Bridging the gap between presynaptic hair cell function and neural sound encoding
- Auditory Neuroscience and Synaptic Nanophysiology Group, Max Planck Institute for Multidisciplinary Sciences, Germany
- Institute for Auditory Neuroscience, University Medical Center Göttingen, Germany
- Collaborative Research Center, University of Göttingen, Germany
- Cluster of Excellence 'Multiscale Bioimaging of Excitable Cells', Germany
Figures
Figure 1 with 1 supplement
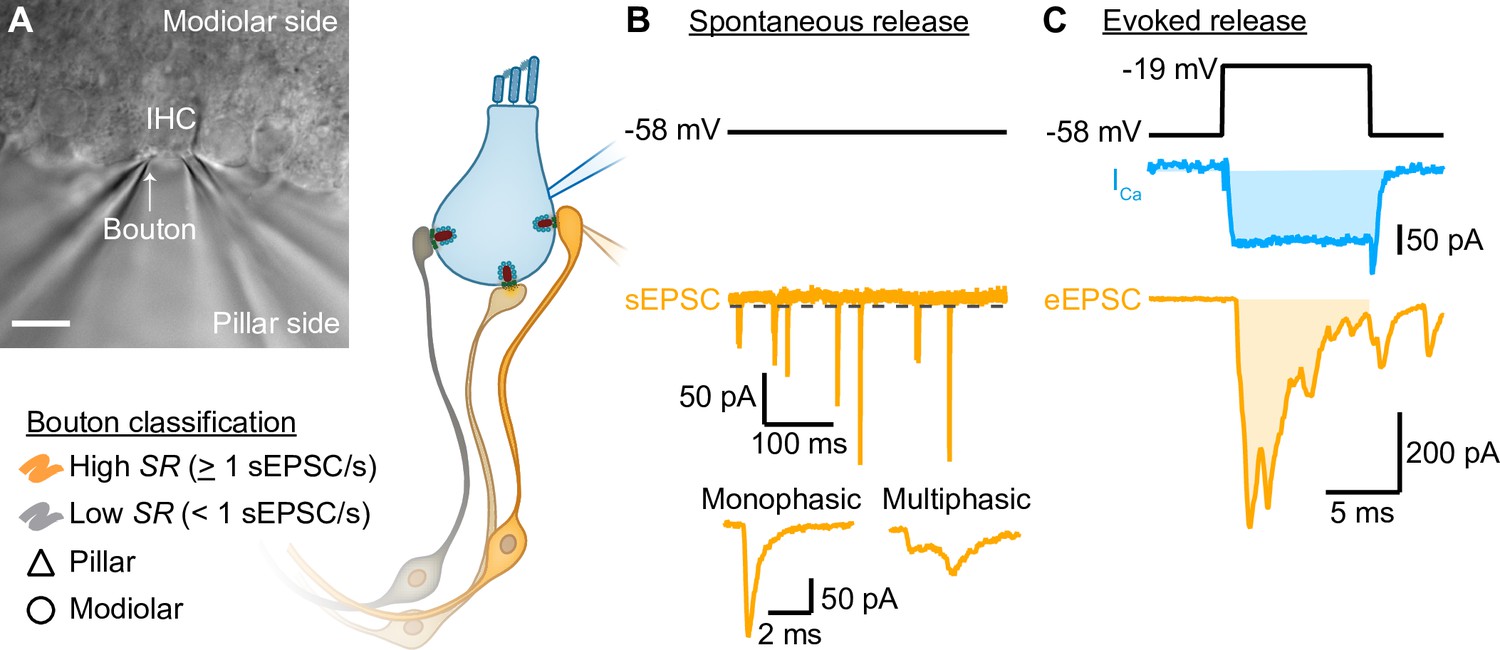
Paired inner hair cell (IHC)- spiral ganglion neuron bouton patch-clamp recordings to study the release properties of individual IHC ribbon synapses as a function of synapse position.
(A) Differential interference contrast (DIC) image of an explanted murine organ of Corti. In this example, supporting cells from the pillar side were removed to gain access to the IHCs and their contacting boutons. The recorded boutons were classified based on their position (△ pillar or ❍ modiolar) and on their spontaneous rate (SR) (Low SR<1 sEPSC/s vs High SR>1 sEPSC/s). Scale bar: 10 µm. (B) Spontaneous release was recorded in absence of stimulation (i.e. IHC holding potential = –58 mV; Supplementary file 1; dashed line represents the threshold for sEPSC detection). sEPSCs were classified as monophasic (a steady rise to peak and monoexponential decay, temporally more compact) or as multiphasic (multiple inflections and slowed raising and decaying kinetics, non-compact). (C) Evoked release: depolarizing pulses (black trace) were used to trigger whole IHC Ca2+ influx (ICa, blue trace) and ensuing release of neurotransmitter that evoked EPSCs (eEPSCs, light orange trace). Ca2+ charge and eEPSC charge were estimated by taking the integral of the currents (shaded light blue and light orange areas).
Figure 1—figure supplement 1
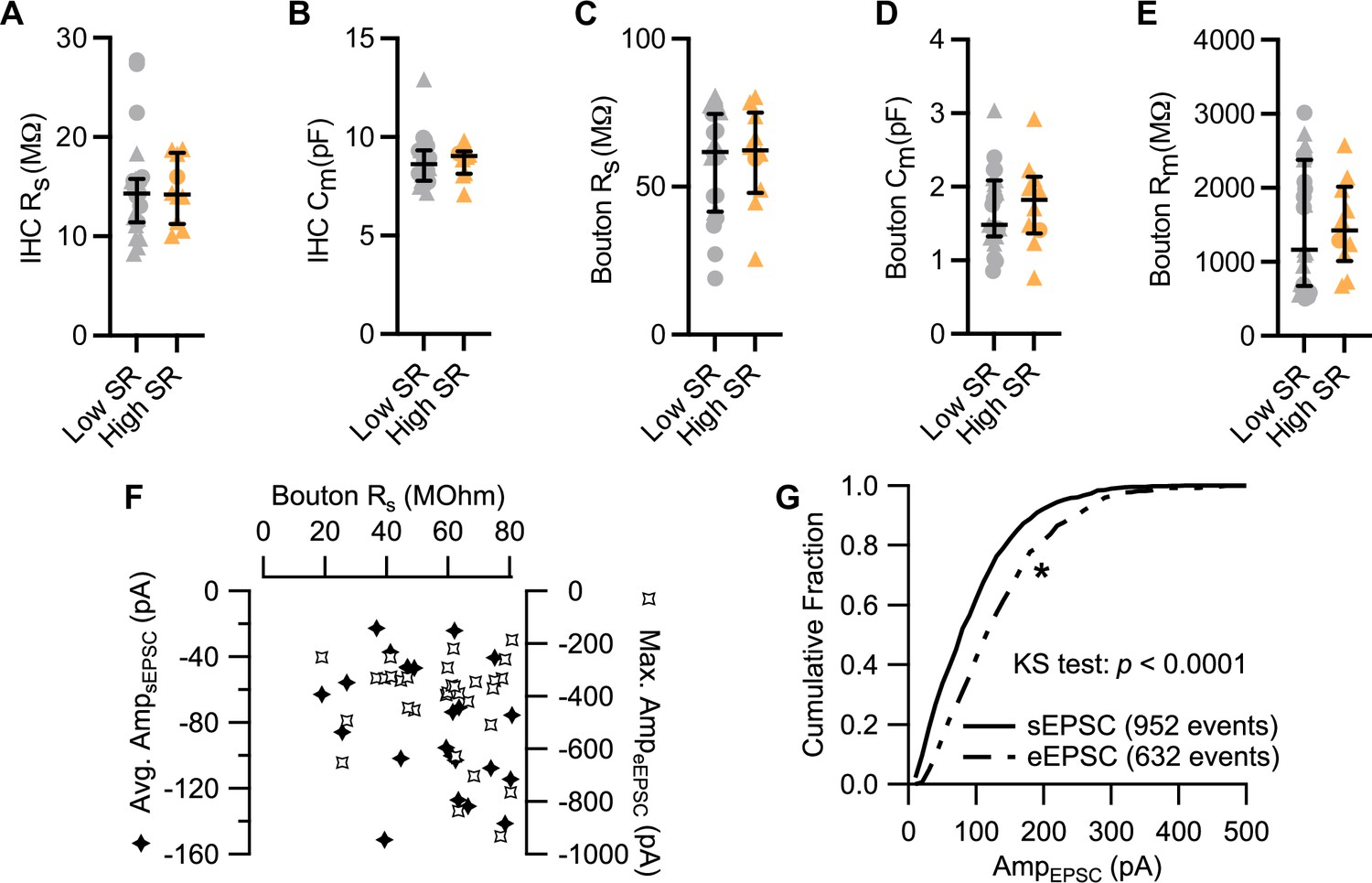
Passive electrical properties of inner hair cells (IHCs) and spiral ganglion neuron (SGN) boutons.
(A–E) Values for IHC’s series resistance (Rs, A) and membrane capacitance (Cm, B), and bouton’s Rs (C), Cm (D) and membrane resistance (Rm, E) were comparable between paired recordings with low and high SR. (F) Average sEPSC amplitudes and maximal evoked EPSCs (eEPSC) amplitudes plotted against bouton Rs. (G) Cumulative fraction of amplitude of sEPSCs and of eEPSCs recorded from the 13 synapses included in Figure 4. The eEPSCs correspond to voltage-evoked EPSCs occurring in the last 20 ms of the Masker stimulus. The two distributions differ significantly (p<0.0001, Kolmogorov-Smirnov test). Average sEPSC amplitude of –97.28±2.22 pA (median 82.10 pA) vs average eEPSC amplitude of –135.8±3.24 pA (median 120.0 pA). Panels A–E show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 2 with 1 supplement
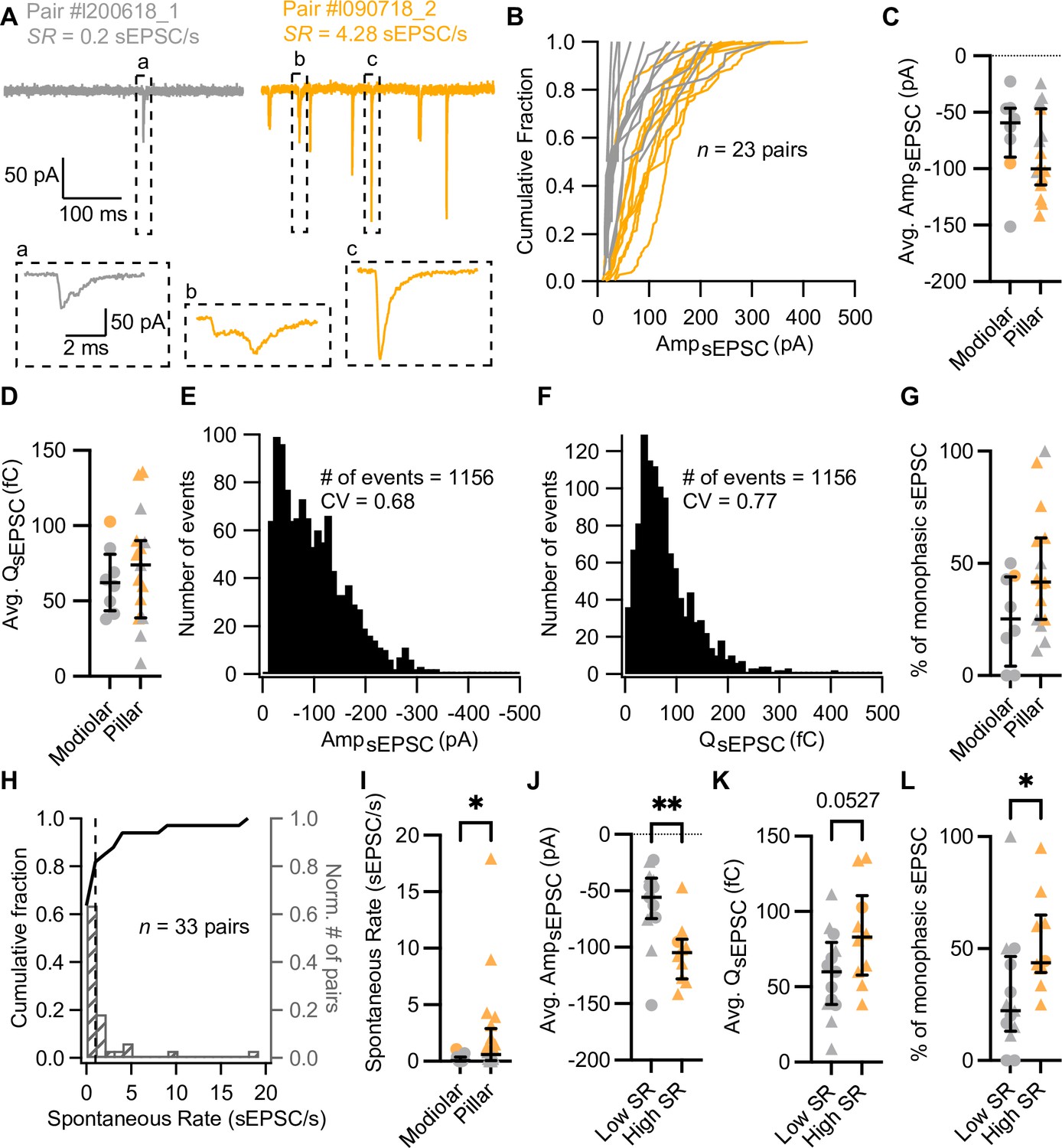
Synapses with high spontaneous release have larger and more monophasic spontaneous EPSCs (sEPSCs).
(A) sEPSCs recorded in the absence of stimulation (i.e. inner hair cell [IHC] holding potential = –58 mV) from two exemplary paired recordings with different spontaneous rate (SR: gray for low SR, orange for high SR). ‘Pair #’ identifies individual paired recordings. Insets show the selected sEPSCs in an expanded time scale. (a, b) correspond to multiphasic sEPSCs, while (c) represents a typical monophasic sEPSC. (B) Cumulative sEPSC amplitude plots for 23 paired synapses that had spontaneous release. (C–D) Average sEPSC amplitude (C) and charge (D) from individual synapses recorded from the pillar or modiolar side of the IHC. (E–F) Pooled sEPSC amplitude (E) and charge (F) distributions show a distinct peak at –40 pA and 40 pC, respectively. Bin size: 10 pA or pC. (G) Percentage of monophasic sEPSCs in pillar and modiolar synapses. (H) Cumulative fraction (left axis) and normalized histogram (right axis) of the spontaneous rate (bin size is 1 sEPSC/s) of 33 pairs. (I) Pillar synapses had higher rates of sEPSCs. (J–L) High SR synapses had significantly larger sEPSC amplitudes (J), a tendency to bigger sEPSC charges (K) and higher percentages of monophasic sEPSCs (L). Panels G, I–L show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 2—figure supplement 1
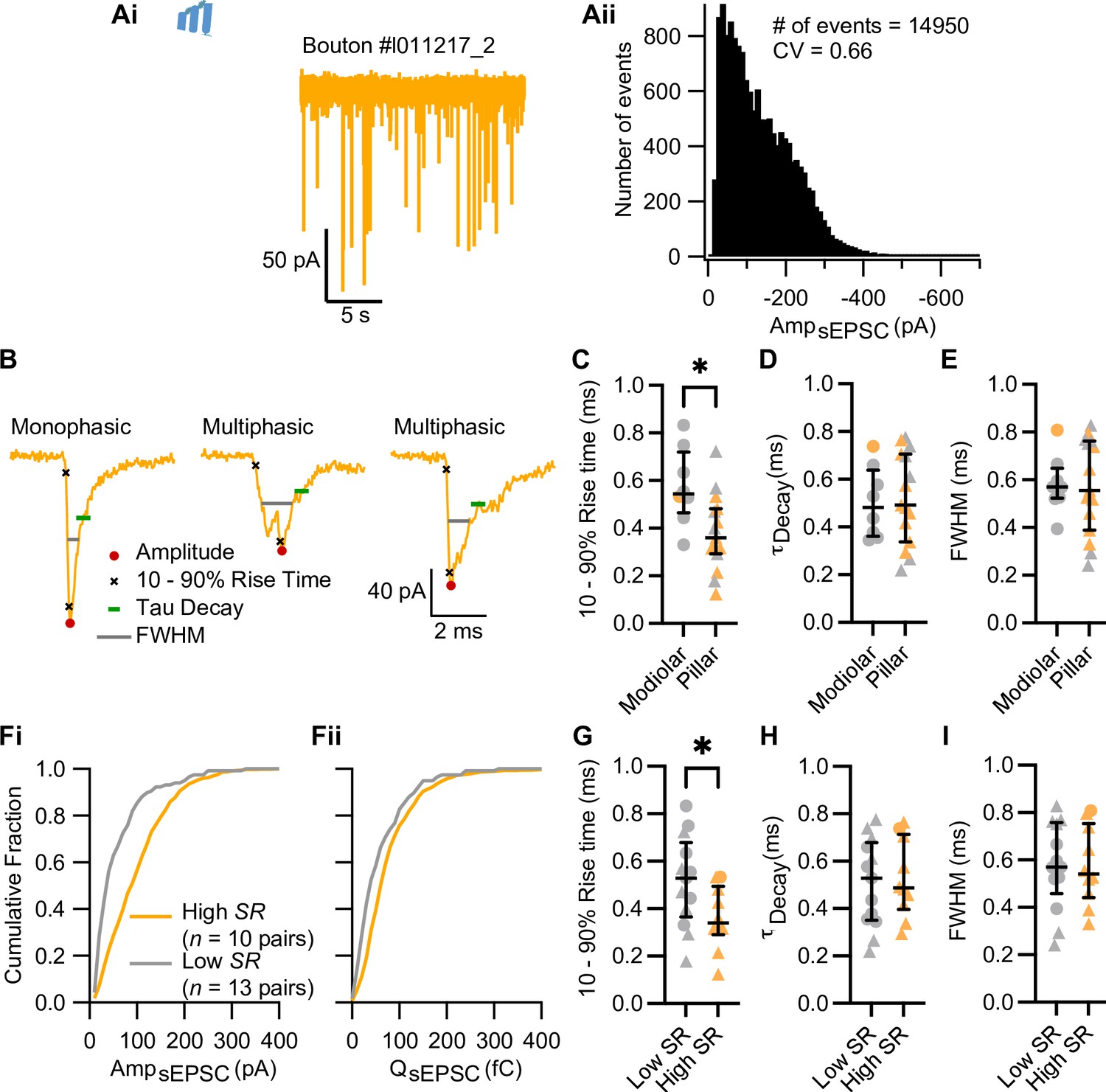
Pillar and high spontaneous release synapses have sEPSCs with faster rising times.
(Ai–Aii) Ruptured-patch whole-cell recordings from postsynaptic spiral ganglion neuron (SGN) boutons (without the inner hair cell [IHC]) displayed spontaneous rates from 0 up to 16.33 sEPSC/s. Pooled sEPSC amplitude (Aii) show a peak at –40 pA, with amplitudes ranging from around –10 to –670 pA. Bin size: 10 pA. (B) Kinetics of sEPSCs, such as amplitude, 10–90% rise time, time constant of decay (τdecay), and full-width half-maximum (FWHM), were calculated with Neuromatic (Rothman and Silver, 2018). (C–E) Pillar synapses had faster 10–90% rise times (C) than modiolar synapses, while their τdecay (D) and FWHM (E) were comparable. (Fi–Fii) Cumulative fraction of amplitude and charge of sEPSCs from low and high SR synapses. (G–I) High SR synapses had faster 10–90% rise times (G) than modiolar synapses, while their τdecay (H) and FWHM (I) were comparable. Panels C–E, G–I show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 3 with 2 supplements
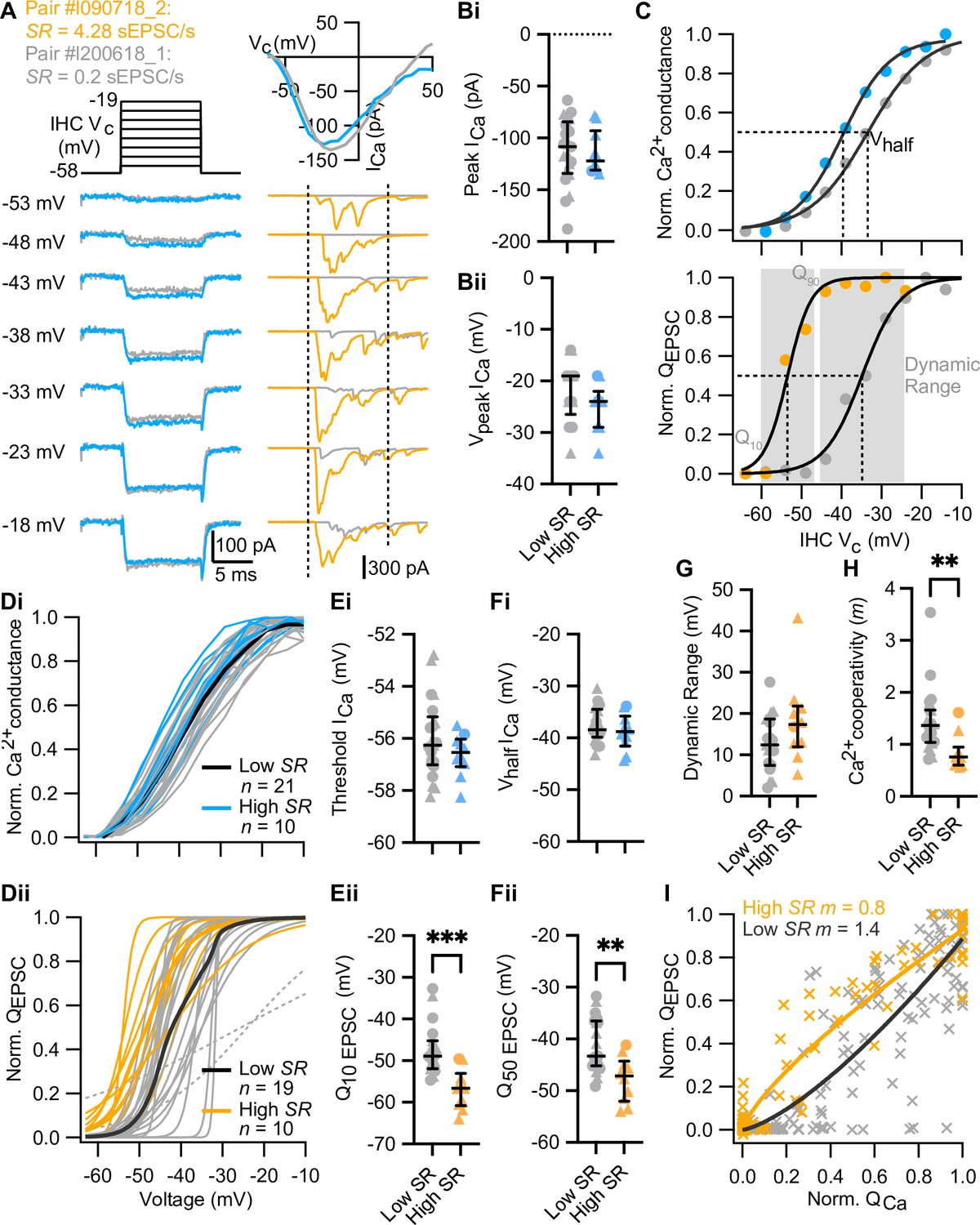
High spontaneous release synapses activate at lower voltages and show tighter Ca2+ channel coupling of synaptic release.
(A) Voltage-protocol (top left), inner hair cell (IHC) Ca2+ current (ICa, bottom left and top right, blue and gray) and evoked EPSCs (eEPSCs) (bottom right, orange and gray) of a high and a low SR pair, respectively, in response to 10 ms depolarizations (dashed vertical lines on top of eEPSC data indicate the onset and offset of the depolarization) to different potentials ranging from –58 to –19 mV in 5 mV steps. The upper right panel shows the current-voltage relationships for the two pairs. (Bi–Bii) The peak of whole-cell Ca2+ current (Bi) and the voltage eliciting maximum Ca2+ current (Bii) of IHCs were comparable between high and low SR synapses. (C) Upper panel: Fractional activation of the Ca2+ channels (blue and gray data points from the examples shown in A) was obtained from the normalized chord conductance. Voltage of half-maximal activation (Vhalf ICa; dotted line) and voltage sensitivity of activation (slope) were determined using a Boltzmann fit (black trace) to the activation curve. Lower panel: Release-intensity curve (orange and gray data points from the examples shown in A) was obtained from the QEPSC for each depolarization step. A sigmoidal function (black trace) was fitted to obtain the voltage of half-maximal synaptic release (Q50 EPSC; dotted line) and the voltage sensitivity of the release (slope), as well as the dynamic range for which the exocytosis changes from 10% to 90% (gray area). (Di–Dii) Voltage dependence of whole-cell Ca2+ channel activation (activation curve; Di) and fits to release-intensity curves (Dii) for 31 synapses. Averages (thick lines) and individual curves (thin lines) are overlaid. The release-intensity curve of two low SR pairs could not be fitted (gray dotted lines). (Ei–Fi) The threshold of Ca2+ influx (Ei) and Vhalf ICa (Fi) did not differ between low and high SR synapses. (Eii–Fii) Voltage of 10% maximum release (Q10 EPSC, Eii) and Q50 EPSC (Fii) were significantly more hyperpolarized in high SR synapses. (G) Dynamic range of release was comparable between low and high SR synapses. (H) Ca2+ cooperativity (m) estimated from fitting a power function to the QEPSC – QCa relationship for each individual synapse (see Figure 3—figure supplement 2) was significantly lower in high SR synapses. (I) Scatter plot of normalized QEPSC vs the corresponding normalized QCa. The solid lines are a least-squares fit of a power function (QEPSC = a(QCa)m) to the data, yielding mhigh SR of 0.8 and mlow SR of 1.4. Panels B, E–H show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 3—figure supplement 1
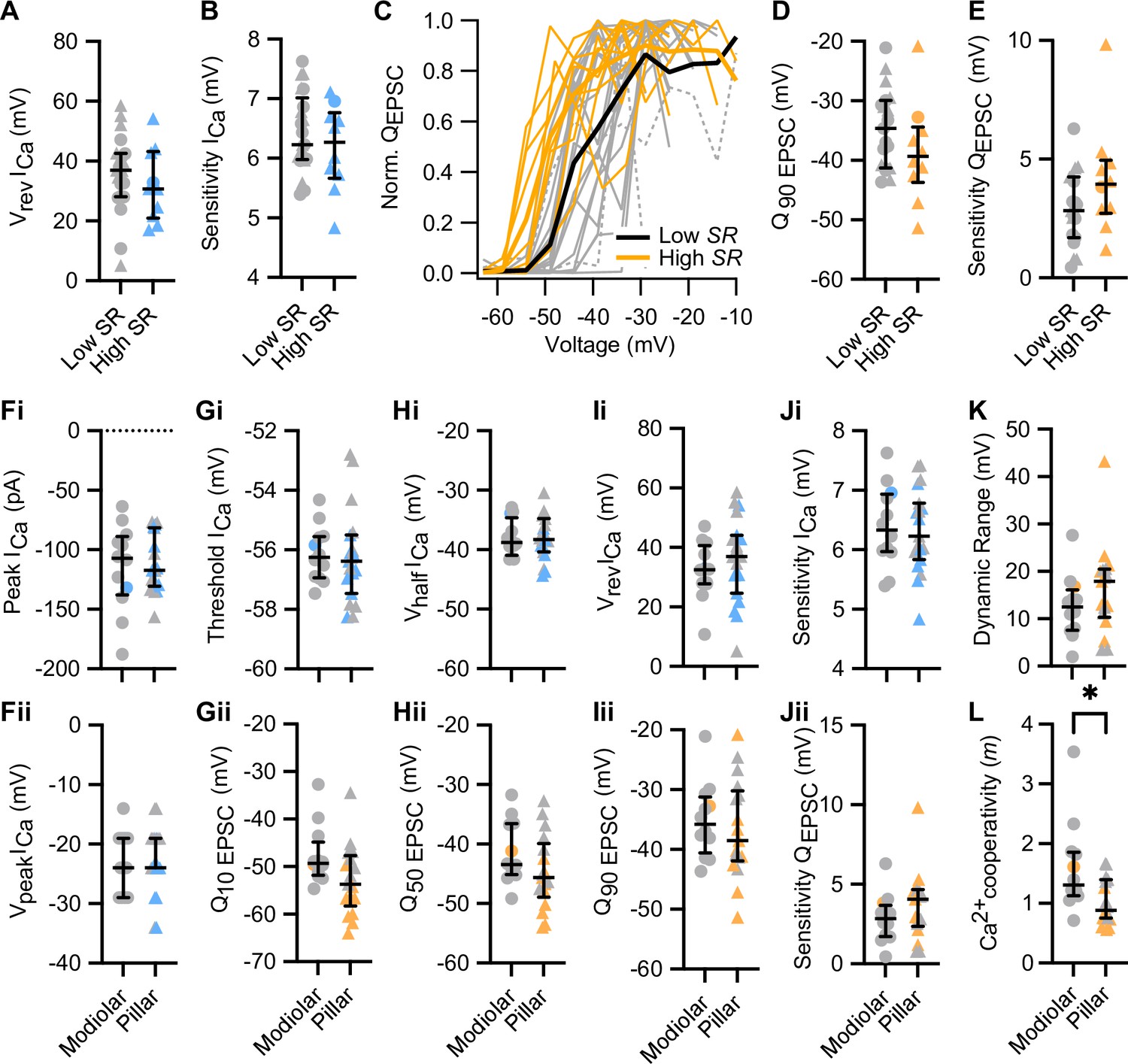
The voltage dependence of synaptic release does not differ significantly between modiolar and pillar synapses, but the Ca2+ dependence does.
(A, B) The reversal potential (A) and the voltage sensitivity of ICa (B) were comparable between low and high SR synapses.(C) Triggered single active zone evoked EPSCs (eEPSCs) (release intensity curves) of 31 pairs of low and high SR. Averages (thick lines) and individual curves (thin lines) are overlaid. The release-intensity curve of two low SR pairs could not be fitted with the sigmoidal function (gray dotted lines). (D–E) Q90 EPSC (D) and voltage sensitivity of the release (E) were comparable between high and low SR synapses. (Fi–Fii) The peak of Ca2+ current (Fi) and the voltage eliciting maximum Ca2+ current (Fii) were comparable between modiolar and pillar synapses. (Gi–Ji) The threshold of Ca2+ influx (Gi), Vhalf ICa (Hi), reversal potential of ICa (Ii), and voltage sensitivity of ICa (Ji) did not differ between modiolar and pillar synapses. (Gii–Jii) Q10 EPSC (Gii), Q50 EPSC (Hii), Q90 EPSC (Iii) and voltage sensitivity of the release (Jii) were comparable between modiolar and pillar synapses. (K) Dynamic range of release was comparable between modiolar and pillar synapse. (L) Ca2+ cooperativity (m) estimated for each individual synapse was significantly lower in pillar synapses. All panels but C show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 3—figure supplement 2
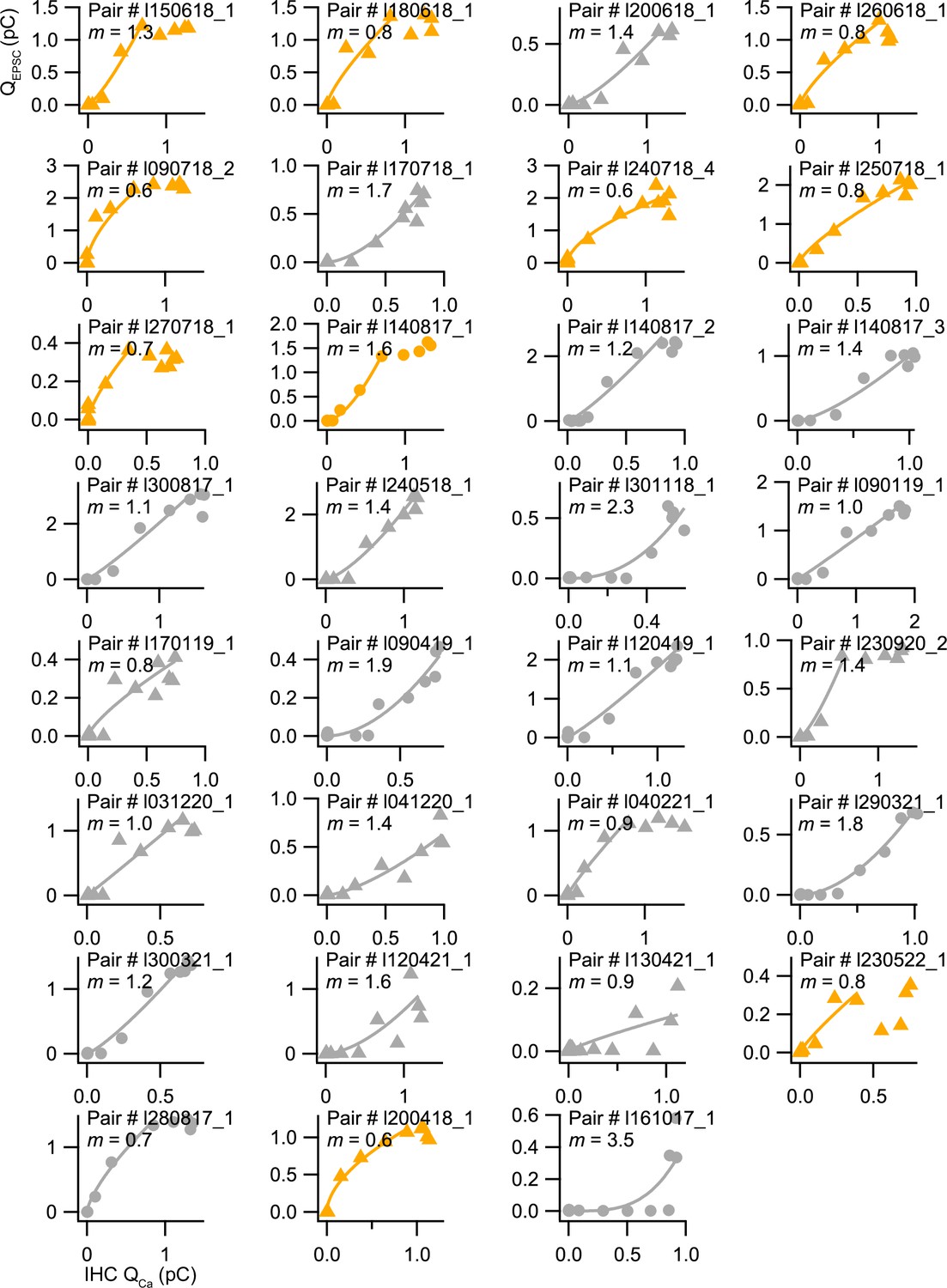
Apparent Ca2+ dependence of neurotransmitter release at individual synapses in the range of inner hair cell (IHC) receptor potentials.
Scatter plots of the EPSC charges (QEPSC) vs the corresponding Ca2+ current integrals (QCa) for each individual synapse in response to 10 ms depolarizations from –58 to –19 mV. The solid line is a least-squares fit of a power function (QEPSC = a(QCa)m) to each pair data. Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 4 with 1 supplement
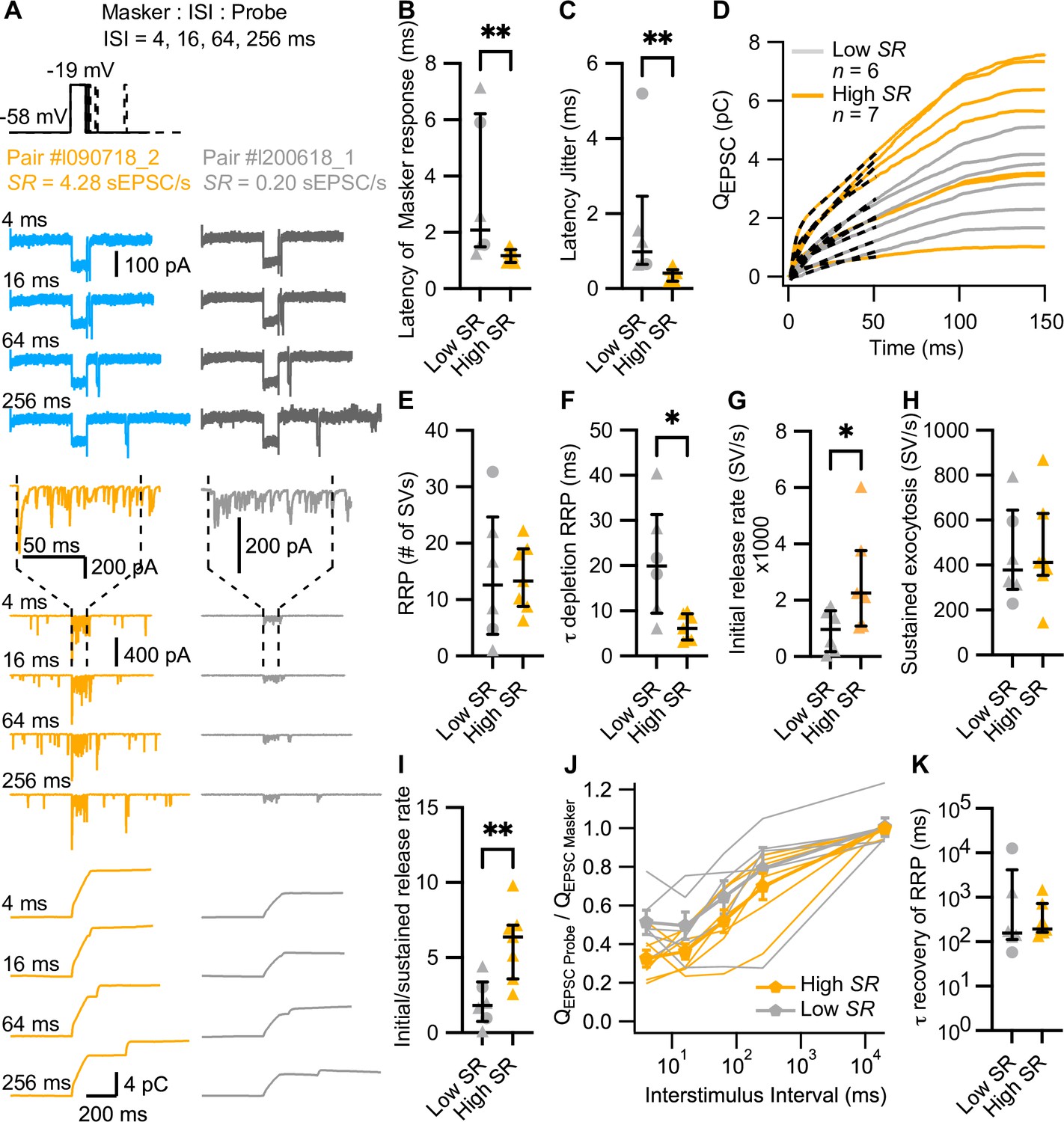
High spontaneous release synapses have shorter synaptic delay and higher initial release rates.
(A) ‘Forward masking’ voltage protocol to study depletion and recovery of RRP and data of an exemplary high SR synapse (left panels, blue and orange) and low SR synapse (right panels, gray): Ca2+ currents (second from top, ICa), evoked EPSCs (eEPSCs) (second from bottom), and QEPSC (bottom). The stimulus (top panel) consists of two sequential voltage steps (‘masker’ and ‘probe’) separated by different interstimulus intervals (ISI). Dashed vertical lines on top of eEPSC data indicate the onset and offset of the masker stimulus. (B) Latencies of the eEPSCs (eEPSConset – Maskeronset) were significantly shorter in high SR than low SR synapses. (C) High SR synapses also had less latency jitter. (D) Pool depletion dynamics were studied by fitting the sum of a single exponential and a line function (black discontinuous line) to the first 50 ms of the average QEPSC trace in response to the masker stimulus. (E–I) RRP, time constant (τ) of depletion, initial release rate, and sustained release were calculated from the fits and the mean QsEPSC for each pair. High SR synapses depleted the RRP with faster time constants (F) and reached higher initial release rates (G) followed by a stronger adaptation (I) (J) Recovery from RRP depletion shown as ratio of QEPSC probe and QEPSC masker (mean ± sem) during the first 10 ms of the stimulus. (K) Time constant of recovery from RRP depletion obtained from single exponential fits to the traces shown in J (see Figure 4—figure supplement 1L). Panels B, C, E–I, and K show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Figure 4—figure supplement 1
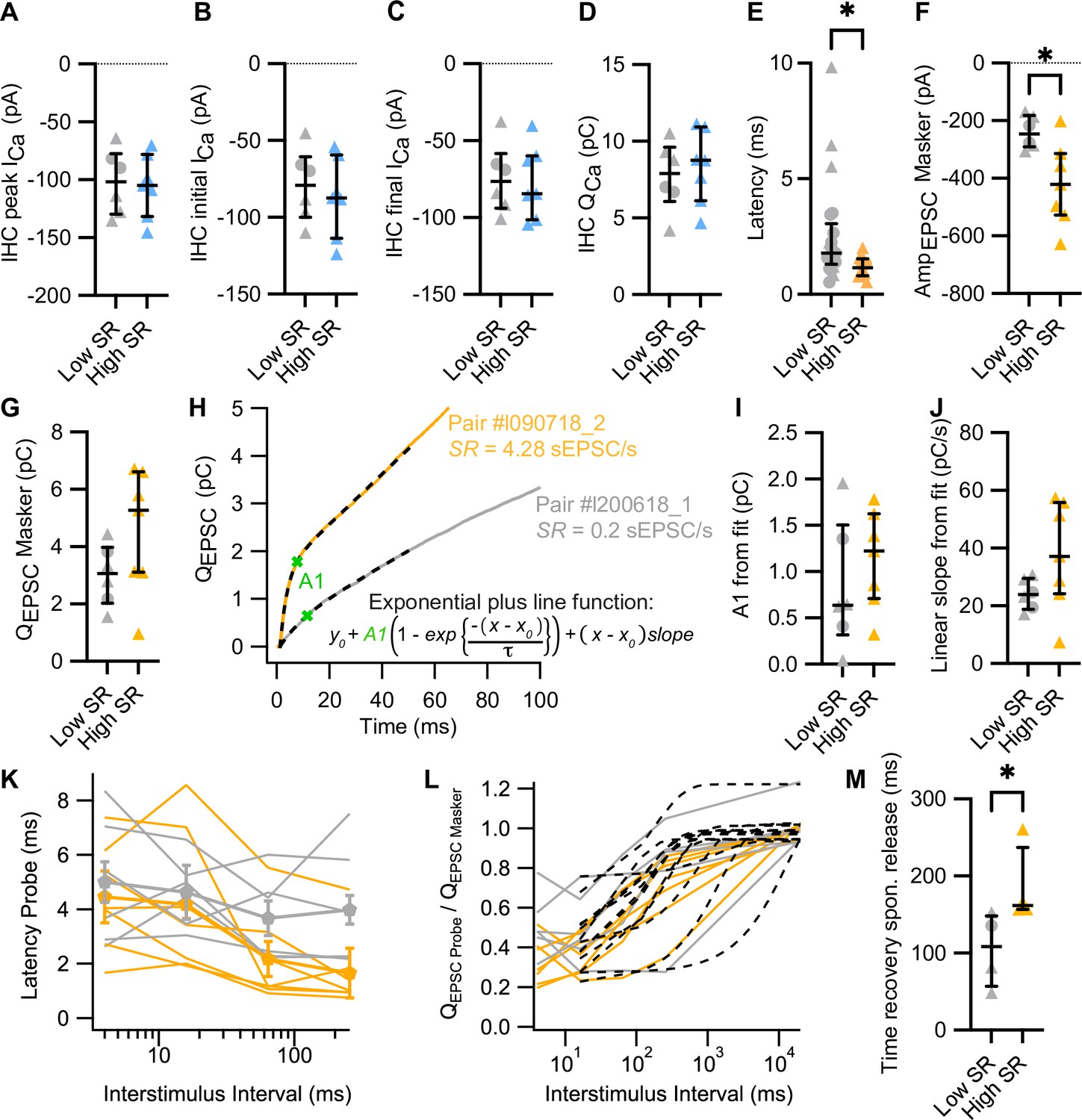
Parameters of synaptic vesicle pool dynamics in forward masking protocols.
(A–D) Presynaptic inner hair cell (IHC) peak (A), initial (B), and final (C) Ca2+ current (ICa) and Ca2+ charge (QCa) during the masker stimuli were comparable regardless the SR of postsynaptic bouton. (E) Latencies of the evoked EPSCs were also significantly shorter in high SR than low SR synapses when compared in a bigger sample size (31 pairs from Figure 4). (F–G) High SR synapses had significantly larger amplitudes of evoked EPSCs in response to the masker stimulus (F), while the EPSC charge (QEPSC) did not differ (G). (H) Exemplary average QEPSC response of a high SR and a low SR synapse to the masker stimuli. We fitted an exponential plus line function to the first 50 ms of the response (discontinuous lines) to study SV pool depletion dynamics. From these fits, we can retrieve information about RRP size (amplitude of the exponential component, A1), RRP depletion time constant (τ of the exponential component), and sustained release (linear slope of the line component). (I–J) Amplitude (I) and linear slope (J) obtained from the fits of the exponential plus line function to the QEPSC from individual pairs (Figure 4E). (K) Latencies of the response to the probe stimuli (eEPSConset – Probeonset) for different ISI. (L) Single exponential fits from 16 to 20,000 ms (black dotted lines) to estimate the recovery kinetics from RRP depletion. (M) The spontaneous release after the probe offset recovered slower in high SR synapses. Panels A–G, I, J, M show individual data points with the median and interquartile range overlaid (line). Synapses were classified as △ pillar or ❍ modiolar, and as Low SR<1 sEPSC/s≤High SR.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Mus musculus) | C57BL/6N | Jackson Laboratory (https://www.jax.org) | RRID:IMSR_JAX:005304 | |
| Software, algorithm | Patchmaster | HEKA Elektronik, (http://www.heka.com/products/products_main.html#soft_pm) | RRID:SCR_000034: | |
| Software, algorithm | Igor Pro software package | WaveMetrics (http://www.wavemetrics.com/products/igorpro/igorpro.htm) | RRID:SCR_000325 | |
| Software, algorithm | Patchers Power Tools | Igor Pro XOP (http://www3.mpibpc.mpg.de/groups/neher/index.php?page=software) | RRID:SCR_001950 | |
| Software, algorithm | NeuroMatic | ThinkRandom (http://www.neuromatic.thinkrandom.com/) | RRID:SCR_004186 | |
| Software, algorithm | Excel | Microsoft (https://www.microsoft.com/en-gb/) | RRID:SCR_016137 | |
| Software, algorithm | GraphPad Prism software | GraphPad Prism (https://graphpad.com) | RRID:SCR_002798 | |
| Software, algorithm | Adobe Illustrator | Adobe (http://www.adobe.com/products/illustrator.html) | RRID:SCR_010279 |
Additional files
-
MDAR checklist
- https://cdn.elifesciences.org/articles/93749/elife-93749-mdarchecklist1-v1.pdf
-
Supplementary file 1
Total time for spontaneous rate calculation.
- https://cdn.elifesciences.org/articles/93749/elife-93749-supp1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Bridging the gap between presynaptic hair cell function and neural sound encoding
eLife 12:RP93749.
https://doi.org/10.7554/eLife.93749.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}