Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorKiyoshi TakedaThe University of Osaka, Osaka, Japan
- Senior EditorWendy GarrettHarvard T.H. Chan School of Public Health, Boston, United States of America
Reviewer #1 (Public review):
Summary:
The authors tried to identify the relationships between gut microbiota, lipid metabolites and the host in type 2 diabetes (T2DM) by using spontaneously developed T2DM in macaques, considered among the best human models.
Strengths:
The authors compared comprehensively the gut microbiota, plasma fatty acids between spontaneous T2DM and the control macaques, and tried verified the results with macaques in high-fat diet-fed mice model.
Weaknesses:
The observed multi-omics on macaques can be done on humans, which weakens the conclusion of the manuscript, unless the observation/data on macaques could cover during the onset of T2DM that would be difficult to obtain from humans.
Regarding the metabolomic analysis on fatty acids, the authors did not include the results obtained form the macaque fecal samples which should be important considering the authors claimed the importance of gut microbiota in the pathogenesis of T2DM. Instead, the authors measured palmitic acid in the mouse model and tried to validate their conclusions with that.
In murine experiments, palmitic acid-containing diet were fed to mice to induce diabetic condition, but this does not mimic spontaneous T2DM in macaques, since the authors did not measure in macaque feces (or at least did not show the data from macaque feces of) palmitic acid or other fatty acids; instead, they assumed from blood metabolome data that palmitic acid would be absorbed from the intestine to affect the host metabolism, and added palmitic acid in the diet in mouse experiments. Here involves the probable leap of logic to support their conclusions and title of the study.
In addition, the authors measured omics data after, but not before, the onset of spontaneous T2DM of macaques. This can reveal microbiota dysbiosis driven purely by disease progression, but does not support the causative effect of gut microbiota on T2DM development that the authors claims.
Author response:
The following is the authors’ response to the original reviews
Reviewer #1 (Public review):
Summary:
The authors tried to identify the relationships among the gut microbiota, lipid metabolites, and the host in type 2 diabetes (T2DM) by using macaques that spontaneously develop T2DM, considered one of the best models of the human disease.
Strengths:
The authors comprehensively compared the gut microbiota and plasma fatty acids between macaques with spontaneous T2DM and control macaques and verified the results with macaques on a high-fat diet-fed mice model.
Weaknesses:
Comment 1: The observed multi-omics of the macaques can be done on humans, which weakens the impact of the conclusion of the manuscript.
We fully acknowledge the critical role of human studies in T2DM research. In our study, the spontaneous T2DM macaque model provided a unique window to address inherent challenges in human studies, including medication interference and environmental heterogeneity. Human studies have struggled to standardize confounding factors such as diet, exercise, and antibiotic use. Moreover, most human T2DM patients receive long-term glucose-lowering medications (e.g., metformin), which directly alter gut microbiota composition and function, masking disease-associated microbial signatures (Sun et al., 2018; Petakh et al., 2023). In contrast, the spontaneous T2DM macaques, untreated with glucose-lowering drugs or antibiotics under strictly controlled conditions, revealed microbiota dysbiosis driven purely by disease progression. Our work bridged the gap between rodent studies and human clinical trials, providing an important clinical reference for guiding targeted interventions, particularly microbiota modulation. We sincerely appreciate the valuable comments. We have added background to the part of the introduction, “In fact, T2DM macaques avoid medication interference and environmental heterogeneity under controlled experimental conditions, and share key pathological features with humans, such as amyloidosis of pancreatic islets, which is absent in mouse models (25, 26), suggesting that T2DM macaques are the optimal animal model for simulating human T2DM and its complications (27).” (Lines 98-103).
References:
Sun L., Xie C., Wang G., Wu Y., Wu Q., Wang X., Liu J., Deng Y., Xia J., et al. 2018) Gut microbiota and intestinal FXR mediate the clinical benefits of metformin Nat. Med 24:1919-1929 https://doi.org/10.1038/s41591-018-0222-4
Petakh P., Kamyshna I., Kamyshnyi A 2023) Effects of metformin on the gut microbiota: A systematic review Mol. metab 77:101805-101805 https://doi.org/10.1016/j.molmet.2023.101805
Comment 2: In addition, the age and sex of the control macaque group did not necessarily match those of the T2DM group, leaving the possibility for compromising the analysis.
Thank you for pointing this out. The availability of spontaneous T2DM macaques is very limited. Wang et al. (2018) identified only nine diabetic macaques among 2,000 screened, and our prior study (Jiang et al., 2022) found merely seven diabetic cases in 1,408 macaques. In this work, we obtained eight spontaneous T2DM macaques with FPG ≥ 7 mmol/L and eight heathy control macaques with FPG ≤ 6.1 mmol/L (three consecutive detections, each detection interval of one month) from a population of 1,698 captive macaques. To avoid confound factors affect the investigated macaques, all macaques were individually housed with standardized diets and environmental controls. While age and sex partially matched, controls originated from the same population to minimize confounding. The T2DM and control groups were matched for age period (5 adult and 3 elder) and had comparable mean ages (mean age of T2DM individuals = 12.88, mean age of control individuals = 11.25) (Table S1). In terms of gender matching, we compared blood metabolome data of 12 healthy adult female and 12 healthy adult male macaques from another study (Liu et al., 2023) and obtained only a small number of differential metabolites that were not associated with tryptophan (Table 1). We acknowledge this limitation and will prioritize matched controls in future studies.
Author response table 1.
List of all differential metabolites.

References:
Wang J., Xu S., Gao J., Zhang L., Zhang Z., Yang W., Li Y., Liao S., Zhou H., Liu P., et al. 2018) SILAC-based quantitative proteomic analysis of the livers of spontaneous obese and diabetic rhesus monkeys Am. J. Physiol-endoc. M 315:E29-E306 https://doi.org/10.1152/ajpendo.00016.2018
Jiang C., Pan X., Luo J., Liu X., Zhang L., Liu Y., Lei G., Hu G., Li J 2022) Alterations in microbiota and metabolites related to spontaneous diabetes and pre-diabetes in rhesus macaques Genes 13:1513 https://doi.org/10.3390/genes13091513
Liu X., Liu X.Y., Wang X.Q., Shang K., Li J.W., Lan Y., Wang J., Li J., et al. 2023). Multi-Omics Analysis Reveals Changes in Tryptophan and Cholesterol Metabolism before and after Sexual Maturation in Captive Macaques BMC Genomics 24:308. https://doi.org/10.1186/s12864-023-09404-3
Comment 3: Regarding the metabolomic analysis, the authors did not include fecal samples which are important, considering the authors' claim about the importance of gut microbiota in the pathogenesis of T2DM.
We thank the reviewer for this suggestion. This study employed untargeted metabolomics on macaque fecal samples to identify metabolites associated with spontaneously developing T2DM. To validate the metabolites identified through the untargeted metabolomic analysis, we conducted targeted medium- and long-chain fatty acid (MLCFA) metabolomics on macaque serum, and we further quantitatively examined the content of palmitic acid (PA) in mice feces, ileum, and serum. Although targeted MLCFA metabolomics was not performed on macaque fecal samples, we performed untargeted metabolomics on macaque feces and confirmed the contribution of PA in mice that underwent fecal microbiota transplantation (FMT) from T2DM macaques. We have added future expectations in the part of the discussion, “Previous studies have shown that insulin-resistant patients exhibit increased fecal monosaccharides associated with microbial carbohydrate metabolism (70). Furthermore, commensal species of Lachnospiraceae actively overproduce long-chain fatty acids during metabolic dysfunction through altered bacterial lipid metabolism. The microbe-derived fatty acids impair intestinal epithelial integrity to exacerbate metabolic dysregulation (71). Given that microbial metabolic activity causally modulates host metabolic homeostasis, the content change of PA was potentially associated with a dynamic equilibrium between host absorption and microbial metabolism. Further integrative studies on the fecal fatty acid metabolome, microbial PA metabolism, and functional pathways will be crucial for delineating causal links between dysbiosis and lipid metabolic dysfunction in T2DM.” (Lines 426-437).
Comment 4: In the mouse experiments, the control group should be given a FMT from control macaques rather than just untreated SPF mice since the fecal microbiota composition is likely very different between macaques and mice.
Thanks for your helpful suggestion. We recognized the importance of a FMT control group and supplemented mouse experiments (using the C57BL/6J strain) with FMT from control macaques (HFT group). Another group of mice without FMT was set as control. Due to the lengthy experimental period, observations were concluded at 30 days post-FMT. We compared changes in the gut microbiota before and after antibiotic treatment in mice (-14D and 0D), and tracked body weight and fasting plasma glucose (FPG) levels from day -14 to day 30. At 30 days after FMT, fecal samples from all groups were collected for 16S rRNA sequencing. Additionally, samples of T2DM microbiota transplant (TP), and control transplant (HTP) were sequenced. Finally, we integrated the 16S sequencing data from the FTPA group (palmitic acid (PA) diet and FMT from T2DM macaques) and FT group (normal diet and FMT from T2DM macaques) at day 30 for combined analysis. The results showed that the antibiotic treatment used in this study effectively depleted the gut microbiota. Following FMT, gut microbial diversity stabilized within 30 days, with similar microbial community proportions between HFT and control groups. Core functional groups of the healthy microbiota (Bacteroidota and Bacillota) stably colonized mice despite host species divergence, confirming that T2DM phenotypes originate specifically from macaque microbiota. Importantly, increased abundance of Lachnospiraceae (including genera Ruminococcus (current name: Mediterraneibacter), Coprococcus, and Clostridium) and the key species Ruminococcus gnavus (current name: Mediterraneibacter gnavus) were also observed in FT group versus HFT group on day 30, validating our original findings. We have added findings in the results, “To eliminate interference from host species divergence in gut microbiota composition, we supplemented mouse experiments using FMT from control macaques (HFT group) (Figure S4A). By day 30, the HFT group exhibited significantly lower body weight than the untreated control group (p < 0.05) (Figure S4B). Throughout the experimental period, FPG levels in both HFT and control groups remained within the normal range (< 6 mmol/L) without significant differences, indicating that transplantation of control macaque microbiota did not induce glycemic alterations (Figure S4C).” (Lines 276-283), and “Integrating 16S rRNA sequencing data from the HFT, FT, and FTPA groups showed that the antibiotic treatment effectively depleted the gut microbiota, resulting in microbial diversity decreased sharply, with the dominant phyla shifting from Bacteroidota and Bacillota to Pseudomonadota (Figure S4D-G). The HFT group restored microbial diversity within 30 days, achieving community proportions comparable to untreated controls. Core functional phyla (Bacteroidota and Bacillota) stably colonized in HFT group (Figure S4D-I). Critically, FT and FTPA groups exhibited increased Lachnospiraceae (including genera Ruminococcus (current name: Mediterraneibacter), Coprococcus, and Clostridium) compared with the HFT group on day 30. In addition, LEfSe comparison identified significant R. gnavus (current name: M. gnavus) enrichment in the FT group (LDA > 3, p < 0.01) (Figure S4J-M).” (Lines 324-334, 825-837). Specifically:
(1) Experimental design: transplant preparation and FMT from control macaques
After single cage feeding and FPG detection, fecal samples from three control macaques were collected and mixed for transplantation preparation. Then, 4 ml diluent (Berland et al., 2021) was added per gram of feces. Sodium L-ascorbic acid (5% (w/v)) and L-cysteine hydrochloride monohydrate (0.1% (w/v)) were added to all suspensions (The sterile diluent of control group was added with the same amount of reagent). The mixture was homogenized and filtered sequentially through 200, 400, and 800 μm sterile mesh screens. The filtrate was centrifuged (600 × g, 5 min), and supernatants were aliquoted (400 μL/tube) for storage at -80°C. For use, the transplant was quickly thawed in a 37℃ water bath.
Specific-pathogen-free male C57BL/6J mice aged 6 weeks were randomized into control and HFT (receiving FMT from control macaques) groups. Mice received antibiotic water (ampicillin, neomycin sulfate, and metronidazole, 1 g/L each) from days -14 to 0. All mice were maintained under standard conditions (12h light/dark, 22-25°C, 40-60% humidity) with sterile diet and twice-daily water changes. Body weight, fasting plasma glucose (FPG) were monitored, and fecal samples were collected throughout the study, with fecal 16S rRNA sequencing performed (Figure S4). The study was approved by the Ethics Committee of College of Life Sciences, Sichuan University, and conducted in accordance with the local legislation and institutional requirements.
(2) Results
Body weight monitoring revealed no significant difference between HFT and control groups before (-14D) and after (0D) antibiotic treatment. By day 30, the HFT group exhibited significantly lower body weight than the untreated control group (p < 0.05) (Figure S4B). Throughout the experimental period, FPG levels in both HFT and control groups remained within the normal range (< 6 mmol/L) without significant differences, indicating that transplantation of control macaque microbiota did not induce glycemic alterations (Figure S4C).
Shannon and Simpson indices showed a significant reduction in gut microbiota diversity after antibiotic treatment (0D) (p < 0.01) (Figure S4D,E). The intestinal microbiota of normal mice (-14D) was predominantly composed of Bacteroidota and Bacillota. After two weeks of antibiotic treatment (0D), microbial diversity decreased sharply compared to the -14D group, with the dominant phyla shifting from Bacteroidota and Bacillota to Pseudomonadota (Author response image 1A; Figure S4L). In healthy gut homeostasis, obligate anaerobes such as Bacillota and Bacteroidota maintain intestinal equilibrium. Antibiotic disruption induced dysbiosis in mice, causing substantial restructuring of fecal microbial composition. During dysbiosis, colon epithelial cells shift to anaerobic glycolysis for energy production, increasing epithelial oxygenation and driving expansion of facultative anaerobic Pseudomonadota (de Nies et al., 2023; Szajewska et al., 2024).
NMDS analysis of integrated 16S rRNA sequencing data of FTPA30D (PA diet and FMT from T2DM macaques) and FT30D (normal diet and FMT from T2DM macaques) revealed high intra-group repeatability among pre-antibiotic (-14D), post-antibiotic (0D), HFT30D, T2DM microbiota transplant (TP), and control transplant (HTP) groups. The 0D group showed maximal separation from other clusters, while the -14D, control30D, and HFT30D clustered closely together, with HFT30D nearest to control30D (Figure S4F). On the day 30, all groups showed restoration of microbiota community structure, and the composition of gut microbiota in HFT30D was basically consistent with the control30D group at all taxonomic levels (Author response image 1A-C). At the phylum level, HFT30D group showed significantly reduced relative abundance of Pseudomonadota and increased abundance of Bacteroidota, Bacillota_A, Bacillota_I, and gut barrier-enhancing Verrucomicrobiota (Author response image 1A). These findings demonstrated that FMT from control macaques effectively restored the gut microbiota of antibiotic-treated mice toward a normative state.
Author response image 1.
Composition of gut microbiota in mice. (A) Phylum level; (B) Family level; (C) Genus level.
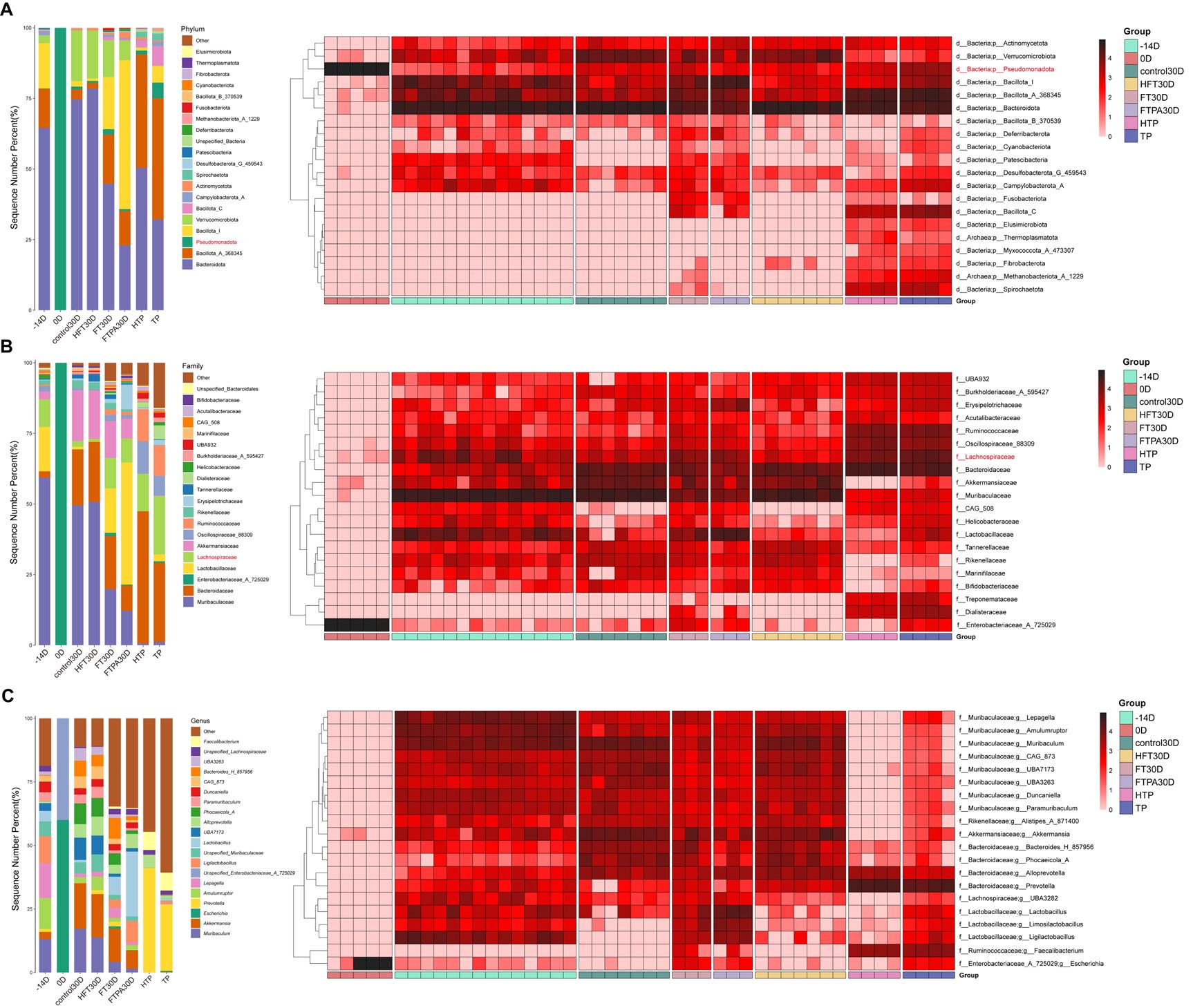
At the phylum level, the FT30D and FTPA30D groups exhibited lower proportions of Bacteroidota/Bacillota compared to the HFT30D (Author response image 1A). Family-level analysis revealed markedly increased abundance of Lactobacillaceae and Lachnospiraceae in FTPA30D and FT30D groups relative to HFT30D, consistent with the changes in the microbiota of spontaneously T2DM macaques (Author response image 1B). Notably, while both HTP and TP groups contained Lachnospiraceae, only FT30D and FTPA30D mice demonstrated significant increase of this family, which was close to that in TP group. Although Muribaculaceae and Bacteroidaceae showed partial recovery in these groups, their relative abundances remained substantially lower than in control30D and HFT30D groups, suggesting that microbiota transplantation from T2DM macaques may reduce specific beneficial taxa while promoting expansion of conditionally pathogenic or metabolically-altered bacteria, such as Lachnospiraceae.
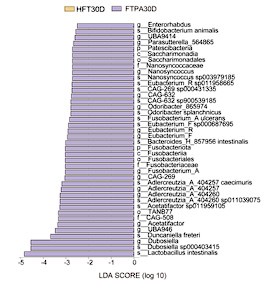
Further analysis of Lachnospiraceae dynamics revealed that at the genus level, most Lachnospiraceae members exhibited higher abundance in the TP group compared to the HTP group. FT30D and FTPA30D groups showed increased abundance of Ruminococcus (current name: Mediterraneibacter), Coprococcus, and Clostridium relative to HFT30D group, consistent with prior analyses (Figure S4). LEfSe comparison between FT30D and HFT30D identified significantly enriched Ruminococcus gnavus (current name: Mediterraneibacter gnavus) in FT30D recipients (LDA > 3, p < 0.01), corroborating earlier findings (Figure S4L). As a mucin-degrading microbe, R. gnavus (current name: M. gnavus) promotes insulin resistance through modulation of tryptamine/phenethylamine levels (Zhai et al., 2023) and exhibits pro-inflammatory properties (Henke et al., 2019; Paone and Cani, 2020). The absence of R. gnavus (current name: M. gnavus) enrichment in FTPA30D was potentially related to differential long-term impacts of T2DM microbiota transplantation across the 30- versus 120-day experimental timelines.
Author response image 2.
Identification of differential microbiota in mice. (A) Linear discriminant analysis Effect Size (LEfSe) analysis between pre-antibiotic (-14D) and post-antibiotic (0D) groups; (B) HFT and FTPA groups; (C) HFT and FT groups.
References:
Berland M., Cadiou J., Levenez F., Galleron N., Quinquis B., Thirion F., Gauthier F., Le ChatelierE., Plaza Oñate F., Schwintner C., et al. 2021) High engraftment capacity of frozen ready-to-use human fecal microbiota transplants assessed in germ-free mice Sci. Rep 11 https://doi.org/10.1038/s41598-021-83638-7
Szajewska H., Scott KP., Meij T de., Forslund-Startceva S.K., Knight R., Koren O., Little P., Johnston B.C., Łukasik J., Suez J., Tancredi D.J., Sanders M.E 2024) Antibiotic-perturbed microbiota and the role of probiotics Nat. Rev. Gastro. Hepat 1-18 https://doi.org/10.1038/s41575-024-01023-x
de Nies L., Kobras C.M., Stracy M 2023) Antibiotic-induced collateral damage to the microbiota and associated infections. Nat. Rev. Microbiol 21:789-804 https://doi.org/10.1038/s41579-023-00936-9
Zhai L., Xiao H., Lin C., Wong H.L.X., Lam Y.Y., Gong M., Wu G., Ning Z., Huang C., Zhang Y., et al. 2023) Gut microbiota-derived tryptamine and phenethylamine impair insulin sensitivity in metabolic syndrome and irritable bowel syndrome Nat. Commun 14 https://doi.org/10 .1038/s41467-023-40552-y
Henke M.T., Kenny D.J., Cassilly C.D., Vlamakis H., Xavier R.J., Clardy J 2019) Ruminococcusgnavus, a member of the human gut microbiome associated with Crohn's disease, produces an inflammatory polysaccharide Proc. Nat. Acad. Sci 116:12672-12677 https://doi.org/10.1073/pnas.1904099116
Paone P., Cani P.D 2020) Mucus barrier, mucins and gut microbiota: the expected slimy partners? Gut 69:2232-2243 https://doi.org/10.1136/gutjnl-2020-322260
Comment 5: Additionally, the palmitic acid-containing diets fed to mice to induce a diabetes-like condition do not mimic spontaneous T2DM in macaques.
Thanks for your helpful suggestion. We agree that the palmitic acid (PA)-containing diet alone could not fully mimic spontaneous T2DM in macaques. In our study, the PA diet was employed in mouse experiments to investigate whether gut microbiota modulates serum PA levels and mediates T2DM progression. Our critical finding revealed that microbiota was essential for enhanced PA absorption, while simply increasing dietary levels of PA did not effectively enhance intestinal uptake. The fecal microbiota transplantation (FMT) combined with PA-diet approach successfully induced prediabetic states in mice, which can be further applied to the induction of T2DM in macaques. We have added future expectations in the part of the discussion, “Our study highlights the essential roles of gut microbiota in T2DM development, which may account for the inability of prior studies to induce T2DM in macaques through high-fat diet intervention alone (28, 29). Furthermore, applying this approach to induce T2DM in macaques will enable deeper investigation into gut-microbiota-driven mechanisms underlying disease pathogenesis.” (Lines 393-398).
Reviewer #1 (Recommendations for the authors):
General comments
Comment 1: The authors used macaques in this study. The author claims that macaques may be the best animal model to investigate the relationships among gut microbiota, lipid metabolites, and the host in type 2 diabetes (T2DM). However, there have already been some studies investigating these relationships in humans (for example, doi: 10.1016/j.cmet.2022.12.013, and doi: 10.1038/s41586-023-06466-x). The authors should cite and discuss these papers.
We thank the reviewer for this suggestion. We have cited the two papers in the part of discussion, “Previous studies have shown that insulin-resistant patients exhibit increased fecal monosaccharides associated with microbial carbohydrate metabolism (70). Furthermore, commensal species of Lachnospiraceae actively overproduce long-chain fatty acids during metabolic dysfunction through altered bacterial lipid metabolism. The microbe-derived fatty acids impair intestinal epithelial integrity to exacerbate metabolic dysregulation (71).” (Lines 426-432).
Specific comments
Major:
Comment 2: (1) First of all, sex and age of the T2DM and control groups are different (Suppl Table 1). Since the size of the captive population is 1,698, the authors should be able to select the factors including the sex and age of the control group to match those of the T2DM group and they should do so.
In this work, we obtained eight spontaneous T2DM macaques with FPG ≥ 7 mmol/L and eight heathy control macaques with FPG ≤ 6.1 mmol/L (three consecutive detections, each detection interval of one month) from a population of 1,698 captive macaques. To avoid confound factors affect the investigated macaques, all macaques were individually housed with standardized diets and environmental controls. While age and sex partially matched, controls originated from the same population to minimize confounding. The T2DM and control groups were matched for age period (5 adult and 3 elder) and had comparable mean ages (mean age of T2DM individuals = 12.88, mean age of control individuals = 11.25) (Table S1). In terms of gender matching, we compared blood metabolome data of 12 healthy adult female and 12 healthy adult male macaques from another study (Liu et al., 2023) and obtained only a very small number of differential metabolites that were not associated with tryptophan (Author response table 1). We acknowledge this limitation and will prioritize matched controls in future studies.
References:
Liu X., Liu X.Y., Wang X.Q., Shang K., Li J.W., Lan Y., Wang J., Li J., et al. 2023). Multi-Omics Analysis Reveals Changes in Tryptophan and Cholesterol Metabolism before and after Sexual Maturation in Captive Macaques BMC Genomics 24:308. https://doi.org/10.1186/s12864-023-09404-3
Comment 3: (2) Are the normal ranges known for the parameters of macaques shown in Table 1? If so, the authors should include those values in Table 1. If not, the authors should show the values of average and SD or SE of all 1,698 individuals as the reference.
We thank the reviewer for this suggestion. In this study, the normal ranges of fasting plasma glucose (FPG), fasting plasma insulin (FPI), homeostasismodel assessment- insulin resistance (HOMA-IR), and glycosylated hemoglobin A1cwe (HbA1c) were referenced against human standards. According to the American Diabetes Association (ADA) for glucose metabolism status and the diagnostic criteria for diabetes, individuals with FPG ≥ 7 mmol/L were diagnosed as T2DM subjects, and individuals with FPG ≤ 6.1 mmol/L were controls. More sensitive assays show a normal fasting plasma insulin level to be under 12 μU/mL (Matsuda and DeFronzo, 1999). HOMA-IR ≥ 2.67 indicated the possibility of insulin resistance, which is used in clinical diagnosis (Lorenzo et al., 2012). HbA1c percentages higher than 6.5% were used as an auxiliary diagnostic index for diabetic macaques (Cowie et al., 2010). The normal ranges of triglycerides (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL), and low-density lipoprotein cholesterol (LDL) were referenced against the blood lipid index of rhesus macaques (Yu et al., 2019). We have added the normal ranges of parameters to Table 1, “FPG: fasting plasma glucose (normal range: ≤ 6.1 mmol/L); FPI: fasting plasma insulin (normal range: ≤ 12 μU/mL); HOMA-IR: homeostasismodel assessment- insulin resistance (normal range: ≤ 2.67); BMI: body mass index; HbA1c: glycosylated hemoglobin A1c (normal range: < 6.5%); TG: triglycerides (normal range: 0.95±0.47 mmol/L); TC: total cholesterol (normal range: 3.06±0.98 mmol/L); HDL: high-density lipoprotein cholesterol (normal range: 1.62±0.46 mmol/L); LDL: low-density lipoprotein cholesterol (normal range: 2.47±0.98 mmol/L). (30, 31, 32, 33).”.
References:
Matsuda M., DeFronzo R.A 1999) Insulin sensitivity indices obtained from oral glucose tolerance testing: comparison with the euglycemic insulin clamp Diabetes care 22:1462-1470 https://doi.org/10.2337/diacare.22.9.1462
Lorenzo C., Hazuda H.P., Haffner S.M 2012) Insulin resistance and excess risk of diabetes in Mexican-Americans: the San Antonio Heart Study J. Clin. Endocr. Metab 97:793-799 https://doi.org/10.1210/jc.2011-2272
Cowie C.C., Rust K.F., Byrd-Holt D.D., Gregg E.W., Ford E.S., Geiss L.S., Bainbridge K.E., Fradkin J.E 2010) Prevalence of diabetes and high risk for diabetes using A1C criteria in the US population in 1988–2006 Diabetes care 33:562-568 https://doi.org/10.2337/dc09-1524
Yu W., Hao X., Yang F., Ma J., Zhao Y., Li Y., Wang J., Xu H., Chen L., Liu Q., et al. 2019) Hematological and biochemical parameters for Chinese rhesus macaque PLoS One 14:e0222338 https://doi.org/10.1371/journal.pone.0222338
Comment 4: (3) The authors measured the fasting plasma glucose (FPG) levels, but it is common to measure whole blood glucose since glucose is consumed during the processing of obtaining plasma which could compromise the results. Please explain why plasma glucose levels were measured.
The criteria for screening spontaneous T2DM macaques were guided by the American Diabetes Association (ADA) for glucose metabolism status and the diagnostic criteria for diabetes. Individuals with FPG ≥ 7 mmol/L were diagnosed as T2DM subjects, and individuals with FPG ≤ 6.1 mmol/L were controls. For the identified subjects, a total of three times of FPG tests were employed, with an interval of one month to reduce the possible error. These individuals were raised in a single cage, and blood samples were collected after an overnight fast at least 12 h. After the three test results meet the standards, venous blood was collected for FPG testing to ensure the reliability of the data to the greatest extent. We have added FPG values of three time to the Table S1.
Comment 5: (4) Since the BMI of the T2DM and control groups did not significantly differ (p>0.05, Table 1), the food intake of the two groups may not significantly differ as well. The authors should examine the food intake data. The food intake is also important in considering the relevance of feeding the PA diet in mice experiments. Were the intake of T2DM macaques including PA more than the control group?
All macaques in this study were individually housed under standardized environments with timed and measured feeding to minimize confounders. Given the non-significant BMI difference between T2DM and control groups, food intake was probably not significantly different. In this study, our findings highlight the essential roles of gut microbiota in T2DM development, and this is probable also the reason that previous studies have failed to induce T2DM in macaques because they have only used a high-fat diet (Ji et al., 2012; Tang, 2020). We agree that PA intake in T2DM macaques warrants focused investigation. Future investigations will incorporate detailed dietary monitoring including palmitic acid (PA) intake and nutrient composition to examine potential relationships between specific dietary components, metabolic parameters, and diabetes progression.
References
Ji F., Jin L., Zeng X., Zhang X., Zhang Y., Sun Y., Gao L., He H., Rao J., Liu X., et al. 2012) Comparison of gene expression between naturally occurring and diet-induced T2DM in cynomolgus monkeys Dongwuxue Yanjiu 33:79–84 https://doi.org/10.3724/SP.J.1141.2012 .01079
Tang MT. 2020) Study on the Role of Glucose and Lipid in the Establishment of Type 2 Diabetic Cynomolgus Monkey Model M.S. Thesis, Dept. Veterinary Med., South China Agricultural Univ. 2020
Comment 6: (5) It may be that the fecal microbiome of the T2DM macaques is involved in the pathogenesis of T2DM; however, it is more important how the gut microbiota compositions were obtained/established by those T2DM macaques. There was no description of when the fecal samples were collected during the course of T2DM. If it was after T2DM symptoms appeared, the authors should perform gut metagenome and also gut metabolome analyses to see the change in those parameters to try to understand how gut microbiome changes are induced leading to T2DM pathogenesis.
The spontaneous T2DM macaques untreated with glucose-lowering drugs or antibiotics, revealed microbiota dysbiosis driven purely by disease progression. After macaques met diagnostic thresholds across three FPG assessments (each detection interval of one month), we collected fresh fecal samples and stored them aseptically at -80 °C until analysis. The scarcity of spontaneous T2DM macaques precludes invasive sampling, restricting tissue collection to naturally deceased diabetic individuals, which prevented us to explicitly define the disease stage of the T2DM individuals. We recognize the scientific value of gut metagenomic and metabolomic analyses to track microbiome evolution during diabetes progression. This study explored the interaction of gut microbiota and metabolites in T2DM macaques, and future studies can continue to investigate its dynamic changes in the disease process of T2DM.
Comment 7: (6) Regarding the fatty acids, the authors only measured them in the plasma, but they also should measure in feces, since the authors focus on gut microbiota; in addition, a recent report showed fecal fatty acids, especially elaidic acid, contributed the pathogenesis of obesity and T2DM by acting on the gut epithelial cells (doi: 10.1016/j.cmet.2022.12.013). Besides, this study showed the link between a Lachnospiraceae species and fecal palmitic and elaidic acids, which the authors also focused on in this manuscript.
We thank the reviewer for this suggestion. This study employed untargeted metabolomics on macaque fecal samples to identify metabolites associated with spontaneously developing T2DM. To validate the metabolites identified through the untargeted metabolomic analysis, we conducted targeted medium- and long-chain fatty acid (MLCFA) metabolomics on macaque serum, and we further quantitatively examined the content of palmitic acid (PA) in mice feces, ileum, and serum. Although targeted MLCFA metabolomics was not performed on macaque fecal samples, we did perform untargeted metabolomics on macaque feces and confirmed the contribution of PA in mice that underwent fecal microbiota transplantation (FMT) from T2DM macaques. We have added future expectations in the part of the discussion, “Previous studies have shown that insulin-resistant individuals exhibit increased fecal monosaccharides associated with microbial carbohydrate metabolism (70). Furthermore, commensal species of Lachnospiraceae actively overproduce long-chain fatty acids during metabolic dysfunction through altered bacterial lipid metabolism. The microbe-derived fatty acids impair intestinal epithelial integrity to exacerbate metabolic dysregulation (71). Given that microbial metabolic activity causally modulates host metabolic homeostasis, the content change of PA was potentially associated with a dynamic equilibrium between host absorption and microbial metabolism. Further integrative studies on the fecal fatty acid metabolome, microbial PA metabolism, and functional pathways will be crucial for delineating causal links between dysbiosis and lipid metabolic dysfunction in T2DM.” (Lines 426-437).
Comment 8: (7) In FMT and PA diet experiments, SPF mice were used as the control group. However, the gut microbiota composition of the SPF mice is markedly different from that of macaques; the difference must be much bigger than the difference between T2DM and healthy control macaques; therefore, mice with FMT from healthy control macaques have to be used as the control group. As mentioned above (in point #4), is the feeding of mice with PA diet a relevant model reflecting the condition observed in macaques in this study?
Thanks for your helpful suggestion. We recognized the importance of a FMT control group and supplemented mouse experiments (using the C57BL/6J strain) with FMT from control macaques (HFT group). Another group of mice without FMT was set as control. Due to the lengthy experimental period, observations were concluded at 30 days post-FMT. We compared changes in the gut microbiota before and after antibiotic treatment in mice (-14D and 0D), and tracked body weight and fasting plasma glucose (FPG) levels from day -14 to day 30. At 30 days after FMT, fecal samples from all groups were collected for 16S rRNA sequencing. Additionally, samples of T2DM microbiota transplant (TP), and control transplant (HTP) were sequenced. Finally, we integrated the 16S sequencing data from the FTPA group (palmitic acid (PA) diet and FMT from T2DM macaques) and FT group (normal diet and FMT from T2DM macaques) at day 30 for combined analysis. The results showed that the antibiotic treatment used in this study effectively depleted the gut microbiota. Following FMT, gut microbial diversity stabilized within 30 days, with similar microbial community proportions between HFT and control groups. Core functional groups of the healthy microbiota (Bacteroidota and Bacillota) stably colonized mice despite host species divergence, confirming that T2DM phenotypes originate specifically from macaque microbiota. Importantly, increased abundance of Lachnospiraceae (including genera Ruminococcus (current name: Mediterraneibacter), Coprococcus, and Clostridium) and the key species Ruminococcus gnavus (current name: Mediterraneibacter gnavus) were also observed in FT group versus HFT group on day 30, validating our original findings. We have added findings in the results, “To eliminate interference from host species divergence in gut microbiota composition, we supplemented mouse experiments using FMT from control macaques (HFT group) (Figure S4A). By day 30, the HFT group exhibited significantly lower body weight than the untreated control group (p < 0.05) (Figure S4B). Throughout the experimental period, FPG levels in both HFT and control groups remained within the normal range (< 6 mmol/L) without significant differences, indicating that transplantation of control macaque microbiota did not induce glycemic alterations (Figure S4C).” (Lines 276-283), and “Integrating 16S rRNA sequencing data from the HFT, FT, and FTPA groups showed that the antibiotic treatment effectively depleted the gut microbiota, resulting in microbial diversity decreased sharply, with the dominant phyla shifting from Bacteroidota and Bacillota to Pseudomonadota (Figure S4D-G). The HFT group restored microbial diversity within 30 days, achieving community proportions comparable to untreated controls. Core functional phyla (Bacteroidota and Bacillota) stably colonized in HFT group (Figure S4D-I). Critically, FT and FTPA groups exhibited increased Lachnospiraceae (including genera Ruminococcus (current name: Mediterraneibacter), Coprococcus, and Clostridium) compared with the HFT group on day 30. In addition, LEfSe comparison identified significant R. gnavus (current name: M. gnavus) enrichment in the FT group (LDA > 3, p < 0.01) (Figure S4J-M).” (Lines 324-334, 825-837).
We agree that the PA-containing diet alone could not fully mimic spontaneous T2DM in macaques. In our study, the PA diet was employed in mouse experiments to investigate whether gut microbiota modulates serum PA levels and mediates T2DM progression. Our critical finding revealed that microbiota was essential for enhanced PA absorption, while simply increasing dietary levels of PA did not effectively enhance intestinal uptake. The FMT combined with PA-diet approach successfully induced prediabetic states in mice, which can be further applied to the induction of T2DM in macaques. We have added future expectations in the part of the discussion, “Our study highlights the essential roles of gut microbiota in T2DM development, which may account for the inability of prior studies to induce T2DM in macaques through high-fat diet intervention alone (28, 29). Furthermore, applying this approach to induce T2DM in macaques will enable deeper investigation into gut-microbiota-driven mechanisms underlying disease pathogenesis.” (Lines 393-398).
Comment 9: FPG was measured here in the mouse experiments, but there was no description of whether mice were under fasting conditions, and this should be clarified. If there are no fasting durations, this should be described in the Materials and Methods section.
As suggested, we have added description to the Materials and Methods section, “Throughout the experiment, body weight and feces were collected every month, FPG was detected every half month under fasting at least 12 h.” (Lines 619-620).
Comment 10: From the PA contents in feces, ileum, and serum in mice (Figures 5A-D), the authors concluded that the absorption of PA was significantly enhanced in the ileum leading to the increase of PA in serum. However, it could also be possible that consumption of PA by gut microbiota occurs at the same time and the authors should discuss the possibility.
We thank the reviewer for spotting this. We have added a discussion to the manuscript, “Previous studies have shown that insulin-resistant individuals exhibit increased fecal monosaccharides associated with microbial carbohydrate metabolism (70). Furthermore, commensal species of Lachnospiraceae actively overproduce long-chain fatty acids during metabolic dysfunction through altered bacterial lipid metabolism. The microbe-derived fatty acids impair intestinal epithelial integrity to exacerbate metabolic dysregulation (71). Given that microbial metabolic activity causally modulates host metabolic homeostasis, the content change of PA was potentially associated with a dynamic equilibrium between host absorption and microbial metabolism. Further integrative studies on the fecal fatty acid metabolome, microbial PA metabolism, and functional pathways will be crucial for delineating causal links between dysbiosis and lipid metabolic dysfunction in T2DM.” (Lines 426-437).
Comment 11: (8) Nomenclature and classification of bacteria has been revised by the List of Prokaryotic names with Standing in Nomenclature (LPSN) (https://lpsn.dsmz.de/) and recognized as Global Core Biodata Resource in 2023. For example, Ruminococcus gnavus is now Mediterraneibacter gnavus. Therefore, the name of microbes should be corrected accordingly; one proposal is to show the revised correct name with the previous name in parenthesis, such as "Mediterraneibacter gnavus (previously Ruminococcus gnavus)".
Thank you for pointing this out. We have corrected the name of microbe, “Ruminococcus (current name: Mediterraneibacter)”, “Ruminococcus gnavus (current name: Mediterraneibacter gnavus), and “R. gnavus (current name: M. gnavus)” (Lines 146, 313, 316-317, 336, 345, 367-368, 401, 404-405, 409, 448, 764-765)
Minor:
Comment 12:
(1) The sentence starting "A total of..." (lines 143-144) seems grammatically wrong; a word such as "represented" should be inserted after "differentially", or alternatively "differentially" should be "differential"?
(2) "medium-and" (line 220) needs a space between "medium-" and "and" to make it "medium- and".
(3) Abbreviations should be spelled out when they appear for the first time in the main text; for example, WBC, NEU, and LYM in line 237.
(4) Should FGP (line 437) be FPG?
(5) What is the definition of "prediabetes" in mice? Is this clearly defined elsewhere?
We sincerely thank the reviewer for careful reading. As suggested, we have improved the statements and revised it according to the requirements:
(1) Line 143: “A total of 21 microbes were identified as differential microbes”.
(2) Line 221: “targeted medium- and long-chain fatty acid”.
(3) Lines 238-239: “white blood cell (WBC)”, “neutrophil (NEU)”, and “lymphocyte (LYM)”.
(4) Line 472: “FPG, HbA1c and FPI were detected”.
(5) Prediabetes or impaired glucose regulation (IGR) is diagnosed when one exhibits blood glucose level higher than normal yet below the diabetic threshold, which is even more prevalent than T2DM in the population (American Diabetes, 2021). Given the higher glycemic diagnostic criteria in mice, we assessed diabetic manifestations integrating physiological and pathological evidence. Compared to control mice, those receiving FMT from T2DM macaques combined with a high-palmitic-acid diet (FTPA group) developed prediabetic characteristics by day 120. Physiological alterations included elevated fasting plasma glucose (FPG), increased fasting plasma insulin (FPI), impaired glucose tolerance, heightened insulin resistance, weight gain, and elevated serum total cholesterol (TC) and triglyceride (TG) levels. Particularly in pathological changes, hepatocytes focal necrosis with inflammatory cell infiltration was commonly observed in FTPA group, alongside decreased volume in pancreatic islets and inflammatory cell infiltration (lines 258-276).
References:
American Diabetes Association 2021) 2. Classification and diagnosis of diabetes: standards of medical care in diabetes—2021 Diabetes care 44:S15-S33 https://doi.org/10.2337/dc21-S002
Reviewer #2 (Public review):
This study analyzes the interaction among the gut microbiota, lipid metabolism, and the host in type 2 diabetes (T2DM) using rhesus macaques. The authors first identified 8 macaques with T2DM from 1698 individuals. Then, they observed in T2DM macaques: dysbiosis by 16S rRNA gene amplicon analysis and shotgun sequencing, imbalanced tryptophan metabolism and fatty acid beta oxidization in the feces by metabolome analysis, increased plasma concentration of palmitic acid by MS analysis, and sn inflammatory gene signature of blood cells by transcriptomic analysis. Finally, they transplanted feces of T2DM macaques into mice and fed them with palmitic acid and showed that those mice became diabetic through increased absorption of palmitic acid in the ileum.
Comment 1: This study clearly shows the interaction among gut microbiota, lipid metabolism, and the host in T2DM. The experiments were well designed and performed, and the data are convincing. One point I would suggest is that in the experiments of mice with FMT, control mice should be those colonized with feces of healthy macaques, but not with no FMT.
See response to Reviewer 1, Public review comment 4.



