Cortical neuroprosthesis-mediated functional ipsilateral control of locomotion in rats with spinal cord hemisection
- Département de Neurosciences, Groupe de recherche sur la Signalisation Neurale etla Circuiterie (SNC) and Centre Interdisciplinaire de Recherche sur le Cerveau etl’Apprentissage (CIRCA), Université de Montréal, Canada
- CIUSSS du Nord-de-l'Île-de-Montréal, Canada
Figures
Figure 1
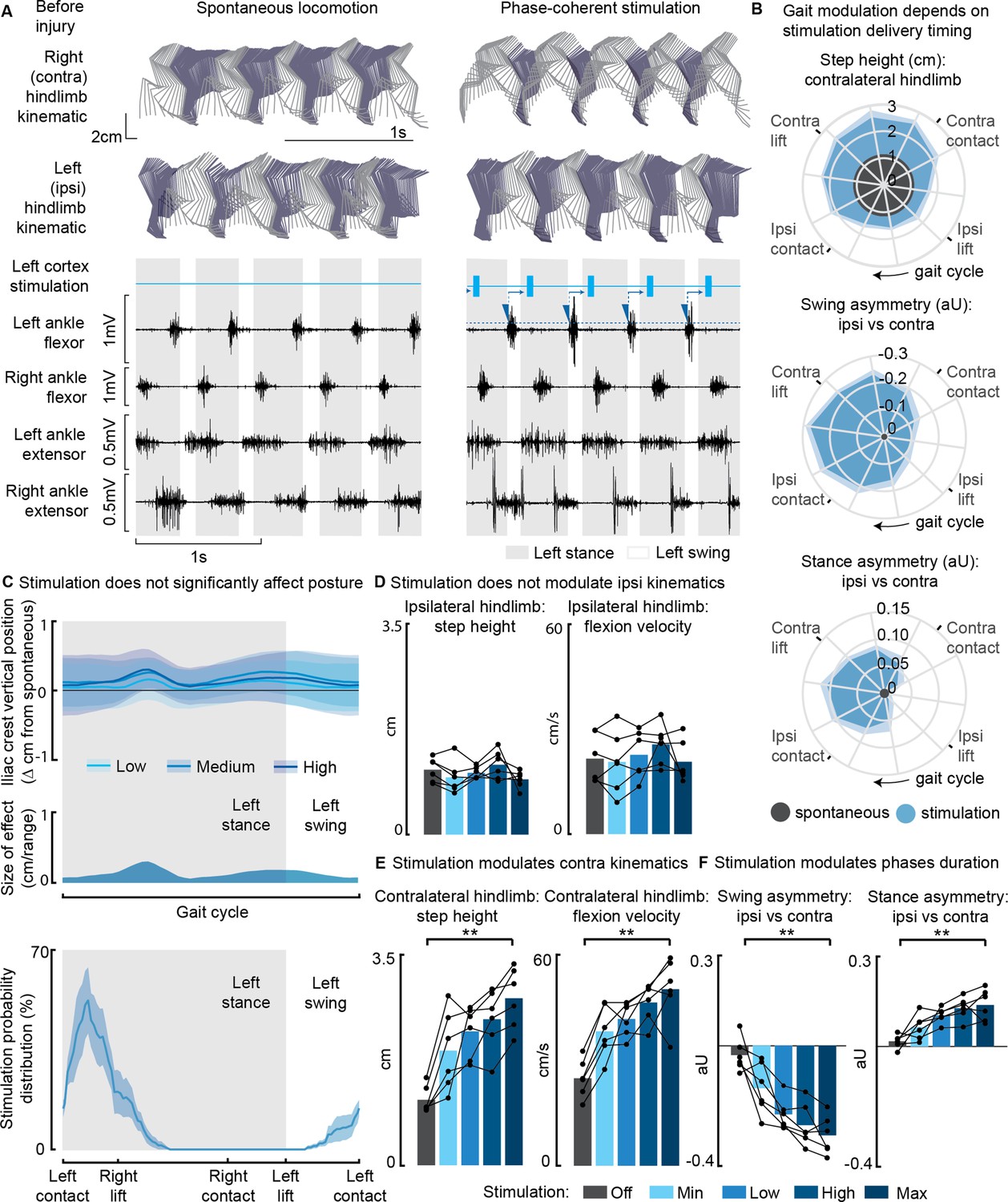
Phase-coherent intracortical stimulation modulated contralateral kinematics in intact rats (n=6 rats).
(A) Stick diagrams and electromyographic (EMG) activity during spontaneous locomotion and phase-coherent stimulation. The stimulation was triggered by ipsilateral ankle flexor activation and was delivered during the late contralateral stance (early ipsilateral stance). On the left ankle trace, the dotted line represents a threshold value for the ankle EMG to cause a trigger, represented by a blue triangle. The stimulation delivery follows a fixed delay indicated with an arrow. (B) Polar plots showing contralateral step height in cm and gait phase asymmetries in arbitrary units (aU, calculated as Δ% length) for stimulation delivered with different timings along the whole gait cycle. Positive asymmetry index values refer to ipsilateral-side dominance. For ease of reading, the radial axis of the swing symmetry plot has been inverted (outer values are negative). For the three polar plots, the most effective kinematic neuromodulation corresponds to the largest radius. The gait cycle progresses clockwise. (C) Analysis of the effects of cortical stimulation on the posture of rats (top) and experimental stimulation distribution (bottom). Posture is shown as the height of the ipsilateral iliac crest during the gait cycle, which was not modulated by increasing cortical stimulation amplitude, indicated as Low, Medium, and High to represent 33%, 66%, and 100% of the functional stimulation range, defined from motor threshold to maximum comfortable stimulus. (D) Characterization of ipsilateral kinematics. Ipsilateral step height and flexion speed were not affected by increasing cortical stimulation amplitudes. (E) Modulation of contralateral kinematics. Contralateral step height and flexion speed were linearly increased with greater stimulation amplitudes. (F) Modulation of bilateral gait phase duration. The absolute values of swing and stance asymmetry indexes were linearly increased with greater stimulation amplitudes. Positive asymmetry index values refer to ipsilateral-side dominance. The data are represented as the mean ± SEM. **p<0.01.
-
Figure 1—source data 1
Source data for Figure 1B, D-F.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig1-data1-v1.xlsx
Figure 2
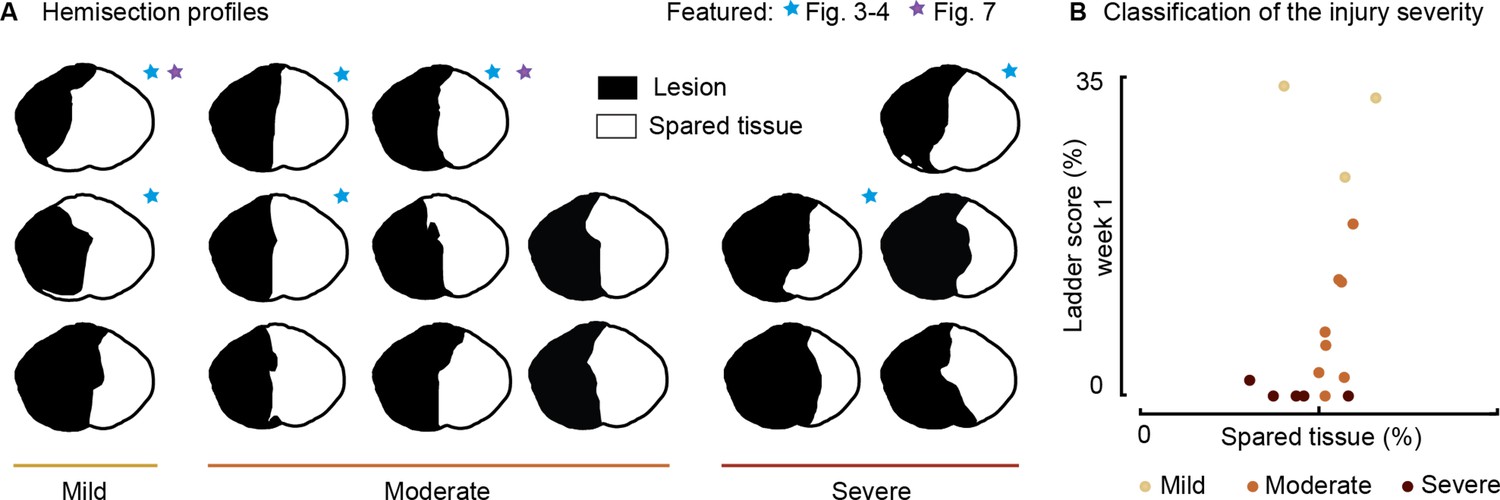
Spinal lesion severity (n=16 rats).
Related to Figures 3, 4 and 7. (A) Hemisection profiles at the epicenter level. (B) Classification of the injury severity. Injury severity groups were defined according to skilled locomotion performance during ladder crossing 7 days after injury. The injuries were classified as mild: left hindlimb>20% correct paw placement, moderate: left hindlimb<20% correct paw placement and right hindlimb>75% correct paw placement and severe: right hindlimb<75% correct paw placement (bilateral deficits).
-
Figure 2—source data 1
Source data for Figure 2B.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig2-data1-v1.xlsx
Figure 3 with 4 supplements
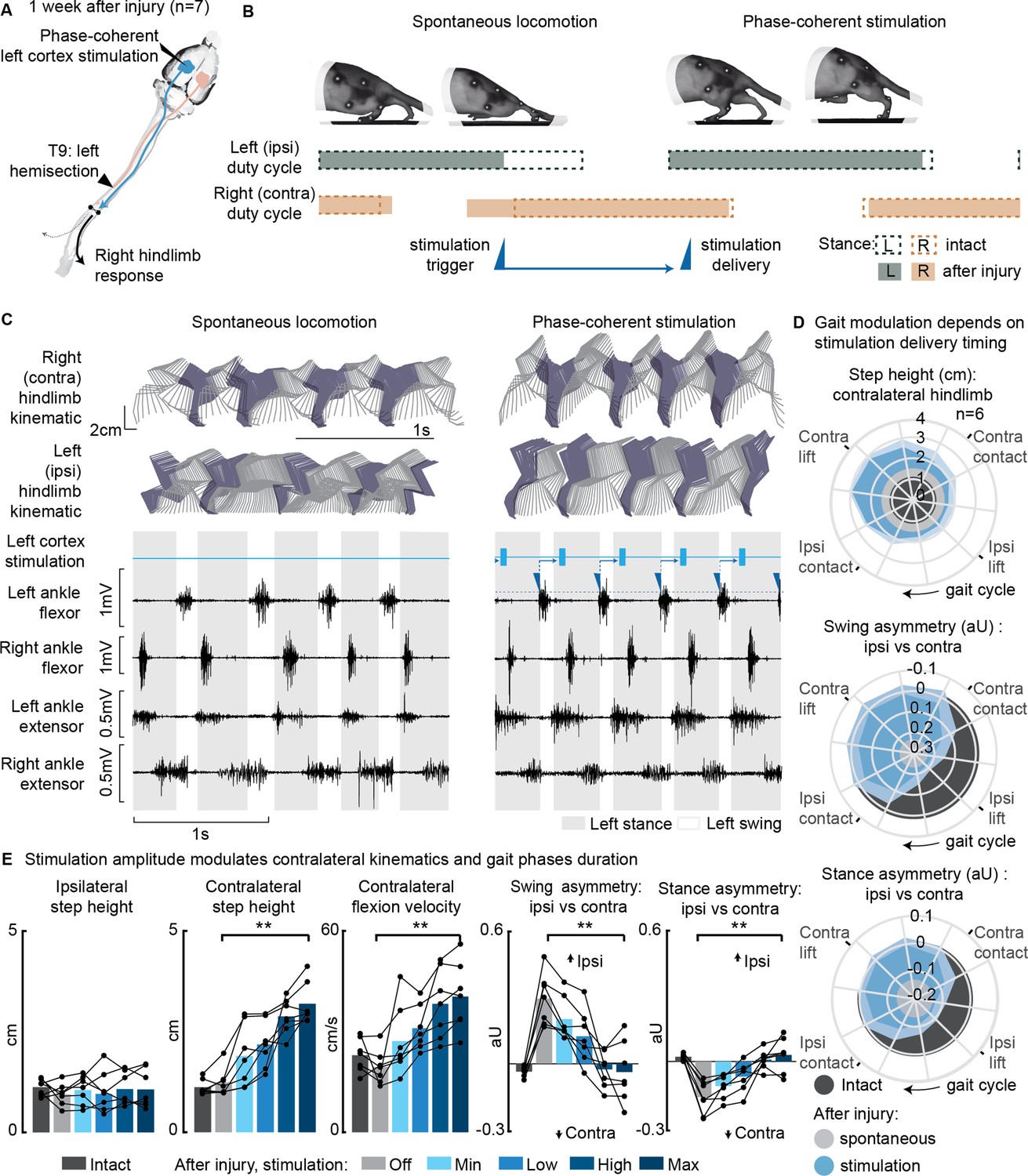
Phase-coherent intracortical stimulation alleviated locomotor deficits 1 week after injury (n=7 rats).
(A) A schematic representation of the injury and neurostimulation model showing the thoracic left hemisection (T9) and left (ipsilesional) motor cortex stimulation. (B) A schematic representation of spontaneous locomotion and phase-coherent stimulation effects on postural changes, gait phase duration and alternation, as well as stimulation trigger and delivery timings. The stimulation, triggered in correspondence with the ipsilateral lift and delivered just before the contralateral lift, resulted in a stronger contralateral swing and a synchronous stronger ipsilateral stance. (C) Stick diagrams and electromyographic (EMG) activity during spontaneous locomotion and phase-coherent stimulation. The stimulation was triggered by ipsilateral ankle flexor activation and was delivered during the late contralateral stance (early ipsilateral stance). (D) Polar plots showing contralateral step height (cm) and gait phase asymmetry variations (aU, calculated as Δ% length) for stimulation delivered at different timings along the whole gait cycle. Positive asymmetry index values refer to ipsilateral-side dominance. For ease of reading, the radial axis of the swing symmetry plot has been inverted (outer values are negative). For the three polar plots, the condition of strongest neuromodulation corresponded to the largest radius. Gait phase symmetry, highly affected during spontaneous locomotion, was recovered for stimulation delivered after the ipsilateral contact and before the contralateral contact. The gait cycle progresses clockwise. (E) The contralateral kinematics and gait phase durations were linearly modulated with increasing stimulation amplitudes. Positive asymmetry index values refer to ipsilateral-side dominance. Phase-coherent stimulation generated an increase in the step height and flexion speed of the contralateral hindlimb and mediated the recovery of the physiological symmetry between the ipsilateral and contralateral swing and stance phases. The data are represented as the mean ± SEM. **p<0.01. The hemisection profiles of the seven rats are identified by a blue star in Figure 2A.
-
Figure 3—source data 1
Source data for Figure 3D and E.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig3-data1-v1.xlsx
Figure 3—figure supplement 1
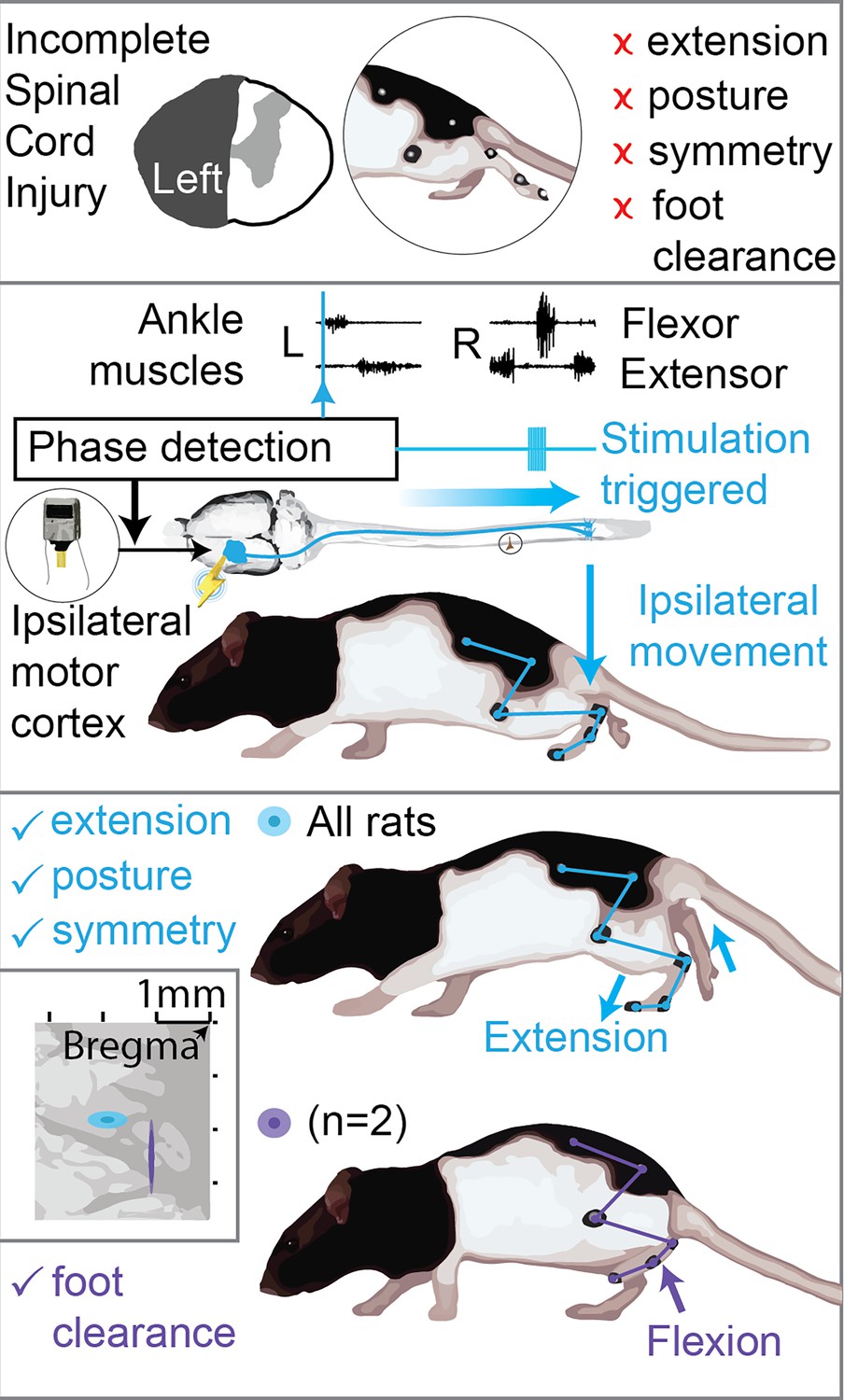
Graphical scheme summarizing the results.
Top: hemisection profiles at the epicenter level. Unilateral spinal cord injury (SCI) causes leg flexion and extension deficits, affecting posture, symmetry and causing leg dragging. Middle: phase-coherent cortical stimulation is obtained by monitoring leg muscle activity and triggering cortical stimulation by reference muscle pattern recognition. In this work we utilize phase-coherent stimulation to unveil ipsilateral cortical control of movement. Bottom: ipsilateral phase-coherent cortical stimulation improves leg extension in all rats and flexion, alleviating foot drop, in two rats displaying a cluster of active sites with unique flexor properties. Related to Figure 3 and Figure 7.
Figure 3—figure supplement 2
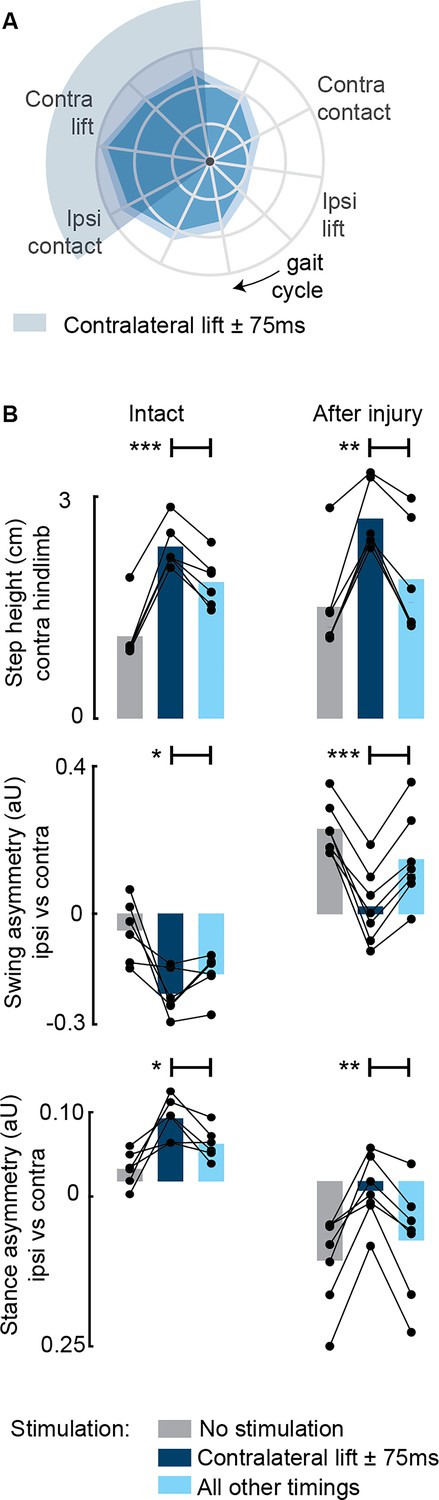
Aggregate timing characterization of phase-coherent stimulation (n=7 rats).
(A) Polar plots showing contralateral step height (cm) for stimulation delivered at different timings along the whole gait cycle. The contralateral lift start time ±75 ms is highlighted with a shaded area. The gait cycle progresses clockwise. (B) Contralateral step height was maximized by stimuli delivered around the time of the contralateral lift. Gait phase symmetry, highly affected during spontaneous locomotion, was recovered for stimulation delivered around the time of the contralateral lift. Positive asymmetry index values refer to left-side dominance. The data are represented as mean values and individual replicates. *, **, *** p<0.05, p<0.01, and p<0.001, respectively.
-
Figure 3—figure supplement 2—source data 1
Source data for Figure 3—figure supplement 2B.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig3-figsupp2-data1-v1.xlsx
Figure 3—figure supplement 3
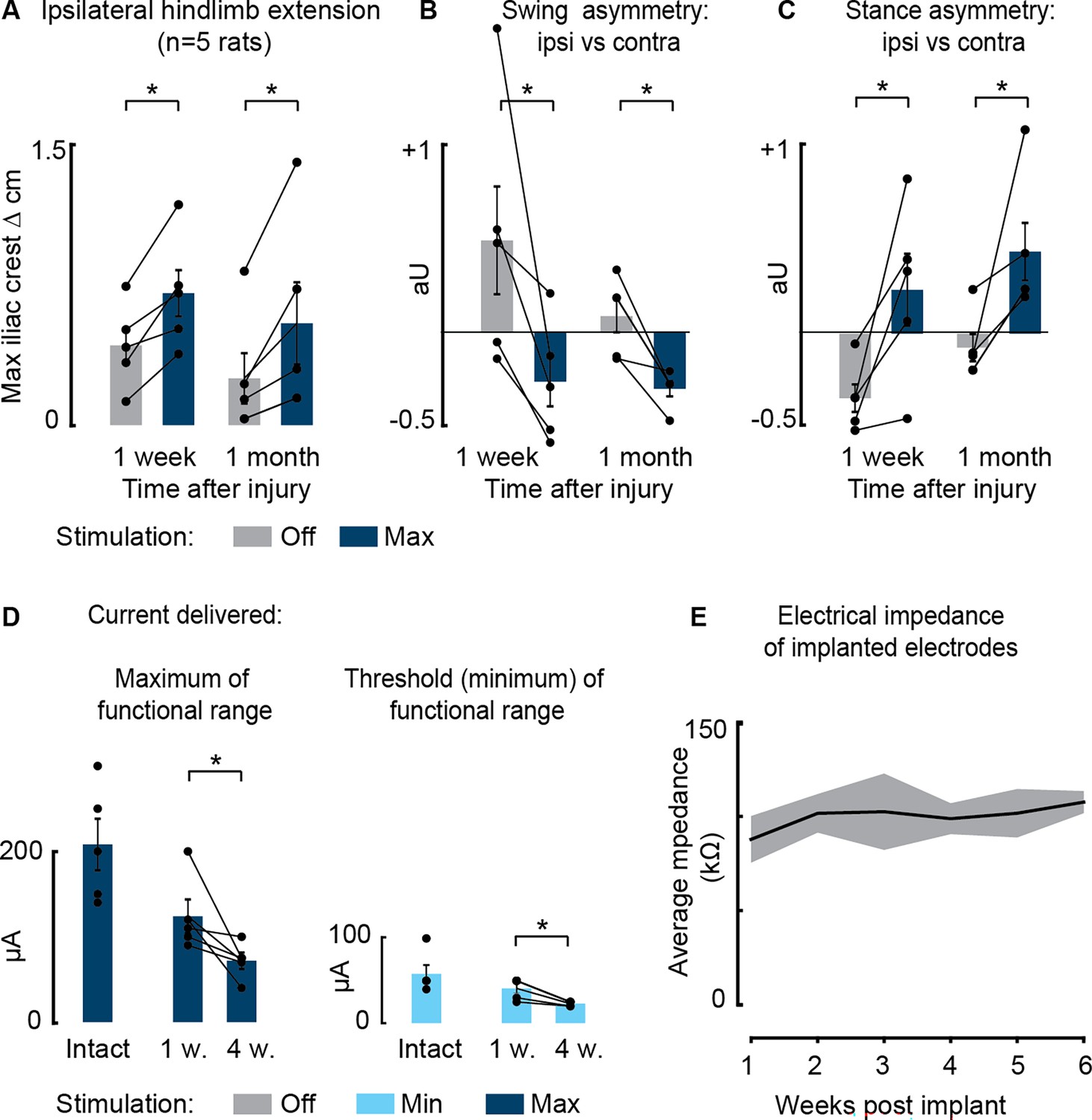
Persistent effects of phase-coherent cortical stimulation 1 month after spinal cord injury (SCI) (n=5 rats).
(A–C) Comparison of maximal stimulation effects on posture (A), swing asymmetry (B), and stance asymmetry (C), 1 week vs. 1 month after SCI. (D) Current delivered to obtain ipsilateral modulation of movement (functional range from the minimal threshold value to visualize motor effect to the maximum value that the rat was able to integrate without discomfort). (E) Average impedance recorded at each electrode array over 6 weeks post-implantation. Data are shown as mean ± SEM. *, p<0.05.
-
Figure 3—figure supplement 3—source data 1
Source data for Figure 3—figure supplement 3.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig3-figsupp3-data1-v1.xlsx
Figure 3—video 1
Cortical neuroprosthesis-mediated control of ipsilateral hindlimb extension.
Treadmill walking during spontaneous locomotion and with phase-coherent stimulation 1 week after injury. The stimulation was delivered during the late right or early left stance and resulted in a flexion of the contralesional hindlimb and a synchronous extension of the ipsilesional hindlimb.
Figure 4
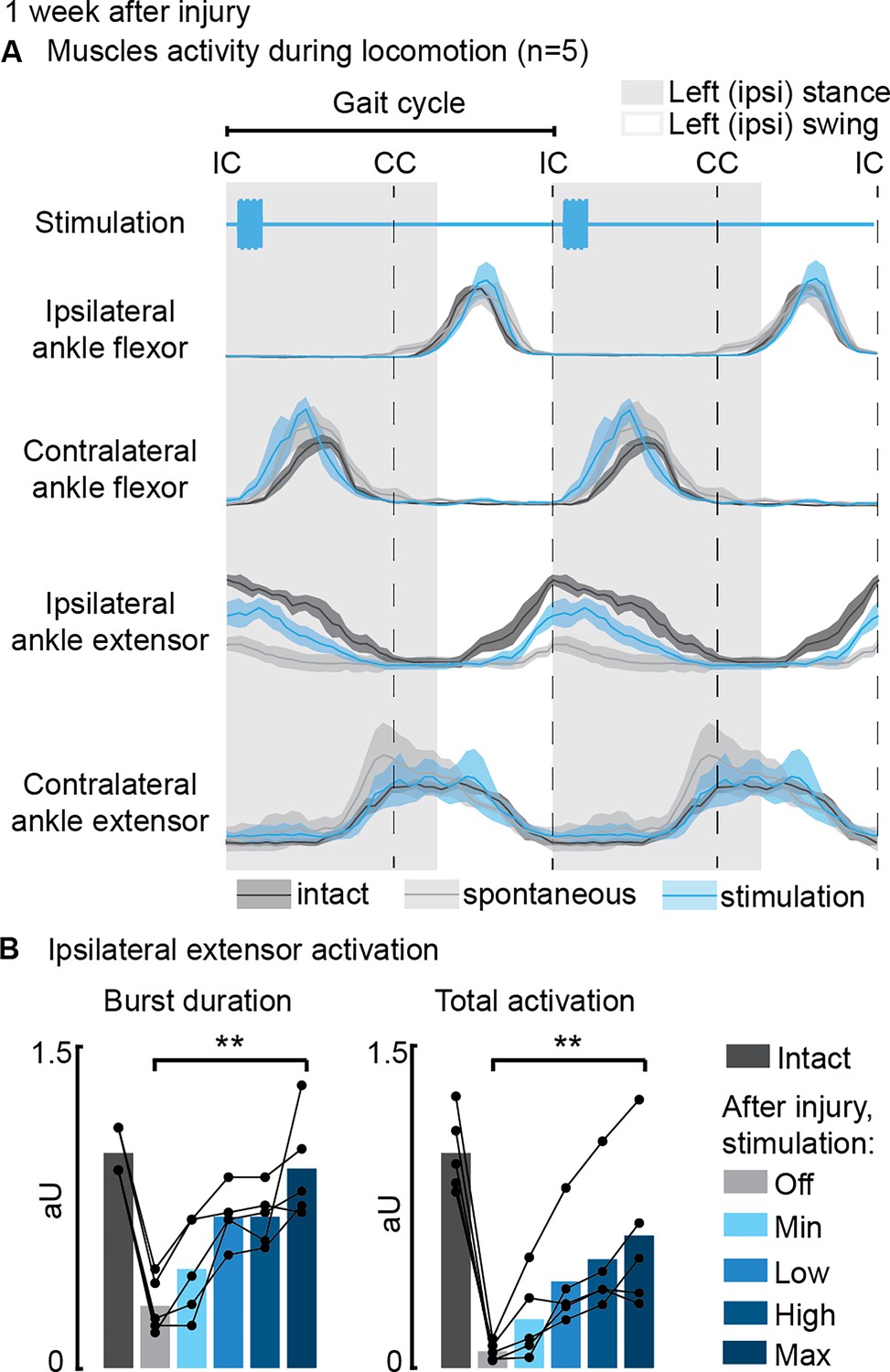
Phase-coherent intracortical stimulation reinstated ipsilateral extension muscle activity 1 week after injury (n=5 recordings for each muscle, from a total of 7 rats, with some muscles unavailable due to implant failure).
(A) Electromyographic (EMG) envelopes during spontaneous locomotion before and after injury as well as phase-coherent stimulation after injury. Activities of the ipsilateral and contralateral ankle flexor (tibialis anterior) and ipsilateral and contralateral ankle extensor (medial gastrocnemius). The gait event references are reported as LC: left contact, RC: right contact. (B) Left medial gastrocnemius activity was modulated by the stimulation. The burst duration and the level of muscle activation were linearly increased with greater stimulation amplitudes. The data are represented as the mean ± SEM. **p<0.01. The hemisection profiles of the seven rats are identified by a blue star in Figure 2A.
-
Figure 4—source data 1
Source data for Figure 4B.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig4-data1-v1.xlsx
Figure 5
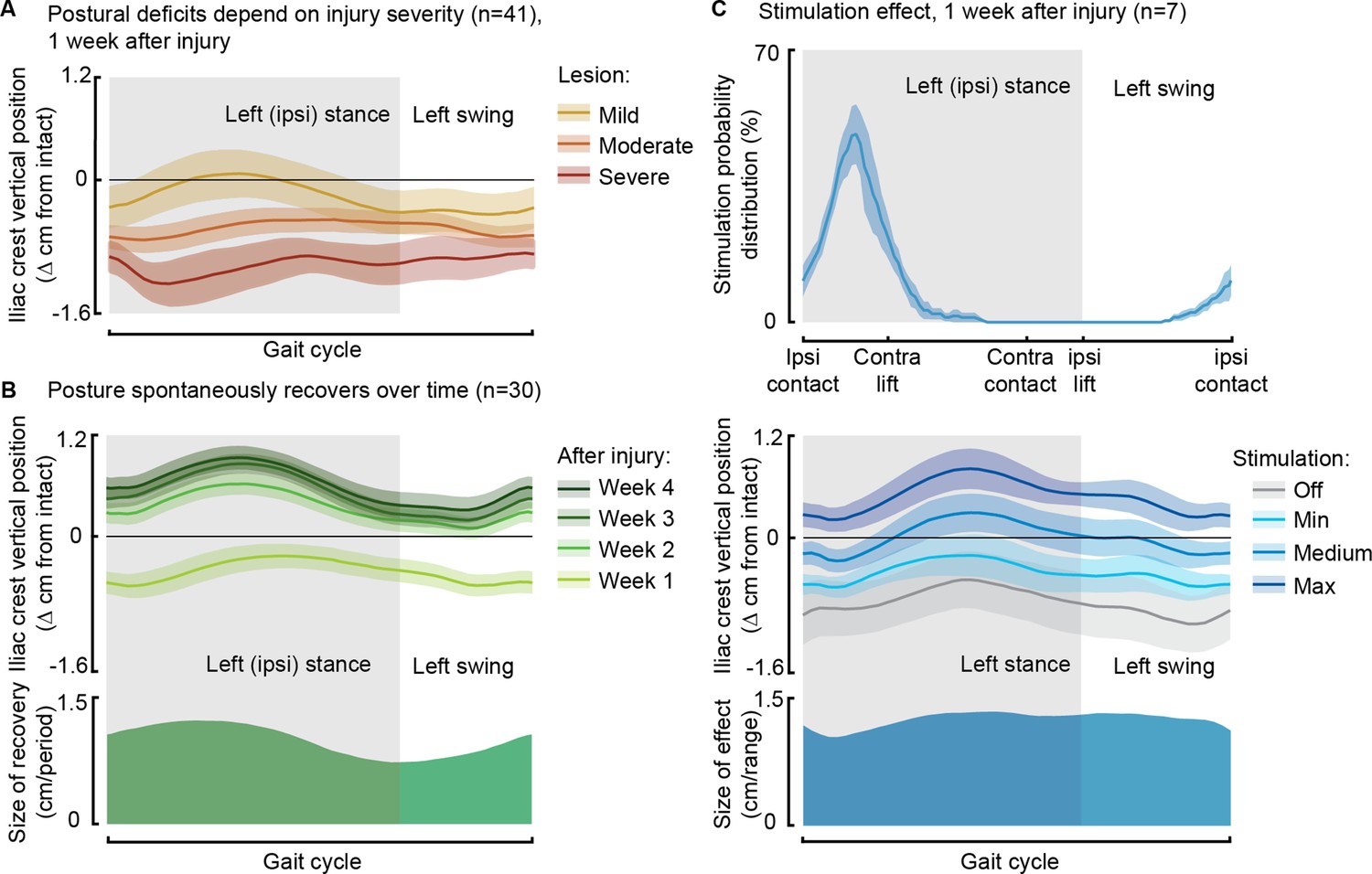
Phase-coherent stimulation improved posture 1 week after injury.
Posture is shown as the height of the ipsilateral iliac crest during locomotion with respect to the spontaneous condition before injury. The data are represented as the mean ± SEM. (A) Postural deficits depend on injury severity. Rats with severe spinal cord injury (SCI) exhibit a weaker posture 1 week after injury. (B) Variation over 1 month of spontaneous recovery. Posture is raised and overcompensated. (C) Effect of phase-coherent stimulation 1 week after injury. Posture is increasingly raised with greater stimulation amplitudes. n=41, 30, or 7 rats, indicated in each panel.
Figure 6 with 1 supplement
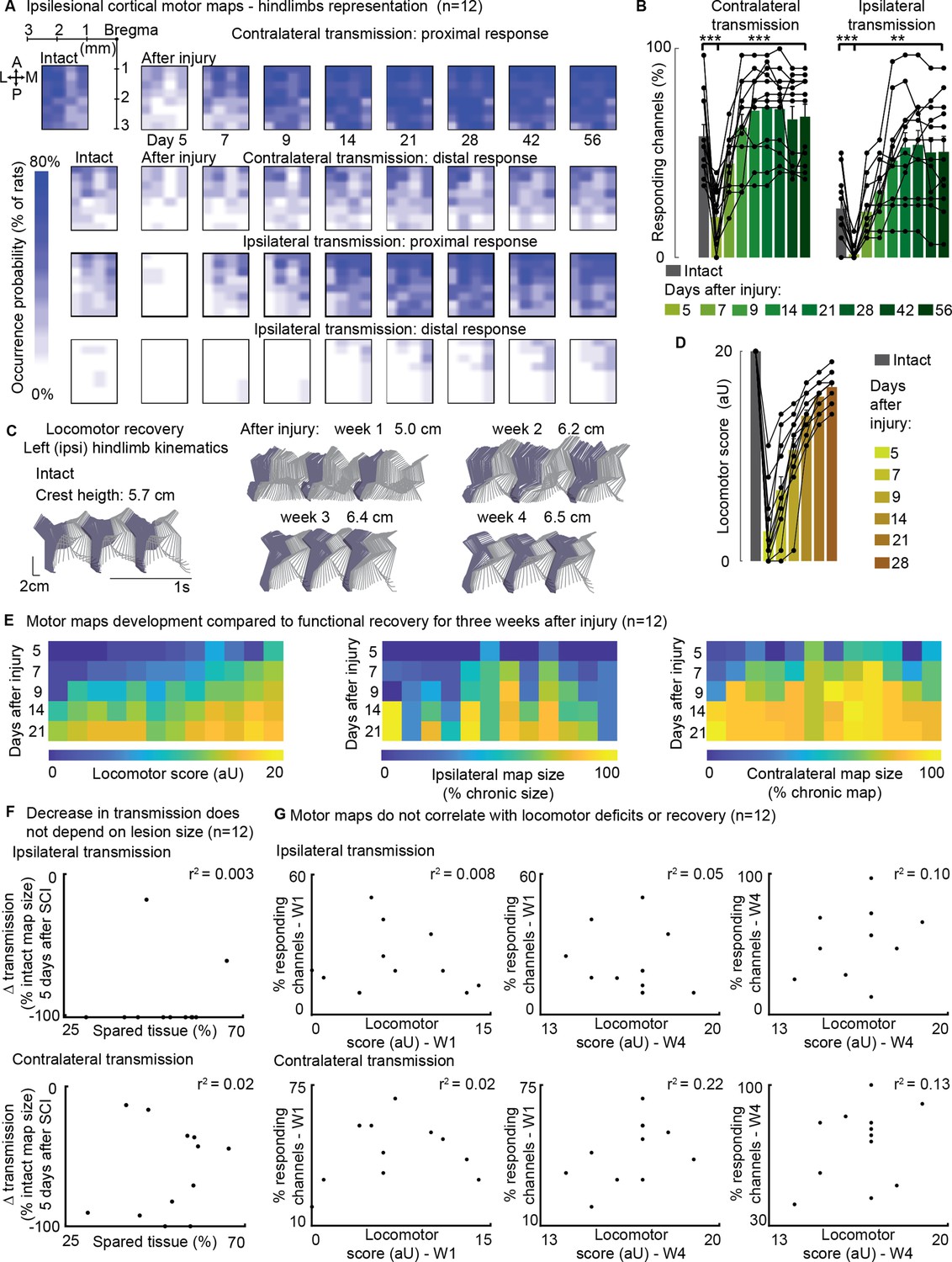
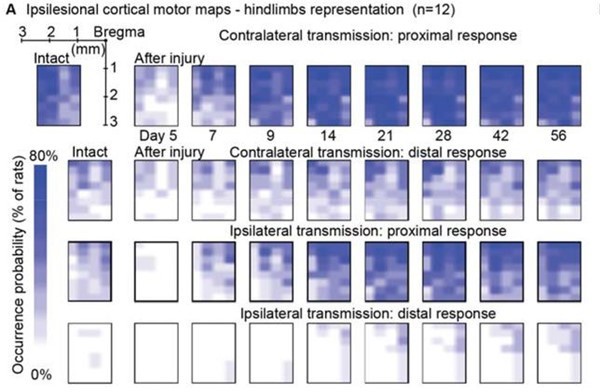
Ipsilateral motor representation of the affected hindlimb was increased in the ipsilesional motor cortex after injury but does not reflect functional recovery (n=12 rats).
The term ‘transmission’ in figure indicates a quantification of the number of responding sites within the array, which is the surface whereby a stimulus transmission to the muscles resulted in a visible hindlimb contraction. (A) Awake cortical motor map representation before injury and up to 2 months after injury. The color intensity is proportional to the probability of evoking proximal/distal ipsi-/contra-lateral responses when stimulating a given site, across 12 animals. Bilateral representation of hindlimb movements increased over time compared to the intact condition. The top left sub-panel carries a representation of the electrode array positioning within the left motor cortex. (B) Quantification of responding channels from the intact condition and up to 2 months after injury. (C) Stick diagrams from treadmill locomotion and iliac crest height before injury and during the first 4 weeks after injury. (D) Quantification of locomotor score from the intact condition and up to 1 month after injury. (E) Corticospinal transmission and locomotor performance. Single rats (columns) are sorted by day 7 locomotor score. The same sorting is maintained for the three sub-panels. Absolute map size (i.e. responding sites on the implanted array, indicated as % of the final, chronic, size) did not correlate with higher or lower locomotor scores across rats, whereas the individual rats’ trend of map size increase for 3 weeks paralleled locomotor recovery. (F) Lack of correlation between map size and lesion size 5 days after injury. An ipsilateral and contralateral decrease in transmission did not parallel the spared tissue at the lesion epicenter. (G) Lack of correlation between map size and locomotor score. Time points are reported as W1: week 1 and W4: week 4. Ipsilateral and contralateral transmission did not correlate with global locomotor recovery measured in an open field. Bars: mean of individual data points.
-
Figure 6—source data 1
Source data for Figure 6.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig6-data1-v1.xlsx
Figure 6—figure supplement 1
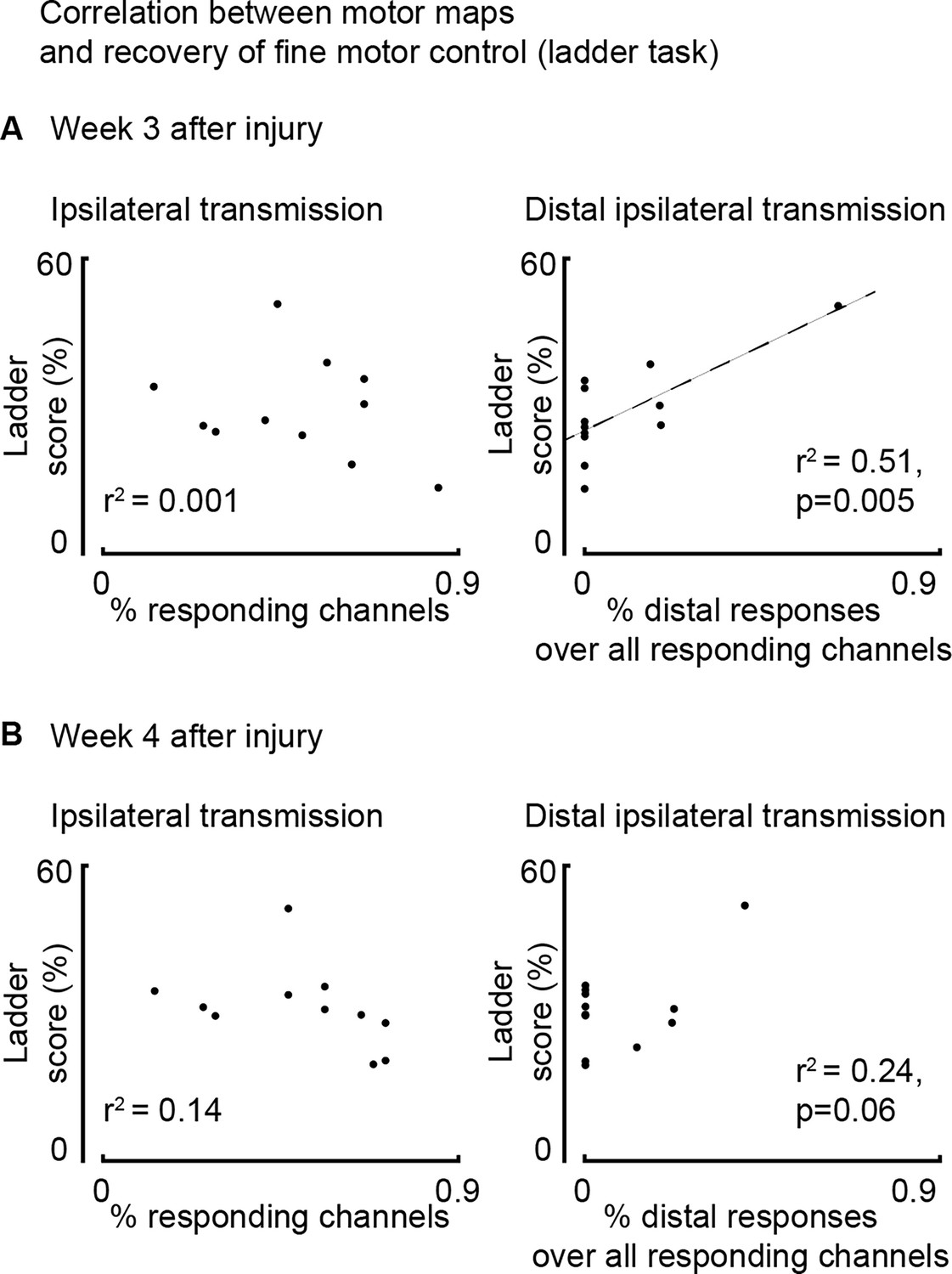
Assessment of correlation between motor maps and recovery of fine motor control evaluated as ladder score (n=12 rats).
(A) Correlation between map size and ladder score 3 weeks after injury. Return of ipsilateral distal transmission (but not general transmission: proximal and distal) paralleled recovery of fine motor control, as assessed by the ladder crossing task. (B) The same assessment performed 4 weeks after injury.
-
Figure 6—figure supplement 1—source data 1
Source data for Figure 6—figure supplement 1.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig6-figsupp1-data1-v1.xlsx
Figure 7 with 1 supplement
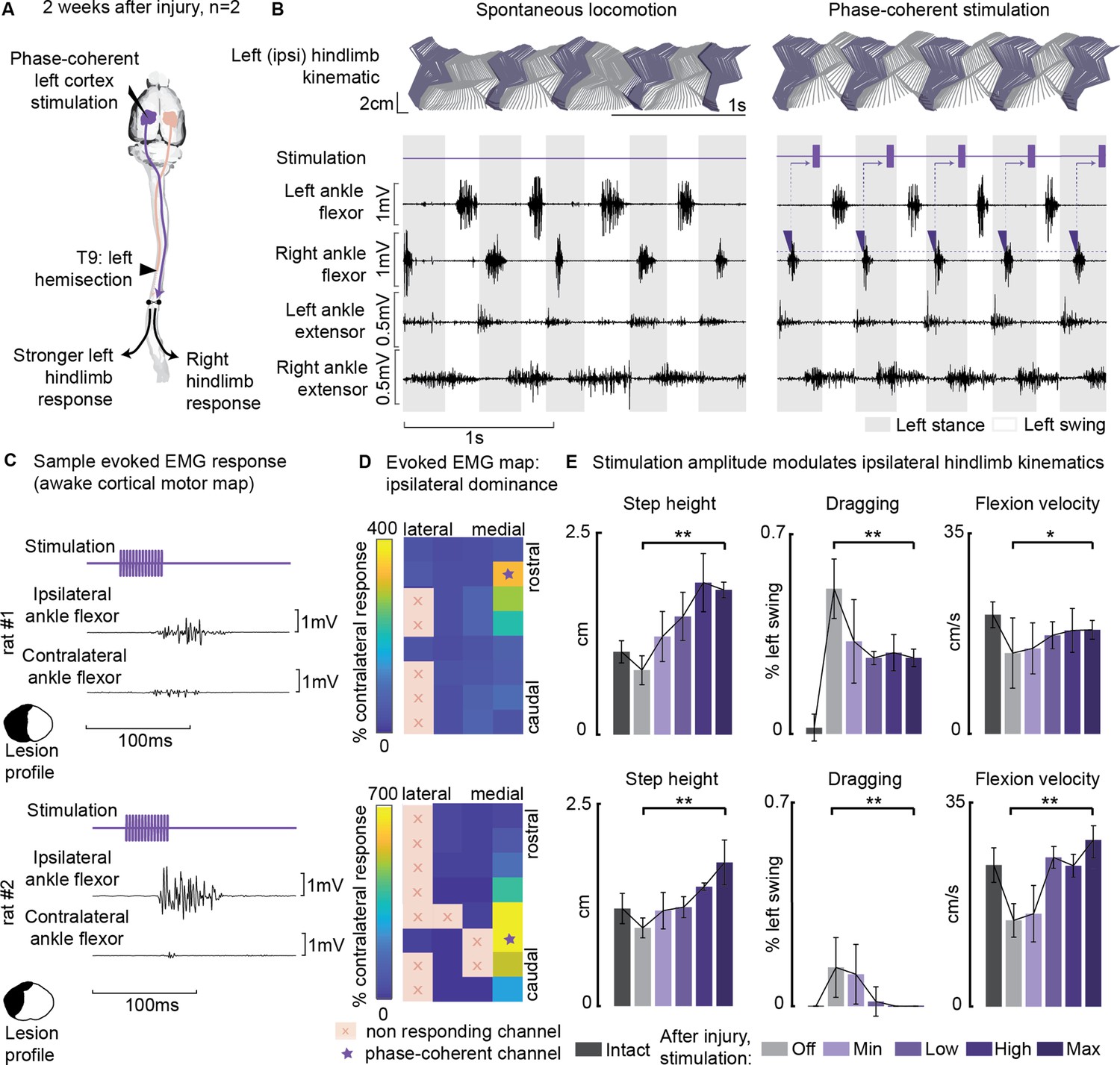
Ipsilesional motor cortex stimulation modulated ipsilateral hindlimb movements 2 weeks after injury (n=10 steps, experiment repeated in two rats).
(A) A schematic representation of the injury and neurostimulation model. After lateral hemisection, ipsilesional motor cortex stimulation evoked ipsilateral responses. (B) Stick diagrams and electromyographic (EMG) activity during spontaneous locomotion and phase-coherent stimulation. The stimulation was triggered by contralateral ankle flexor activation and was delivered during the late ipsilateral stance. (C) Samples of single train stimulation of specific channels that preferentially evoked ipsilateral muscle activation in two different animals. (D) Ipsilateral dominance of EMG responses. Awake cortical motor maps were obtained as a ratio between ipsilateral and contralateral tibialis anterior activation. Channels that presented an ipsilateral preference were located in the most medial region of the map and identified by a star. (E) Phase-coherent stimulation modulated ipsilateral kinematics and reduced the foot drop deficit. Ipsilateral step height, flexion speed, and dragging alleviation linearly increased with greater stimulation amplitudes. Two subjects are presented independently, 10 steps per condition. The hemisection profiles of the two rats are identified by a purple star in Figure 2A. The data are represented as the mean ± SEM. *, ** p<0.05 and <0.01, respectively.
-
Figure 7—source data 1
Source data for Figure 7D and E.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig7-data1-v1.xlsx
Figure 7—video 1
Cortical neuroprosthesis-mediated control of ipsilateral hindlimb flexion.
Treadmill walking during spontaneous locomotion and with phase-coherent stimulation 2 weeks after injury. The stimulation was delivered during the late left or early right stance and resulted in a flexion of the ipsilesional hindlimb.
Figure 8
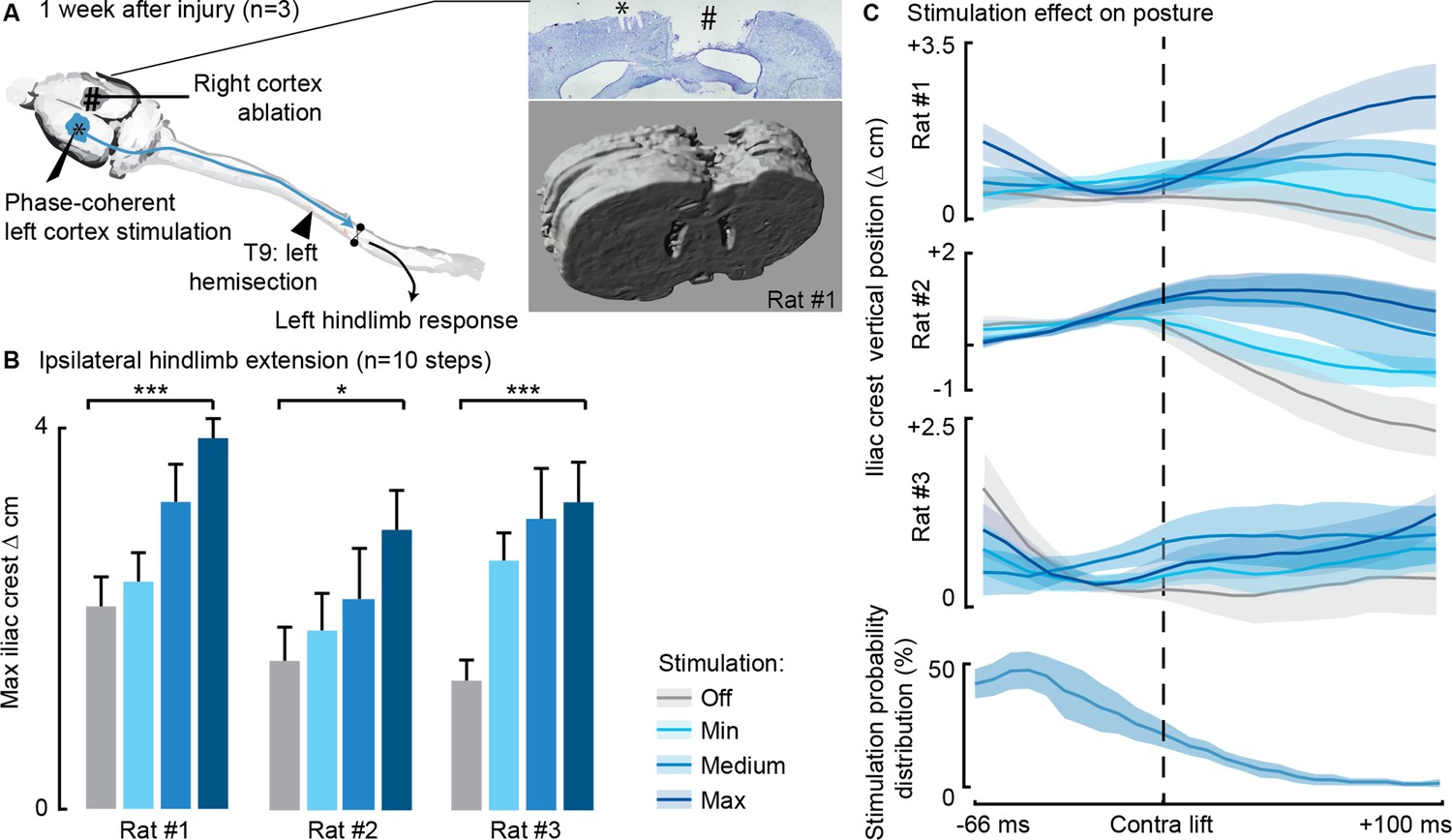
Phase-coherent stimulation of the ipsilesional motor cortex alleviated locomotor deficits even after ablation of the contralateral motor cortex (n=3 rats).
(A) Schematic representation of the injury and neurostimulation model. After lateral hemisection, ipsilesional motor cortex stimulation evoked ipsilateral responses, even when the contralateral motor cortex was ablated. Right inset: top, Cresyl violet staining of a coronal brain slice of rat #1. *, electrode traces in the left cortex. #, right cortex ablation. (B–C) Phase-coherent stimulation modulated ipsilateral kinematics. Posture linearly increased with greater stimulation amplitudes. Three rats are presented independently, 10 steps per condition. The data are represented as the mean ± SEM. *, *** p<0.05 and <0.001, respectively.
-
Figure 8—source data 1
Source data for Figure 8B.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig8-data1-v1.xlsx
Figure 9 with 1 supplement
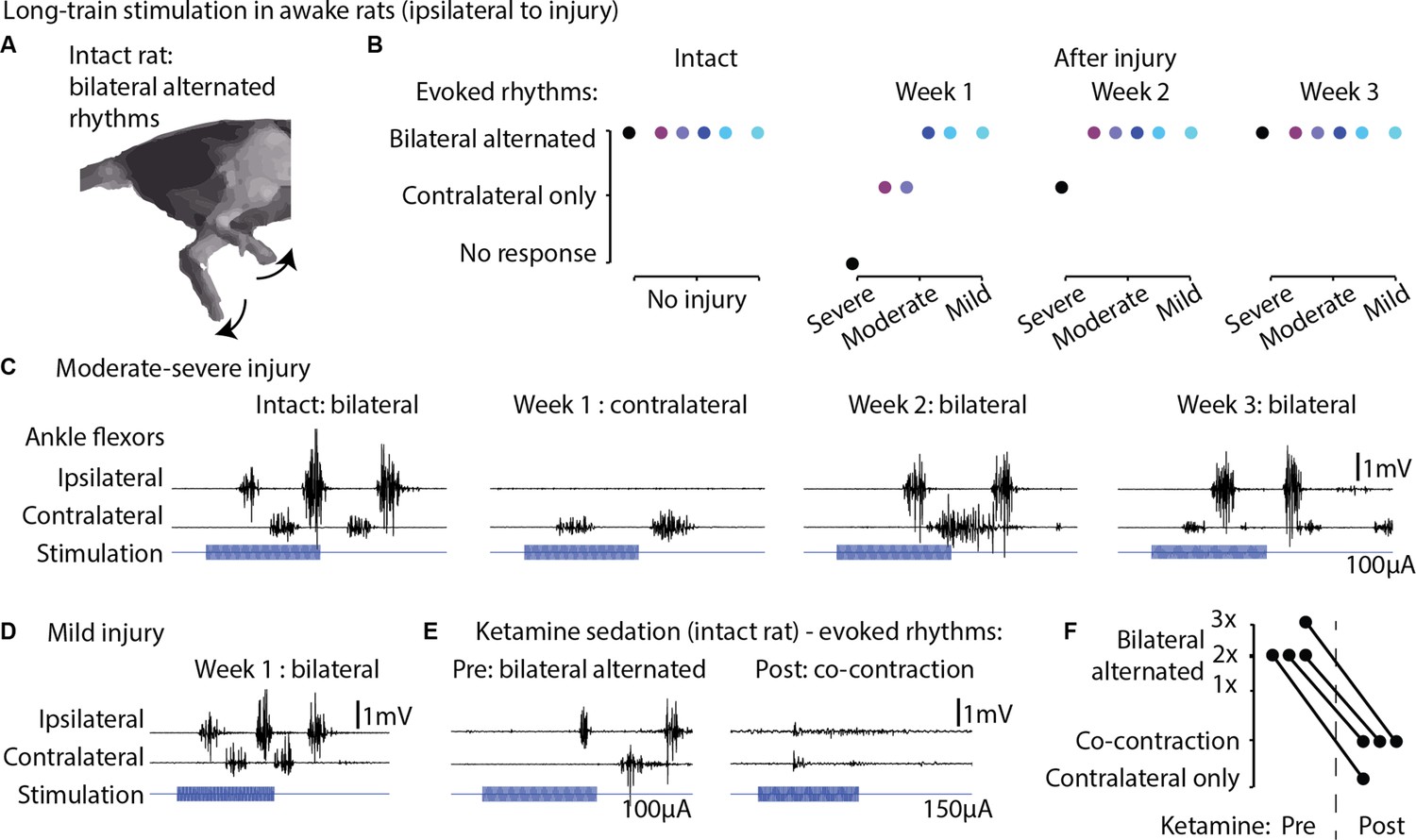
Long-train intracortical stimulation in awake rats elicited alternated bilateral rhythms.
(A) Schematic representation of the locomotor-like rhythmic movements evoked by long-train (250 ms) cortical stimulation (amplitude 100 µA). Evoked rhythms are characterized by alternated hindlimb movements. (B) In six intact rats, stimulation of the left motor cortex generated bilateral alternated hindlimb rhythms. After spinal cord injury (SCI), rats are sorted by injury severity, using their ladder score at week 1 for ranking. One week after injury, long-train cortical stimulation failed to evoke bilateral alternated rhythms in half of the cohort. In two of these rats, contralateral rhythms were still present and bilateral alternated rhythms were recovered by week 2. In the most severe rat, contralateral-only rhythms were evoked on week 2 and bilateral alternated rhythms on week 3. For the remaining half of the cohort, long-train cortical stimulation recruited bilateral alternated rhythms at all tested time points. (C) Stimulus-synchronized ankle flexor electromyographic (EMG) traces from one rat with a moderate-severe injury, showing loss (week 1) and following recovery (weeks 2–3) of ipsilateral evoked hindlimb rhythms. (D) Stimulus-synchronized EMG trace from one rat with mild injury, showing that bilateral alternated evoked rhythms are preserved at week 1. (E) Stimulus-synchronized EMG trace from four intact rats before and after ketamine sedation, showing transient loss of bilateral alternated rhythms. (F) Loss of bilateral alternated rhythms in four rats after ketamine sedation. 1X, 2X, 3X: number of complete repetitions of alternating movements produced by long-train cortical stimulation (amplitude 150 µA).
-
Figure 9—source data 1
Source data for Figure 9B and F.
- https://cdn.elifesciences.org/articles/92940/elife-92940-fig9-data1-v1.xlsx
Figure 9—video 1
Long-train cortical stimulation recruits spinal locomotor circuits.
Hindlimb responses to long-train intracortical stimulation in the intact state, with and without ketamine sedation, and at two time points (1 and 2 weeks) after spinal cord injury (SCI). The stimulation was delivered in resting rats manually supported at torso and forelimbs, with the hindlimbs left free to move.
Author response image 1

Threshold analysis.
Author response image 2
Occurrence probability analysis, for comparison.
Additional files
-
Supplementary file 1
List of animals engaged in each experiment.
Rats marked with * did not receive left motor cortex implantation. They were included in the study for establishing spontaneous changes in posture over time (Figure 5A and B).
- https://cdn.elifesciences.org/articles/92940/elife-92940-supp1-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/92940/elife-92940-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cortical neuroprosthesis-mediated functional ipsilateral control of locomotion in rats with spinal cord hemisection
eLife 12:RP92940.
https://doi.org/10.7554/eLife.92940.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}