Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorJihwan ParkGwangju Institute of Science and Technology, Gwangju, Republic of Korea
- Senior EditorMa-Li WongState University of New York Upstate Medical University, Syracuse, United States of America
Reviewer #1 (Public review):
Summary:
The manuscript, titled Hippocampal Single-Cell RNA Atlas of Chronic Methamphetamine Abuse-Induced Cognitive Decline in Mice, focuses on single-cell RNA sequencing (scRNA-seq) analysis following chronic methamphetamine (METH) treatment in mice. The authors propose two hypotheses: (1) METH induces neuroinflammation involving T and NKT cells, and (2) METH alters neuronal stem cell differentiation.
Strengths:
The authors provide a substantial dataset with numerous replicates, offering valuable resources to the research community.
Weaknesses:
Concerns remain regarding the interpretation of the data and the appropriateness of the statistical analyses.
Although the authors provided detailed responses to the reviewer's concerns, I am still concerned that several key issues have not yet been fully addressed in the revised manuscript.
First, in Figure 5, the authors state that neural stem cells (NSCs) preferentially differentiate into astrocytes rather than neuroblasts following METH treatment. However, based on the presented trajectories, it is difficult to visually confirm differences in the relative proportions of astrocyte versus neuroblast differentiation between the control and METH-treated conditions. The current figures do not provide a quantitative or clearly interpretable comparison of lineage allocation that would support this conclusion.
Moreover, in Figures 5C and 5F, the inferred pseudotime trajectories differ both the starting cell populations and the intermediate and terminal cell identities. As a result, the trajectories are not directly comparable between the control and METH conditions. Under these circumstances, it is inappropriate to interpret gene expression changes as occurring along equivalent differentiation paths, and the current analysis does not convincingly support the stated conclusions regarding altered NSC differentiation.
If the authors intend to claim differential gene expression associated with altered differentiation trajectories, the analysis should at minimum present the expression of the same set of genes (e.g., Bsg, Ccl4, Fos, Sox11, Flt1, Hspb1, Igfbp7, and Tmsb10) plotted along a matched trajectory (for example, NSC-to-astrocyte or NSC-to-neuroblast lineages) in both control and METH-treated samples, so that readers can directly compare expression dynamics across conditions.
In addition, several statements throughout the manuscript describing changes in cell-type proportions are not supported by corresponding statistical analyses. For example, in Figure 2C (around line 430), the authors report changes in cell proportions of ~0.1% or 2-3%. Without appropriate statistical testing, it is unclear whether such marginal differences are biologically meaningful or reproducible. The authors should either provide statistical testing (e.g., sample-level proportion analysis with p-values or confidence intervals) or revise the text to describe these findings as descriptive rather than significant changes.
Finally, the reported decrease in astrocyte proportion following METH exposure (from 6.6% to 5.5%), together with the lack of reported changes in neuroblast proportions, appears inconsistent with the trajectory-based conclusion that NSCs preferentially differentiate into astrocytes in METH-treated mice. This apparent discrepancy should be clarified or the conclusions appropriately tempered.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
(1) Concerns persist regarding the interpretation of data and the validation of experiments. First, the presence of T cells, NKT cells, and neutrophils in both the control and METH-treated hippocampi suggests that blood contamination rather than immune cell infiltration is the cause. Since the authors claim that METH disrupts the blood-brain barrier, increasing the infiltration of these immune cells, identifying the source of these immune cells is critical.
We sincerely appreciate the valuable suggestions you have provided. Your professional perspective impresses us. Based on your suggestion, we conducted a systematic review and in-depth analysis of the experimental process.
As you have pointed out, we believe that the T cells, NK cells and neutrophils detected in the single-cell sequencing of the mouse hippocampus may have a blood-derived origin. However, this does not mean that the presence of these cell types in the control group is abnormal, because in many literature, these cells can also be found in the hippocampus of control mice. Nevertheless, clarifying the origin and location of these cells will help to further strengthen the persuasiveness of the research hypothesis. Although there is currently no systematic discussion on the role of such cells in the field of methamphetamine neurotoxicity research, we believe that the relevant findings still have certain reference value for subsequent research in this field.
Our response is based on the following description:
(1) Insufficient perfusion during the extraction of the hippocampus may lead to a certain degree of blood contamination.
Given that the single-cell sequencing technique employed in this study can detect all the mRNA of the entire cell, in order to ensure that the cells are in the optimal physiological state and to minimize the stress response caused by the experimental operation on the cells, we perfused the anesthetized mice with cold PBS for approximately 3 min (this has been supplemented in the Materials and methods Line165-166), and completed the rapid dissection and collection of the mouse hippocampus on the ice surface within 2 min, and immediately placed it in an appropriate amount of tissue preservation solution for storage. The time of tissue perfusion might be insufficient or the perfusion volume might not be adequate, resulting in the incomplete expulsion of all the blood. Subsequently, the decomposition operations of the tissue samples were all carried out in the preservation solution or PBS buffer, which to some extent reduced the potential interference of blood components on the experimental results. Additionally, T cells, NKT cells and neutrophils in the capillary perivascular spaces of the hippocampal tissue might still remain and be successfully captured, and were reflected in the final sequencing data.
(2) The presence of T cells, NKT cells, and neutrophils in the brain tissue of normal mice has been supported by existing literature. Moreover, several studies have specifically described the localization of these immune cell types within the brain parenchyma.
Contemporary studies have completely changed the view of brain immunity from envisioning the brain as isolated and inaccessible to peripheral immune cells to an organ in close physical and functional communication with the immune system for its maintenance, function, and repair. Circulating immune cells reside in special niches in the brain’s borders, the choroid plexus, meninges, and perivascular spaces, from which they patrol and sense the brain in a remote manner [1].
A large-scale mouse brain cell atlas study also reported that approximately 8% of non-neuronal cells are immune cells, including microglia, boundary-associated macrophages, lymphocytes, dendritic cells, and monocytes [2].
Hang Yao et al. demonstrated through flow cytometry that neutrophils were present in the hippocampal tissues of both healthy control mice and depressed mice (Fig.2 H) [3]. Wei Su et al. identified through single-cell sequencing that dendritic cells, neutrophils, macrophages, T cells, and NKT cells were present in the brain tissues of non-transgenic (Non-Tg) control mice (Fig.1a-b), and the localization of these cells was explicitly characterized as brain parenchyma in the study [4]. Tomomi M Yoshida et al. discovered through immunohistochemistry (IHC) and single-cell sequencing techniques that there were a certain number of CD3+ and CD4+ T cells in the hippocampus and other regions of the brain, and they observed that these cells were located outside the blood vessels. (Fig.1a-c, g) [5].
(3) Both the analysis of immune cells within blood vessels and those in the brain parenchyma contribute to elucidating the immune effects in the hippocampal microenvironment under chronic METH exposure, as well as their interactions with other cell types. At present, the understanding of the neurotoxicity of methylphenidate and the immune system is still limited to the central resident immune cells, such as microglia, astrocytes and oligodendrocytes [6]. Adaptive immune cells and myeloid cells recruited from the circulation have also been implicated in brain development, function, and aging. Their depletion during developmental stages can disrupt critical neural processes, including glial cell maturation, neuronal activity, and myelinogenesis. However, the precise developmental stage at which lymphocyte infiltration into the central nervous system occurs remains to be elucidated [7].
Our data results indicate that during chronic METH abuse, T cells are more active and participate in the regulation of cytokines through complement signaling. At the same time, the frequency of cell communication between endothelial cells and epithelial cells is increased. Moreover, microglia upregulated the processes of cell chemotaxis and migration, as well as the communication with immune cells such as T cells, and to some extent, this also suggests an enhanced infiltration of T cells. However, we also recognize that the current conclusions regarding immune cell infiltration based on sequencing data and literature reports lack the support of experimental data. Currently, we are conducting morphological analysis using the same batch of brain tissue samples to further validate the relevant findings.
Immune fluorescence staining and flow cytometry can be utilized to further determine the locations of these immune cells in the hippocampus. The classical pathways through which peripheral immune cells enter the brain mainly include the BBB and the choroid plexus. In June 2025, Kim N. Green et al. published a study in Neuron, further revealing that during the developmental stage and in cases of inflammatory diseases, immune cells can also infiltrate the brain parenchyma through a newly identified channel - the medial ventricle, thereby further confirming that these cells have the ability to migrate to the central nervous system under specific physiological or pathological conditions [8].
(1) Castellani G, Croese T, Peralta Ramos JM, Schwartz M. Transforming the understanding of brain immunity. Science. 2023 Apr 7;380(6640):eabo7649. doi: 10.1126/science.abo7649.
(2) Zhang M, Pan X, Jung W, Halpern AR, Eichhorn SW, Lei Z, Cohen L, Smith KA, Tasic B, Yao Z, Zeng H, Zhuang X. Molecularly defined and spatially resolved cell atlas of the whole mouse brain. Nature. 2023 Dec;624(7991):343-354. doi: 10.1038/s41586-023-06808-9.
(3) Yao H, Jiang SY, Jiao YY, Zhou ZY, Zhu Z, Wang C, Zhang KZ, Ma TF, Hu G, Du RH, Lu M. Astrocyte-derived CCL5-mediated CCR5+ neutrophil infiltration drives depression pathogenesis. Sci Adv. 2025 May 23;11(21):eadt6632. doi: 10.1126/sciadv.adt6632.
(4) Su W, Saravia J, Risch I, Rankin S, Guy C, Chapman NM, Shi H, Sun Y, Kc A, Li W, Huang H, Lim SA, Hu H, Wang Y, Liu D, Jiao Y, Chen PC, Soliman H, Yan KK, Zhang J, Vogel P, Liu X, Serrano GE, Beach TG, Yu J, Peng J, Chi H. CXCR6 orchestrates brain CD8+ T cell residency and limits mouse Alzheimer's disease pathology. Nat Immunol. 2023 Oct;24(10):1735-1747. doi: 10.1038/s41590-023-01604-z.
(5) Yoshida TM, Nguyen M, Zhang L, Lu BY, Zhu B, Murray KN, Mineur YS, Zhang C, Xu D, Lin E, Luchsinger J, Bhatta S, Waizman DA, Coden ME, Ma Y, Israni-Winger K, Russo A, Wang H, Song W, Al Souz J, Zhao H, Craft JE, Picciotto MR, Grutzendler J, Distasio M, Palm NW, Hafler DA, Wang A. The subfornical organ is a nucleus for gut-derived T cells that regulate behaviour. Nature. 2025 Jul;643(8071):499-508. doi: 10.1038/s41586-025-09050-7.
(6) Shi S, Sun Y, Zan G, Zhao M. The interaction between central and peripheral immune systems in methamphetamine use disorder: current status and future directions. J Neuroinflammation. 2025 Feb 15;22(1):40. doi: 10.1186/s12974-025-03372-z.
(7) Castellani G, Croese T, Peralta Ramos JM, Schwartz M. Transforming the understanding of brain immunity. Science. 2023 Apr 7;380(6640):eabo7649. doi: 10.1126/science.abo7649.
(8) Hohsfield LA, Kim SJ, Barahona RA, Henningfield CM, Mansour K, Vallejo KD, Tsourmas KI, Kwang NE, Ghorbanian Y, Angulo JAA, Gao P, Pachow C, Inlay MA, Walsh CM, Xu X, Lane TE, Green KN. Identification of the velum interpositum as a meningeal-CNS route for myeloid cell trafficking into the brain. Neuron. 2025 May 28:S0896-6273(25)00351-4. doi: 10.1016/j.neuron.2025.05.004.
(2) Secondly, the pseudotime analysis, which suggests altered neural stem cell (NSC) differentiation, is not conclusively supported by the current data and requires further validation.
We sincerely appreciate your valuable feedback, which we find highly relevant and constructive. It is important to acknowledge that the sequencing data presented in our study currently lacks experimental validation. Nevertheless, considering that existing research on the effects of METH on neural stem cell differentiation predominantly emphasizes observational phenomena and remains limited in terms of in vivo experimental evidence and mechanistic investigations, we aim to contribute our analytical findings as a reference for further scholarly exploration in this field.
Our study utilized pseudotime analysis (powered by Monocle2) to reconstruct an "imaginary timeline" (pseudo-time) based on intercellular gene expression similarities, thereby modeling the dynamic state transitions of cells during continuous biological processes. Drawing upon single-cell RNA sequencing data captured as "snapshots" from hippocampal astrocytes, neural stem cells, and neuroblasts in mice four weeks after METH exposure, we applied computational algorithms to integrate the originally discrete cellular states into a continuous pseudo-time trajectory. This approach was employed to elucidate the differentiation stages of these cell populations, identify potential branching points in their developmental pathways, and uncover the key regulatory genes driving the differentiation process. Pseudotime analysis, as a computational approach grounded in mathematical modeling, yields inferences that are contingent upon the underlying assumptions of the algorithms employed. Consequently, experimental validation through methodologies such as time-series sampling and lineage tracing is essential to substantiate the derived biological interpretations. In light of the insufficiency of such empirical verification to date, our conclusions concerning alterations in the dynamic behavior of neural stem cell differentiation remain preliminary and require further experimental support.
In Figures 5C and 5F, we present the expression profiles of the four genes exhibiting the most statistically significant differences across the differentiation trajectory. In Figures 5B and 5E, we conducted GO and KEGG functional enrichment analyses on the genes that showed significant differential expression at different differentiation stages. While no studies within the current METH research domain have reported on the potential effects of these genes on neural stem cell differentiation, emerging evidence from related fields provides preliminary insights into their functional roles. For instance, the Flt1 gene (also known as VEGFR1), referred to as the vascular endothelial growth factor receptor, has been demonstrated to play a critical role in the conversion of Müller glial cells into neurons within the zebrafish retina [1], serves as a critical regulator in promoting definitive neural stem cell survival [2]. Furthermore, it substantiates the intricate interconnection between neurons, neural stem cells, and vascular cells, as identified in our cell communication analysis. Hsp1b gene plays a significant role in ferroptosis and autophagy processes of nerve cells[3, 4], and may be closely related to the self-renewal ability of neural stem cell, while METH may impair neural stem cell function by disrupting autophagy, leading to reduced self-renewal capacity and altered differentiation potential [5]. In METH group, Sox11 has been shown to play a critical role in early differentiation and neuronal growth, both during perinatal development and in adult neurogenesis [6] Fos gene plays a critical regulatory role in the differentiation of neural stem cells into neurons and in modulating neuronal functional activities [7]; Alterations in Ccl5 expression levels may indicate astrocyte-mediated inflammatory responses, which could represent one of the underlying mechanisms through which METH promotes the differentiation of neural stem cells into astrocytes.
Thank you very much for your thoughtful questions and valuable suggestions. These suggestions have helped us gain a deeper understanding of the areas where we can improve, and have guided us toward more meaningful directions for future research.
(1) Mitra S, Devi S, Lee MS, Jui J, Sahu A, Goldman D. Vegf signaling between Müller glia and vascular endothelial cells is regulated by immune cells and stimulates retina regeneration. Proc Natl Acad Sci U S A. 2022 Dec 13;119(50):e2211690119. doi: 10.1073/pnas.2211690119.
(2) Wada T, Haigh JJ, Ema M, Hitoshi S, Chaddah R, Rossant J, Nagy A, van der Kooy D. Vascular endothelial growth factor directly inhibits primitive neural stem cell survival but promotes definitive neural stem cell survival. J Neurosci. 2006 Jun 21;26(25):6803-12. doi: 10.1523/JNEUROSCI.0526-06.2006.
(3) Meng J, Fang J, Bao Y, Chen H, Hu X, Wang Z, Li M, Cheng Q, Dong Y, Yang X, Zou Y, Zhao D, Tang J, Zhang W, Chen C. The biphasic role of Hspb1 on ferroptotic cell death in Parkinson's disease. Theranostics. 2024 Aug 1;14(12):4643-4666. doi: 10.7150/thno.98457.
(4) Sisto A, van Wermeskerken T, Pancher M, Gatto P, Asselbergh B, Assunção Carreira ÁS, De Winter V, Adami V, Provenzani A, Timmerman V. Autophagy induction by piplartine ameliorates axonal degeneration caused by mutant HSPB1 and HSPB8 in Charcot-Marie-Tooth type 2 neuropathies. Autophagy. 2025 May;21(5):1116-1143. doi: 10.1080/15548627.2024.2439649.
(5) Gu C, Wang Z, Luo W, Ling H, Cui X, Deng T, Li K, Huang W, Xie Q, Tao B, Qi X, Peng X, Ding J, Qiu P. Impaired olfactory bulb neurogenesis mediated by Notch1 contributes to olfactory dysfunction in mice chronically exposed to methamphetamine. Cell Biol Toxicol. 2025 Feb 20;41(1):46. doi: 10.1007/s10565-025-10004-y.
(6) Rasetto NB, Giacomini D, Berardino AA, Waichman TV, Beckel MS, Di Bella DJ, Brown J, Davies-Sala MG, Gerhardinger C, Lie DC, Arlotta P, Chernomoretz A, Schinder AF. Transcriptional dynamics orchestrating the development and integration of neurons born in the adult hippocampus. Sci Adv. 2024 Jul 19;10(29):eadp6039. doi: 10.1126/sciadv.adp6039.
(7) Pagin M, Pernebrink M, Pitasi M, Malighetti F, Ngan CY, Ottolenghi S, Pavesi G, Cantù C, Nicolis SK. FOS Rescues Neuronal Differentiation of Sox2-Deleted Neural Stem Cells by Genome-Wide Regulation of Common SOX2 and AP1(FOS-JUN) Target Genes. Cells. 2021 Jul 12;10(7):1757. doi: 10.3390/cells10071757.
Reviewer #2 (Public review):
(1) Despite this potential novelty, the study has numerous weaknesses. Notably, single-cell RNA sequencing was unable to capture an adequate number of neuronal populations. Neurons accounted for only approximately 0.6% of the total nuclei, representing a significant underrepresentation compared to their actual physiological proportion. Given that the behavioral effects of METH are likely mediated by neuronal dysfunction, readers would reasonably expect to see transcriptional changes in neurons. The authors should explain why they were unable to capture a sufficient number of neurons and justify how this incomplete dataset can still provide meaningful scientific insights for researchers studying METH-induced hippocampal damage and behavioral alterations.
Thank you sincerely for bringing this important issue to our attention.
Firstly, this represents an unavoidable technical bottleneck. The single-cell sequencing (scRNA-seq) we perform involves the detection of mRNA at the whole-cell level, a process that necessitates cells with high structural integrity, robust viability, and minimal exposure to external stimuli. During the preparation of single-cell suspensions, mature neurons due to their highly differentiated state, morphological rigidity, and excessively long axons often fail to maintain structural integrity. These cells typically undergo death during the dissociation process, lose viability, and are subsequently excluded prior to sequencing. To retain a substantial amount of neuron-related data, an alternative technique single-cell nuclear sequencing (snRNA-seq) should be employed. This method does not necessitate cell viability and focuses exclusively on the nuclei of individual cells, thereby capturing mRNA information solely from the nuclear compartment. Consequently, mRNA data originating from the cytoplasm and organelles will not be represented.
Secondly, numerous studies have shown that the neurological damage caused by chronic exposure to methamphetamine exhibits a high degree of similarity in clinical manifestations and pathogenesis to neurodegenerative diseases (such as Alzheimer's disease, Parkinson's disease, etc.) [1-4].
We fully acknowledge the central role of neurons in cognitive functions and the pathogenesis of cognitive disorders. However, despite decades of neuron-centric research that has yielded significant advancements, major challenges remain in elucidating disease origins, identifying early pathological events, and developing effective therapeutic strategies. For example, current models fail to adequately explain early disease events. Many pathological hallmarks of cognitive disorders such as amyloid plaques, neurofibrillary tangles, and α-synuclein aggregation emerge in the extracellular space long before overt neuronal loss or dysfunction occurs, and are increasingly recognized to be initiated or modulated by non-neuronal cells, including astrocytes and microglia [5]. Furthermore, the critical contribution of the neural microenvironment is often overlooked. Neuronal function and survival are highly dependent on this microenvironment, which is predominantly established and maintained by non-neuronal cell types such as astrocytes, oligodendrocytes, microglia, vascular endothelial cells, pericytes, and interstitial cells and matrix [6-10]. Additionally, systemic factors such as metabolic dysregulation, peripheral inflammation, and vascular pathology are closely associated with cognitive disorders. These factors often initially impact non-neuronal cells, particularly those forming the blood-brain barrier (e.g., endothelial cells) or mediating immune responses (e.g., microglia), before exerting downstream effects on neurons [11,12]. Finally, current therapeutic approaches for neuron face significant limitations, highlighting an urgent need for novel intervention strategies.
During the development of neurodegenerative chronic diseases, although the structural or functional abnormalities of neurons are the direct factors leading to clinical symptoms (such as cognitive decline), this process is often regulated by various auxiliary cell types such as glial cells, immune cells, and stromal cells, and constitutes a complex pathological mechanism network. It is worth noting that the chronic and persistent progression of the disease usually results from the failure of these auxiliary cells to effectively provide support and nutrition to neurons, and even in some pathological states, they transform into effector cells that promote neuronal damage [13,14]. In recent years, a growing number of evidence has demonstrated that glial cells, immune cells, and stromal cells exert critical regulatory functions in the pathogenesis of neurodegenerative diseases. These cell types not only contribute to the maintenance of neural microenvironmental homeostasis during the early stages of disease progression but also display substantial functional heterogeneity in modulating inflammatory responses, synaptic plasticity, the repair of neuronal injury, linking genetic risks with environmental factors and the pathogenic mechanism of pathological protein propagation [15-19]. These research results indicate that they have the potential to become key therapeutic targets in clinical interventions: 1. compared to neurons themselves, they are more susceptible to being targeted by drugs or biological agents (such as antibodies), and have higher accessibility; 2. Non-neuronal cells (especially glial cells) exhibit high plasticity and reactivity during the course of diseases, providing an opportunity window for intervening in their functional states (such as inhibiting harmful activation and promoting protective functions); 3. they can serve as early intervention targets before irreversible damage occurs to neurons, helping to prevent or delay the progression of the disease;4. intervention methods targeting these targets are diverse, including immunomodulation, anti-inflammatory, vascular protection, and metabolic regulation strategies, which are usually more feasible in practical applications than directly protecting the fragile neurons.
Early pharmacological studies have extensively characterized the neurotoxic effects of METH, including the induction of autophagy, apoptosis, oxidative stress, endoplasmic reticulum stress, and dopaminergic neurotoxicity [20]. However, therapeutic options and pharmacological interventions for METH abuse remain limited [21]. In recent years, increasing attention has been directed toward the impact of METH on non-neuronal cells. Research into mechanisms such as neuroinflammatory responses, blood-brain barrier disruption, and immune modulation is progressively contributing to a more comprehensive understanding of METH-induced neural injury [22-24]. Moreover, METH is a substance that induces widespread damage across multiple organ systems and diverse cell types throughout the body. Beyond its effects on neurons, various cell types exhibit distinct responses to METH exposure, which differ significantly depending on the duration of exposure. Our research dataset encompasses high-quality whole-cell mRNA sequencing data from multiple cell types within the hippocampus of mice subjected to chronic METH exposure, offering substantial data support and a robust foundation for in-depth investigation into the pathological mechanisms underlying METH-induced neurodamage.
Thirdly, the selection of scRNA-seq was guided by our experimental objectives and prior research experience. Our earlier investigations have primarily centered on astrocytes, endothelial cells, and microglia. This single-cell sequencing study is intended to enhance our understanding of these neural support cells, comprehensively explore their underlying mechanisms and cellular interactions, and ultimately provide a solid foundation and reference for future research. However, our experience and infrastructure in the field of neuronal research remain relatively limited. To ensure the generation of high-quality data and to systematically advance the experimental objectives, we have prioritized the analysis of the neural microenvironment as the central focus of this study.
Fourthly, the hippocampal region is a brain area with highly specialized and collaborative characteristics, which can be further divided into the ventral hippocampus, the dorsal hippocampus, and multiple subregions such as DG, CA1, CA2, and CA3. The neurons in these subregions exhibit strong heterogeneity, and the experimental methods we currently adopt are still unable to precisely distinguish the neurons in these different regions, which may to some extent affect the accuracy of data interpretation. To address the impact of neuronal heterogeneity, we believe that single-cell spatial transcriptomics technology can be adopted for in-depth research. However, due to the high cost of this technology, it is currently difficult to apply it in our research group.
(1) Lappin JM. Rare but relevant: Methamphetamine and Parkinson's disease. Addiction. 2025 Apr;120(4):797-800. doi: 10.1111/add.16695. Epub 2024 Oct 22. PMID: 39434702.
(2) Lappin JM, Darke S. Methamphetamine and heightened risk for early-onset stroke and Parkinson's disease: A review. Exp Neurol. 2021 Sep;343:113793. doi: 10.1016/j.expneurol.2021.113793. Epub 2021 Jun 21. PMID: 34166684.
(3) Shukla M, Vincent B. The multi-faceted impact of methamphetamine on Alzheimer's disease: From a triggering role to a possible therapeutic use. Ageing Res Rev. 2020 Jul;60:101062. doi: 10.1016/j.arr.2020.101062.
(4) Shrestha P, Katila N, Lee S, Seo JH, Jeong JH, Yook S. Methamphetamine induced neurotoxic diseases, molecular mechanism, and current treatment strategies. Biomed Pharmacother. 2022 Oct;154:113591. doi: 10.1016/j.biopha.2022.113591.
(5) Gabitto MI, et al.. Integrated multimodal cell atlas of Alzheimer's disease. Nat Neurosci. 2024 Dec;27(12):2366-2383. doi: 10.1038/s41593-024-01774-5.
(6) Stogsdill JA, Harwell CC, Goldman SA. Astrocytes as master modulators of neural networks: Synaptic functions and disease-associated dysfunction of astrocytes. Ann N Y Acad Sci. 2023 Jul;1525(1):41-60. doi: 10.1111/nyas.15004.
(7) Terreros-Roncal J, et al.. Impact of neurodegenerative diseases on human adult hippocampal neurogenesis. Science. 2021 Nov 26;374(6571):1106-1113. doi: 10.1126/science.abl5163.
(8) Zhu K, Fu Y, Zhao Y, Niu B, Lu H. Perineuronal nets: Role in normal brain physiology and aging, and pathology of various diseases. Ageing Res Rev. 2025 Jun;108:102756. doi: 10.1016/j.arr.2025.102756.
(9) Depp C, Doman JL, Hingerl M, Xia J, Stevens B. Microglia transcriptional states and their functional significance: Context drives diversity. Immunity. 2025 May 13;58(5):1052-1067. doi: 10.1016/j.immuni.2025.04.009.
(10) Sweeney MD, Zhao Z, Montagne A, Nelson AR, Zlokovic BV. Blood-Brain Barrier: From Physiology to Disease and Back. Physiol Rev. 2019 Jan 1;99(1):21-78. doi: 10.1152/physrev.00050.2017.
(11) Nation DA, et al.. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat Med. 2019 Feb;25(2):270-276. doi: 10.1038/s41591-018-0297-y.
(12) Montagne A, Zhao Z, Zlokovic BV. Alzheimer's disease: A matter of blood-brain barrier dysfunction? J Exp Med. 2017 Nov 6;214(11):3151-3169. doi: 10.1084/jem.20171406. Epub 2017 Oct 23.
(13) Huang Q, Wang Y, Chen S, Liang F. Glycometabolic Reprogramming of Microglia in Neurodegenerative Diseases: Insights from Neuroinflammation. Aging Dis. 2024 May 7;15(3):1155-1175. doi: 10.14336/AD.2023.0807.
(14) Shi FD, Yong VW. Neuroinflammation across neurological diseases. Science. 2025 Jun 19;388(6753):eadx0043. doi: 10.1126/science.adx0043.
(15) Xu X, Mei B, Yang Y, Li J, Weng J, Yang Y, Zhu Q, Zhang H, Liu X. Astrocytes Lingering at a Crossroads: Neuroprotection and Neurodegeneration in Neurocognitive Dysfunction. Int J Biol Sci. 2025 Apr 28;21(7):3122-3143. doi: 10.7150/ijbs.109315.
(16) Bedolla A, et al.. Adult microglial TGFβ1 is required for microglia homeostasis via an autocrine mechanism to maintain cognitive function in mice. Nat Commun. 2024 Jun 21;15(1):5306. doi: 10.1038/s41467-024-49596-0.
(17) Castellani G, Croese T, Peralta Ramos JM, Schwartz M. Transforming the understanding of brain immunity. Science. 2023 Apr 7;380(6640):eabo7649. doi: 10.1126/science.abo7649.
(18) Chen YH, Jin SY, Yang JM, Gao TM. The Memory Orchestra: Contribution of Astrocytes. Neurosci Bull. 2023 Mar;39(3):409-424. doi: 10.1007/s12264-023-01024-x.
(19) Deng Q, Wu C, Parker E, Liu TC, Duan R, Yang L. Microglia and Astrocytes in Alzheimer's Disease: Significance and Summary of Recent Advances. Aging Dis. 2024 Aug 1;15(4):1537-1564. doi: 10.14336/AD.2023.0907.
(20) Jayanthi S, Daiwile AP, Cadet JL. Neurotoxicity of methamphetamine: Main effects and mechanisms. Exp Neurol. 2021 Oct;344:113795. doi: 10.1016/j.expneurol.2021.113795.
(21) Paulus MP, Stewart JL. Neurobiology, Clinical Presentation, and Treatment of Methamphetamine Use Disorder: A Review. JAMA Psychiatry. 2020 Sep 1;77(9):959-966. doi: 10.1001/jamapsychiatry.2020.0246.
(22) Shi S, Sun Y, Zan G, Zhao M. The interaction between central and peripheral immune systems in methamphetamine use disorder: current status and future directions. J Neuroinflammation. 2025 Feb 15;22(1):40. doi: 10.1186/s12974-025-03372-z.
(23) Pang L, Wang Y. Overview of blood-brain barrier dysfunction in methamphetamine abuse. Biomed Pharmacother. 2023 May;161:114478. doi: 10.1016/j.biopha.2023.114478.
(24) Shaerzadeh F, Streit WJ, Heysieattalab S, Khoshbouei H. Methamphetamine neurotoxicity, microglia, and neuroinflammation. J Neuroinflammation. 2018 Dec 12;15(1):341. doi: 10.1186/s12974-018-1385-0.
(2) Another significant weakness of this study is the lack of a cohesive hypothesis or overarching conclusion regarding how METH impacts neural populations. The authors provide a largely descriptive account of transcriptional alterations across various cell types, but the manuscript lacks clear, biologically meaningful conclusions. This descriptive approach makes it difficult for readers to identify the key findings or take-home messages. To improve clarity and impact, the authors should focus on developing and presenting a few plausible hypotheses or mechanistic scenarios regarding METH-induced neurotoxicity, grounded in their scRNA-seq data. Including schematic figures to illustrate these hypotheses would also help readers better understand and interpret the study.
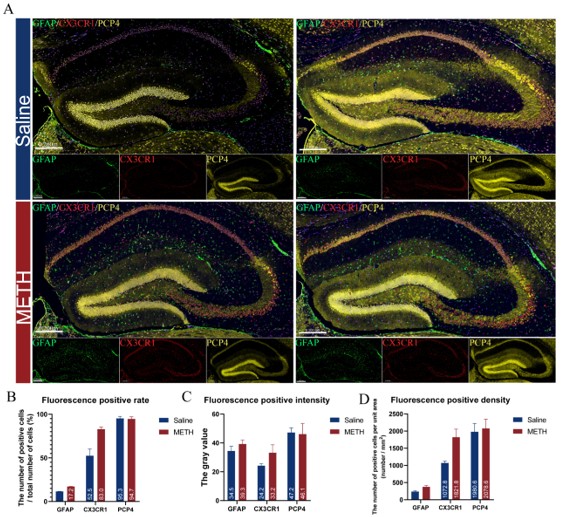
We sincerely appreciate your valuable comments on our article. As you pointed out, the current research lacks experimental verification to further support our conclusions. To enhance the clarity and readability of the mechanism explanation, we have added several hypothetical diagrams (such as Figures.7, 8, and 9) in the discussion section to present the biological mechanisms reflected by the data more intuitively. Additionally, relevant verification work is underway, such as marking specific cell types with marker proteins. Author response image 1 shows some of our preliminary experimental results that have not been published yet, and their trends are consistent with the conclusions of this article. However, since the complete verification still requires a certain period of time, to ensure the rigor of the data, these results have not been included in the current manuscript for the time being. Finally, we would like to thank you again for your constructive suggestions.
Author response image 1.
(3) The final major weakness of this study is its poor readability. It appears that the authors did not adequately proofread the manuscript, as there are numerous typographical errors (e.g., line 333: trisulting; line 756: essencial), unsupported scientific claims lacking citations (e.g., lines 485, 503, 749-753), and grammatically incorrect sentences (e.g., lines 470-472, 540-543, 749-753). In addition, many paragraphs are unorganized and overly descriptive, which further hinders clarity. Some figures are also problematic - too small in size and overcrowded with text in fonts that are difficult to read. It is recommended that the authors carry out quality control. There are too many typographical and grammatical errors to list individually; the authors should carefully review and revise the entire manuscript to address all of these issues.
We truly appreciate your thoughtful feedback and sincerely apologize for any inconvenience experienced by you and other readers.
The text of this research manuscript was manually entered, which unfortunately resulted in some spelling and grammatical errors. In response, we have carefully revised the entire manuscript using word processing tools in the second version. Meanwhile, we have restructured and organized some lengthy paragraphs to enhance the clarity and readability of the content.
Regarding the issue you raised about certain viewpoints lacking citation support, we have added the necessary references to those sections and reviewed the entire text to ensure all scientific claims are properly supported.
As for the image clarity, we made sure the submitted images met the 600dpi resolution requirement. However, we acknowledge that there were clarity issues in the final published version. We have since re-adjusted and re-uploaded the images to improve their quality.
We are committed to continuously improving the manuscript and enhancing the overall quality of our academic presentation. Thank you sincerely for your kind attention to our work, your careful review, and the valuable suggestions you provided.
Reviewer #3 (Public review):
(1) While the bioinformatics analyses are extensive, the study is primarily descriptive at the molecular level. The absence of experimental validation, such as targeted mRNA/protein quantification and gene knockdown/overexpression to confirm the causal relationship between these identified genes and METH-induced cognitive deficits, is a notable limitation.
We sincerely appreciate your valuable comments and suggestions. Indeed, there are still certain limitations in our manuscript in some aspects. It may not be able to systematically answer specific questions, and it is also difficult to fully clarify the functional roles of certain genes or specific cell types through experimental evidence.
Although our manuscript still has certain limitations, we believe that the publication of this research is expected to provide new perspectives and theoretical support for the in-depth exploration of METH toxicity damage-related fields, thereby promoting the progress of research in this direction:
(1) At present, the single-cell sequencing datasets on chronic damage caused by METH are still relatively limited, especially in terms of studies at the whole-cell level. Our dataset is expected to fill the research gap in this field to some extent, providing reference and support for subsequent related research.
(2) During the sampling process of the sequencing experiment, we ensured high cell viability and sequencing quality. The experiment exhibited good reproducibility (each group consisted of 10 mice, and 2 mice from each group were selected to mix their hippocampal tissues into one sample), and the obtained data had high credibility.
(3) The effects of METH have a wide distribution pattern across various organs and tissues. Through single-cell sequencing data, the common and differential expression patterns of related genes under different conditions can be systematically analyzed, which is helpful for future targeted knockout studies of these genes and provides a predictive basis for the evaluation of intervention measures, thereby enabling precise regulation of gene functions.
(4) This is conducive to the orderly implementation of our subsequent research plans. Our subsequent research plan can be further developed based on a specific aspect of this study. We are indeed planning to do exactly that. During our earlier research on astrocytes, we discovered that astrocytes have two phenotypes (protective and inflammatory) in neuroinflammation. Given that astrocytes in the hippocampus show great variability depending on their location, the cells they come into contact with, and the stimuli they receive, we aim to investigate the changes in the function of astrocyte subpopulations in chronic METH-induced cognitive impairment. We focused on the role of the cAMP signaling pathway in the transformation of astrocyte phenotypes and attempted to link changes in astrocyte energy metabolism to their inflammatory phenotype. In addition, we found that endothelial cells can be easily distinguished into many subpopulations, which are related to their specific functions in immune responses, material transport, vascular growth regulation, energy metabolism, and other processes. We believe that single-cell technology can help us find the key mechanisms and intervention targets of chronic METH abuse-induced damage with greater precision.
(2) While the discussion extensively covers the functional implications of specific molecular pathways and cell types, it would greatly benefit from a comparison of these findings with existing RNA sequencing data from other METH models in hippocampal tissue.
We are very grateful for your professional suggestions, which have been of great help in improving the quality of our manuscript. We agree that comparing our findings with existing RNA sequencing data from other METH models in hippocampal tissue would strengthen the discussion. In response to your suggestion, we have actively reviewed relevant literature and databases, and attempted to request the database administrators and original authors for the download and use of the relevant data. However, as data integration still requires some time, we may not be able to conduct a detailed analysis of the data in this revised version. We can only discuss the conclusions of some authors.
Palsamy Periyasamy et al. published a scRNA sequencing (live-cell) study on chronic METH exposure almost at the same time as us. They also adopted a similar gradual incremental 4-week METH exposure model and conducted sequencing analysis on glial cells in the cerebral cortex of mice [1]. The changes they observed in the circadian rhythm, adherens junctions, Rap1 signaling pathway, and cAMP signaling pathway (Disscusion, Lines 892-897) in the cortical astrocytes were also similar in the astrocytes of the hippocampal region that we studied. Similarly, in oligodendrocytes, we observed an upregulation trend of key genes regulating the circadian rhythm, such as Per2, Per3, and Nr1d1 (Disscusion, Lines 916-939). This result is consistent with their research findings. Non etheless, we believe that the changes in oligodendrocytes in terms of metabolic regulation and axonal function homeostasis are more significant.
Pingming Qiu et al. further confirmed the correlation between the NF-κB signaling pathway in hippocampal astrocytes under METH action and neuroinflammation, neuroinjury, and learning and memory impairments in mice by integrating the GEO dataset [2]. This conclusion is also consistent with the sequencing results and analysis conclusions we obtained (Results, Lines 473-476).
In terms of the neuro-immune system disorder caused by chronic METH exposure, our research findings are consistent with those of Biao Wang et al [3]. We both observed that METH exposure may involve the participation of related immune cells (such as T cells, monocytes) and may be related to the regulation of the innate immune response and the homeostasis of myeloid cells, etc. Through the identification and analysis of cell subtypes, we further revealed that these signals may be closely related to the interaction between microglia and other immune cells mediated by MHC molecules (Disscusion, Lines 870-894).
Currently, the research results related to METH are still scattered and lack systematicness. There are differences among the research models, and there are relatively few studies on chronic exposure and in vivo experiments. Sequencing data sets with strong correlations are also scarce. We hope that this dataset can comprehensively and elaborately depict the molecular map of the hippocampus of mice after chronic METH exposure (although due to technical limitations, mature neurons die during dissociation, thus making it impossible to obtain the relevant data). In addition, we also hope to integrate the single-cell sequencing data and spatial transcriptome data of the hippocampus of mice after chronic METH exposure, providing a reliable data foundation and theoretical support for subsequent research in this field.
Finally, we would like to express our sincere gratitude for your valuable suggestions and support. Although we still need some time to further refine the manuscript based on your opinions, we sincerely hope that more readers will provide us with constructive feedback to promote the continuous improvement and deepening of this research.
(1) Oladapo A, Deshetty UM, Callen S, Buch S, Periyasamy P. Single-Cell RNA-Seq Uncovers Robust Glial Cell Transcriptional Changes in Methamphetamine-Administered Mice. Int J Mol Sci. 2025 Jan 14;26(2):649. doi: 10.3390/ijms26020649.
(2) Li K, Ling H, Wang X, Xie Q, Gu C, Luo W, Qiu P. The role of NF-κB signaling pathway in reactive astrocytes among neurodegeneration after methamphetamine exposure by integrated bioinformatics. Prog Neuropsychopharmacol Biol Psychiatry. 2024 Feb 8;129:110909. doi: 10.1016/j.pnpbp.2023.110909.
(3) Wu L, Liu X, Jiang Q, Li M, Liang M, Wang S, Wang R, Su L, Ni T, Dong N, Zhu L, Guan F, Zhu J, Zhang W, Wu M, Chen Y, Chen T, Wang B. Methamphetamine-induced impairment of memory and fleeting neuroinflammation: Profiling mRNA changes in mouse hippocampus following short-term and long-term exposure. Neuropharmacology. 2024 Dec 15;261:110175. doi: 10.1016/j.neuropharm.2024.110175.
(3) The conclusion that "prolonged METH use may progressively impair cognitive function" may not be uniformly supported by the behavioral data: Figures 1C and F (discrimination and preference indexes) exhibited that the 4-week test further declined in the METH group compared to the 2-week. In contrast, Figure 1E and H present a contradictory pattern.
Thank you very much for pointing this out. Your observation is very detailed and constructive. Regarding the conclusion "prolonged use of METH may progressively impair cognitive function", our main basis is the discrimination index and preference index shown in Figures 1C and 1F. These two indicators are usually calculated based on the total exploration time of new and old objects by mice. They are widely adopted as important references for cognitive function assessment in many relevant literature [1-3], thus providing strong support for our conclusion. The exploration frequency data we provided can, on the one hand, reflect the curiosity of mice towards new things, and on the other hand, can be calculated as the average time of each exploration by "total exploration time / exploration frequency", thereby evaluating their learning interest and the degree of their focus during exploration. We believe this is also of certain significance for reflecting the effect of METH on learning. As for the fact that there is no statistically significant difference in the exploration frequency of new and old objects in the 4-week-old mice in Figure 1H, we are also regretful about this. This might be due to the fact that our tests allow mice to freely explore in a stress-free environment, and there are significant differences among individual mice within the group. However, the mean values still show certain differences between the two groups. Compared to the mice at 2 weeks, the mice at 4 weeks have undergone a NOR test once and may have formed memories, which were retained in the subsequent assessment after four weeks. Moreover, we believe that injecting normal saline to the control group mice for a long time may affect their emotional state, because they cannot obtain the same pleasure as that brought by METH from the injection behavior.
(1) Riva M, Moriceau S, Morabito A, Dossi E, Sanchez-Bellot C, Azzam P, Navas-Olive A, Gal B, Dori F, Cid E, Ledonne F, David S, Trovero F, Bartolomucci M, Coppola E, Rebola N, Depaulis A, Rouach N, de la Prida LM, Oury F, Pierani A. Aberrant survival of hippocampal Cajal-Retzius cells leads to memory deficits, gamma rhythmopathies and susceptibility to seizures in adult mice. Nat Commun. 2023 Mar 18;14(1):1531. doi: 10.1038/s41467-023-37249-7.
(2) Lu Y, Chen X, Liu X, Shi Y, Wei Z, Feng L, Jiang Q, Ye W, Sasaki T, Fukunaga K, Ji Y, Han F, Lu YM. Endothelial TFEB signaling-mediated autophagic disturbance initiates microglial activation and cognitive dysfunction. Autophagy. 2023 Jun;19(6):1803-1820. doi: 10.1080/15548627.2022.2162244.
(3) Arroyo-García LE, Tendilla-Beltrán H, Vázquez-Roque RA, Jurado-Tapia EE, Díaz A, Aguilar-Alonso P, Brambila E, Monjaraz E, De La Cruz F, Rodríguez-Moreno A, Flores G. Amphetamine sensitization alters hippocampal neuronal morphology and memory and learning behaviors. Mol Psychiatry. 2021 Sep;26(9):4784-4794. doi: 10.1038/s41380-020-0809-2.



