PKCδ is an activator of neuronal mitochondrial metabolism that mediates the spacing effect on memory consolidation
- Energy & Memory, Brain Plasticity Unit, CNRS, ESPCI Paris, PSL Research University, France
Figures
Figure 1 with 1 supplement
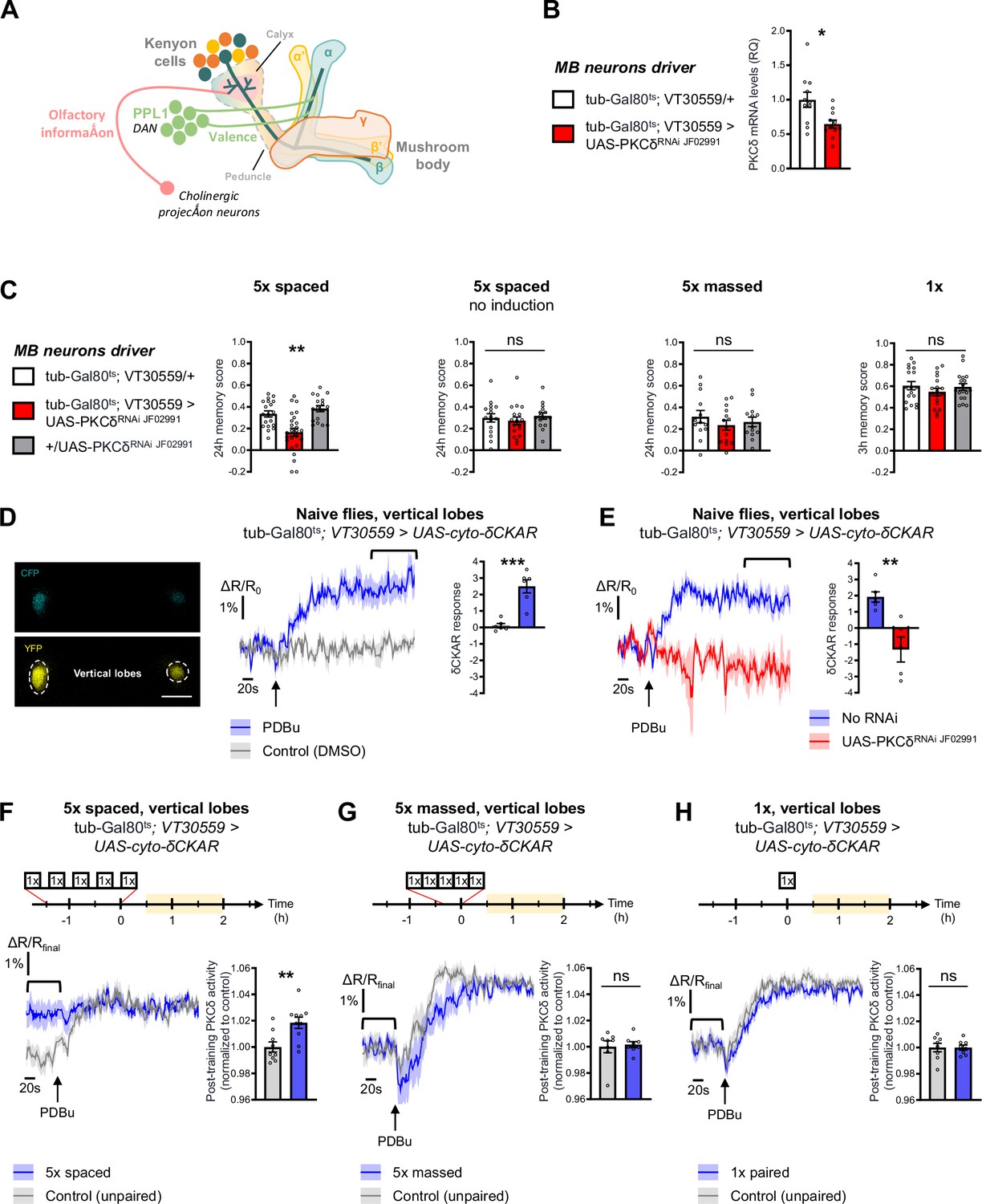
PKCδ is required in MB neurons for LTM formation.
(A) Schematic representation of Drosophila MB. The MB includes ~2000 intrinsic neurons per brain hemisphere. Their cell bodies are located in the dorsal posterior part of the brain. MB neurons each send a single neurite into the neuropil, which first traverses the calyx, a dendritic region where MB neurons receive olfactory input from projections neurons, and then extends into a long axonal branch. The bundled axons of MB neurons form a fascicle called the peduncle, which traverses the brain to the anterior part, where axons branch to form medial and vertical lobes according to three major branching patterns – α/β, α’/β’ and γ (Aso et al., 2014) – that define as many neuronal categories. The MB lobes receive input from dopaminergic neurons (the DANs), which signal stimuli of positive and negative valences in a region-specific manner (Aso et al., 2014). During associative learning, dopamine release on coincidentally odorant-activated MB output synapses modulates the synaptic drive to the network of MB output neurons, which bias subsequent odor-driven behavior (Heisenberg, 2003; Hige, 2018). Aversive LTM induced by spaced training is more specifically encoded within the α/β neurons (Pascual and Préat, 2001; Séjourné et al., 2011; Yu et al., 2006), and we previously showed that LTM retrieval involves the depression of an attraction-mediated pathway efferent from the MB vertical lobes (Aso et al., 2014; Bouzaiane et al., 2015; Dolan et al., 2018; Séjourné et al., 2011). However, according to another recent study, LTM retrieval mobilizes in parallel another MB output circuit efferent from the medial lobes (Jacob and Waddell, 2020). (B) Expression of PKCδ RNAi in adult MB neurons induced a significant reduction in the mRNA level of PKCδ measured by RT-qPCR in fly heads. Relative Quantification (RQ) was performed, indicating the foldchange of mRNA levels relative to the control genotype (n=11, t20=2.83, p=0.010). (C) PKCδ knockdown in adult MB neurons impaired memory after 5 x spaced conditioning (n=17–25, F2,58=12.59, p<0.0001). Without the induction of PKCδ RNAi expression, memory formed after 5 x spaced conditioning was normal (n=15–17, F2,45=0.41, p=0.67). Memory formed after 5 x massed training (n=13–14, F2,37=0.65, p=0.53) and 1 x training (n=18, F2,51=0.81, p=0.45) was normal in flies knocked down for PKCδ in adult MB neurons. (D) The cyto-δCKAR sensor was expressed in adult MB neurons and visualized in the CFP and YFP channels. Cytosolic PKCδ activity levels are recorded within the vertical lobes of the MBs (indicated with dashed line). Scale bar = 50 µm (valid for both channels). In naive flies, application of 250 µM of PDBu (black arrow), a pharmacological activator of PKCδ, resulted in the increase of the cyto-δCKAR response, reaching a plateau, as compared to the DMSO control (n=6, t10=5.66, p=0.0002). Quantification of the mean cyto-δCKAR response was performed 280 s after PDBu application on a time window of 560 s (black line). (E) In naive flies, application of 250 µM of PDBu (black arrow) resulted in an increase in the cyto-δCKAR response that is abolished when PKCδ is knocked down in adult MB neurons (n=5–6, t9=4.18, p=0.0024). Quantification of the mean cyto-δCKAR response was performed 280 s after PDBu application on a time window of 560 s (black line). (F) To compare post-conditioning cytosolic PKCδ activities (between 30 min and 2 hr post-conditioning, in yellow on the imaging time frame), cyto-δCKAR traces were normalized to the plateau value reached after addition of PDBu (saturation of the sensor), thus the activity level of cytosolic PKCδ is estimated as the cyto-δCKAR signal value before PDBu application. Cytosolic PKCδ activity is increased in the vertical lobes after 5 x spaced associative paired conditioning as compared to a non-associative spaced conditioning (unpaired) protocol (n=9–10, t17=3.18, p=0.0055). Quantification of the mean post-training PKCδ activity was performed on a time window of 120 s before PDBu application (black line). (G) After 5 x massed paired conditioning, cytosolic PKCδ activity was not changed as compared to 5 x massed unpaired conditioning (n=8, t14=0.33, p=0.75). (H) Similarly, after 1 x paired conditioning, cytosolic PKCδ activity was not changed as compared to 1 x unpaired conditioning (n=8, t14=0.0041, p=0.99). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by one-way ANOVA with post hoc testing by the Tukey pairwise comparisons test (C) or by unpaired two-sided t-test (B, D–H). Asterisks refer to the least significant p-value of post hoc comparison between the genotype of interest and the genotypic controls (C), or to the p-value of the unpaired t-test comparison (B, D–H) using the following nomenclature: *p<0.05, **p<0.01, ***p<0.001, ns: not significant, p>0.05. See also Figure 1—figure supplement 1 and Table 1.
-
Figure 1—source data 1
Source data displayed on Figure 1.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig1-data1-v1.xlsx
Figure 1—figure supplement 1
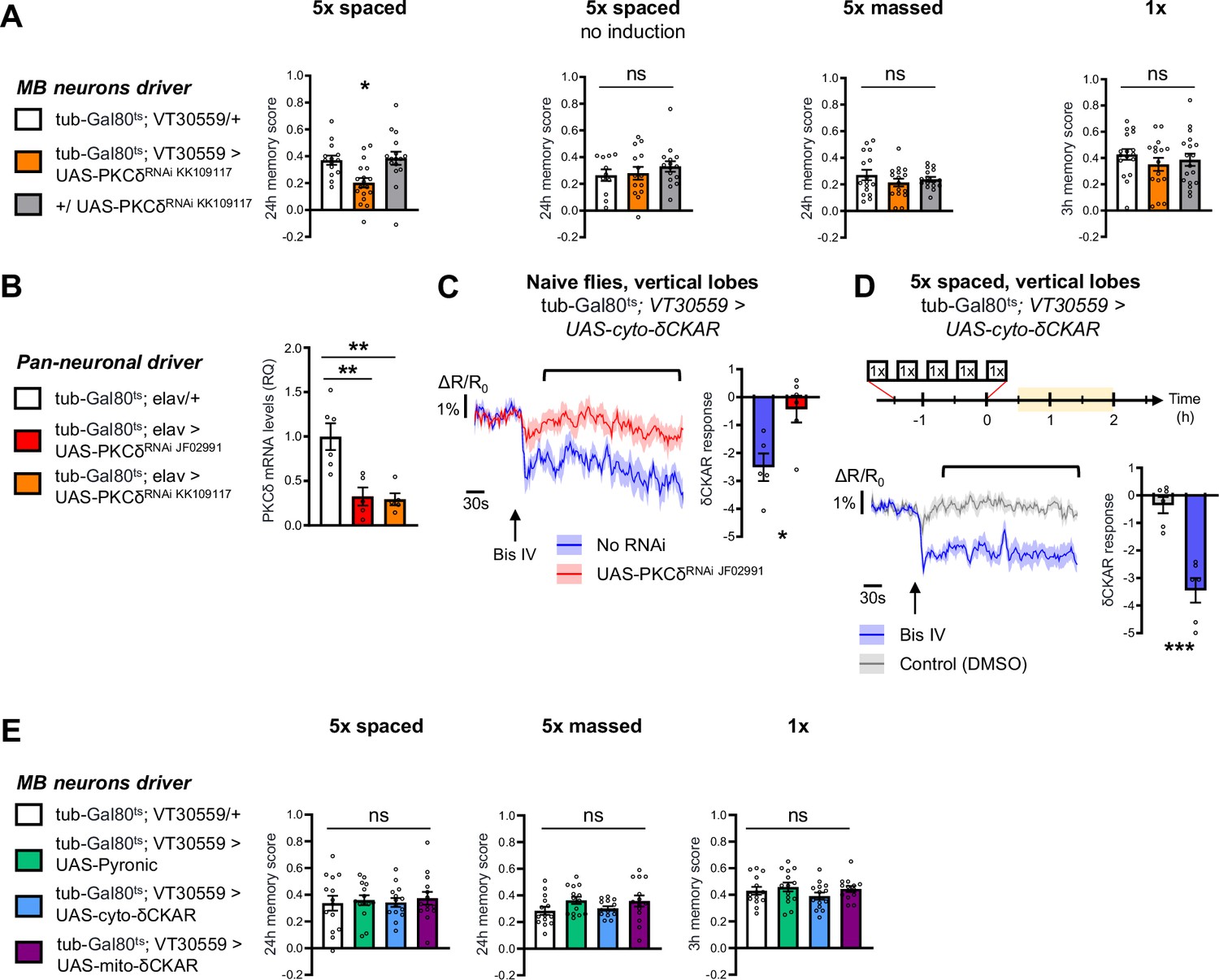
Control experiments for behavior analysis and δCKAR imaging experiments.
(A) Behavior analysis with a second non-overlapping PKCδ RNAi: 24 hr memory after 5 x spaced conditioning was specifically impaired after induction of the PKCδ RNAi expression in adult MB (n=14–18, F2,45=6.76, p=0.027), and it was not impacted without induction (n=11–15, F2,37=0.62, p=0.54). No memory defect was found after 5 x massed training (n=16, F2,45=0.90, p=0.42) or 1 x training (n=17–18, F2,48=0.66, P=0.52) in flies knocked down for PKCδ in adult MB. (B) Pan-neuronal expression of either PKCδ RNAi at the adult stage significantly reduced PKCδ mRNA levels. Relative Quantification (RQ) was performed, indicating the foldchange of mRNA levels relative to the control genotype (n=5–6, F2,13=11.85, p=0.0012). (C) In naive flies, application of 5 µM of Bis IV (black arrow), a PKC inhibitor, resulted in a decrease in the cyto-δCKAR response, and this response was abolished when PKCδ was knocked down (n=5–6, t9=3.01, p=0.015). Quantification of the mean cyto-δCKAR response was performed 20 s after Bis IV application on a time window of 250 s (black line). (D) After 5 x spaced training, Bis IV injection also decreased the cyto-δCKAR response as compared to the DMSO control (n=6, t10=4.83, p=0.0007). (E) Memory formed after either 5 x spaced (n=14, F3,52=0.14, p=0.94), or 5 x massed (n=14, F3,52=1.69, p=0.18), or 1 x training (n=14, F3,52=1.07, p=0.37) was not impaired by the expression in the MB neurons at adult stage of the imaging probes used in this study: Pyronic, cyto-δCKAR and mito-δCKAR. Data are expressed as mean ± SEM with dots as individual values, and were analyzed either by one-way ANOVA with post hoc testing by the Tukey pairwise comparisons test (A–B and E), or by an unpaired two-sided t-test (C–D). Asterisks refer to the least significant p-value of post hoc comparison between the genotype of interest and the genotypic controls (A–B, E) or to the p-value of the unpaired t-test comparison (C–D) using the following nomenclature: *p<0.05, **p<0.01, ***p<0.001, ns: not significant, p>0.05.
-
Figure 1—figure supplement 1—source data 1
Source data displayed on Figure 1—figure supplement 1.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig1-figsupp1-data1-v1.xlsx
Figure 2
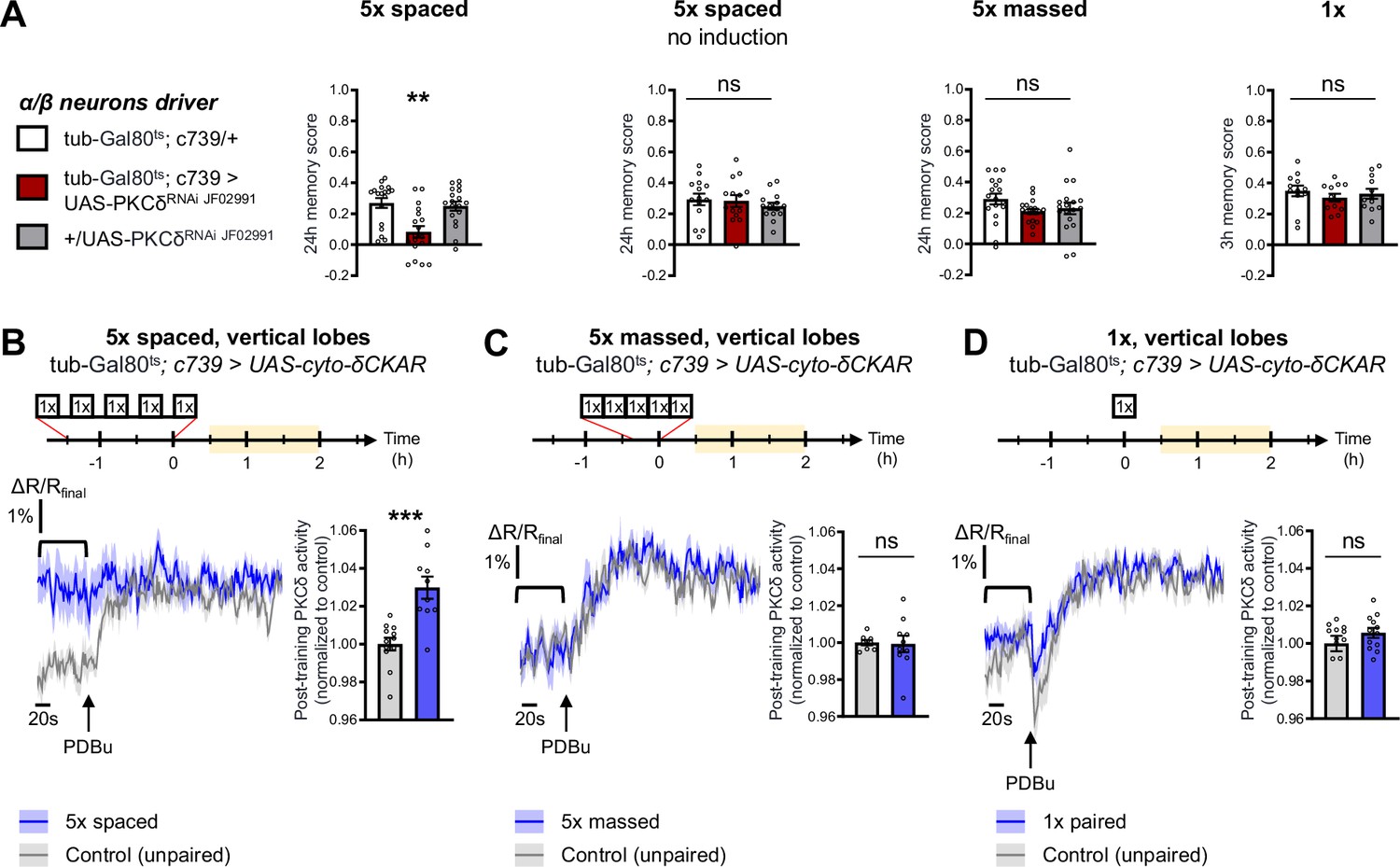
PKCδ is required in α/β neurons for LTM formation.
(A) When PKCδ was knocked down specifically in the α/β neurons of MBs, memory after 5 x spaced conditioning was impaired (n=18, F2,51=9.52, p=0.0003). Without PKCδ RNAi induction, memory after 5 x spaced conditioning was normal (n=14, F2,39=0.47, p=0.63). Memory was normal after 5 x massed training (n=18, F2,51=1.771, p=0.1805) as well as after 1 x training (n=12, F2,33=0.51, p=0.61) in flies expressing PKCδ RNAi in adult α/β MB neurons. (B) Using the same approach as detailed in Figure 1F, the post-training activity of cytosolic PKCδ was measured specifically in the α/β neurons, between 30 min and 2 hr post-conditioning (in yellow on the imaging time frame). After 5 x spaced paired conditioning, cytosolic PKCδ activity was increased in the α lobes as compared to unpaired conditioning (n=10–12, t20=4.58, p=0.0002). (C) Following 5 x massed conditioning, cytosolic PKCδ activity in the α lobes was not changed compared to unpaired conditioning (n=8–10, t16=0.13, p=0.90). (D) After 1 x paired conditioning, cytosolic PKCδ activity in the α lobes also remained unchanged compared to unpaired conditioning (n=12, t22=1.16, p=0.26). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by one-way ANOVA with post hoc testing by the Tukey pairwise comparisons test (A) or by unpaired two-sided t-test (B–D). Asterisks refer to the least significant P-value of a post hoc comparison between the genotype of interest and the genotypic controls (A) or to the p-value of the unpaired t-test comparison (B–D) using the following nomenclature: **p<0.01, p***<0.001, ns: not significant, p>0.05. See also Table 1.
-
Figure 2—source data 1
Source data displayed on Figure 2.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig2-data1-v1.xlsx
Figure 3 with 1 supplement
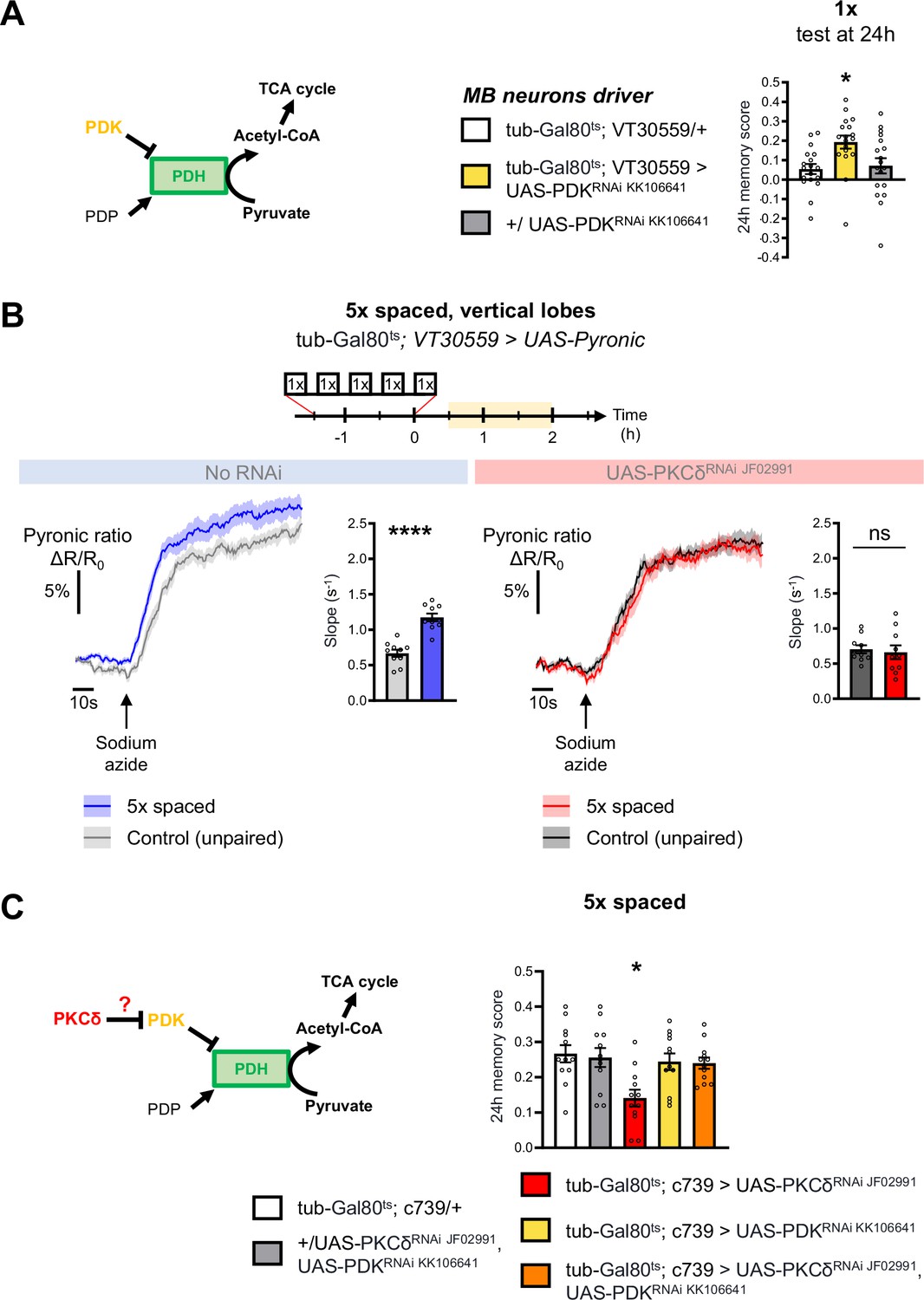
PKCδ regulates mitochondrial pyruvate metabolism upon LTM formation.
(A) Left panel: schema of the regulation of the pyruvate dehydrogenase (PDH) complex. In mitochondria, PDHC catalyzes the conversion of pyruvate to acetyl-CoA, which enters the tricarboxylic acid cycle (TCA). PDH can be inactivated through phosphorylation by PDK. In contrast, PDP can activate PDH via its dephosphorylation. Right panel: flies expressing an RNAi against PDK in MB neurons exclusively at the adult stage showed increased memory measured at 24 hr after a single cycle of training as compared to their genotypic controls (n=18, F2,51=5.09, p=0.0097). (B) The pyruvate sensor Pyronic was expressed in adult MB neurons and the pyruvate FRET signal was quantified in the vertical lobes. In control flies, spaced training elicited a faster pyruvate accumulation in axons of MB neurons after sodium azide application (5 mM; black arrow) as compared to non-associative unpaired training (left panel, slope measurement n=10, t18=6.751, p<0.0001). PKCδ knockdown in adult MB neurons impaired the spaced training induced increase in pyruvate accumulation in axons of MB neurons following sodium azide application (right panel, slope measurement n=10, t18=0.38, p=0.71). As for Figures 1 and 2, imaging was performed between 30 min and 2 hr post-conditioning, represented in yellow on the imaging time frame. (C) Left panel: schema of our model, asking whether PKCδ intervene upstream of the PDH complex. Here, we show that PKCδ regulates PDH activity via PDK inhibition. Right panel: after 5 x spaced conditioning, flies coexpressing the PDK and PKCδ RNAi in the α/β MB neurons at the adult stage exhibited normal memory formation as compared to genotypic controls. Flies solely expressing the PKCδ RNAi in α/β neurons at the adult stage exhibited the reported LTM defect, whereas flies that were only knocked down for PDK in adult α/β neurons formed normal memory (n=12, F4,55=4.75, p=0.0023). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by unpaired two-sided t-test (B) or by one-way ANOVA with post hoc testing by the Tukey pairwise comparisons test (A,C). Asterisks refer to the p-value of the unpaired t-test comparison or to the least significant p-value of post hoc comparison between the genotype of interest and the genotypic controls using the following nomenclature: *p<0.05, ****p<0.0001, ns: not significant, p>0.05. See also Figure 3—figure supplement 1.
-
Figure 3—source data 1
Source data displayed on Figure 3.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig3-data1-v1.xlsx
Figure 3—figure supplement 1
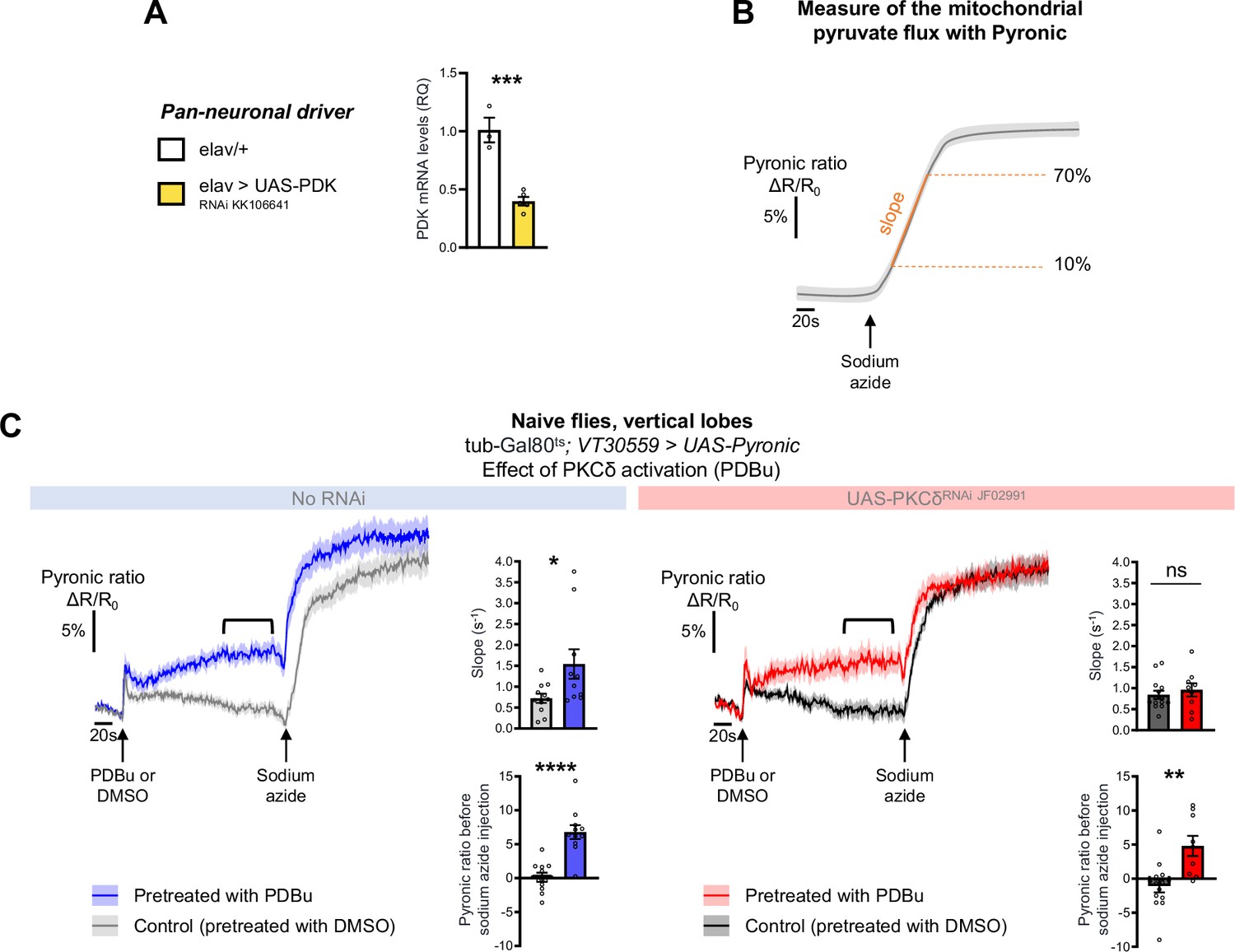
PKCδ activation by PDBu application increases the MB neuronal pyruvate flux.
(A) Pan-neuronal expression of PDK RNAi significantly reduced PDK mRNA levels, as compared to the control genotype (n=3–5, t6=6.66, p=0.0006). (B) Schema representing the measure of the mitochondrial pyruvate flux using the pyruvate FRET sensor Pyronic. Here, mitochondrial respiration is blocked by sodium azide at the beginning of the recording, thereby stopping pyruvate mitochondrial uptake. The expected kinetic of pyruvate accumulation following the azide treatment is represented in grey, showing that pyruvate accumulates from the arrest of its mitochondrial uptake until saturation of the sensor. The rate of pyruvate accumulation (i.e. the slope of the measured kinetic, measured between 10 and 70% of the plateau) reflects the rate at which pyruvate was consumed by mitochondria for energy production before their blockade. (C) The Pyronic probe was expressed in adult MB neurons and the pyruvate FRET signal was quantified in the vertical lobes. In naive control flies, PDBu pretreatment (250 µM, 3 min) elicited a faster pyruvate accumulation following sodium azide application (5 mM, black arrows) as compared to flies pretreated with the DMSO solvent alone (left panel, slope measurement n=10–11, t19=2.31, p=0.032). This PDBu pretreatment induced increase in pyruvate accumulation is abolished when PKCδ is knocked down in MB neurons as compared to DMSO pretreatment (right panel, slope measurement n=9–14, t21=0.67, p=0.51). Notably, in control flies, PDBu injection is followed by a progressive increase in the measured Pyronic ratio (black line) as compared to DMSO injection (n=11, t20=5.44, p<0.0001). This effect is not sensitive to PKCδ knockdown (n=9–14, t21=3.54, p=0.0019). As PDBu is known to activate other PKCs (Wu-Zhang et al., 2012), this observation likely reflects a PDBu off-target effect independent of PKCδ on pyruvate neuronal levels. Data are expressed as mean ± SEM with dots as individual values, and were analyzed by unpaired two-sided t-test. Asterisks refer to the p-value of the unpaired t-test comparison using the following nomenclature: *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, ns: not significant, p>0.05.
-
Figure 3—figure supplement 1—source data 1
Source data displayed on Figure 3—figure supplement 1.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig3-figsupp1-data1-v1.xlsx
Figure 4 with 1 supplement
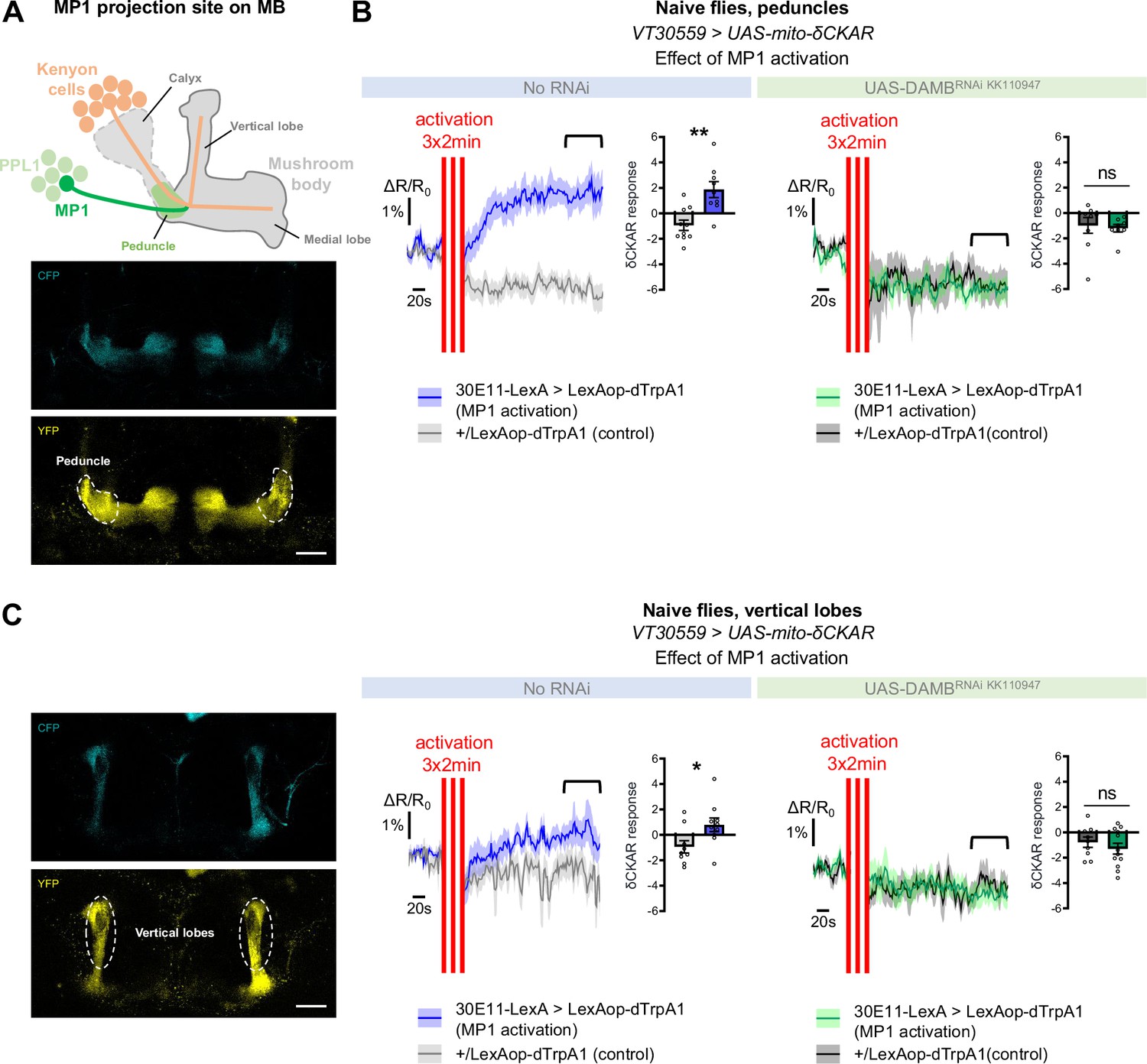
MP1 neurons control PKCδ activity in the MB neurons via DAMB signaling.
(A) Top panel: Schema of the MB structure and its afferent MP1 neuron. The dopaminergic neuron MP1 (depicted in green) in the PPL1 cluster projects onto the peduncle area of the MBs. Lower panel: The mito-δCKAR sensor was expressed in adult MB neurons and visualized in the CFP and YFP channels. The region of recording for panel B is at the level of the MB peduncles (dashed line). Scale bar = 30 µm (valid for both channels). (B) Naive flies expressing the mito-δCKAR sensor in MB neurons together with the dTrpA1 (a heat-sensitive cation channel) in MP1 neurons (30E11-LexA driver) were subjected to a thermal treatment consisting of three 2 min periods at 30 °C separated by 2 min (red vertical lines); control flies expressed the mito-δCKAR in MB neurons but not the dTrpA1 channel (no 30E11-LexA driver). Mitochondrial PKCδ activity was recorded before (baseline) and immediately after the activation periods. Quantification of the mean mito-δCKAR response was performed 120 s after the last cycle of thermal activation on a time window of 480 s (black line). In naive flies expressing the dTrpA1 channel in MP1 neurons, activation of MP1 increased mitochondrial PKCδ activity as compared to control flies (n=9–10, t17=3.83, p=0.0013). When DAMB was knocked down in MB neurons, MP1 activation failed to increase mitochondrial PKCδ activity as compared to control flies (n=8–9, t15=0.31, p=0.76). (C) The mito-δCKAR sensor was expressed in adult MB neurons and mitochondrial PKCδ activity was recorded in the vertical lobes (dashed line). As in the peduncle region, in naive flies expressing the dTrpA1 channel in MP1 neurons activation of MP1 increased mitochondrial PKCδ activity in the vertical lobes as compared to control flies (n=9–10, t17=2.37, p=0.030). DAMB knock-down in the MB neurons also prevented any increase of PKCδ mitochondrial activity in the vertical lobes upon the thermogenic activation of MP1 neurons, as compared to the genotypic control (n=9–12, t19=0.83, p=0.42). Data are expressed as mean ± SEM with dots as individual values and were analyzed by unpaired two-sided t-test. Asterisks refer to the p-value of the unpaired t-test comparison using the following nomenclature: *p<0.05, **p<0.01, ns: not significant, p>0.05. Scale bar = 30 µm (valid for both channels). See also Figure 4—figure supplement 1 and Video 1.
-
Figure 4—source data 1
Source data displayed on Figure 4.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig4-data1-v1.xlsx
Figure 4—figure supplement 1
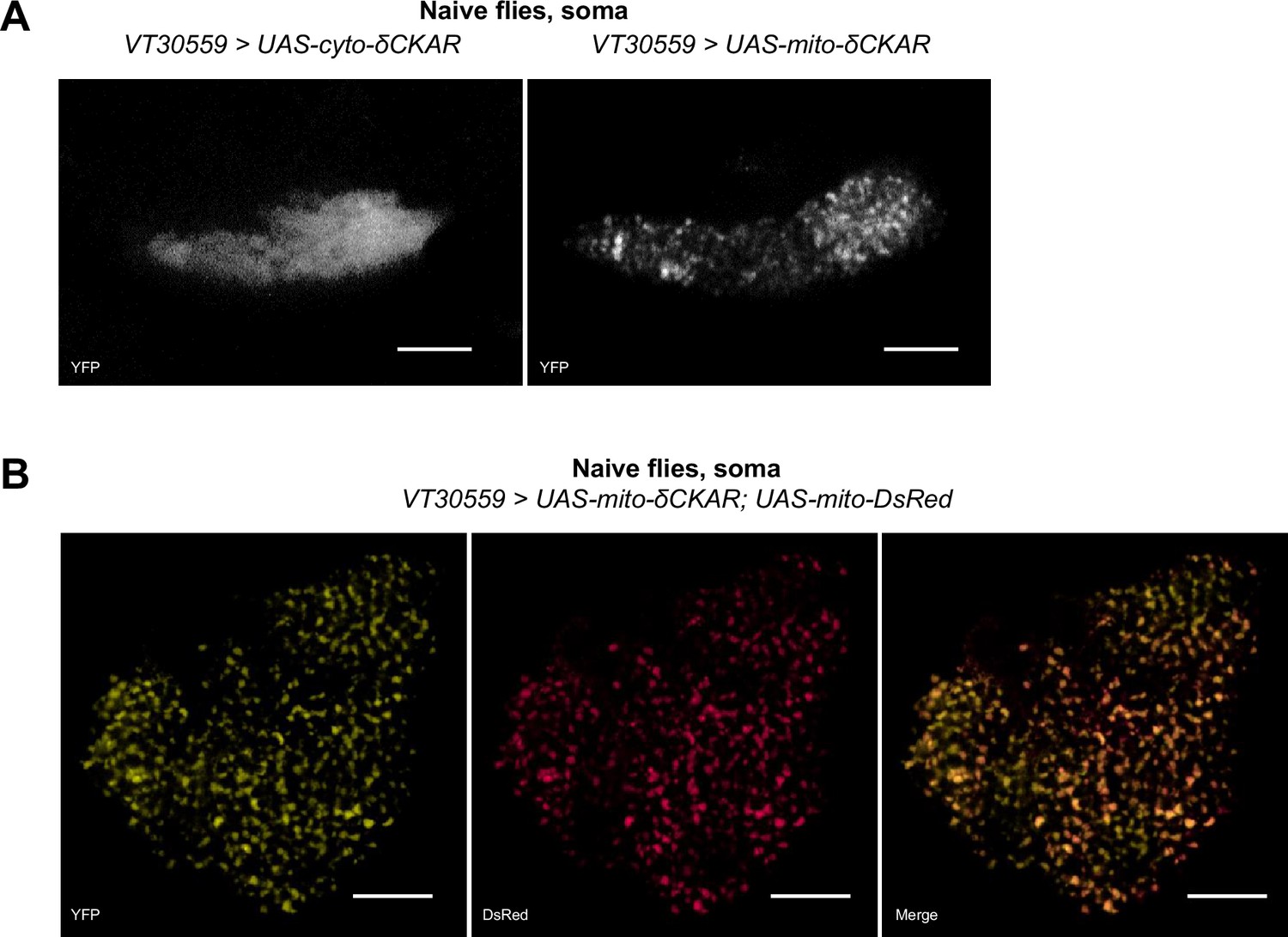
Subcellular addressing of the cyto-δCKAR and mito-δCKAR probes.
(A) Comparison of fluorescence sub localization in the soma of MB neurons in flies expressing either cyto-δCKAR or mito-δCKAR under the control of the VT30559 driver. Mito-δCKAR punctate signal, suggests a mitochondrial localization of the probe, in contrast to the diffuse signal observed with cyto-δCKAR. This comparison was done in the soma area of the MBs because the density of mitochondria in the lobes is too important to allow their discrimination. Images were acquired by two-photon microscopy and the cyto-and mito-δCKAR sensors were visualized through the YFP channel. (B) Coexpression of mito-δCKAR and mito-DsRed (DsRed is addressed to mitochondria using the COX8 targeting sequence Lutas et al., 2012) in the MB neurons shows a clear colocalization of the YFP and DsRed signals. The image acquired by confocal microscopy shows a single plan located in the middle of a MB soma of a representative fly. Scale bars = 20 µm. See full stack on Video 1.
Figure 5 with 1 supplement
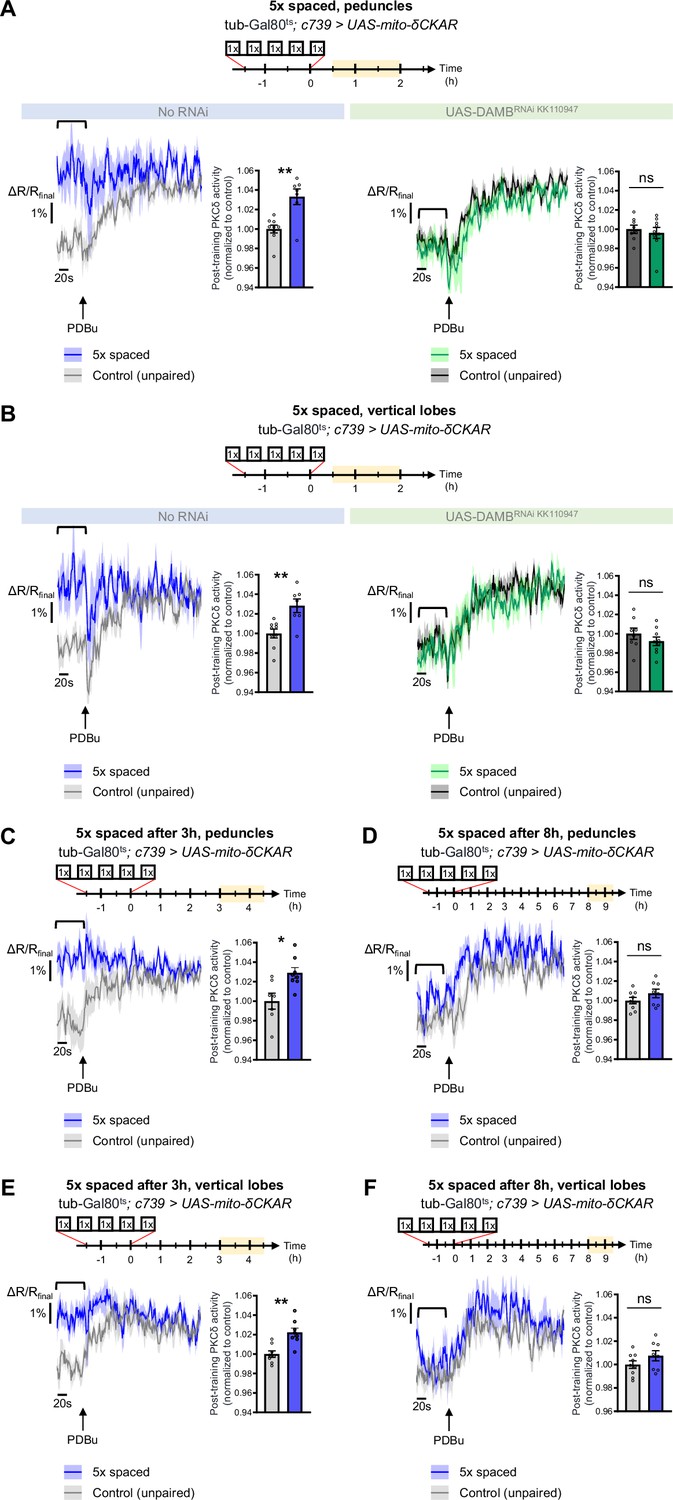
PKCδ translocates to the mitochondria of α/β neurons upon LTM formation.
The mito-δCKAR sensor was expressed in adult α/β neurons and the post-training activity of mitochondrial PKCδ was measured in the peduncle (A) or vertical lobes (B) in the same flies, between 30 min and 2 hr post-conditioning (in yellow on the imaging time frame). The same approach was used as in Figure 1F–H with PDBu application to reach saturation level of the sensor and quantification of the mean post-training PKCδ activity performed on a time-window of 120 s before PDBu application (black line). (A) After 5 x spaced paired conditioning, mitochondrial PKCδ activity in the peduncle was increased as compared to unpaired conditioning (left panel, n=7–9, t14=3.56, p=0.0032). When DAMB was knocked down in adult α/β neurons, mitochondrial PKCδ activity was not changed after 5 x spaced paired conditioning as compared to unpaired conditioning (right panel, n=8–9, t15=1.09, p=0.29). (B) At the level of the vertical lobes, after 5 x spaced paired conditioning, mitochondrial PKCδ activity was increased as compared to unpaired conditioning (left panel n=7–9, t14=3.93, p=0.0015). When DAMB was knocked down in adult α/β neurons, mitochondrial PKCδ activity was not changed after 5 x spaced paired conditioning, as compared to unpaired conditioning (right panel n=8–9, t15=0.52, p=0.61). (C) The post-training activity of mitochondrial PKCδ was measured at the level of the peduncle, between 3 hr and 4 hr 30 min after 5 x spaced conditioning. At that timepoint, PKCδ mitochondrial activity was still increased as compared to 5 x spaced unpaired conditioning (n=7–8, t14=3.01, p=0.010). (D) However, 8 h to 9 h 30 min after 5 x spaced conditioning, PKCδ mitochondrial activity in the peduncle was not significantly different from its 5 x spaced unpaired control (n=8–9, t15=1.02, p=0.33). (E) Similarly, in the α lobe, between 3 hr and 4 hr 30 min after 5 x spaced conditioning, PKCδ mitochondrial activity was still increased as compared to 5 x spaced unpaired conditioning (n=8, t14=3.99, p=0.0014), whereas (F) 8 hr to 9 hr 30 min after 5 x spaced conditioning, PKCδ mitochondrial activity in the α lobe was not significantly different from its 5 x spaced unpaired control (n=8–9, t15=1.40, p=0.18). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by unpaired two-sided t-test. Asterisks refer to the p-value of the unpaired t-test comparison using the following nomenclature: *p<0.05, **p<0.01, ns: not significant, p>0.05. See also Figure 5—figure supplement 1.
-
Figure 5—source data 1
Source data displayed on Figure 5.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig5-data1-v1.xlsx
Figure 5—figure supplement 1
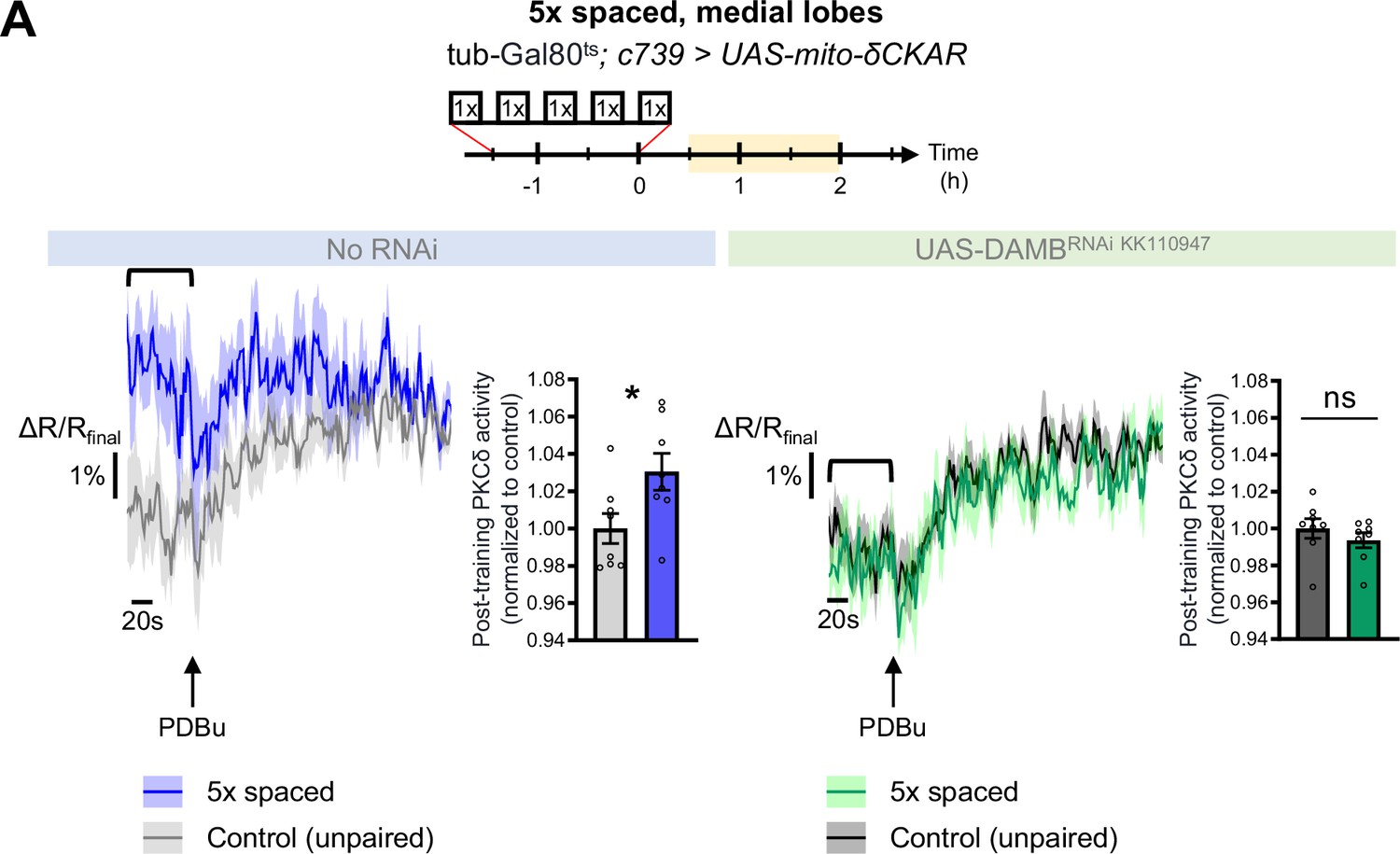
PKCδ also translocates to the mitochondria of the β lobe upon LTM formation.
(A) The post-training activity of mitochondrial PKCδ was measured in the medial lobes between 30 min and 2 hr post-conditioning (in yellow on the imaging time frame), in the same flies as in Figure 5A–B. PKCδ activity was increased in the medial lobes after 5 x spaced conditioning as compared to unpaired conditioning (left panel n=8, t14=2.38, p=0.032). On the other hand, PKCδ activity was unchanged in the medial lobes after 5 x spaced conditioning upon the knock-down of DAMB in adult α/β neurons, as compared to unpaired conditioning (right panel n=8, t14=0.95, p=0.36). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by unpaired two-sided t-test. Asterisks refer to the p-value of the unpaired t-test comparison using the following nomenclature: *p<0.05, ns: not significant, p>0.05.
-
Figure 5—figure supplement 1—source data 1
Source data displayed on Figure 5—figure supplement 1.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig5-figsupp1-data1-v1.xlsx
Figure 6 with 1 supplement
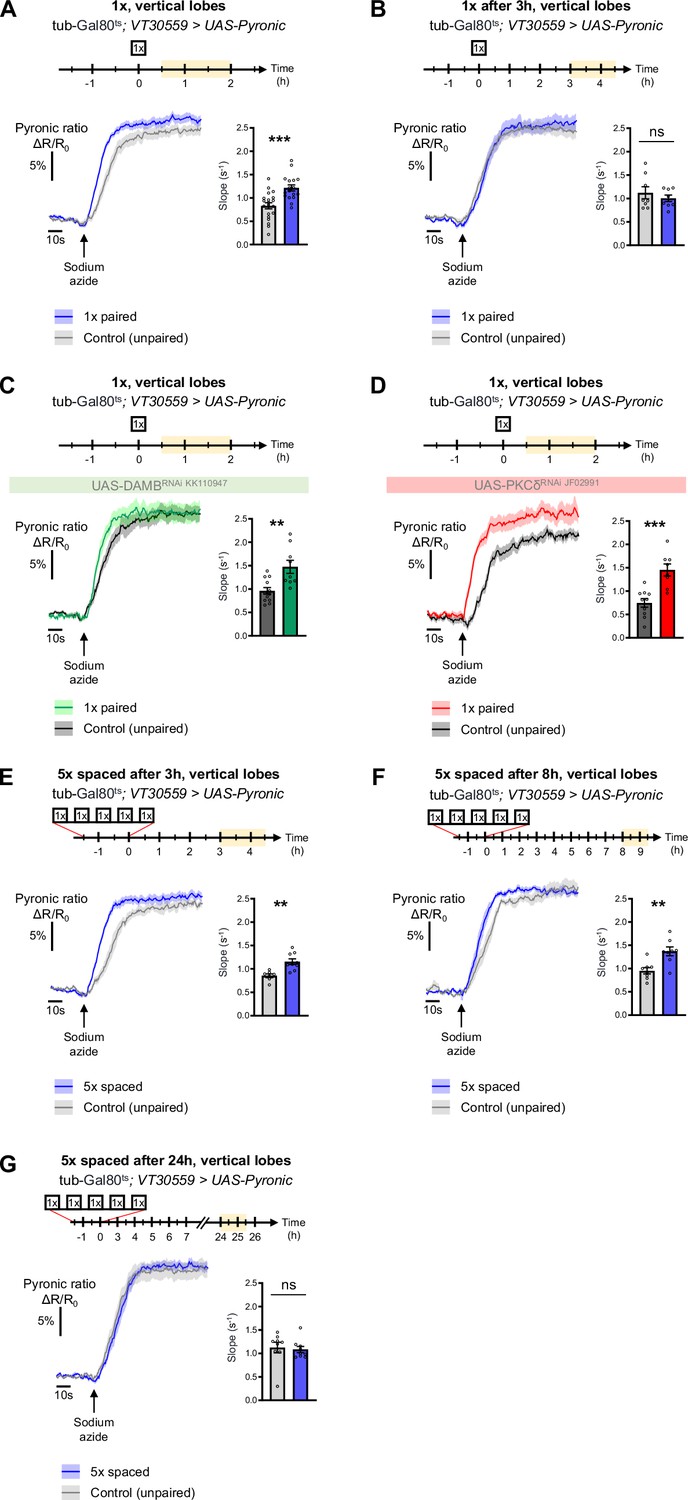
The DAMB/PKCδ axis is specific to metabolic activation during LTM formation.
The pyruvate sensor Pyronic was expressed in adult MB neurons and pyruvate accumulation was measured after conditioning in the vertical lobes. For each panel, the period of imaging after conditioning is indicated in yellow on the imaging time frame. (A) 30 min to 2 hr after 1 x paired conditioning, an increased pyruvate flux was measured in control flies at the level of the vertical lobes, as compared to non-associative unpaired 1 x training (slope measurement n=17–20, t35=4.08, p=0.0003). Here, we pooled the data obtained from control flies acquired in parallel to DAMB and PKCδ knockdown flies (panels C and D). (B) 3 hr to 4 hr 30 min after 1 x paired conditioning of control flies the rate of pyruvate accumulation was similar in the vertical lobes as compared to unpaired conditioning (slope measurement n=8, t14=0.80, p=0.44). (C) 30 min to 2 hr after 1 x paired conditioning, the pyruvate flux was still increased when DAMB was knocked down in the MBs at the adult stage (slope measurement n=9–12, t19=3.53, p=0.0022). (D) Similarly, PKCδ knockdown in adult MB neurons did not impair the 1 x conditioning induced increase in pyruvate accumulation in MB neuron axons (slope measurement n=8–10, t16=4.57, p=0.0003). (E) 3 hr to 4 hr 30 min after the last cycle of 5 x spaced conditioning of control flies, the pyruvate flux was increased in the vertical lobes as compared to 5 x spaced unpaired conditioning (slope measurement n=9–7, t14=3.89, p=0.0016). (F) 8 hr to 9 hr 30 min after the last cycle of 5 x spaced conditioning of control flies, the pyruvate flux was still increased in the vertical lobes as compared to 5 x spaced unpaired conditioning (slope measurement n=8, t14=3.63, p=0.0027). (G) 24 hr after the last cycle of 5 x spaced conditioning of control flies, the pyruvate flux was similar as compared to 5 x spaced unpaired conditioning at the level of the vertical lobes (slope measurement n=9, t16=0.33, p=0.75). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by unpaired two-sided t-test. Asterisks refer to the p-value of the unpaired t-test comparison using the following nomenclature: **p<0.01, ***p<0.001, ns: not significant, p>0.05. See also Figure 6—figure supplement 1.
-
Figure 6—source data 1
Source data displayed on Figure 6.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig6-data1-v1.xlsx
Figure 6—figure supplement 1
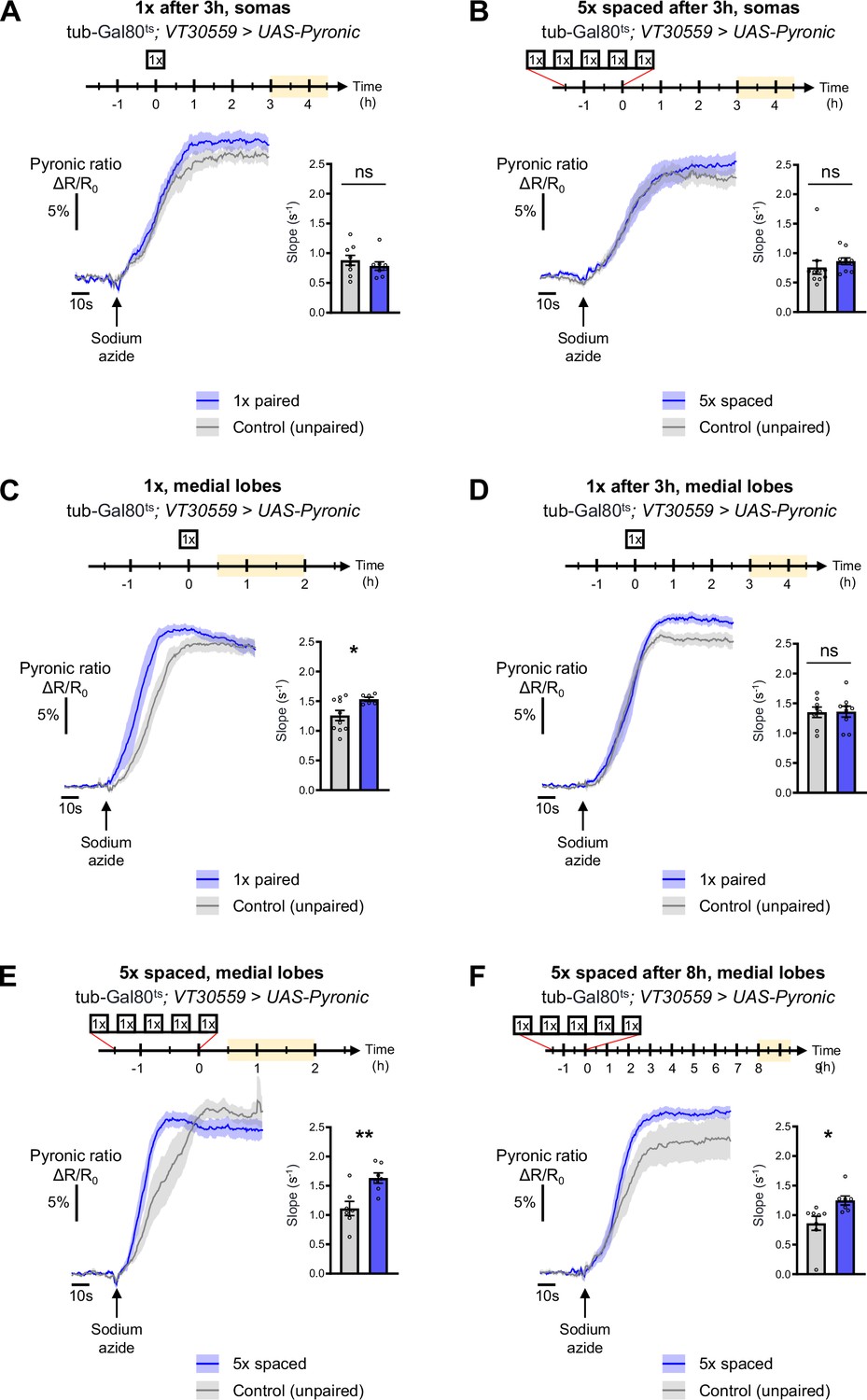
Additional characterization of the temporal dynamic of the pyruvate flux following conditioning in the somas and medial lobes.
(A) At the level of the MB somas, 3 hr to 4 hr 30 min after 1 x conditioning, the pyruvate flux of control flies was not increased as compared to 5 x spaced unpaired conditioning (slope measurement n=8–9, t15=0.85, p=0.41). (B) 3 hr to 4 hr 30 min after 5 x spaced conditioning, the pyruvate flux of control flies was not increased as compared to 5 x spaced unpaired conditioning at the level of the MB somas (slope measurement n=10, t18=0.81, p=0.43). (C) 30 min to 2 hr after 1 x paired conditioning, an increased pyruvate flux was measured in control flies at the level of the medial lobes as compared to unpaired 1 x training (slope measurement n=6–10, t14=2.37, p=0.033). (D) 3 hr to 4 hr 30 min after 1 x paired conditioning of control flies the rate of pyruvate accumulation in the medial lobes was similar as compared to unpaired conditioning (slope measurement n=8–9, t15=0.081, p=0.94). (E) 30 min to 2 hr after 5 x spaced conditioning, the pyruvate flux of control flies was increased in the medial lobes as compared to 5 x spaced unpaired conditioning (slope measurement n=7, t12=3.46, p=0.0047). (F) 8 hr to 9 hr 30 min after the last cycle of 5 x spaced conditioning of control flies, the pyruvate flux was still increased in the medial lobes as compared to 5 x spaced unpaired conditioning (slope measurement n=7–8, t13=2.62, p=0.021). Data are expressed as mean ± SEM with dots as individual values, and were analyzed by unpaired two-sided t-test. Asterisks refer to the p-value of the unpaired t-test comparison using the following nomenclature: *p<0.05, **p<0.01, ns: not significant, p>0.05.
-
Figure 6—figure supplement 1—source data 1
Source data displayed on Figure 6—figure supplement 1.
- https://cdn.elifesciences.org/articles/92085/elife-92085-fig6-figsupp1-data1-v1.xlsx
Figure 7
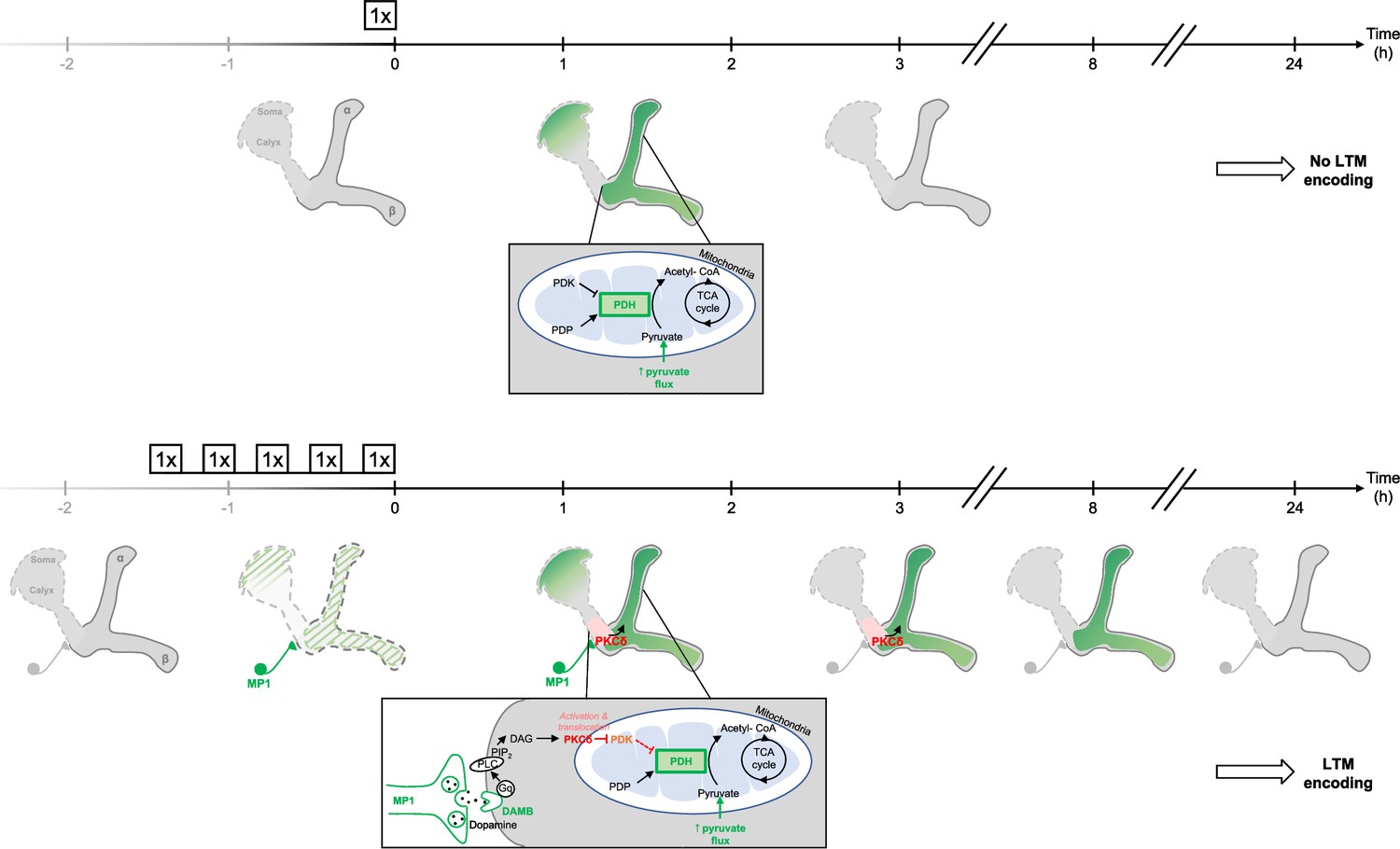
Schema of the energy-based gating of LTM, illustrating the role of PKCδ downstream of the DAMB signaling cascade.
After 1 x training, the pyruvate flux to the mitochondria in the MB neurons somas, the vertical lobes (Rabah et al., 2023), and in the medial lobes is increased. This increased energy state is not maintained, and after 3 hr, the MBs are back to their basal energy state. Upon spaced training, in addition to the mechanism occurring after 1 x training, the enhanced oscillatory activity of MP1 neurons activates DAMB signaling in the efferent α/β MB neurons. The DAMB receptor preferentially couples with Gq and the produced DAG activates PKCδ, which results in its translocation to mitochondria. There, PKCδ can activate the PDH, by releasing the inhibition exerted by PDK. The enhanced PDH activity promotes an increase in mitochondrial pyruvate flux, and this enhanced energy state gates memory consolidation, thereby enabling LTM formation. After 3 hr, the lasting effect of activated PKCδ on mitochondrial metabolism maintains a high pyruvate flux in the vertical and medial lobes, while it comes back to its baseline levels in the somas. After 8 hr, whereas PKCδ activation has ended, the high energy state of the MBs is still maintained, which ultimately results in the gating of LTM. At 24 hr, once LTM has been encoded and can be tested by behavioral assays, the MB neurons mitochondrial metabolism is back to its basal state, indicating that the upregulation of pyruvate metabolism does not represents the memory trace in itself.
Videos
Video 1
Video of mito-δCKAR and mito-DsRed colocalization z-stack.
Video of the full z-stack composed of 28 images showing the entire MB soma of the representative fly shown on Figure 4—figure supplement 1B. The YFP channel is on the left, DsRed is in the middle, and the merged channels are shown on the right. Scale bar = 20 µm.
Tables
Table 1
Sensory acuity controls in PKCδ knockdown flies.
Control experiments for olfactory acuity and electric shock avoidance: the expression of either of the two PKCδ RNAi used in this study, in MB neurons at the adult stage had no significant effect on olfactory acuity, or on the avoidance of electric shocks.
| Genotypes | Shock avoidance | Naive odor avoidance | ||||
|---|---|---|---|---|---|---|
| Octanol | Methylcyclohexanol | |||||
| Mean ± s.e.m. | Statistics | Mean ± s.e.m. | Statistics | Mean ± s.e.m. | Statistics | |
| tubGal80ts;VT30559/+ | 0.63±0.057 | F2,33=2.60; p=0.09; n=12 | 0.74±0.053 | F2,27=5.60; p=0.0092*; n=9–11 | 0.72±0.069 | F2,27=1.27; p=0.30; n=9–11 |
| tubGal80ts;VT30559 >UAS-PKCδRNAi JF02991 | 0.76±0.033 | 0.53±0.070 | 0.60±0.082 | |||
| UAS-PKCδRNAi JF02991/+ | 0.64±0.039 | 0.44±0.073 | 0.56±0.069 | |||
| tubGal80ts;VT30559/+ | 0.47±0.052 | F2,33=0.38; p=0.69; n=12 | 0.63±0.040 | F2,33=2.76; p=0.078; n=12 | 0.66±0.046 | F2,33=0.64; p=0.53; n=12 |
| tubGal80ts;VT30559 >UAS-PKCδRNAi KK109117 | 0.42±0.039 | 0.70±0.033 | 0.67±0.034 | |||
| UAS-PKCδRNAi KK109117/+ | 0.43±0.047 | 0.57±0.047 | 0.61±0.050 | |||
| tubGal80ts,c739/+ | 0.45±0.060 | F2,39=0.61; p=0.55; n=14 | 0.58±0.050 | F2,33=2.48; p=0.099; n=12 | 0.46±0.044 | F2,33=0.94; p=0.40; n=12 |
| tubGal80ts,c739 >UAS-PKCδRNAi JF02991 | 0.40±0.053 | 0.45±0.036 | 0.47±0.051 | |||
| UAS-PKCδRNAi JF02991/+ | 0.49±0.059 | 0.48±0.045 | 0.54±0.048 | |||
-
*
Tukey post hoc comparison between the genotype of interest and controls are not significant: tubGal80ts;VT30559 >UAS-PKCδRNAi JF02991 vs tubGal80ts;VT30559/+: ns tubGal80ts;VT30559 >UAS-PKCδRNAi JF02991 vs UAS-PKCδRNAi JF02991/+: ns tubGal80ts;VT30559/+vs UAS-PKCδRNAi JF02991/+: **.
-
Table 1—source data 1
Source data displayed on Table 1.
- https://cdn.elifesciences.org/articles/92085/elife-92085-table1-data1-v1.xlsx
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (D. melanogaster) | tubulin-Gal80ts;VT30559-Gal4 | Plaçais et al., 2017 | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | tubulin-Gal80ts;c739-Gal4 | Turrel et al., 2018 | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | tubulin-Gal80ts;elav-Gal4 | Silva et al., 2022 | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | 30E11-LexA | Bloomington Drosophila Stock Center | BDSC:54209; FLYB: FBst0054209; RRID:BDSC_54209 | FlyBase symbol: P{GMR30E11-lexA} |
| Genetic reagent (D. melanogaster) | UAS-PKCδRNAi JF02991 | Bloomington Drosophila Stock Center | BDSC:28355; FLYB: FBst0028355; RRID:BDSC_28355 | FlyBase symbol: P{TRiP.JF02991} |
| Genetic reagent (D. melanogaster) | UAS-PKCδRNAi KK109117 | Vienna Drosophila Resource Center | VDRC:101421; FLYB: FBti0121612; RRID:VDRC_101421 | FlyBase symbol: P{KK109117} |
| Genetic reagent (D. melanogaster) | UAS-DAMBRNAi KK110947 | Vienna Drosophila Resource Center | VDRC:105324; FLYB: FBst0477151; RRID:VDRC_105324 | FlyBase symbol: P{KK110947} |
| Genetic reagent (D. melanogaster) | UAS-PDKRNAi KK106641 | Vienna Drosophila Resource Center | VDRC:106641; FLYB: FBst0478465; RRID:VDRC_106641 | FlyBase symbol: P{KK107950} |
| Genetic reagent (D. melanogaster) | LexAop-dTrpA1 | Liu et al., 2012 | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | tubulin-Gal80ts;VT30559-Gal4, UAS-Pyronic | Plaçais et al., 2017 | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | 30E11-LexA;VT30559-Gal4 | Plaçais et al., 2017 | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | UAS-cyto-δCKAR | This paper | N/A | Available upon request; see “generation of transgenic flies” in Material and methods |
| Genetic reagent (D. melanogaster) | UAS-mito-δCKAR | This paper | N/A | Available upon request; see “generation of transgenic flies” in Material and methods |
| Genetic reagent (D. melanogaster) | UAS-cyto-δCKAR;UAS-PKCδRNAi JF01991 | This paper | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | UAS-mito-δCKAR;UAS-DAMBRNAi KK110947 | This paper | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | UAS-mito-δCKAR;UAS-DAMBRNAi KK110947;LexAop-dTrpA1 | This paper | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | UAS-PDKRNAi KK106641;UAS-PKCδRNAi JF02991 | This paper | N/A | Available upon request |
| Genetic reagent (D. melanogaster) | UAS-mito-DsRed | Bloomington Drosophila Stock Center | BDSC:93056; FLYB: FBst0093056; RRID:BDSC_93056 | FlyBase symbol: P{UAS-DsRed.mito} |
| Genetic reagent (D. melanogaster) | UAS-mito-δCKAR;UAS-mito-DsRed | This paper | N/A | Available upon request |
| Recombinant DNA reagent | pcDNA3-deltaCKAR | Addgene | #31526 RRID:Addgene_31526 | |
| Recombinant DNA reagent | pJFRC-MUH | Addgene | #26213 RRID:Addgene_26213 | |
| Recombinant DNA reagent | pJRFC-MUH-UAS-deltaCKAR | This paper | N/A | cyto-δCKAR construct used to generate the UAS- cyto-δCKAR Drosophila line |
| Recombinant DNA reagent | pJRFC-MUH-UAS-mitodelta CKAR | This paper | N/A | mito-δCKAR construct used to generate the UAS- mito-δCKAR Drosophila line |
| Sequence-based reagent | Primer PKCδ forward | This paper, DRSC Fly Primer Bank | PCR Primer Pair PP14953 | 5’-GGCACCAAACACCCGTATCT-3’ |
| Sequence-based reagent | Primer PKCδ reverse | This paper, DRSC Fly Primer Bank | PCR Primer Pair PP14953 | 5’-CCCATAGAATCTGGCTCGCT-3’ |
| Sequence-based reagent | Primer PDK forward | This paper, DRSC Fly Primer Bank | PCR Primer Pair PP14510 | 5’-CCTCGCCCCTCTCGATAAAG-3’ |
| Sequence-based reagent | Primer PDK reverse | This paper, DRSC Fly Primer Bank | PCR Primer Pair PP14510 | 5’-TCGAACAGGCAGTTCCTTGC-3’ |
| Sequence-based reagent | Primer tub forward | Turrel et al., 2018 | N/A | 5’-TTGTCGCGTGTGAAACACTTC-3’ |
| Sequence-based reagent | Primer tub reverse | Turrel et al., 2018 | N/A | 5’-CTGGACACCAGCCTGACCAAC-3’ |
| Commercial assay or kit | RNeasy Plant Mini Kit | QIAGEN | Cat. #74904 | |
| Commercial assay or kit | RNA MinElute Cleanup Kit | QIAGEN | Cat. #74204 | |
| Commercial assay or kit | SuperScript III First-Strand Kit | Thermofisher Invitrogen | Cat. #18080–051 | |
| Commercial assay or kit | SYBR Green I Master mix | Roche | Cat. # 04729692001 | |
| Chemical compound, drug | 3-octanol (99%) | Sigma-Aldrich | Cat. #153095 | |
| Chemical compound, drug | 4-methylcyclohexanol (98%) | Sigma-Aldrich | Cat. #218405 | |
| Chemical compound, drug | Paraffine GPR Rectapur | VWR | Cat. #24679.360 | |
| Chemical compound, drug | NaCl | Sigma-Aldrich | Cat. #S9625 | |
| Chemical compound, drug | KCl | Sigma-Aldrich | Cat. #P3911 | |
| Chemical compound, drug | MgCl2 | Sigma-Aldrich | Cat. #M9272 | |
| Chemical compound, drug | CaCl2 | Sigma-Aldrich | Cat. #C3881 | |
| Chemical compound, drug | D-trehalose | Sigma-Aldrich | Cat. #9531 | |
| Chemical compound, drug | Sucrose | Sigma-Aldrich | Cat. #S9378 | |
| Chemical compound, drug | HEPES-NaOH | Sigma-Aldrich | Cat. #H7637 | |
| Chemical compound, drug | Phorbol 12,13-dibutyrate (PDBu) | Tocris | Cat. #4153 | |
| Chemical compound, drug | Bisindolylmaleimide IV (Bis IV) | Sigma-Aldrich | Cat. #B3306 | |
| Chemical compound, drug | Sodium Azide | Sigma-Aldrich | Cat. #71289 | |
| Chemical compound, drug | Phosphate Buffered Saline (PBS) | Sigma-Aldrich | Cat. #P4417 | |
| Chemical compound, drug | Paraformaldehyde 16% | Life technologies | Cat. #P36965 | |
| Software, algorithm | Prism 8 | GraphPad software, v8.4.3 | RRID:SCR_002798 | |
| Software, algorithm | Fiji | ImageJ 1.52 p | RRID:SCR_002285 | |
| Software, algorithm | Affinity Photo | Affinity Photo software, v1.10.5 | RRID:SCR_016951 |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
PKCδ is an activator of neuronal mitochondrial metabolism that mediates the spacing effect on memory consolidation
eLife 13:RP92085.
https://doi.org/10.7554/eLife.92085.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}