The components of an electrical synapse as revealed by expansion microscopy of a single synaptic contact
- Dominick P. Purpura Department of Neuroscience, Albert Einstein College of Medicine, United States
Figures
Figure 1 with 2 supplements
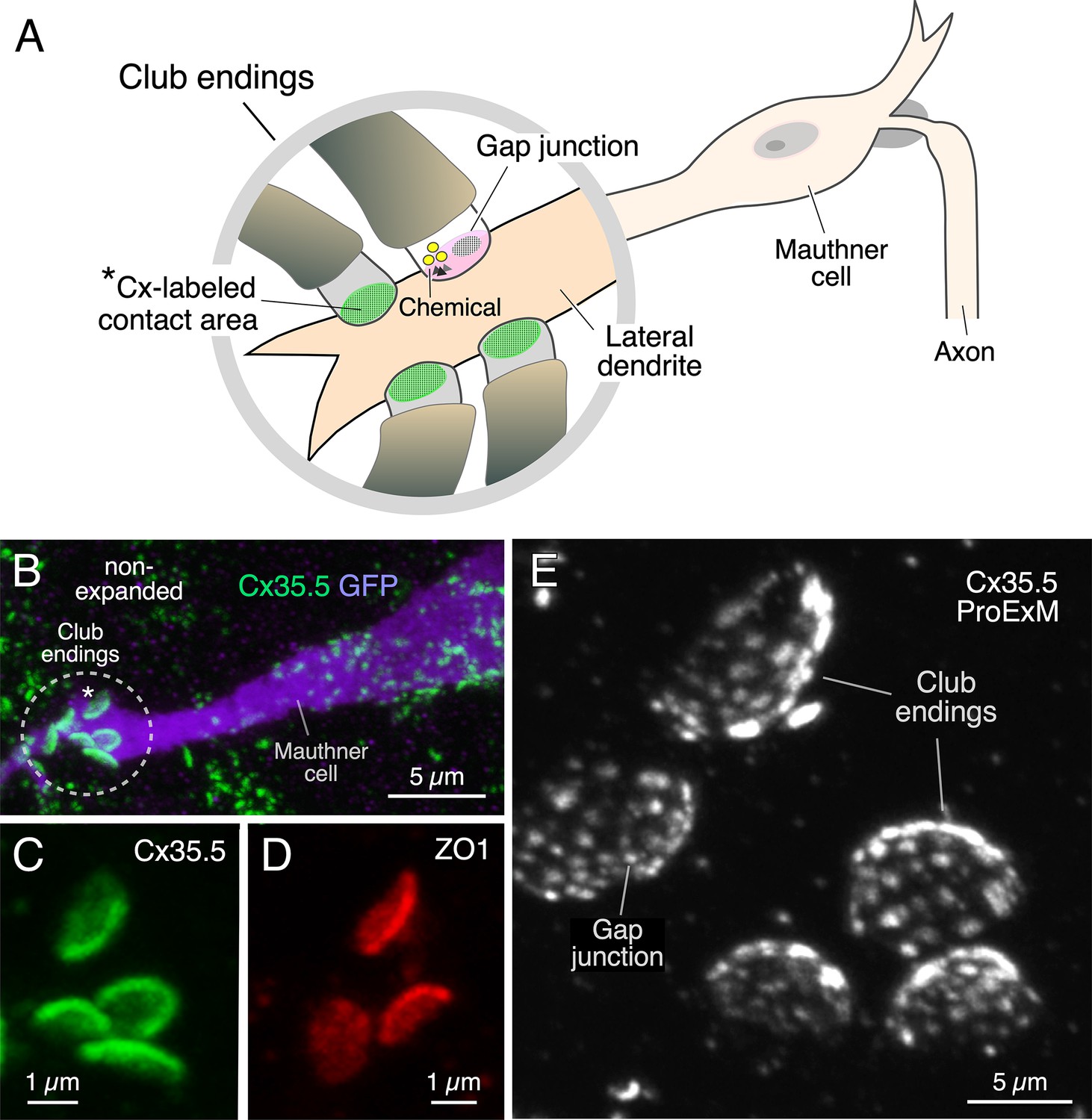
Expansion microscopy of club ending (CE) contact areas in larval zebrafish.
(A) The cartoon illustrates the auditory afferents that terminate as single CEs, each containing gap junctions (GJs, green) and specializations for chemical transmission (Chemical), on the distal portion of the lateral dendrite of the Mauthner (M-) cell. Synaptic contact areas labeled with connexin antibody (see B and C) are represented in green. (B) Confocal image with anti-GFP (purple) and anti-Cx35/36 (green), which labels both Cx35.5 and Cx35.1, showing a long stretch of the lateral dendrite of the M-cell (projection of 34 confocal z-sections at 0.39 µm z-step size), revealing the contact areas (indicated by the asterisk here and the cartoon of A) of several CEs. (C, D) Contact areas of individual CEs labeled with anti-Cx35/36 (C, green; projection of 12 sections at 0.39 µm z-step size) and anti-ZO1 (D, red; projection of 4 sections at 0.39 µm z-step size). (E) Protein-retention expansion microscopy (ProExM) with anti-Cx35.5 increases the size of CE synaptic contact areas, enabling the visualization of intrasynaptic components (projection of 19 sections at 0.88 µm z-step size). The scale bars represent actual dimensions and, therefore, ProExM images were not adjusted for expansion factor.
Figure 1—figure supplement 1
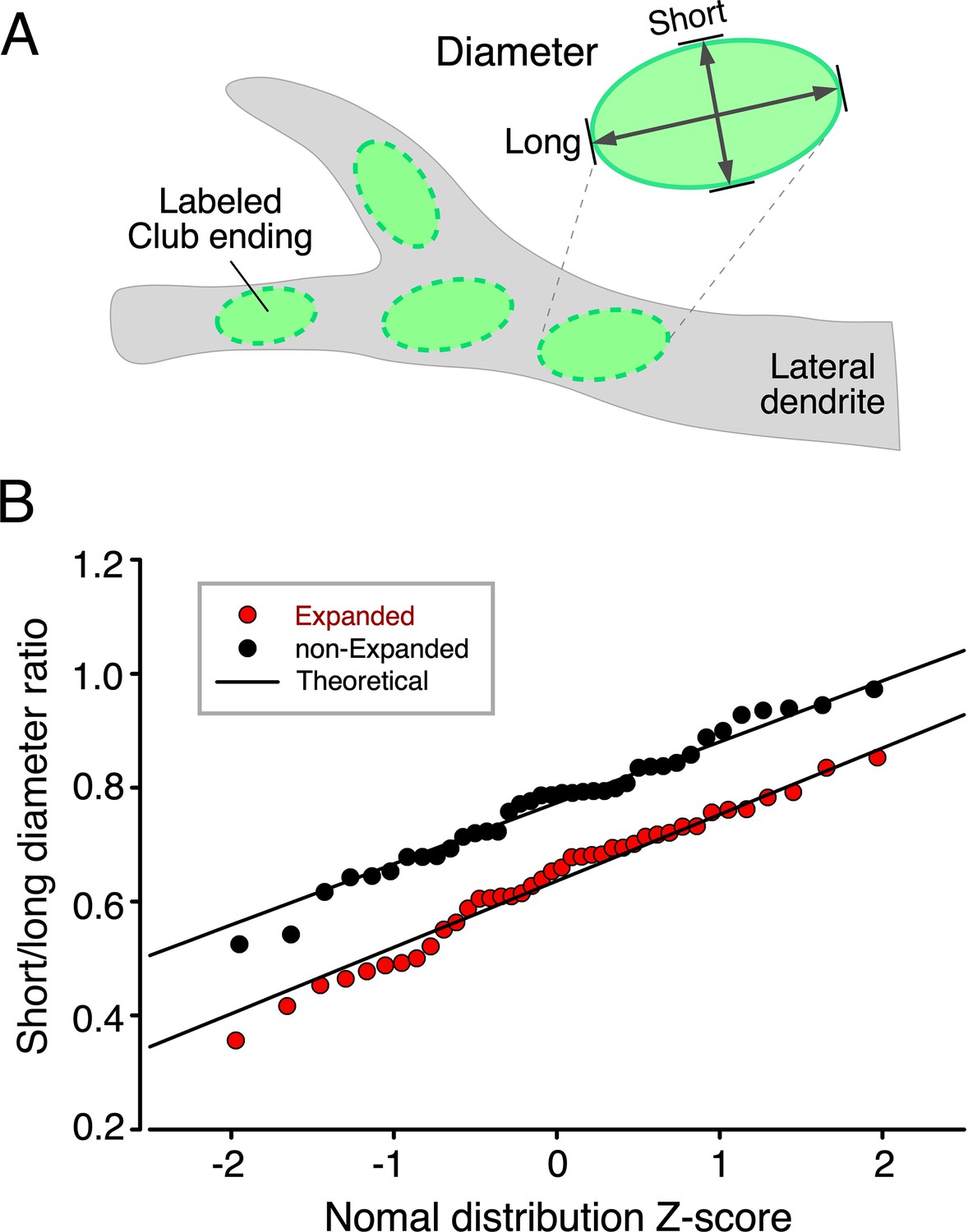
Short–long-diameter ratio probability plots.
(A) Cartoon illustrates labeled club ending (CE) contact areas in the lateral dendrite of the M-cell and the measurement of their short and long diameters. (B) Short–long-diameter ratios obtained from expanded (red circles) and non-expanded (black circles) CE contact areas are shown plotted against their standardized normal Z-scores calculated from sampled cumulative probabilities. Straight lines (Theoretical) show data expected from normally distributed short–long ratios derived from sample means and standard deviations. In both expanded and non-expanded cases, experimental distributions fit theoretical normal distributions well. It may also be observed that the variability of ratios is nearly identical for expanded and non-expanded data, indicating that the expansion process had no selective effects across the CE population.
Figure 1—video 1
Expansion microscopy with anti-Cx35.5.
Figure 2
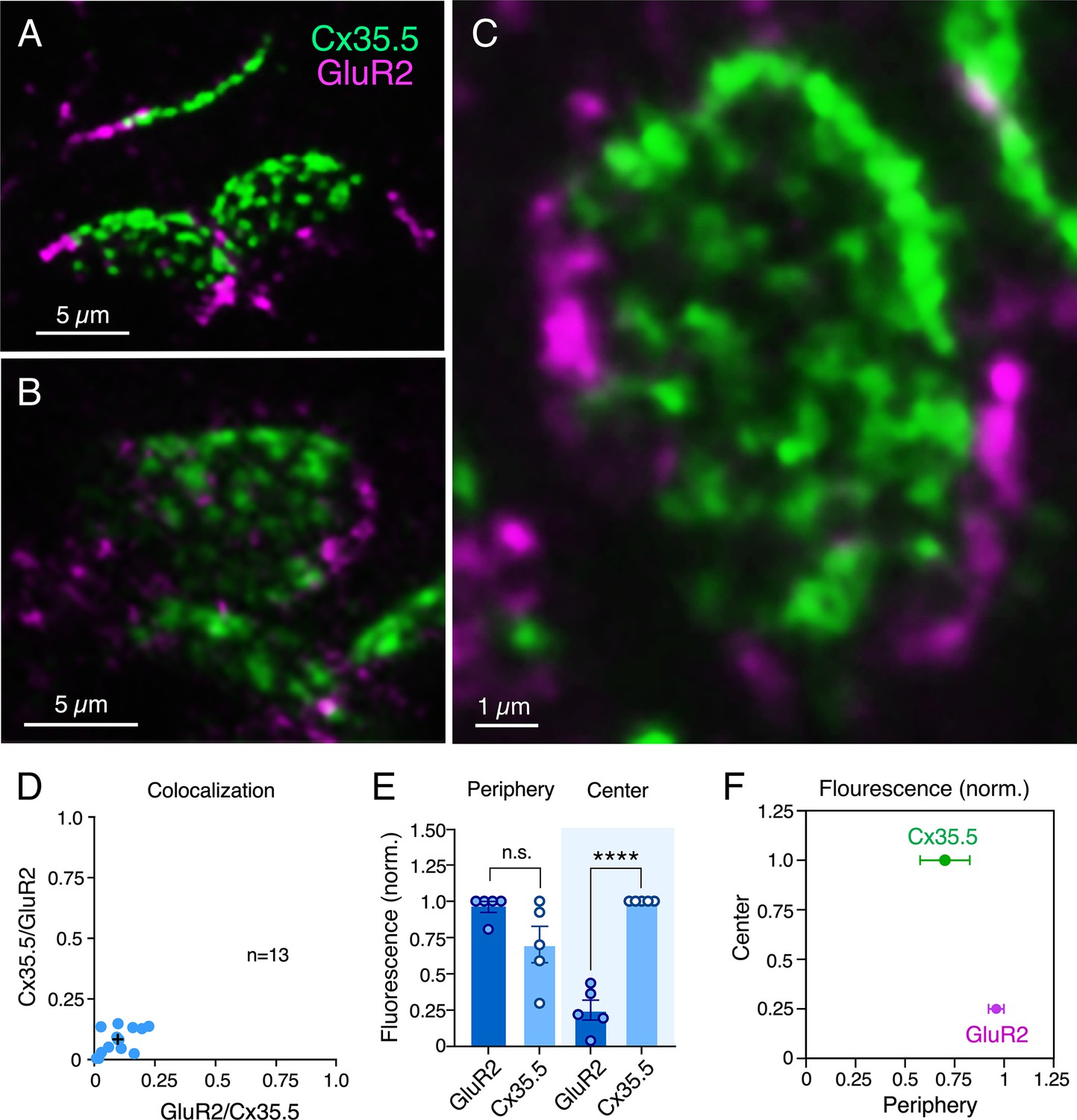
Electrical and chemical transmitting areas are mutually exclusive.
(A, B) Expanded synaptic contact areas labeled with anti-Cx35.5 (green) and anti-GluR2 (magenta) (A: projection of 18 sections at 0.55 µm z-step size; B: projection of 15 sections at 0.50 µm z-step size). (C) ‘En face’ view of an expanded synaptic contact area showing that GluR2 labeling is restricted to the periphery of the contact, whereas Cx35.5 labeling is distributed throughout the whole contact area (projection of 46 sections at 0.65 µm z-step size). (D) Graph shows the lack of colocalization (see ‘Materials and methods’) between Cx35.5 and GluR2 fluorescence at individual club ending (CE) contacts, determined by the Manders’ colocalization coefficient: GluR2/Cx35.5 0.10 ± 0.020 (x-axis); Cx35.5/GluR2 0.08 ± 0.015 (y-axis), n = 13 CEs from five fish. Cross mark indicates the average value. (E) Quantification of fluorescence over area for Cx35.5 and GluR2 at the ‘Center” (central ¾) and ‘Periphery’ (remaining ¼) of the CE contact area. Values of fluorescence/area are represented as normalized to those of Cx35.5 in the center (higher value): GluR2 center: 0.25 ± 0.069; Cx35.5 periphery: 0.70 ± 0.126; GluR2 periphery: 0.96 ± 0.039 (n = 5 CEs from five fish). While fluorescence for Cx35.5 and GluR2 is not significantly different in the periphery (n.s.), Cx35.5 distinctly predominates over GluR2 at the center (Student’s t-test, p<0.0001). (F) Graphical description of the center vs. periphery distribution of Cx35.5 and GluR2 for the data described in (E). The scale bars represent actual dimensions; expanded images were not adjusted for expansion factor.
Figure 3 with 1 supplement
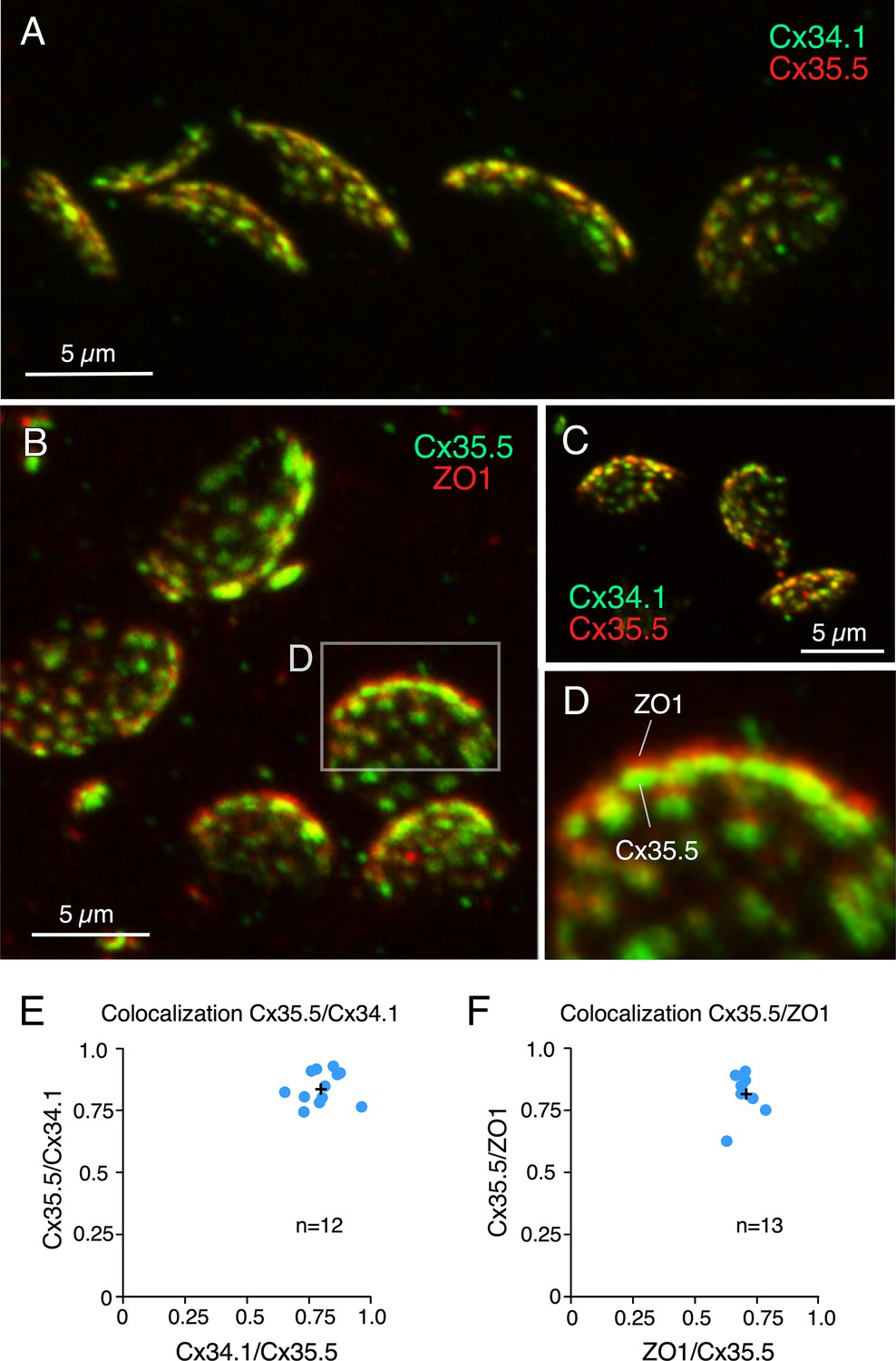
Labeling for gap junction proteins reveals the presence of multiple puncta at expanded club ending (CE) synaptic contact areas.
(A) CE synaptic contact areas labeled with anti-Cx34.1 and anti-Cx35.5 (projection of 69 sections at 0.65 µm z-step size). (B) Contact areas labeled with anti-Cx35.5 and anti-ZO1. Same experiment as Figure 1E (projection of 19 sections at 0.88 µm z-step size). (C) Labeling with anti-Cx34.1 and anti-Cx35.5 (projection of 50 sections at 0.40 µm z-step size). (D) Magnification of the boxed region in (B) showing a side view of an expanded synaptic contact area labeled for Cx35.5 and ZO1. (E) Graph showing colocalization of Cx35.5 and Cx34.1 fluorescence at individual CEs determined by the Manders’ coefficient: Cx34.1/Cx35.5 0.80 ± 0.023 (x-axis); Cx35.5/Cx34.1 0.84 ± 0.019 (y-axis), n = 12 CEs from six fish. (F) Colocalization of Cx35.5 and ZO1 fluorescence at individual CEs. Manders’ coefficient: ZO1/Cx35.5 0.71 ± 0.012 (x-axis); Cx35.5/ZO1 0.82 ± 0.021 (y-axis), n = 13 CEs from seven fish. Cross mark indicates the average value. The scale bars represent actual dimensions; expanded images were not adjusted for expansion factor.
Figure 3—video 1
Expansion microscopy with anti-Cx35.5 and anti-ZO1.
Figure 4
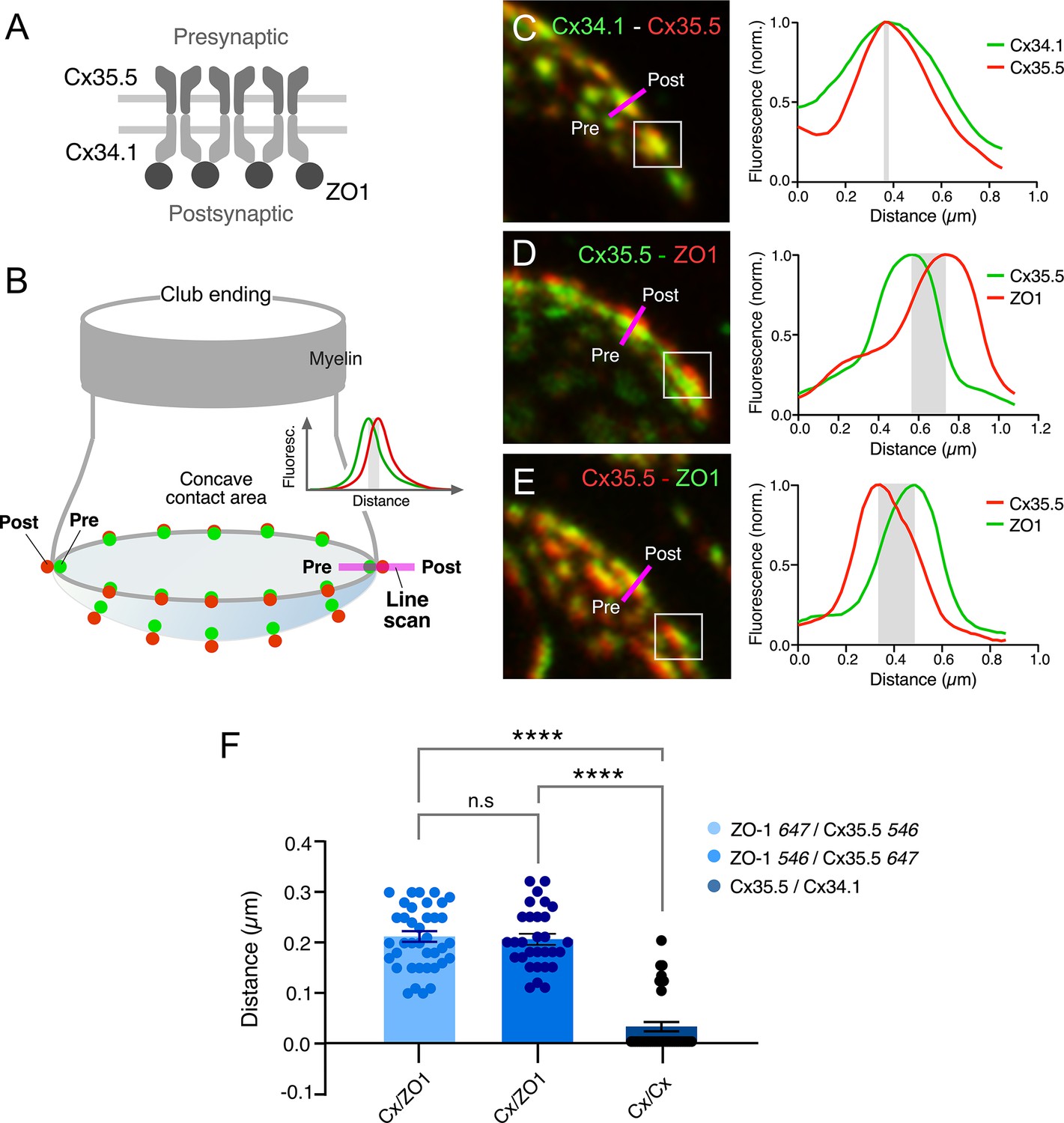
Expansion microscopy reveals the molecular components of gap junction (GJ) plaques at club ending (CE) synaptic contact areas.
(A) Schematic representation of the molecular organization of GJs between CEs (presynaptic) and the M-cell (postsynaptic). The presynaptic and postsynaptic hemichannels are formed by Cx35.5 and Cx34.1, respectively. The scaffolding protein, ZO1, is postsynaptic and interacts with Cx34.1. (B) Cartoon of a CE terminal illustrating the concavity of its contact area with the M-cell. The concavity determines differences in the relative position of presynaptic (green) vs. postsynaptic (red) labeling at different points throughout the contact area. Puncta located in the periphery of the contact are ideally aligned to determine colocalization of fluorescence at individual puncta (line scan, inset). (C–E) Line scan of puncta at expanded contact areas showing colocalization of presynaptic Cx35.5 and postsynaptic Cx34.1 (C) (projection of 69 sections at 0.65 µm z-step size), and presynaptic Cx35.5 and postsynaptic ZO1 (D–E) (D: projection of 26 sections at 0.86 µm z-step size; E: projection of 55 sections at 0.60 µm z-step size). The example in (C) is part of the experiment illustrated in Figure 3A. The magenta lines indicate the position of the line scan in each case. The fluorescence intensity profiles for each fluorophore are illustrated on the right side of each panel. As a control, secondary antibodies were swapped in (E). (F) Bar graph illustrates the distance between the peaks of fluorescence intensity profiles for Cx35.5-Cx34.1 labeling (with either 647Atto or 546Alexa-Cx35.5 vs. either 647Atto or 546Alexa-Cx34.1: 0.03 ± 0.010 µm, n = 37 puncta from six fish) and Cx35.5-ZO1 labeling (546Alexa-ZO1 vs. 647Atto-Cx35.5: 0.21 ± 0.011 µm, n = 30 puncta from three fish). Secondary antibodies were swapped as control (647Atto-ZO1 vs. 546Alexa-Cx35.5: 0.21 ± 0.011 µm, n = 39 puncta from eight fish). Bars represent ± SEM (ANOVA analysis with Tukey’s multiple comparison test correction; ****p<0.0001). The scale bars represent actual dimensions; expanded images were not adjusted for expansion factor.
Figure 5
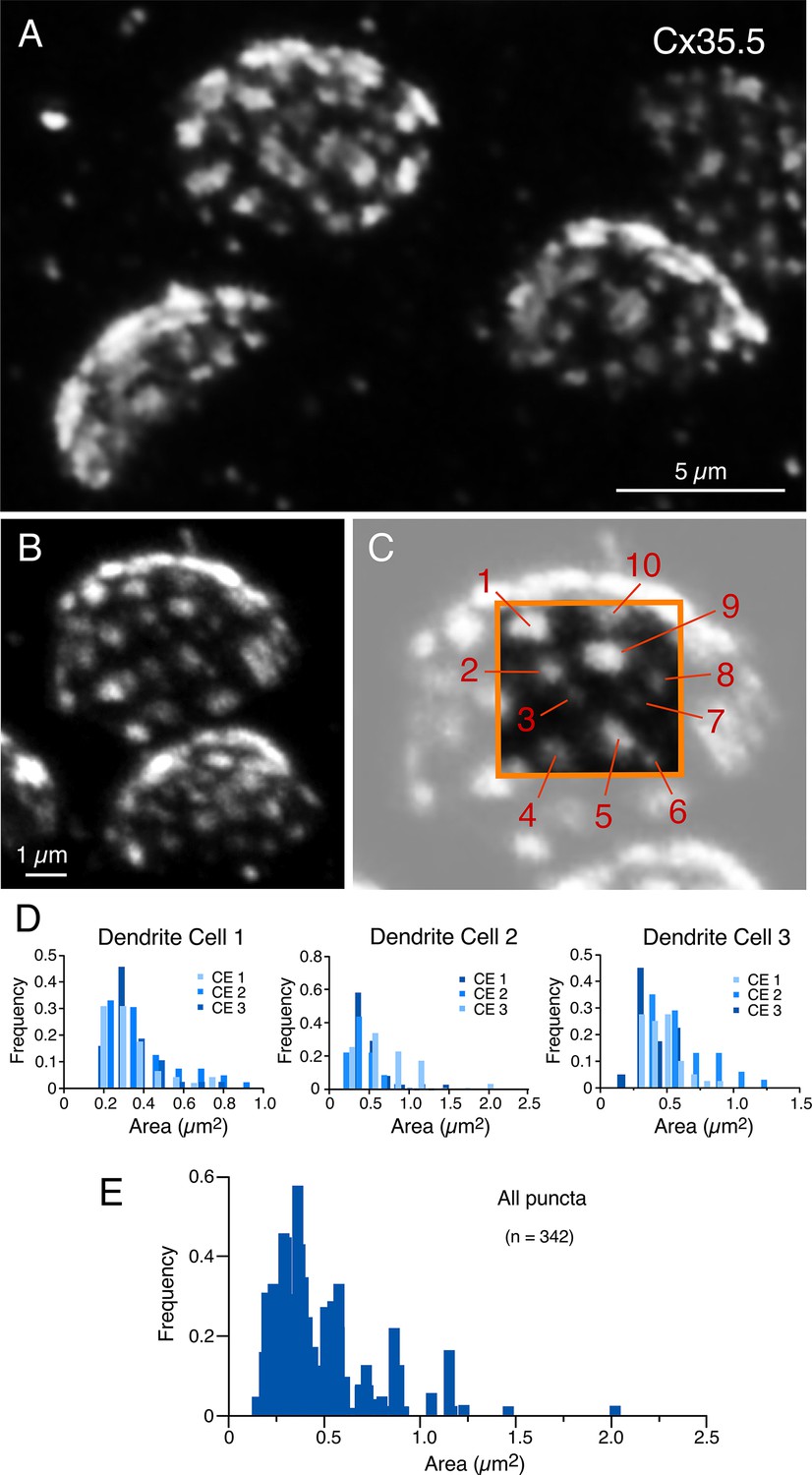
Expansion reveals the presence of multiple, variably sized, gap junctions.
(A, B) ‘En face’ views of expanded club ending (CE) contact areas labeled with anti-Cx35.5 showing multiple puncta with high variability of their size (A: projection of 18 sections at 0.60 µm z-step size; B: projection of 19 sections at 0.88 µm z-step size). (C) Magnification of the CE contact area at the top of (B) (light gray). The area enclosed by the orange box illustrates the wide variability in puncta size, labeled 1–10 (to better highlight the variability in puncta size, the image delimited by the orange box was cropped from and placed on the same region of the lighter image). Panels (B) and (C) are the same experiment as Figures 1E and 3B–D, demonstrating the ability of expansion microscopy for providing multiple layers of information within the same experiment. (D) Frequency histograms summarize the number and size distribution of puncta labeled for Cx35.5 obtained from three dendrites, each belonging to a different fish (bar graphs labeled as ‘Dendrite cell 1’, ‘Dendrite cell 2’, ‘Dendrite cell 3’). Each histogram illustrates, overlapped in different shades of blue, the values obtained from the analysis of three ‘en face’ views of CE terminals. Histograms show similar variability in number and size for all nine terminals. (E) Frequency histogram summarizes the average values resulting from the analysis of all nine ‘en face’ CE views. The scale bars represent actual dimensions; expanded images were not adjusted for the expansion factor.
Figure 6
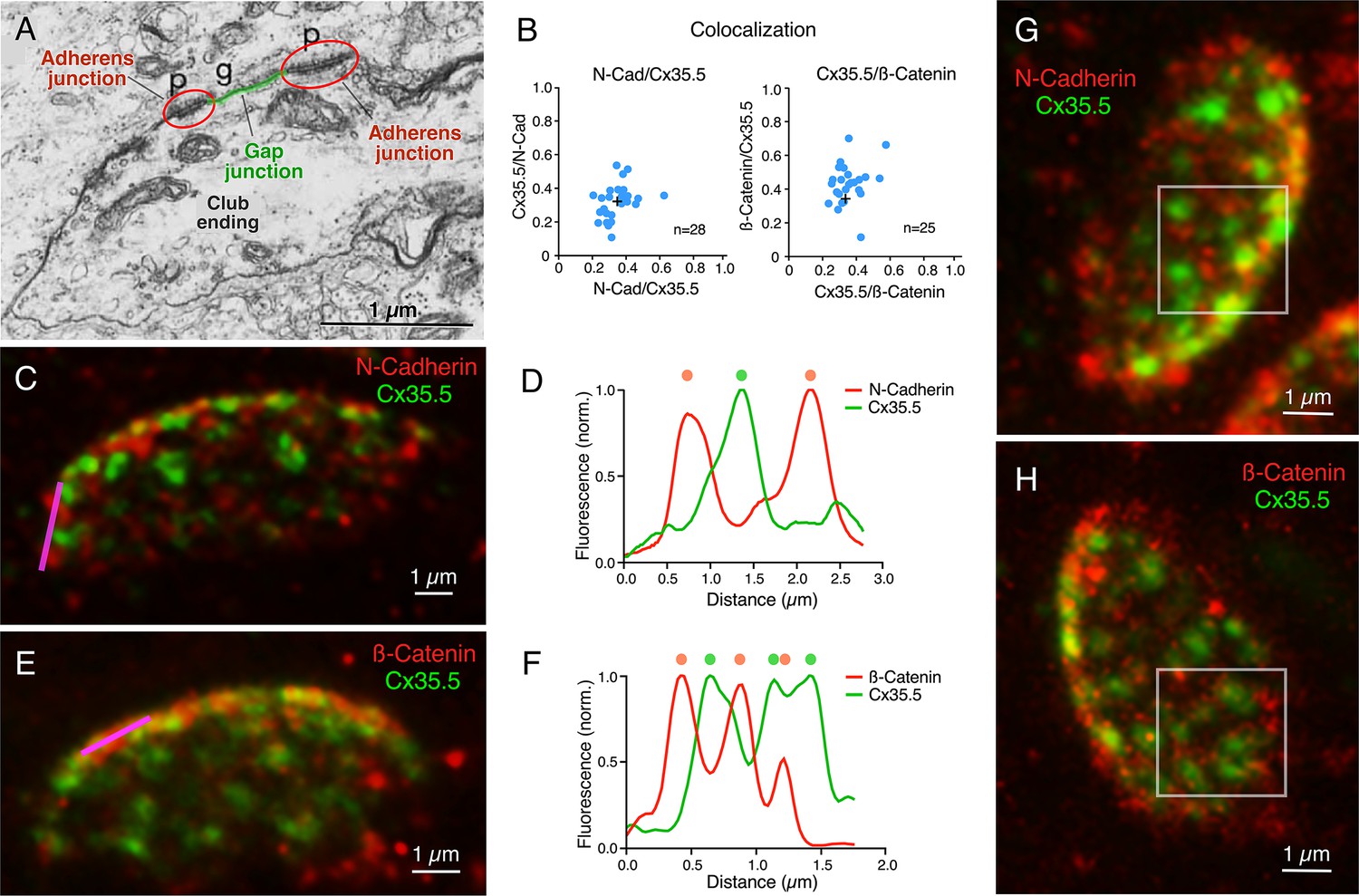
Gap junctions (GJs) at club endings (CEs) are associated with adherens junctions (AJs).
(A) Electron micrograph of a CE obtained in a 6 days post fertilization (dpf) zebrafish showing a GJ (g, highlighted in green) surrounded by AJs (p, encircled in red). ((A) was reproduced from Figure 5 of Kimmel et al., 1981, with permission from John Wiley and Sons. It is not covered by the CC-BY 4.0 license and further reproduction of this panel would need permission from the copyright holder; Kimmel et al., 1981.) (B) GJ and AJ proteins do not colocalize. Left: Cx35.5 and N-cadherin labeling show a low index of colocalization. Manders’ coefficient: N-cadherin/Cx35.5 0.35 ± 0.016 (x-axis); Cx35.5/N-cadherin 0.32 ± 0.019 (y-axis), n = 28 CEs from 10 fish. Right: Cx35.5 and β-catenin labeling also show low colocalization. Manders’ coefficient: Cx35.5/β-catenin 0.36 ± 0.017 (x-axis); β-catenin/Cx35.5 0.43 ± 0.024 (y-axis), n = 25 CEs from 12 fish. Cross mark indicates the average value. (C) Expansion microscopy of a CE contact area labeled for N-cadherin (red) and Cx35.5 (green) (projection of 47 sections at 0.40 µm z-step size). (D) Fluorescence profiles for N-cadherin and Cx35.5 obtained with a line scan (magenta line in C) are mutually exclusive. The small degree of colocalization observed in (B) is likely due to fluorophore amplification and the close spatial association between GJs and AJs, as shown in (A). (E) Image shows an expanded CE contact area labeled for β-catenin (red) and Cx35.5 (green) (projection of 12 sections at 0.60 µm z-step size). (F) Line scan (magenta line in E) shows that labeling for β-catenin and Cx35.5 is also mutually exclusive. (G, H) ‘En face’ view of the expanded contact area double-labeled for N-cadherin and Cx35.5, and β-catenin and Cx35.5, respectively, showing the close association of GJs and AJs throughout the synaptic contact area. Insets: the boxed areas in the ‘en face’ images highlight the mutually exclusive labeling (G: projection of 24 sections at 0.50 µm z-step size; H: projection of 16 sections at 0.60 µm z-step size). The scale bars represent actual dimensions; expanded images were not adjusted for the expansion factor.
© 1981, John Wiley and Sons. Figure 6A is reproduced from Figure 5 from Kimmel et al., 1981, with permission from John Wiley and Sons. It is not covered by the CC-BY 4.0 licence and further reproduction of this panel would need permission from the copyright holder.
Figure 7
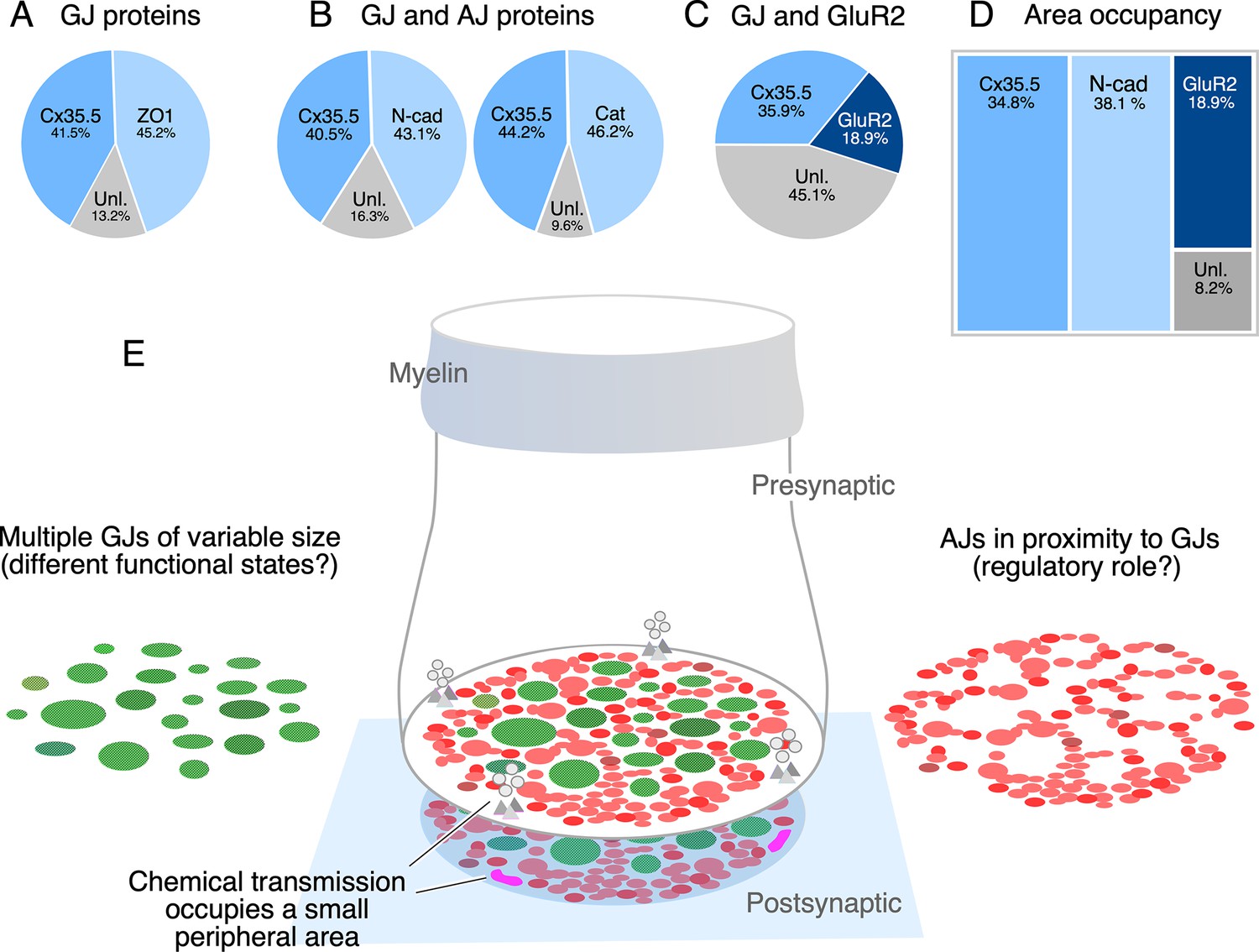
The electrical synapse at the club ending (CE) combines multiple gap junctions (GJs) with adherens junctions (AJs).
(A) Double labeling with anti-Cx35.5 and anti-ZO1 shows a similar proportion of fluorescence at CEs (ZO1 45.24% ± 2.709; Cx35.5 41.53% ± 4.058; unlabeled 13.23%; n = 8 CEs from six fish). The region of interest (ROI) for analysis of fluorescence here and (B) was defined by the outline of Cx35.5 labeling of CEs. (B) Double labeling for N-cadherin and Cx35.5 (left), and for β-catenin and Cx35 (right) also shows similar proportionality (N-Cad 43.20% ± 3.334; Cx35.5 40.48% ± 3.041; unlabeled 16.32%; n = 13 from five fish; β-catenin 46.21% ± 2.728; Cx35.5 44.18% ± 1.671; unlabeled 9.61%; n = 13 CEs from seven fish). (C) Double labeling for Cx35.5 and GluR2 shows lack of proportionality, with Cx35.5 fluorescence occupying the majority of the CE contact area (GluR2 18.99% ± 1.601; Cx35.5 35.92% ± 2.087; unlabeled 45.10%; n = 10 CEs from five fish). ROI was defined in this case by the outline of GluR2 labeling. (D) Tree plot illustrating the area occupancy (fluorescence/contact area) for AJ (N-Cad = 38.1%), GJ (Cx35 = 34.8%), and glutamatergic (GluR2=18.9%) labeling at individual CEs (values normalized to ROI outlined by GluR2 labeling). The unlabeled area represents 8.1% of the contact’s surface. (E) The cartoon summarizes the synaptic components identified at a single CE contact. While chemical synapses are restricted to a small and peripheral area of the contact (presynaptic vesicles and release sites are represented in gray, postsynaptic receptor areas in magenta), most of its contact surface is occupied by multiple GJs (green) of variable size, which are interleaved and closely associated to AJs (red).
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The components of an electrical synapse as revealed by expansion microscopy of a single synaptic contact
eLife 13:e91931.
https://doi.org/10.7554/eLife.91931




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}