Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorAndrew KingUniversity of Oxford, Oxford, United Kingdom
- Senior EditorAndrew KingUniversity of Oxford, Oxford, United Kingdom
Reviewer #1 (Public review):
Summary:
Mazer & Yovel 2025 dissect the inverse problem of how echolocators in groups manage to navigate their surroundings despite intense jamming using computational simulations.
The authors show that despite the 'noisy' sensory environments that echolocating groups present, agents can still access some amount of echo-related information and use it to navigate their local environment. It is known that echolocating bats have strong small and large-scale spatial memory that plays an important role for individuals. The results from this paper also point to the potential importance of an even lower-level, short-term role of memory in the form of echo 'integration' across multiple calls, despite the unpredictability of echo detection in groups. The paper generates a useful basis to think about the mechanisms in echolocating groups for experimental investigations too.
Strengths:
(1) The paper builds on biologically well-motivated and parametrised 2D acoustics and sensory simulation setup to investigate the various key parameters of interest
(2) The 'null-model' of echolocators not being able to tell apart objects & conspecifics while echolocating still shows agents successfully emerge from groups - even though the probability of emergence drops severely in comparison to cognitively more 'capable' agents. This is nonetheless an important result showing the direction-of-arrival of a sound itself is the 'minimum' set of ingredients needed for echolocators navigating their environment.
(3) The results generate an important basis in unraveling how agents may navigate in sensorially noisy environments with a lot of irrelevant and very few relevant cues.
(4) The 2D simulation framework is simple and computationally tractable enough to perform multiple runs to investigate many variables - while also remaining true to the aim of the investigation.
Weaknesses:
There are a few places in the paper that can be misunderstood or don't provide complete details. Here is a selection:
(1) Line 61: '... studies have focused on movement algorithms while overlooking the sensory challenges involved' : This statement does not match the recent state of the literature. While the previous models may have had the assumption that all neighbours can be detected, there are models that specifically study the role of limited interaction arising from a potential inability to track all neighbours due to occlusion, and the effect of responding to only one/few neighbours at a time e.g. Bode et al. 2011 R. Soc. Interface, Rosenthal et al. 2015 PNAS, Jhawar et al. 2020 Nature Physics.
(2) The word 'interference' is used loosely places (Line 89: '...took all interference signals...', Line 319: 'spatial interference') - this is confusing as it is not clear whether the authors refer to interference in the physics/acoustics sense, or broadly speaking as a synonym for reflections and/or jamming.
(3) The paper discusses original results without reference to how they were obtained or what was done. The lack of detail here must be considered while interpreting the Discussion e.g. Line 302 ('our model suggests...increasing the call-rate..' - no clear mention of how/where call-rate was varied) & Line 323 '..no benefit beyond a certain level..' - also no clear mention of how/where call-level was manipulated in the simulations.
Reviewer #2 (Public review):
This manuscript describes a detailed model of bats flying together through a fixed geometry. The model considers elements that are faithful to both bat biosonar production and reception and the acoustics governing how sound moves in the air and interacts with obstacles. The model also incorporates behavioral patterns observed in bats, like one-dimensional feature following and temporal integration of cognitive maps. From a simulation study of the model and comparison of the results with the literature, the authors gain insight into how often bats may experience destructive interference of their acoustic signals and those of their peers, and how much such interference may actually negatively affect the groups' ability to navigate effectively. The authors use generalized linear models to test the significance of the effects they observe.
In terms of its strengths, the work relies on a thoughtful and detailed model that faithfully incorporates salient features, such as acoustic elements like the filter for a biological receiver and temporal aggregation as a kind of memory in the system. At the same time, the authors' abstract features are complicating without being expected to give additional insights, as can be seen in the choice of a two-dimensional rather than three-dimensional system. I thought that the level of abstraction in the model was perfect, enough to demonstrate their results without needless details. The results are compelling and interesting, and the authors do a great job discussing them in the context of the biological literature.
The most notable weakness I found in this work was that some aspects of the model were not entirely clear to me. For example, the directionality of the bat's sonar call in relation to its velocity. Are these the same? If so, what is the difference between phi_target and phi_tx in the model equations? What is a bat's response to colliding with a conspecific (rather than a wall)? From the statistical side, it was not clear if replicate simulations were performed. If they were, which I believe is the right way due to stochasticity in the model, how many replicates were used, and are the standard errors referred to throughout the paper between individuals in the same simulation or between independent simulations, or both?
Overall, I found these weaknesses to be superficial and easily remedied by the authors. The authors presented well-reasoned arguments that were supported by their results, and which were used to demonstrate how call interference impacts the collective's roost exit as measured by several variables. As the authors highlight, I think this work is valuable to individuals interested in bat biology and behavior, as well as to applications in engineered multi-agent systems like robotic swarms.
Reviewer #3 (Public review):
Summary:
The authors describe a model to mimic bat echolocation behavior and flight under high-density conditions and conclude that the problem of acoustic jamming is less severe than previously thought, conflating the success of their simulations (as described in the manuscript) with hard evidence for what real bats are actually doing. The authors base their model on two species of bats that fly at "high densities" (defined by the authors as colony sizes from tens to tens of thousands of individuals and densities of up to 33.3 bats/m2), Pipistrellus kuhli and Rhinopoma microphyllum. This work fits into the broader discussion of bat sensorimotor strategies during collective flight, and simulations are important to try to understand bat behavior, especially given a lack of empirical data. However, I have major concerns about the assumptions of the parameters used for the simulation, which significantly impact both the results of the simulation and the conclusions that can be made from the data. These details are elaborated upon below, along with key recommendations the authors should consider to guide the refinement of the model.
Strengths:
This paper carries out a simulation of bat behavior in dense swarms as a way to explain how jamming does not pose a problem in dense groups. Simulations are important when we lack empirical data. The simulation aims to model two different species with different echolocation signals, which is very important when trying to model echolocation behavior. The analyses are fairly systematic in testing all ranges of parameters used and discussing the differential results.
Weaknesses:
The justification for how the different foraging phase call types were chosen for different object detection distances in the simulation is unclear. Do these distances match those recorded from empirical studies, and if so, are they identical for both species used in the simulation? What reasoning do the authors have for a bat using the same call characteristics to detect a cave wall as they would for detecting a small insect? Additionally, details on the signal creation are also absent, but based on the sample spectrogram in Figure 2A, it appears that the authors used a synthetic linear FM chirp characterized by the call parameters. This simplification of the echolocation signals for these species is not representative of the true emitted signals, which are nonlinear FM for not only the species used within this simulation--PK (Schnitzler et al., 1987; Kalko and Schnitzler 1993 and RM (Schmidt and Joermann 1986)-but also for many other bat species that form large aggregations and undergo dense emergence. Furthermore, echolocation calls of bats emitted during dense emergence flights (see Gillam et al 2010) can be very much different from those emitted during foraging calls, so limiting the simulation to foraging calls may not be valid. Why did the authors not use actual waveforms of calls produced by these species during dense emergence to use biologically relevant signals in their simulation?
The two species modeled have different calls. In particular, the bandwidth varies by a factor of 10, meaning the species' sonars will have different spatial resolutions. Range resolution is about 10x better for PK compared to RM, but the authors appear to use the same thresholds for "correct detection" for both, which doesn't seem appropriate. Also, the authors did not mention incorporating/correcting for/exploiting Doppler, which leads me to assume they did not model it.
The success of the simulation may very well be due to variation in the calls of the bats, which ironically enough demonstrates the importance of a jamming avoidance response in dense flight. This explains why the performance of the simulation falls when bats are not able to distinguish their own echoes from other signals. For example, in Figure C2, there are calls that are labeled as conspecific calls and have markedly shorter durations and wider bandwidths than others. These three phases for call types used by the authors may be responsible for some (or most) of the performance of the model since the correlation between different call types is unlikely to exceed the detection threshold. But it turns out this variation in and of itself is what a jamming avoidance response may consist of. So, in essence, the authors are incorporating a jamming avoidance response into their simulation.
The authors claim that integration over multiple pings (though I was not able to determine the specifics of this integration algorithm) reduces the masking problem. Indeed, it should: if you have two chances at detection, you've effectively increased your SNR by 3dB.
They also claim - although it is almost an afterthought - that integration dramatically reduces the degradation caused by false echoes. This also makes sense: from one ping to the next, the bat's own echo delays will correlate extremely well with the bat's flight path. Echo delays due to conspecifics will jump around kind of randomly. However, the main concern is regarding the time interval and number of pings of the integration, especially in the context of the bat's flight speed. The authors say that a 1s integration interval (5-10 pings) dramatically reduces jamming probability and echo confusion. This number of pings isn't very high, and it occurs over a time interval during which the bat has moved 5-10m. This distance is large compared to the 0.4m distance-to-obstacle that triggers an evasive maneuver from the bat, so integration should produce a latency in navigation that significantly hinders the ability to avoid obstacles. Can the authors provide statistics that describe this latency, and discussion about why it doesn't seem to be a problem?
The authors are using a 2D simulation, but this very much simplifies the challenge of a 3D navigation task, and there is an explanation as to why this is appropriate. Bat densities and bat behavior are discussed per unit area when realistically it should be per unit volume. In fact, the authors reference studies to justify the densities used in the simulation, but these studies were done in a 3D world. If the authors have justification for why it is realistic to model a 3D world in a 2D simulation, I encourage them to provide references justifying this approach.
The focus on "masking" (which appears to be just in-band noise), especially relative to the problem of misassigned echoes, is concerning. If the bat calls are all the same waveform (downsweep linear FM of some duration, I assume - it's not clear from the text), false echoes would be a major problem. Masking, as the authors define it, just reduces SNR. This reduction is something like sqrt(N), where N is the number of conspecifics whose echoes are audible to the bat, so this allows the detection threshold to be set lower, increasing the probability that a bat's echo will exceed a detection threshold. False echoes present a very different problem. They do not reduce SNR per se, but rather they cause spurious threshold excursions (N of them!) that the bat cannot help but interpret as obstacle detection. I would argue that in dense groups the mis-assignment problem is much more important than the SNR problem.
The criteria set for flight behavior (lines 393-406) are not justified with any empirical evidence of the flight behavior of wild bats in collective flight. How did the authors determine the avoidance distances? Also, what is the justification for the time limit of 15 seconds to emerge from the opening? Instead of an exit probability, why not instead use a time criterion, similar to "How long does it take X% of bats to exit?" What is the empirical justification for the 1-10 calls used for integration? The "average exit time for 40 bats" is also confusing and not well explained. Was this determined empirically? From the simulation? If the latter, what are the conditions? Does it include masking, no masking, or which species?
Author response:
Public Reviews:
Reviewer #1 (Public review):
We thank the reviewer for his valuable input and careful assessment, which have significantly improved the clarity and rigor of our manuscript.
Summary:
Mazer & Yovel 2025 dissect the inverse problem of how echolocators in groups manage to navigate their surroundings despite intense jamming using computational simulations.
The authors show that despite the 'noisy' sensory environments that echolocating groups present, agents can still access some amount of echo-related information and use it to navigate their local environment. It is known that echolocating bats have strong small and large-scale spatial memory that plays an important role for individuals. The results from this paper also point to the potential importance of an even lower-level, short-term role of memory in the form of echo 'integration' across multiple calls, despite the unpredictability of echo detection in groups. The paper generates a useful basis to think about the mechanisms in echolocating groups for experimental investigations too.
Strengths:
(1) The paper builds on biologically well-motivated and parametrised 2D acoustics and sensory simulation setup to investigate the various key parameters of interest
(2) The 'null-model' of echolocators not being able to tell apart objects & conspecifics while echolocating still shows agents successfully emerge from groups - even though the probability of emergence drops severely in comparison to cognitively more 'capable' agents. This is nonetheless an important result showing the direction-of-arrival of a sound itself is the 'minimum' set of ingredients needed for echolocators navigating their environment.
(3) The results generate an important basis in unraveling how agents may navigate in sensorially noisy environments with a lot of irrelevant and very few relevant cues.
(4) The 2D simulation framework is simple and computationally tractable enough to perform multiple runs to investigate many variables - while also remaining true to the aim of the investigation.
Weaknesses:
There are a few places in the paper that can be misunderstood or don't provide complete details. Here is a selection:
(1) Line 61: '... studies have focused on movement algorithms while overlooking the sensory challenges involved' : This statement does not match the recent state of the literature. While the previous models may have had the assumption that all neighbours can be detected, there are models that specifically study the role of limited interaction arising from a potential inability to track all neighbours due to occlusion, and the effect of responding to only one/few neighbours at a time e.g. Bode et al. 2011 R. Soc. Interface, Rosenthal et al. 2015 PNAS, Jhawar et al. 2020 Nature Physics.
We appreciate the reviewer's comment and the relevant references. We have revised the manuscript accordingly to clarify the distinction between studies that incorporate limited interactions and those that explicitly analyze sensory constraints and interference. We have refined our statement to acknowledge these contributions while maintaining our focus on sensory challenges beyond limited neighbor detection, such as signal degradation, occlusion effects, and multimodal sensory integration (see lines 61-64):
While collective movement has been extensively studied in various species, including insect swarming, fish schooling, and bird murmuration (Pitcher, Partridge and Wardle, 1976; Partridge, 1982; Strandburg-Peshkin et al., 2013; Pearce et al., 2014; Rosenthal, Twomey, Hartnett, Wu, Couzin, et al., 2015; Bastien and Romanczuk, 2020; Davidson et al., 2021; Aidan, Bleichman and Ayali, 2024), as well as in swarm robotics agents performing tasks such as coordinated navigation and maze-solving (Faria Dias et al., 2021; Youssefi and Rouhani, 2021; Cheraghi, Shahzad and Graffi, 2022), most studies have focused on movement algorithms , often assuming full detection of neighbors (Parrish and Edelstein-Keshet, 1999; Couzin et al., 2002, 2005; Sumpter et al., 2008; Nagy et al., 2010; Bialek et al., 2012; Gautrais et al., 2012; Attanasi et al., 2014). Some models have incorporated limited interaction rules where individuals respond to one or a few neighbors due to sensory constraints (Bode, Franks and Wood, 2011; Jhawar et al., 2020). However, fewer studies explicitly examine how sensory interference, occlusion, and noise shape decision-making in collective systems (Rosenthal et al., 2015).
(2) The word 'interference' is used loosely places (Line 89: '...took all interference signals...', Line 319: 'spatial interference') - this is confusing as it is not clear whether the authors refer to interference in the physics/acoustics sense, or broadly speaking as a synonym for reflections and/or jamming.
To improve clarity, we have revised the manuscript to distinguish between different types of interference:
· Acoustic interference (jamming): Overlapping calls that completely obscure echo detection, preventing bats from perceiving necessary environmental cues.
· Acoustic interference (masking): Partial reduction in signal clarity due to competing calls.
· Spatial interference: Physical obstruction by conspecifics affecting movement and navigation.
We have updated the manuscript to use these terms consistently and explicitly define them in relevant sections (see lines 87-94 and 329-330). This distinction ensures that the reader can differentiate between interference as an acoustic phenomenon and its broader implications in navigation.
(3) The paper discusses original results without reference to how they were obtained or what was done. The lack of detail here must be considered while interpreting the Discussion e.g. Line 302 ('our model suggests...increasing the call-rate..' - no clear mention of how/where call-rate was varied) & Line 323 '..no benefit beyond a certain level..' - also no clear mention of how/where call-level was manipulated in the simulations.
All tested parameters, including call rate dynamics and call intensity variations, are detailed in the Methods section and Tables 1 and 2. Specifically:
· Call Rate Variation: The Inter-Pulse Interval (IPI) was modeled based on documented echolocation behavior, decreasing from 100 msec during the search phase to 35 msec (~28 calls per second) at the end of the approach phase, and to 5 msec (200 calls per second) during the final buzz (see Table 2). This natural variation in call rate was not manually manipulated in the model but emerged from the simulated bat behavior.
· Call Intensity Variation: The tested call intensity levels (100, 110, 120, 130 dB SPL) are presented in Table 1 under the “Call Level” parameter. The effect of increasing call intensity was analyzed in relation to exit probability, jamming probability, and collision rate. This is now explicitly referenced in the Discussion.
We have revised the manuscript to explicitly reference these aspects in the Results and Discussion sections.
Reviewer #2 (Public review):
We are grateful for the reviewer’s insightful feedback, which has helped us clarify key aspects of our research and strengthen our conclusions.
This manuscript describes a detailed model of bats flying together through a fixed geometry. The model considers elements that are faithful to both bat biosonar production and reception and the acoustics governing how sound moves in the air and interacts with obstacles. The model also incorporates behavioral patterns observed in bats, like one-dimensional feature following and temporal integration of cognitive maps. From a simulation study of the model and comparison of the results with the literature, the authors gain insight into how often bats may experience destructive interference of their acoustic signals and those of their peers, and how much such interference may actually negatively affect the groups' ability to navigate effectively. The authors use generalized linear models to test the significance of the effects they observe.
In terms of its strengths, the work relies on a thoughtful and detailed model that faithfully incorporates salient features, such as acoustic elements like the filter for a biological receiver and temporal aggregation as a kind of memory in the system. At the same time, the authors' abstract features are complicating without being expected to give additional insights, as can be seen in the choice of a two-dimensional rather than three-dimensional system. I thought that the level of abstraction in the model was perfect, enough to demonstrate their results without needless details. The results are compelling and interesting, and the authors do a great job discussing them in the context of the biological literature.
The most notable weakness I found in this work was that some aspects of the model were not entirely clear to me.
For example, the directionality of the bat's sonar call in relation to its velocity. Are these the same?
For simplicity, in our model, the head is aligned with the body, therefore the direction of the echolocation beam is the same as the direction of the flight.
Moreover, call directionality (directivity) is not directly influenced by velocity. Instead, directionality is estimated using the piston model, as described in the Methods section. The directionality is based on the emission frequency and is thus primarily linked to the behavioral phases of the bat, with frequency shifts occurring as the bat transitions from search to approach to buzz phases. During the approach phase, the bat emits calls with higher frequencies, resulting in increased directionality. This is supported by the literature (Jakobsen and Surlykke, 2010; Jakobsen, Brinkløv and Surlykke, 2013). This phase is also associated with a natural reduction in flight speed, which is a well-documented behavioral adaptation in echolocating bats (Jakobsen et al., 2024).
To clarify this in the manuscript, we have updated the text to explicitly state that directionality follows phase-dependent frequency changes rather than being a direct function of velocity, see lines 460-465.
If so, what is the difference between phi_target and phi_tx in the model equations?
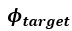 represents the angle between the bat and the reflected object (target).
represents the angle between the bat and the reflected object (target).
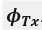 the angle [rad], between the masking bat and target (from the transmitter’s perspective)
the angle [rad], between the masking bat and target (from the transmitter’s perspective)
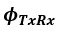 refers to the angle between the transmitting conspecific and the receiving focal bat, from the transmitter’s point of view.
refers to the angle between the transmitting conspecific and the receiving focal bat, from the transmitter’s point of view.
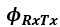 represents the angle between the receiving bat and the transmitting bat, from the receiver’s point of view.
represents the angle between the receiving bat and the transmitting bat, from the receiver’s point of view.
These definitions have been explicitly stated in the revised manuscript to prevent any ambiguity (lines 467-468). Additionally, a Supplementary figure demonstrating the geometrical relations has been added to the manuscript.
Author response image 1.
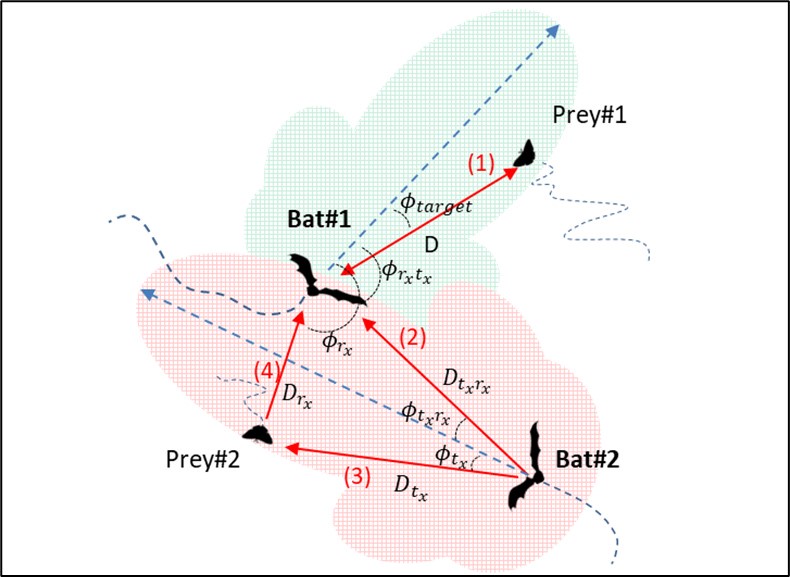
What is a bat's response to colliding with a conspecific (rather than a wall)?
In nature, minor collisions between bats are common and typically do not result in significant disruptions to flight (Boerma et al., 2019; Roy et al., 2019; Goldstein et al., 2024).Given this, our model does not explicitly simulate the physical impact of a collision event. Instead, during the collision event the bat keeps decreasing its velocity and changing its flight direction until the distance between bats is above the threshold (0.4 m). We assume that the primary cost of such interactions arises from the effort required to avoid collisions, rather than from the collision itself. This assumption aligns with observations of bat behavior in dense flight environments, where individuals prioritize collision avoidance rather than modeling post-collision dynamics.
From the statistical side, it was not clear if replicate simulations were performed. If they were, which I believe is the right way due to stochasticity in the model, how many replicates were used, and are the standard errors referred to throughout the paper between individuals in the same simulation or between independent simulations, or both?
The number of repetitions for each scenario is detailed in Table 1, but we included it in a more prominent location in the text for clarity. Specifically, we now state (Lines 274-275):
"The number of repetitions for each scenario was as follows: 1 bat: 240; 2 bats: 120; 5 bats: 48; 10 bats: 24; 20 bats: 12; 40 bats: 12; 100 bats: 6."
Regarding the reported standard errors, they are calculated across all individuals within each scenario, without distinguishing between different simulation trials.
We clarified in the revised text (Lines 534-535 in Statistical Analysis)
Overall, I found these weaknesses to be superficial and easily remedied by the authors. The authors presented well-reasoned arguments that were supported by their results, and which were used to demonstrate how call interference impacts the collective's roost exit as measured by several variables. As the authors highlight, I think this work is valuable to individuals interested in bat biology and behavior, as well as to applications in engineered multi-agent systems like robotic swarms.
Reviewer #3 (Public review):
We sincerely appreciate the reviewer’s thoughtful comments and the time invested in evaluating our work, which have greatly contributed to refining our study.
We would like to note that in general, our model often simplifies some of the bats’ abilities, under the assumption that if the simulated bats manage to perform this difficult task with simpler mechanisms, real better adapted bats will probably perform even better. This thought strategy will be repeated in several of the answers below.
Summary:
The authors describe a model to mimic bat echolocation behavior and flight under high-density conditions and conclude that the problem of acoustic jamming is less severe than previously thought, conflating the success of their simulations (as described in the manuscript) with hard evidence for what real bats are actually doing. The authors base their model on two species of bats that fly at "high densities" (defined by the authors as colony sizes from tens to tens of thousands of individuals and densities of up to 33.3 bats/m2), Pipistrellus kuhli and Rhinopoma microphyllum. This work fits into the broader discussion of bat sensorimotor strategies during collective flight, and simulations are important to try to understand bat behavior, especially given a lack of empirical data. However, I have major concerns about the assumptions of the parameters used for the simulation, which significantly impact both the results of the simulation and the conclusions that can be made from the data. These details are elaborated upon below, along with key recommendations the authors should consider to guide the refinement of the model.
Strengths:
This paper carries out a simulation of bat behavior in dense swarms as a way to explain how jamming does not pose a problem in dense groups. Simulations are important when we lack empirical data. The simulation aims to model two different species with different echolocation signals, which is very important when trying to model echolocation behavior. The analyses are fairly systematic in testing all ranges of parameters used and discussing the differential results.
Weaknesses:
The justification for how the different foraging phase call types were chosen for different object detection distances in the simulation is unclear. Do these distances match those recorded from empirical studies, and if so, are they identical for both species used in the simulation?
The distances at which bats transition between echolocation phases are identical for both species in our model (see Table 2). These distances are based on well-documented empirical studies of bat hunting and obstacle avoidance behavior (Griffin, Webster and Michael, 1958; Simmons and Kick, 1983; Schnitzler et al., 1987; Kalko, 1995; Hiryu et al., 2008; Vanderelst and Peremans, 2018). These references provide extensive evidence that insectivorous bats systematically adjust their echolocation calls in response to object proximity, following the characteristic phases of search, approach, and buzz.
To improve clarity, we have updated the text to explicitly state that the phase transition distances are empirically grounded and apply equally to both modeled species (lines 430-447).
What reasoning do the authors have for a bat using the same call characteristics to detect a cave wall as they would for detecting a small insect?
In echolocating bats, call parameters are primarily shaped by the target distance and echo strength. Accordingly, there is little difference in call structure between prey capture and obstacles-related maneuvers, aside from intensity adjustments based on target strength (Hagino et al., 2007; Hiryu et al., 2008; Surlykke, Ghose and Moss, 2009; Kothari et al., 2014). In our study, due to the dense cave environment, the bats are found to operate in the approach phase nearly all the time, which is consistent with natural cave emergence, where they are navigating through a cluttered environment rather than engaging in open-space search. For one of the species (Rhinopoma M.), we also have empirical recordings of individuals flying under similar conditions (Goldstein et al., 2024). Our model was designed to remain as simple as possible while relying on conservative assumptions that may underestimate bat performance. If, in reality, bats fine-tune their echolocation calls even earlier or more precisely during navigation than assumed, our model would still conservatively reflect their actual capabilities.
We actually used logarithmically frequency modulated (FM) chirps, generated using the MATLAB built-in function chirp(t, f0, t1, f1, 'logarithmic'). This method aligns with the nonlinear FM characteristics of Pipistrellus kuhlii (PK) and Rhinopoma microphyllum (RM) and provides a realistic approximation of their echolocation signals. We acknowledge that this was not sufficiently emphasized in the original text, and we have now explicitly highlighted this in the revised version to ensure clarity (sell Lines 447-449 in Methods).
The two species modeled have different calls. In particular, the bandwidth varies by a factor of 10, meaning the species' sonars will have different spatial resolutions. Range resolution is about 10x better for PK compared to RM, but the authors appear to use the same thresholds for "correct detection" for both, which doesn't seem appropriate.
The detection process in our model is based on Saillant’s method using a filter bank, as detailed in the paper (Saillant et al., 1993; Neretti et al., 2003; Sanderson et al., 2003). This approach inherently incorporates the advantages of a wider bandwidth, meaning that the differences in range resolution between the species are already accounted for within the signal-processing framework. Thus, there is no need to explicitly adjust the model parameters for bandwidth variations, as these effects emerge from the applied method.
Also, the authors did not mention incorporating/correcting for/exploiting Doppler, which leads me to assume they did not model it.
The reviewer is correct. To maintain model simplicity, we did not incorporate the Doppler effect or its impact on echolocation. The exclusion of Doppler effects was based on the assumption that while Doppler shifts can influence frequency perception, their impact on jamming and overall navigation performance is minor within the modelled context.
The maximal Doppler shifts expected for the bats in this scenario are of ~ 1kHz. These shifts would be applied variably across signals due to the semi-random relative velocities between bats, leading to a mixed effect on frequency changes. This variability would likely result in an overall reduction in jamming rather than exacerbating it, aligning with our previous statement that our model may overestimate the severity of acoustic interference. Such Doppler shifts would result in errors of 2-4 cm in localization (i.e., 200-400 micro-seconds) (Boonman, Parsons and Jones, 2003).
We have now explicitly highlighted this in the revised version (see Lines 468-470).
The success of the simulation may very well be due to variation in the calls of the bats, which ironically enough demonstrates the importance of a jamming avoidance response in dense flight. This explains why the performance of the simulation falls when bats are not able to distinguish their own echoes from other signals. For example, in Figure C2, there are calls that are labeled as conspecific calls and have markedly shorter durations and wider bandwidths than others. These three phases for call types used by the authors may be responsible for some (or most) of the performance of the model since the correlation between different call types is unlikely to exceed the detection threshold. But it turns out this variation in and of itself is what a jamming avoidance response may consist of. So, in essence, the authors are incorporating a jamming avoidance response into their simulation.
We fully agree that the natural variations in call design between the phases contribute significantly to interference reduction (see our discussion in a previous paper in Mazar & Yovel, 2020). However, we emphasize that this cannot be classified as a Jamming Avoidance Response (JAR). In our model, bats respond only to the physical presence of objects and not to the acoustic environment or interference itself. There is no active or adaptive adjustment of call design to minimize jamming beyond the natural phase-dependent variations in call structure. Therefore, while variation in call types does inherently reduce interference, this effect emerges passively from the modeled behavior rather than as an intentional strategy to avoid jamming.
The authors claim that integration over multiple pings (though I was not able to determine the specifics of this integration algorithm) reduces the masking problem. Indeed, it should: if you have two chances at detection, you've effectively increased your SNR by 3dB.
The reviewer is correct. Indeed, integration over multiple calls improves signal-to-noise ratio (SNR), effectively increasing it by approximately 3 dB per doubling of observations. The specifics of the integration algorithm are detailed in the Methods section, where we describe how sensory information is aggregated across multiple time steps to enhance detection reliability.
They also claim - although it is almost an afterthought - that integration dramatically reduces the degradation caused by false echoes. This also makes sense: from one ping to the next, the bat's own echo delays will correlate extremely well with the bat's flight path. Echo delays due to conspecifics will jump around kind of randomly. However, the main concern is regarding the time interval and number of pings of the integration, especially in the context of the bat's flight speed. The authors say that a 1s integration interval (5-10 pings) dramatically reduces jamming probability and echo confusion. This number of pings isn't very high, and it occurs over a time interval during which the bat has moved 5-10m. This distance is large compared to the 0.4m distance-to-obstacle that triggers an evasive maneuver from the bat, so integration should produce a latency in navigation that significantly hinders the ability to avoid obstacles. Can the authors provide statistics that describe this latency, and discussion about why it doesn't seem to be a problem?
As described in the Methods section, the bat’s collision avoidance response does not solely rely on the integration process. Instead, the model incorporates real-time echoes from the last calls, which are used independently of the integration process for immediate obstacle avoidance maneuvers. This ensures that bats can react to nearby obstacles without being hindered by the integration latency. The slower integration on the other hand is used for clustering, outlier removal and estimation wall directions to support the pathfinding process, as illustrated in Supplementary Figure 1.
Additionally, our model assumes that bats store the physical positions of echoes in an allocentric coordinate system (x-y). The integration occurs after transforming these detections from a local relative reference frame to a global spatial representation. This allows for stable environmental mapping while maintaining responsiveness to immediate changes in the bat’s surroundings.
See lines 518-523 in the revied version.
The authors are using a 2D simulation, but this very much simplifies the challenge of a 3D navigation task, and there is an explanation as to why this is appropriate. Bat densities and bat behavior are discussed per unit area when realistically it should be per unit volume. In fact, the authors reference studies to justify the densities used in the simulation, but these studies were done in a 3D world. If the authors have justification for why it is realistic to model a 3D world in a 2D simulation, I encourage them to provide references justifying this approach.
We acknowledge that this is a simplification; however, from an echolocation perspective, a 2D framework represents a worst-case scenario in terms of bat densities and maneuverability:
· Higher Effective Density: A 2D model forces all bats into a single plane rather than distributing them through a 3D volume, increasing the likelihood of overlap in calls and echoes and making jamming more severe. As described in the text: the average distance to the nearest bat in our simulation is 0.27m (with 100 bats), whereas reported distances in very dense colonies are 0.5m, as observed in Myotis grisescens and Tadarida brasiliensis (Fujioka et al., 2021; Sabol and Hudson, 1995; Betke et al., 2008; Gillam et al, 2010)
· Reduced Maneuverability: In 3D space, bats can use vertical movement to avoid obstacles and conspecifics. A 2D constraint eliminates this degree of freedom, increasing collision risk and limiting escape options.
Thus, our 2D model provides a conservative difficult test case, ensuring that our findings are valid under conditions where jamming and collision risks are maximized. Additionally, the 2D framework is computationally efficient, allowing us to perform multiple simulation runs to explore a broad parameter space and systematically test the impact of different variables.
To address the reviewer’s concern, we have clarified this justification in the revised text and will provide supporting references where applicable: (see Methods lines 407-412)
The focus on "masking" (which appears to be just in-band noise), especially relative to the problem of misassigned echoes, is concerning. If the bat calls are all the same waveform (downsweep linear FM of some duration, I assume - it's not clear from the text), false echoes would be a major problem. Masking, as the authors define it, just reduces SNR. This reduction is something like sqrt(N), where N is the number of conspecifics whose echoes are audible to the bat, so this allows the detection threshold to be set lower, increasing the probability that a bat's echo will exceed a detection threshold. False echoes present a very different problem. They do not reduce SNR per se, but rather they cause spurious threshold excursions (N of them!) that the bat cannot help but interpret as obstacle detection. I would argue that in dense groups the mis-assignment problem is much more important than the SNR problem.
There is substantial literature supporting the assumption that bats can recognize their own echoes and distinguish them from conspecific signals (Schnitzler and Bioscience, 2001; Kazial, Burnett and Masters, 2001; Burnett and Masters, 2002; Kazial, Kenny and Burnett, 2008; Chili, Xian and Moss, 2009; Yovel et al., 2009; Beetz and Hechavarría, 2022). However, we acknowledge that false echoes may present a major challenge in dense groups. To address this, we explicitly tested the impact of the self-echo identification assumption in our study see Results Figure 4: The impact of confusion on performance, and lines 345-355 in the Discussion.
Furthermore, we examined a full confusion scenario, where all reflected echoes from conspecifics were misinterpreted as obstacle reflections (i.e., 100% confusion). Our results show that this significantly degrades navigation performance, supporting the argument that echo misassignment is a critical issue. However, we also explored a simple mitigation strategy based on temporal integration with outlier rejection, which provided some improvement in performance. This suggests that real bats may possess additional mechanisms to enhance self-echo identification and reduce false detections. See lines XX in the manuscript for further discussion.
The criteria set for flight behavior (lines 393-406) are not justified with any empirical evidence of the flight behavior of wild bats in collective flight. How did the authors determine the avoidance distances? Also, what is the justification for the time limit of 15 seconds to emerge from the opening? Instead of an exit probability, why not instead use a time criterion, similar to "How long does it take X% of bats to exit?"
While we acknowledge that wild bats may employ more complex behaviors for collision avoidance, we chose to implement a simplified decision-making rule in our model to maintain computational tractability.
The avoidance distances (1.5 m from walls and 0.4 m from other bats) were selected as internal parameters to ensure coherent flight trajectories while maintaining a reasonable collision rate. These distances provide a balance between maneuverability and stability, preventing erratic flight patterns while still enabling effective obstacle avoidance. In the revised paper, we have added supplementary figures illustrating the effect of model parameters on performance, specifically focusing on the avoidance distance.
The 15-second exit limit was determined as described in the text (Lines 403-404): “A 15-second window was chosen because it is approximately twice the average exit time for 40 bats and allows for a second corrective maneuver if needed.” In other words, it allowed each bat to circle the ‘cave’ twice to exit even in the most crowded environment. This threshold was set to keep simulation time reasonable while allowing sufficient time for most bats to exit successfully.
We acknowledge that the alternative approach suggested by the reviewer—measuring the time taken for a certain percentage of bats to exit—is also valid. However, in our model, some outlier bats fail to exit and continue flying for many minutes, Such simulations would lead to excessive simulation times making it difficult to generate repetitions and not teaching us much – they usually resulted from the bat slightly missing the opening (see video S1. Our chosen approach ensures practical runtime constraints while still capturing relevant performance metrics.
What is the empirical justification for the 1-10 calls used for integration?
The "average exit time for 40 bats" is also confusing and not well explained. Was this determined empirically? From the simulation? If the latter, what are the conditions? Does it include masking, no masking, or which species?
Previous studies have demonstrated that bats integrate acoustic information received sequentially over several echolocation calls (2-15), effectively constructing an auditory scene in complex environments (Ulanovsky and Moss, 2008; Chili, Xian and Moss, 2009; Moss and Surlykke, 2010; Yovel and Ulanovsky, 2017; Salles, Diebold and Moss, 2020). Additionally, bats are known to produce echolocation sound groups when spatiotemporal localization demands are high (Kothari et al., 2014). Studies have documented call sequences ranging from 2 to 15 grouped calls (Moss et al., 2010), and it has been hypothesized that grouping facilitates echo segregation.
We did not use a single integration window - we tested integration sizes between 1 and 10 calls and presented the results in Figure 3A. This range was chosen based on prior empirical findings and to explore how different levels of temporal aggregation impact navigation performance. Indeed, the results showed that the performance levels between 5-10 calls integration window (Figure 3A)
Regarding the average exit time for 40 bats, this value was determined from our simulations, where it represents the mean time for successful exits under standard conditions with masking.
We have revised the text to clarify these details see, lines 466.
References:
Aidan, Y., Bleichman, I. and Ayali, A. (2024) ‘Pausing to swarm: locust intermittent motion is instrumental for swarming-related visual processing’, Biology letters, 20(2), p. 20230468. Available at: https://doi.org/10.1098/rsbl.2023.0468.
Attanasi, A. et al. (2014) ‘Collective Behaviour without Collective Order in Wild Swarms of Midges’. Edited by T. Vicsek, 10(7). Available at: https://doi.org/10.1371/journal.pcbi.1003697.
Bastien, R. and Romanczuk, P. (2020) ‘A model of collective behavior based purely on vision’, Science Advances, 6(6). Available at: https://doi.org/10.1126/sciadv.aay0792.
Beetz, M.J. and Hechavarría, J.C. (2022) ‘Neural Processing of Naturalistic Echolocation Signals in Bats’, Frontiers in Neural Circuits, 16, p. 899370. Available at: https://doi.org/10.3389/FNCIR.2022.899370/BIBTEX.
Betke, M. et al. (2008) ‘Thermal Imaging Reveals Significantly Smaller Brazilian Free-Tailed Bat Colonies Than Previously Estimated’, Journal of Mammalogy, 89(1), pp. 18–24. Available at: https://doi.org/10.1644/07-MAMM-A-011.1.
Bialek, W. et al. (2012) ‘Statistical mechanics for natural flocks of birds’, Proceedings of the National Academy of Sciences, 109(13), pp. 4786–4791. Available at: https://doi.org/10.1073/PNAS.1118633109.
Bode, N.W.F., Franks, D.W. and Wood, A.J. (2011) ‘Limited interactions in flocks: Relating model simulations to empirical data’, Journal of the Royal Society Interface, 8(55), pp. 301–304. Available at: https://doi.org/10.1098/RSIF.2010.0397.
Boerma, D.B. et al. (2019) ‘Wings as inertial appendages: How bats recover from aerial stumbles’, Journal of Experimental Biology, 222(20). Available at: https://doi.org/10.1242/JEB.204255/VIDEO-3.
Boonman, A.M., Parsons, S. and Jones, G. (2003) ‘The influence of flight speed on the ranging performance of bats using frequency modulated echolocation pulses’, The Journal of the Acoustical Society of America, 113(1), p. 617. Available at: https://doi.org/10.1121/1.1528175.
Burnett, S.C. and Masters, W.M. (2002) ‘Identifying Bats Using Computerized Analysis and Artificial Neural Networks’, North American Symposium on Bat Research, 9.
Cheraghi, A.R., Shahzad, S. and Graffi, K. (2022) ‘Past, Present, and Future of Swarm Robotics’, in Lecture Notes in Networks and Systems. Available at: https://doi.org/10.1007/978-3-030-82199-9_13.
Chili, C., Xian, W. and Moss, C.F. (2009) ‘Adaptive echolocation behavior in bats for the analysis of auditory scenes’, Journal of Experimental Biology, 212(9), pp. 1392–1404. Available at: https://doi.org/10.1242/jeb.027045.
Couzin, I.D. et al. (2002) ‘Collective Memory and Spatial Sorting in Animal Groups’, Journal of Theoretical Biology, 218(1), pp. 1–11. Available at: https://doi.org/10.1006/jtbi.2002.3065.
Couzin, I.D. et al. (2005) ‘Effective leadership and decision-making in animal groups on the move’, Nature, 433(7025), pp. 513–516. Available at: https://doi.org/10.1038/nature03236.
Davidson, J.D. et al. (2021) ‘Collective detection based on visual information in animal groups’, Journal of the Royal Society, 18(180), p. 2021.02.18.431380. Available at: https://doi.org/10.1098/rsif.2021.0142.
Faria Dias, P.G. et al. (2021) ‘Swarm robotics: A perspective on the latest reviewed concepts and applications’, Sensors. Available at: https://doi.org/10.3390/s21062062.
Fujioka, E. et al. (2021) ‘Three-Dimensional Trajectory Construction and Observation of Group Behavior of Wild Bats During Cave Emergence’, Journal of Robotics and Mechatronics, 33(3), pp. 556–563. Available at: https://doi.org/10.20965/jrm.2021.p0556.
Gautrais, J. et al. (2012) ‘Deciphering Interactions in Moving Animal Groups’, PLOS Computational Biology, 8(9), p. e1002678. Available at: https://doi.org/10.1371/JOURNAL.PCBI.1002678.
Gillam, E.H. et al. (2010) ‘Echolocation behavior of Brazilian free-tailed bats during dense emergence flights’, Journal of Mammalogy, 91(4), pp. 967–975. Available at: https://doi.org/10.1644/09-MAMM-A-302.1.
Goldstein, A. et al. (2024) ‘Collective Sensing – On-Board Recordings Reveal How Bats Maneuver Under Severe 4 Acoustic Interference’, Under Review, pp. 1–25.
Griffin, D.R., Webster, F.A. and Michael, C.R. (1958) ‘THE ECHOLOCATION OF FLYING INSECTS BY BATS ANIMAL BEHAVIOUR , Viii , 3-4’.
Hagino, T. et al. (2007) ‘Adaptive SONAR sounds by echolocating bats’, International Symposium on Underwater Technology, UT 2007 - International Workshop on Scientific Use of Submarine Cables and Related Technologies 2007, pp. 647–651. Available at: https://doi.org/10.1109/UT.2007.370829.
Hiryu, S. et al. (2008) ‘Adaptive echolocation sounds of insectivorous bats, Pipistrellus abramus, during foraging flights in the field’, The Journal of the Acoustical Society of America, 124(2), pp. EL51–EL56. Available at: https://doi.org/10.1121/1.2947629.
Jakobsen, L. et al. (2024) ‘Velocity as an overlooked driver in the echolocation behavior of aerial hawking vespertilionid bats’. Available at: https://doi.org/10.1016/j.cub.2024.12.042.
Jakobsen, L., Brinkløv, S. and Surlykke, A. (2013) ‘Intensity and directionality of bat echolocation signals’, Frontiers in Physiology, 4 APR(April), pp. 1–9. Available at: https://doi.org/10.3389/fphys.2013.00089.
Jakobsen, L. and Surlykke, A. (2010) ‘Vespertilionid bats control the width of their biosonar sound beam dynamically during prey pursuit’, 107(31). Available at: https://doi.org/10.1073/pnas.1006630107.
Jhawar, J. et al. (2020) ‘Noise-induced schooling of fish’, Nature Physics 2020 16:4, 16(4), pp. 488–493. Available at: https://doi.org/10.1038/s41567-020-0787-y.
Kalko, E.K. V. (1995) ‘Insect pursuit, prey capture and echolocation in pipistrelle bats (Microchirptera)’, Animal Behaviour, 50(4), pp. 861–880.
Kazial, K.A., Burnett, S.C. and Masters, W.M. (2001) ‘ Individual and Group Variation in Echolocation Calls of Big Brown Bats, Eptesicus Fuscus (Chiroptera: Vespertilionidae) ’, Journal of Mammalogy, 82(2), pp. 339–351. Available at: https://doi.org/10.1644/1545-1542(2001)082<0339:iagvie>2.0.co;2.
Kazial, K.A., Kenny, T.L. and Burnett, S.C. (2008) ‘Little brown bats (Myotis lucifugus) recognize individual identity of conspecifics using sonar calls’, Ethology, 114(5), pp. 469–478. Available at: https://doi.org/10.1111/j.1439-0310.2008.01483.x.
Kothari, N.B. et al. (2014) ‘Timing matters: Sonar call groups facilitate target localization in bats’, Frontiers in Physiology, 5 MAY. Available at: https://doi.org/10.3389/fphys.2014.00168.
Moss, C.F. and Surlykke, A. (2010) ‘Probing the natural scene by echolocation in bats’, Frontiers in Behavioral Neuroscience. Available at: https://doi.org/10.3389/fnbeh.2010.00033.
Nagy, M. et al. (2010) ‘Hierarchical group dynamics in pigeon flocks’, Nature 2010 464:7290, 464(7290), pp. 890–893. Available at: https://doi.org/10.1038/nature08891.
Neretti, N. et al. (2003) ‘Time-frequency model for echo-delay resolution in wideband biosonar’, The Journal of the Acoustical Society of America, 113(4), pp. 2137–2145. Available at: https://doi.org/10.1121/1.1554693.
Parrish, J.K. and Edelstein-Keshet, L. (1999) ‘Complexity, Pattern, and Evolutionary Trade-Offs in Animal Aggregation’, Science, 284(5411), pp. 99–101. Available at: https://doi.org/10.1126/SCIENCE.284.5411.99.
Partridge, B.L. (1982) ‘The Structure and Function of Fish Schools’, 246(6), pp. 114–123. Available at: https://doi.org/10.2307/24966618.
Pearce, D.J.G. et al. (2014) ‘Role of projection in the control of bird flocks’, Proceedings of the National Academy of Sciences of the United States of America, 111(29), pp. 10422–10426. Available at: https://doi.org/10.1073/pnas.1402202111.
Pitcher, T.J., Partridge, B.L. and Wardle, C.S. (1976) ‘A blind fish can school’, Science, 194(4268), pp. 963–965. Available at: https://doi.org/10.1126/science.982056.
Rosenthal, S.B., Twomey, C.R., Hartnett, A.T., Wu, H.S., Couzin, I.D., et al. (2015) ‘Revealing the hidden networks of interaction in mobile animal groups allows prediction of complex behavioral contagion’, Proceedings of the National Academy of Sciences of the United States of America, 112(15), pp. 4690–4695. Available at: https://doi.org/10.1073/pnas.1420068112.
Rosenthal, S.B., Twomey, C.R., Hartnett, A.T., Wu, H.S. and Couzin, I.D. (2015) ‘Revealing the hidden networks of interaction in mobile animal groups allows prediction of complex behavioral contagion’, Proceedings of the National Academy of Sciences of the United States of America, 112(15), pp. 4690–4695. Available at: https://doi.org/10.1073/PNAS.1420068112/-/DCSUPPLEMENTAL/PNAS.1420068112.SAPP.PDF.
Roy, S. et al. (2019) ‘Extracting interactions between flying bat pairs using model-free methods’, Entropy, 21(1). Available at: https://doi.org/10.3390/e21010042.
Sabol, B.M. and Hudson, M.K. (1995) ‘Technique using thermal infrared-imaging for estimating populations of gray bats’, Journal of Mammalogy, 76(4). Available at: https://doi.org/10.2307/1382618.
Saillant, P.A. et al. (1993) ‘A computational model of echo processing and acoustic imaging in frequency- modulated echolocating bats: The spectrogram correlation and transformation receiver’, The Journal of the Acoustical Society of America, 94(5). Available at: https://doi.org/10.1121/1.407353.
Salles, A., Diebold, C.A. and Moss, C.F. (2020) ‘Echolocating bats accumulate information from acoustic snapshots to predict auditory object motion’, Proceedings of the National Academy of Sciences of the United States of America, 117(46), pp. 29229–29238. Available at: https://doi.org/10.1073/PNAS.2011719117/SUPPL_FILE/PNAS.2011719117.SAPP.PDF.
Sanderson, M.I. et al. (2003) ‘Evaluation of an auditory model for echo delay accuracy in wideband biosonar’, The Journal of the Acoustical Society of America, 114(3), pp. 1648–1659. Available at: https://doi.org/10.1121/1.1598195.
Schnitzler, H., Bioscience, E.K.- and 2001, undefined (no date) ‘Echolocation by insect-eating bats: we define four distinct functional groups of bats and find differences in signal structure that correlate with the typical echolocation ’, academic.oup.comHU Schnitzler, EKV KalkoBioscience, 2001•academic.oup.com [Preprint]. Available at: https://academic.oup.com/bioscience/article-abstract/51/7/557/268230 (Accessed: 17 March 2025).
Schnitzler, H.-U. et al. (1987) ‘The echolocation and hunting behavior of the bat,Pipistrellus kuhli’, Journal of Comparative Physiology A, 161(2), pp. 267–274. Available at: https://doi.org/10.1007/BF00615246.
Simmons, J.A. and Kick, S.A. (1983) ‘Interception of Flying Insects by Bats’, Neuroethology and Behavioral Physiology, pp. 267–279. Available at: https://doi.org/10.1007/978-3-642-69271-0_20.
Strandburg-Peshkin, A. et al. (2013) ‘Visual sensory networks and effective information transfer in animal groups’, Current Biology. Cell Press. Available at: https://doi.org/10.1016/j.cub.2013.07.059.
Sumpter, D.J.T. et al. (2008) ‘Consensus Decision Making by Fish’, Current Biology, 18(22), pp. 1773–1777. Available at: https://doi.org/10.1016/J.CUB.2008.09.064.
Surlykke, A., Ghose, K. and Moss, C.F. (2009) ‘Acoustic scanning of natural scenes by echolocation in the big brown bat, Eptesicus fuscus’, Journal of Experimental Biology, 212(7), pp. 1011–1020. Available at: https://doi.org/10.1242/JEB.024620.
Theriault, D.H. et al. (no date) ‘Reconstruction and analysis of 3D trajectories of Brazilian free-tailed bats in flight’, cs-web.bu.edu [Preprint]. Available at: https://cs-web.bu.edu/faculty/betke/papers/2010-027-3d-bat-trajectories.pdf (Accessed: 4 May 2023).
Ulanovsky, N. and Moss, C.F. (2008) ‘What the bat’s voice tells the bat’s brain’, Proceedings of the National Academy of Sciences of the United States of America, 105(25), pp. 8491–8498. Available at: https://doi.org/10.1073/pnas.0703550105.
Vanderelst, D. and Peremans, H. (2018) ‘Modeling bat prey capture in echolocating bats : The feasibility of reactive pursuit’, Journal of theoretical biology, 456, pp. 305–314.
Youssefi, K.A.R. and Rouhani, M. (2021) ‘Swarm intelligence based robotic search in unknown maze-like environments’, Expert Systems with Applications, 178. Available at: https://doi.org/10.1016/j.eswa.2021.114907.
Yovel, Y. et al. (2009) ‘The voice of bats: How greater mouse-eared bats recognize individuals based on their echolocation calls’, PLoS Computational Biology, 5(6). Available at: https://doi.org/10.1371/journal.pcbi.1000400.
Yovel, Y. and Ulanovsky, N. (2017) ‘Bat Navigation’, The Curated Reference Collection in Neuroscience and Biobehavioral Psychology, pp. 333–345. Available at: https://doi.org/10.1016/B978-0-12-809324-5.21031-6.



