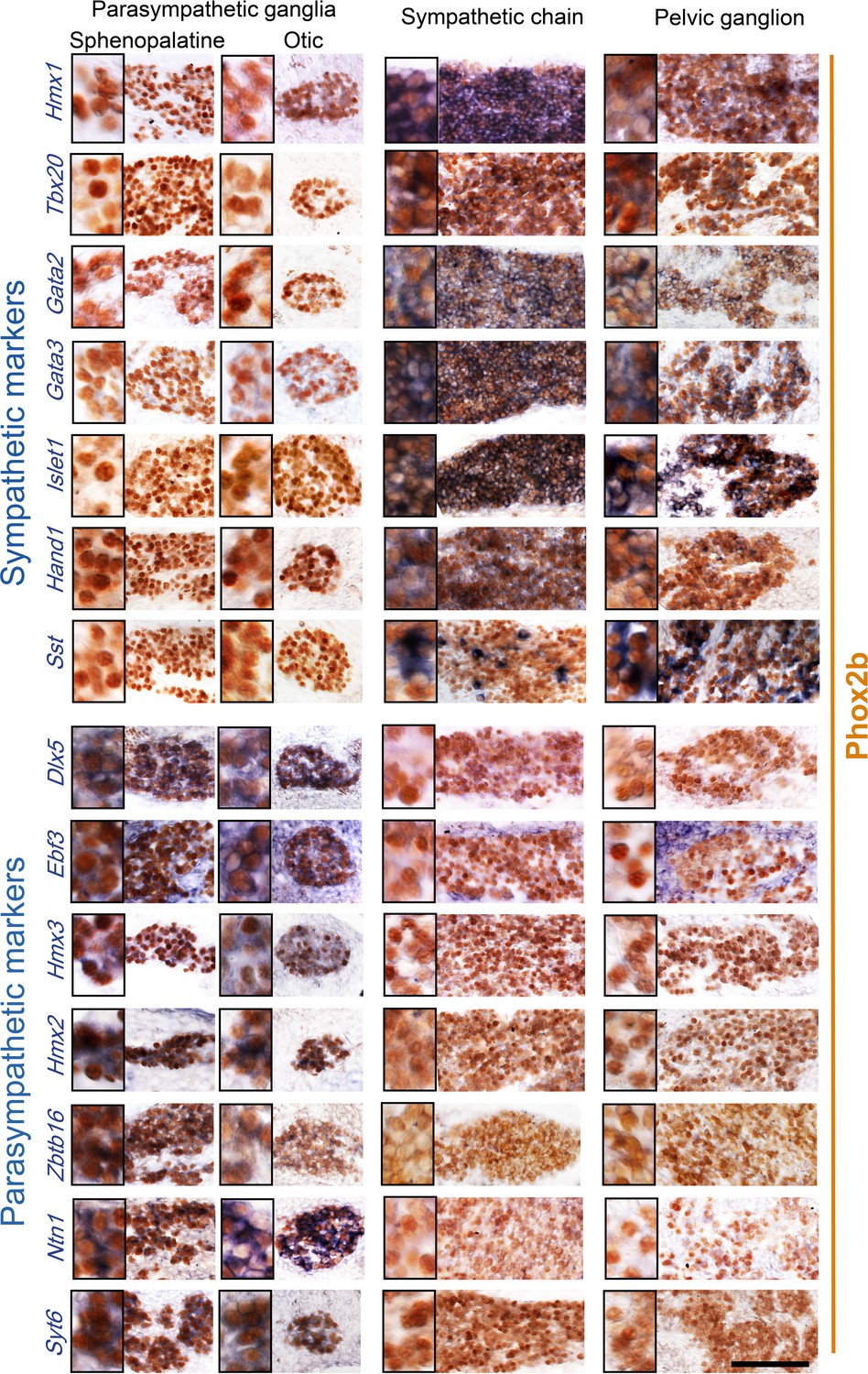
Pelvic ganglion cells express sympathetic but not parasympathetic markers.
Combined immunohistochemistry for Phox2b and in situ hybridization for seven sympathetic markers (upper panels) including six transcription factors or seven parasympathetic markers (lower panels) including five transcription factors, in two parasympathetic ganglia (sphenopalatine and otic), the lumbar sympathetic chain, and the pelvic ganglion, at low and high magnifications (inset on the left) in E16.5 embryos. Ebf3 is expressed in both, the parasympathetic ganglia and the mesenchyme surrounding all ganglia. Sst is expressed in a salt and pepper fashion. Zbtb16, a zinc-finger transcriptional repressor, appeared after the 100 highest scorer gene of our screen, but was spotted as expressed in the sphenopalatine in Genepaint. Some transcription factors detected by the RNA sequencing screen at P5 (Satb2, Dlx6) were expressed below the detection limit by in situ hybridization at E16.5. Scale bar: 100μm.
Figure 3
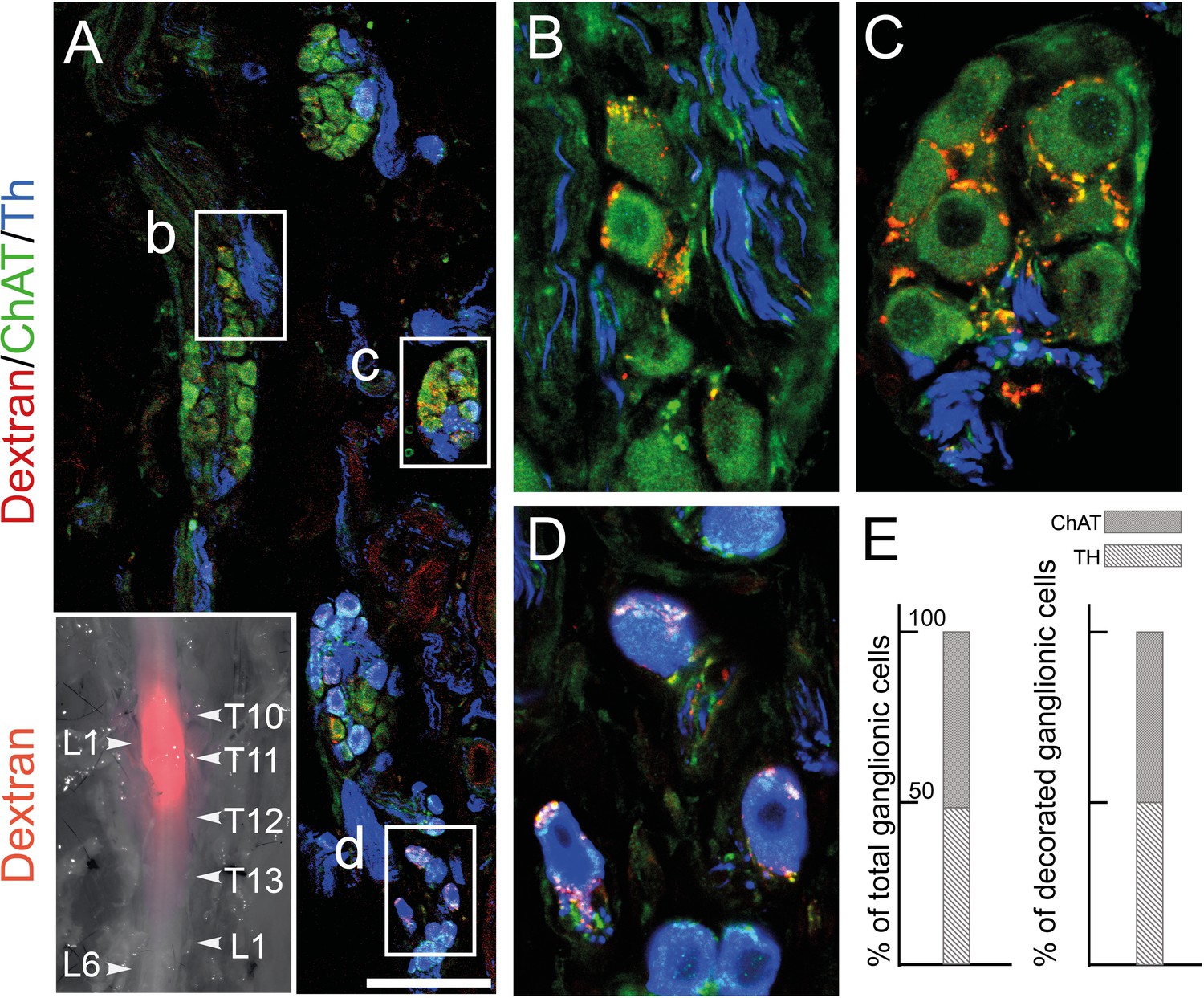
The lumbar outflow targets both cholinergic and noradrenergic pelvic ganglionic cells.
(A–E) Section (A, low magnification, B–D high magnifications of selected regions) through the pelvic ganglion of an adult male mouse stereotactically injected with Dextran at the L1 level of the lumbar spinal cord (inset) and showing dextran filled boutons decorating both choline acetyltransferase (CHAT)+ (B–C) and tyrosine hydroxylase (TH)+ cells (D). Whether they are filled by Dextran or not, cholinergic boutons (green), presumably from spinal preganglionics (lumbar or sacral), are present on most cells. In the inset, levels of the vertebral column are indicated on the right, levels of the spinal cord on the left. (E) Quantification of TH and CHAT cells among total or bouton-decorated ganglionic cells. CHAT+ cells represent 51% of total cells and 50% of decorated cells (for a total of 3186 counted cells, among which 529 decorated cells, on 48 sections in four mice). Scale bar in A: 100μm.
Figure 4
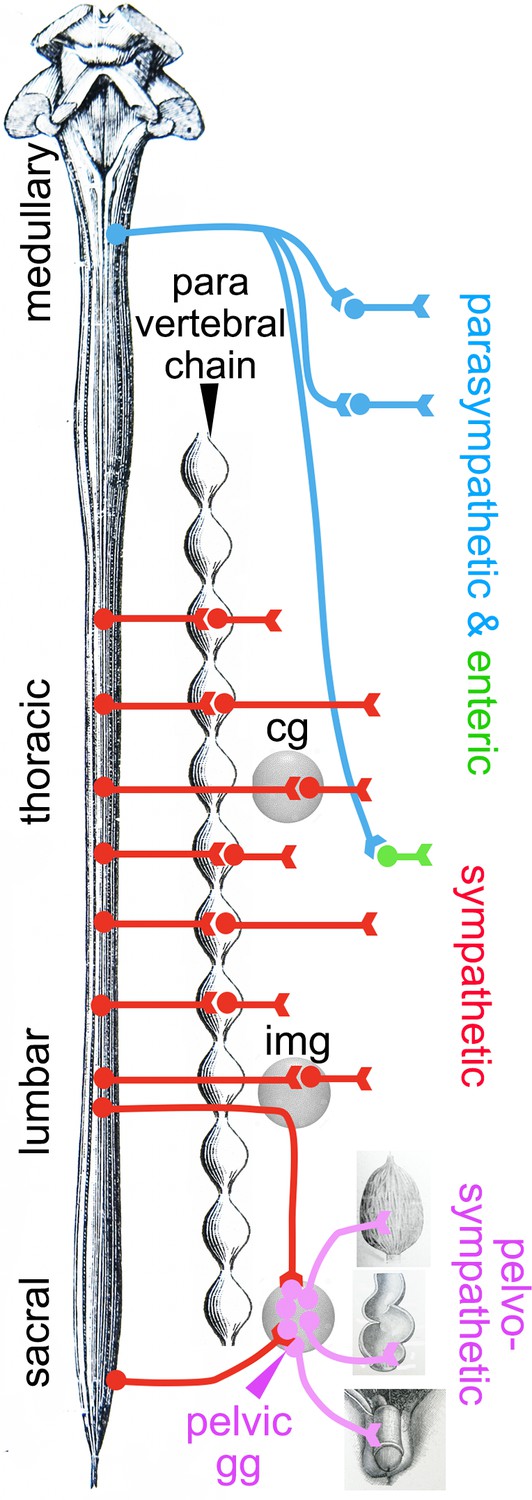
Deployment of the divisions of the autonomic nervous system on the rostro-caudal axis.
Cg, celiac ganglion; img, inferior mesenteric ganglion; pelvic gg, pelvic ganglion. Only the target organs of the pelvo-sympathetic pathway are represented. The adrenal medulla is omitted. The pelvic ganglion is shown with its lumbar input (through the hypogastric nerve) and sacral input (through the pelvic nerve).
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (Mus musculus) | Phox2b::Cre mouse line | D’Autréaux et al., 2011 | BAC transgenic line expressing Cre under the control of the Phox2b promoter | |
| Genetic reagent (Mus musculus) | Rosalox-stop-lox-tdTomato (RosatdT) mouse line | Madisen et al., 2010 | Knock-in line expressing the reporter gene tdTomato from the Rosa locus in a Cre-dependent manner | |
| Antibody (primary) | α-Phox2b rabbit polyclonal | Pattyn et al., 1997 | IHC and IF (1:500) | |
| Antibody (primary) | α-TH rabbit polyclonal | Invitrogen | OPA1-04050 | IF (1:1000) |
| Antibody (primary) | α-Choline acetyltransferase (ChAT) goat polyclonal | Thermo Fisher | PA1-9027 | IF (1:100) |
| Antibodies (secondary) | α-Rabbit PK goat polyclonal | Vector Laboratories | PK-4005 | IHC (1:200) |
| Antibodies (secondary) | Anti-goat 647 donkey polyclonal | Thermo Fisher | A-21447 | IF (1:500) |
| Antibodies (secondary) | Anti-rabbit 488 donkey polyclonal | Thermo Fisher | A-21206 | IF (1:500) |
| Antibodies (secondary) | α-Rabbit Cy3 donkey polyclonal | Jackson | 711-165-152 | IF (1:500) |
| Recombinant DNA reagent | Ebf3 (plasmid) | Gift of S Garel | ||
| Recombinant DNA reagent | Gata3 (plasmid) | Gift of JD Engel | ||
| Recombinant DNA reagent | Hand1 (plasmid) | Gift of P Cserjesi | ||
| Recombinant DNA reagent | Hmx2 (plasmid) | Gift of EE Turner | ||
| Recombinant DNA reagent | Hmx3 (plasmid) | Gift of S Mansour | ||
| Recombinant DNA reagent | Islet1 (plasmid) | Tiveron et al., 1996 (10.1523/JNEUROSCI.16-23-07649.1996) | ||
| Recombinant DNA reagent | Tbx20 (plasmid) | Dufour et al., 2006 (10.1073/pnas.0600805103) | ||
| Recombinant DNA reagent | Sst (plasmid) | Clone Image ID #4981984 | ||
| Sequence-based reagent | Dlx5_F | This paper | PCR primers | 5’ -GACGCAAA CACAGGTGAAAATCTGG-3’ |
| Sequence-based reagent | Dlx5_R | This paper | PCR primers | 5’-GGGCGGGGC TCTCTGAAATG-3’ |
| Sequence-based reagent | Gata2_F | This paper | PCR primers | 5’-TTGTGTTCTT GGGGTCCTTC-3’ |
| Sequence-based reagent | Gata2_R | This paper | PCR primers | 5’-GCTTCTGTGG CAACGTACAA-3’ |
| Sequence-based reagent | Hmx1_F | This paper | PCR primers | 5’-CGTTCGCCAC TATCCAAACGGG-3’ |
| Sequence-based reagent | Hmx1_R | This paper | PCR primers | 5’-TGTCAGGACT TAGACCACCTCCG-3’ |
| Sequence-based reagent | Ntn1_F | This paper | PCR primers | 5’-CTTCCTCACC GACCTCAATAAC-3’ |
| Sequence-based reagent | Ntn1_R | This paper | PCR primers | 5’-GCGATTTAG GTGACACTATAGTTGTGCCTACAGTCACACAC C-3’ |
| Sequence-based reagent | Syt6_F | This paper | PCR primers | 5’-GTGGTCTTCT TGTCCCGTGT-3’ |
| Sequence-based reagent | Syt6_R | This paper | PCR primers | 5’-CATGTGCTTA CAGGGTGTGG-3’ |
| Sequence-based reagent | Zbtb16_F | This paper | PCR primers | 5’-ATGAAAACAT ACGGGTGTGAA-3’ |
| Sequence-based reagent | Zbtb16_R | This paper | PCR primers | 5’-CCAAGGCCAA GTAACTATCAGG-3’ |
| Chemical compound, drug | Tetramethyl-rhodamine-conjugated dextran | Thermo Fisher | D3308 | For tracing experiments |
| Chemical compound, drug | NBT-BCIP solution | Sigma | B1911 | For ISH experiments |
| Chemical compound, drug | 3,3’-Diaminobenzidine (DAB) | Sigma | D12384 | For IHC experiments |
| Software, algorithm | Cell Ranger software | 10x Genomics | 6.0.1 |
Additional files
-
Supplementary file 1
Table of the 500 highest-scoring genes for each of the 254 possible dichotomizations of ganglia and pelvic clusters (i.e. partitioning the ganglia and pelvic clusters into two mutually exclusive sets, subset_1 and subset_2), ordered by score, and after removal, for each gene, of all dichotomizations below the highest scoring one (resulting in the ranking of 7593 genes).
Each row in the table indicates the gene name, the score, a description of the dichotomization (under the field ‘group.compar.string’, in the format {subset_1}vs{subset_2}), and each ganglion or pelvic cluster, marked with 1 or 0 to indicate that the cluster belongs, respectively, to subset_1 or subset_2.
- https://cdn.elifesciences.org/articles/91576/elife-91576-supp1-v2.xlsx
-
Supplementary file 2
Violin plots of the top 100 highest-scoring genes, displaying the SCT-normalized values for each of the eight ganglion or pelvic cluster: stellate, sphenopalatine, pelvic_1, coeliac, lumbar, and pelvic_2 color-coded as indicated.
- https://cdn.elifesciences.org/articles/91576/elife-91576-supp2-v2.pdf
-
Supplementary file 3
Bar plots of the top 100 highest-scoring genes, displaying the proportion of cells (ranging between 0 and 1) in each ganglion or pelvic cluster that express at least one read of a given gene.
- https://cdn.elifesciences.org/articles/91576/elife-91576-supp3-v2.pdf
-
MDAR checklist
- https://cdn.elifesciences.org/articles/91576/elife-91576-mdarchecklist1-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The pelvic organs receive no parasympathetic innervation
eLife 12:RP91576.
https://doi.org/10.7554/eLife.91576.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}