Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorJohn TuthillUniversity of Washington, Seattle, United States of America
- Senior EditorAlbert CardonaUniversity of Cambridge, Cambridge, United Kingdom
Reviewer #1 (Public review):
Summary:
The study by McKim et al seeks to provide a comprehensive description of the connectivity of neurosecretory cells (NSCs) using a high-resolution electron microscopy dataset of the fly brain and several single-cell RNA seq transcriptomic datasets from the brain and peripheral tissues of the fly. They use connectomic analyses to identify discrete functional subgroups of NSCs and describe both the broad architecture of the synaptic inputs to these subgroups as well as some of the specific inputs including from chemosensory pathways. They then demonstrate that NSCs have very few traditional presynapses consistent with their known function as providing paracrine release of neuropeptides. Acknowledging that EM datasets can't account for paracrine release, the authors use several scRNAseq datasets to explore signaling between NSCs and characterize widespread patterns of neuropeptide receptor expression across the brain and several body tissues. The thoroughness of this study allows it to largely achieve it's goal and provides a useful resource for anyone studying neurohormonal signaling.
Strengths:
The strengths of this study are the thorough nature of the approach and the integration of several large-scale datasets to address short-comings of individual datasets. The study also acknowledges the limitations that are inherent to studying hormonal signaling and provides interpretations within the the context of these limitations.
Weaknesses:
Overall, the framing of this paper needs to be shifted from statements of what was done to what was found. Each subsection, and the narrative within each, is framed on topics such as "synaptic output pathways from NSC" when there are clear and impactful findings such as "NSCs have sparse synaptic output". Framing the manuscript in this way allows the reader to identify broad takeaways that are applicable to other model system. Otherwise, the manuscript risks being encyclopedic in nature. An overall synthesis of the results would help provide the larger context within which this study falls.
The cartoon schematic in Figure 5A (which is adapted from a 2020 review) has an error. This schematic depicts uniglomerular projection neurons of the antennal lobe projecting directly to the lateral horn (without synapsing in the mushroom bodies) and multiglomerular projection neurons projecting to the mushroom bodies and then lateral horn. This should be reversed (uniglomerular PNs synapse in the calyx and then further project to the LH and multiglomerular PNs project along the mlACT directly to the LH) and is nicely depicted in a Strutz et al 2014 publication in eLife.
Reviewer #2 (Public review):
Summary:
The authors aim to provide a comprehensive description of the neurosecretory network in the adult Drosophila brain. They sought to assign and verify the types of 80 neurosecretory cells (NSCs) found in the publicly available FlyWire female brain connectome. They then describe the organization of synaptic inputs and outputs across NSC types and outline circuits by which olfaction may regulate NSCs, and by which Corazon-producing NSCs may regulate flight behavior. Leveraging existing transcriptomic data, they also describe the hormone and receptor expressions in the NSCs and suggest putative paracrine signaling between NSCs. Taken together, these analyses provide a framework for future experiments, which may demonstrate whether and how NSCs, and the circuits to which they belong, may shape physiological function or animal behavior.
Strengths:
This study uses the FlyWire female brain connectome (Dorkenwald et al. 2023) to assign putative cell types to the 80 neurosecretory cells (NSCs) based on clustering of synaptic connectivity and morphological features. The authors then verify type assignments for selected populations by matching cluster sizes to anatomical localization and cell counts using immunohistochemistry of neuropeptide expression and markers with known co-expression.
The authors compare their findings to previous work describing the synaptic connectivity of the neurosecretory network in larval Drosophila (Huckesfeld et al., 2021), finding that there are some differences between these developmental stages. Direct comparisons between adults and larvae are made possible through direct comparison in Table 1, as well as the authors' choice to adopt similar (or equivalent) analyses and data visualizations in the present paper's figures.
The authors extract core themes in NSC synaptic connectivity that speak to their function: different NSC types are downstream of shared presynaptic outputs, suggesting the possibility of joint or coordinated activation, depending on upstream activity. NSCs receive some but not all modalities of sensory input. NSCs have more synaptic inputs than outputs, suggesting they predominantly influence neuronal and whole-body physiology through paracrine and endocrine signaling.
The authors outline synaptic pathways by which olfactory inputs may influence NSC activity and by which Corazon-releasing NSCs may regulate flight. These analyses provide a basis for future experiments, which may demonstrate whether and how such circuits shape physiological function or animal behavior.
The authors extract expression patterns of neuropeptides and receptors across NSC cell types from existing transcriptomic data (Davie et al., 2018) and present the hypothesis that NSCs could be interconnected via paracrine signaling. The authors also catalog hormone receptor expression across tissues, drawing from the Fly Cell Atlas (Li et al., 2022).
Weaknesses:
The clustering of NSCs by their presynaptic inputs and morphological features, along with corroboration with their anatomical locations, distinguished some, but not all cell types. The authors attempt to distinguish cell types using additional methodologies: immunohistochemistry (Figure 2), retrograde trans-synaptic labeling, and characterization of dense core vesicle characteristics in the FlyWire dataset (Figure 1, Supplement 1). However, these corroborating experiments often lacked experimental replicates, were not rigorously quantified, and/or were presented as singular images from individual animals or even individual cells of interest. The assignments of DH44 and DMS types remain particularly unconvincing.
The authors present connectivity diagrams for visualization of putative paracrine signaling between NSCs based on their peptide and receptor expression patterns. These transcriptomic data alone are inadequate for drawing these conclusions, and these connectivity diagrams are untested hypotheses rather than results. The authors do discuss this in the Discussion section.
Reviewer #3 (Public review):
Summary:
The manuscript presents an ambitious and comprehensive synaptic connectome of neurosecretory cells (NSC) in the Drosophila brain, which highlights the neural circuits underlying hormonal regulation of physiology and behaviour. The authors use EM-based connectomics, retrograde tracing, and previously characterised single-cell transcriptomic data. The goal was to map the inputs to and outputs from NSCs, revealing novel interactions between sensory, motor, and neurosecretory systems. The results are of great value for the field of neuroendocrinology, with implications for understanding how hormonal signals integrate with brain function to coordinate physiology.
The manuscript is well-written and provides novel insights into the neurosecretory connectome in the adult Drosophila brain. Some, additional behavioural experiments will significantly strengthen the conclusions.
Strengths:
(1) Rigorous anatomical analysis
(2) Novel insights on the wiring logic of the neurosecretory cells.
Weaknesses:
(1) Functional validation of findings would greatly improve the manuscript.
Author response:
Public Reviews:
Reviewer #1 (Public review):
Summary:
The study by McKim et al seeks to provide a comprehensive description of the connectivity of neurosecretory cells (NSCs) using a high-resolution electron microscopy dataset of the fly brain and several single-cell RNA seq transcriptomic datasets from the brain and peripheral tissues of the fly. They use connectomic analyses to identify discrete functional subgroups of NSCs and describe both the broad architecture of the synaptic inputs to these subgroups as well as some of the specific inputs including from chemosensory pathways. They then demonstrate that NSCs have very few traditional presynapses consistent with their known function as providing paracrine release of neuropeptides. Acknowledging that EM datasets can't account for paracrine release, the authors use several scRNAseq datasets to explore signaling between NSCs and characterize widespread patterns of neuropeptide receptor expression across the brain and several body tissues. The thoroughness of this study allows it to largely achieve it's goal and provides a useful resource for anyone studying neurohormonal signaling.
Strengths:
The strengths of this study are the thorough nature of the approach and the integration of several large-scale datasets to address short-comings of individual datasets. The study also acknowledges the limitations that are inherent to studying hormonal signaling and provides interpretations within the the context of these limitations.
Weaknesses:
Overall, the framing of this paper needs to be shifted from statements of what was done to what was found. Each subsection, and the narrative within each, is framed on topics such as "synaptic output pathways from NSC" when there are clear and impactful findings such as "NSCs have sparse synaptic output". Framing the manuscript in this way allows the reader to identify broad takeaways that are applicable to other model system. Otherwise, the manuscript risks being encyclopedic in nature. An overall synthesis of the results would help provide the larger context within which this study falls.
We agree with the reviewer and will replace all the subsection titles as suggested.
The cartoon schematic in Figure 5A (which is adapted from a 2020 review) has an error. This schematic depicts uniglomerular projection neurons of the antennal lobe projecting directly to the lateral horn (without synapsing in the mushroom bodies) and multiglomerular projection neurons projecting to the mushroom bodies and then lateral horn. This should be reversed (uniglomerular PNs synapse in the calyx and then further project to the LH and multiglomerular PNs project along the mlACT directly to the LH) and is nicely depicted in a Strutz et al 2014 publication in eLife.
We thank the reviewer for spotting this error. We will modify the schematic as suggested.
Reviewer #2 (Public review):
Summary:
The authors aim to provide a comprehensive description of the neurosecretory network in the adult Drosophila brain. They sought to assign and verify the types of 80 neurosecretory cells (NSCs) found in the publicly available FlyWire female brain connectome. They then describe the organization of synaptic inputs and outputs across NSC types and outline circuits by which olfaction may regulate NSCs, and by which Corazon-producing NSCs may regulate flight behavior. Leveraging existing transcriptomic data, they also describe the hormone and receptor expressions in the NSCs and suggest putative paracrine signaling between NSCs. Taken together, these analyses provide a framework for future experiments, which may demonstrate whether and how NSCs, and the circuits to which they belong, may shape physiological function or animal behavior.
Strengths:
This study uses the FlyWire female brain connectome (Dorkenwald et al. 2023) to assign putative cell types to the 80 neurosecretory cells (NSCs) based on clustering of synaptic connectivity and morphological features. The authors then verify type assignments for selected populations by matching cluster sizes to anatomical localization and cell counts using immunohistochemistry of neuropeptide expression and markers with known co-expression.
The authors compare their findings to previous work describing the synaptic connectivity of the neurosecretory network in larval Drosophila (Huckesfeld et al., 2021), finding that there are some differences between these developmental stages. Direct comparisons between adults and larvae are made possible through direct comparison in Table 1, as well as the authors' choice to adopt similar (or equivalent) analyses and data visualizations in the present paper's figures.
The authors extract core themes in NSC synaptic connectivity that speak to their function: different NSC types are downstream of shared presynaptic outputs, suggesting the possibility of joint or coordinated activation, depending on upstream activity. NSCs receive some but not all modalities of sensory input. NSCs have more synaptic inputs than outputs, suggesting they predominantly influence neuronal and whole-body physiology through paracrine and endocrine signaling.
The authors outline synaptic pathways by which olfactory inputs may influence NSC activity and by which Corazon-releasing NSCs may regulate flight. These analyses provide a basis for future experiments, which may demonstrate whether and how such circuits shape physiological function or animal behavior.
The authors extract expression patterns of neuropeptides and receptors across NSC cell types from existing transcriptomic data (Davie et al., 2018) and present the hypothesis that NSCs could be interconnected via paracrine signaling. The authors also catalog hormone receptor expression across tissues, drawing from the Fly Cell Atlas (Li et al., 2022).
Weaknesses:
The clustering of NSCs by their presynaptic inputs and morphological features, along with corroboration with their anatomical locations, distinguished some, but not all cell types. The authors attempt to distinguish cell types using additional methodologies: immunohistochemistry (Figure 2), retrograde trans-synaptic labeling, and characterization of dense core vesicle characteristics in the FlyWire dataset (Figure 1, Supplement 1). However, these corroborating experiments often lacked experimental replicates, were not rigorously quantified, and/or were presented as singular images from individual animals or even individual cells of interest. The assignments of DH44 and DMS types remain particularly unconvincing.
We thank the reviewer for this comment. We would like to clarify that the images presented in Figure 2 and Figure 1 Supplement 1 are representative images based on at least 5 independent samples. We will clarify this in the figure caption and methods. The electron micrographs showing dense core vesicle (DCV) characteristics (Figure 1 Supplement E-G) are also representative images based on examination of multiple neurons. However, we agree with the reviewer that a rigorous quantification would be useful to showcase the differences between DCVs from NSC subtypes. Therefore, we have now performed a quantitative analysis of the DCVs in putative m-NSCDH44 (n=6), putative m-NSCDMS (n=6) and descending neurons (n=4) known to express DMS. For consistency, we examined the cross section of each cell where the diameter of nuclei was the largest. We quantified the mean gray value of at least 50 DCV per cell. Our analysis shows that mean gray values of putative m-NSCDMS and DMS descending neurons are not significantly different, whereas the mean gray values of m-NSCDH44 are significantly larger. This analysis is in agreement with our initial conclusion.
Author response image 1.
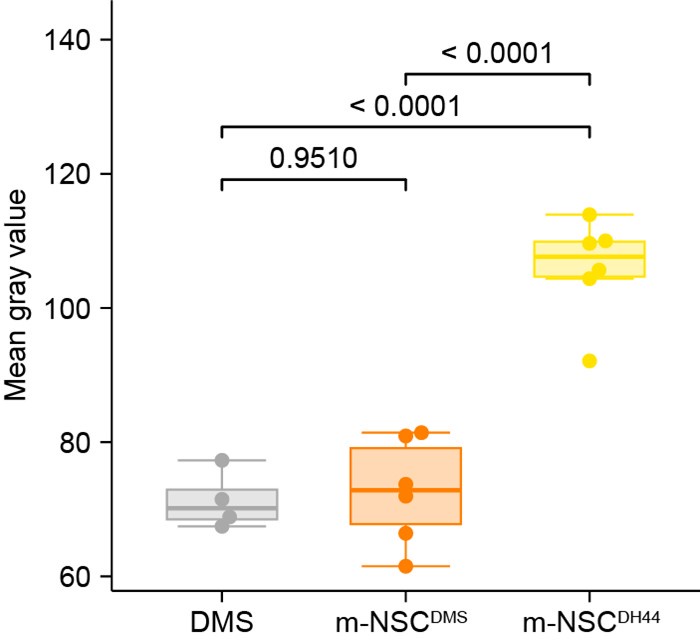
The authors present connectivity diagrams for visualization of putative paracrine signaling between NSCs based on their peptide and receptor expression patterns. These transcriptomic data alone are inadequate for drawing these conclusions, and these connectivity diagrams are untested hypotheses rather than results. The authors do discuss this in the Discussion section.
We fully agree with the reviewer and will further elaborate on the limitations of our approach in the revised manuscript. However, there is a very high-likelihood that a given NSC subtype can signal to another NSC subtype using a neuropeptide if its receptor is expressed in the target NSC. This is due to the fact that all NSC axons are part of the same nerve bundle (nervi corpora cardiaca) which exits the brain. The axons of different NSCs form release sites that are extremely close to each other. Neuropeptides from these release sites can easily diffuse via the hemolymph to peripheral tissues that (e.g. fat body and ovaries) that are much further away from the release sites on neighboring NSCs. We believe that neuropeptide receptors are expressed in NSCs near these release sites where they can receive inputs not just from the adjacent NSCs but also from other sources such as the gut enteroendocrine cells. Hence, neuropeptide diffusion is not a limiting factor preventing paracrine signaling between NSCs and receptor expression is a good indicator for putative paracrine signaling.
Reviewer #3 (Public review):
Summary:
The manuscript presents an ambitious and comprehensive synaptic connectome of neurosecretory cells (NSC) in the Drosophila brain, which highlights the neural circuits underlying hormonal regulation of physiology and behaviour. The authors use EM-based connectomics, retrograde tracing, and previously characterised single-cell transcriptomic data. The goal was to map the inputs to and outputs from NSCs, revealing novel interactions between sensory, motor, and neurosecretory systems. The results are of great value for the field of neuroendocrinology, with implications for understanding how hormonal signals integrate with brain function to coordinate physiology.
The manuscript is well-written and provides novel insights into the neurosecretory connectome in the adult Drosophila brain. Some, additional behavioural experiments will significantly strengthen the conclusions.
Strengths:
(1) Rigorous anatomical analysis
(2) Novel insights on the wiring logic of the neurosecretory cells.
Weaknesses:
(1) Functional validation of findings would greatly improve the manuscript.
We agree with this reviewer that assessing the functional output from NSCs would improve the manuscript. Given that we currently lack genetic tools to measure hormone levels and that behaviors and physiology are modulated by NSCs on slow timescales, it is difficult to assess the immediate functional impact of the sensory inputs to NSC using approaches such as optogenetics. However, since l-NSCCRZ are the only known cell type that provide output to descending neurons, we will functionally test this output pathway using different behavioral assays recommended by this reviewer.



