Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorAlex NechiporukOregon Health and Science University, Portland, United States of America
- Senior EditorDidier StainierMax Planck Institute for Heart and Lung Research, Bad Nauheim, Germany
Reviewer #1 (Public review):
Summary:
Palardy and colleagues examine how transcription factors of the SoxB1 family alter patterning within the zebrafish posterior lateral line primordium and subsequent formation of neuromast organs along the body of the developing fish. They describe how expression of soxb genes changes when Wnt and Fgf signaling pathways are altered, and in addition, how outputs of these signalling pathways change when soxb gene expression is disrupted. Together, experiments suggest a model where the expression of SoxB genes counteracts Wnt signaling. Support comes from the combined inhibition of both pathways, partially restoring the pattern of neuromast deposition. Together, the work reveals an additional layer of control over Wnt and Fgf signals that together ensure proper posterior lateral line development.
Strengths:
The authors provide a clear analysis of changes in RNA expression after systematic manipulation of gene expression and signaling pathways to construct a plausible model of how Sox factors regulate primordium patterning.
Weaknesses:
There is little attempt to capture the variation of expression patterns with each manipulation. Photomicrographs are examples, with little quantification.
While the combined loss of soxb functions shows more severe phenotypes, it is not exactly clear what underlies the apparent redundancy. It would be helpful if the soxb gene family member expression was reported after loss of each. Expression of sox1a is shown in sox2 mutants in Figure 4, but other combinations are not reported. This additional analysis would clarify whether there are alterations in expression that influence apparent redundancy.
Reviewer #2 (Public review):
Summary:
This manuscript seeks to determine the molecular basis of tissue patterning in the collectively migrating cells of the zebrafish posterior lateral line primordium. In particular, the authors examine the cross-regulation of canonical Wnt signaling, Fgf signaling, and the SoxB1 family members Sox1a, Sox2, and Sox3 in the migrating primordium. Using a combination of mutant lines, morphino (MO) knock down, pharmacological inhibition, and dominant-negative inhibition, the authors propose a model in which Sox2 and Sox3 in the trailing region of the primordium restricts Wnt signaling to the leading region, facilitating the formation of rosettes and the deposition of the first formed neuromast downstream of Fgf pathway activity. In contrast, sox1a is expressed in the leading region of the primordium, and the sox1ay590 -/- mutant shows little phenotype on its own. Together, the authors propose a multistep signaling loop that regulates tissue patterning during lateral line collective cell migration.
Strengths:
The zebrafish posterior lateral line primordium is a well-established model for the study of collective cell migration that is useful for genetic manipulation and live imaging. The manuscript seeks to understand the complex reciprocal regulation of signaling pathways that regulate tissue patterning of collectively migrating cells.
Weaknesses:
(1) The primary tools used in this study are inadequate to support the author's conclusions.
A. The authors state that the phenotype of the sox2y589 homozygous mutant line described in this manuscript changed across generations, but do not specify which generation is used for any given experiment. The sox2y589 mutant line is not properly verified in this manuscript, which could be done by examining ant-Sox2 antibody labeling, Western blot analysis, or complementation to the existing sox2x50 line described in Gou et al., 2018a and Gou et al., 2018b. There are also published sox1a mutant lines Lekk, et al., 2019.
B. The authors acknowledge that the sox2 MO1 used in this manuscript also alters sox3 function, but do not redo the experiments with a specific sox2 MO. In addition, the authors show that the anti-Sox2 and anti-Sox3 antibody labeling is reduced but not absent in sox2 MO1 and sox3 MO-injected embryos, but do not show antibody labeling of the sox2 MO and sox3 MO-double injected embryos to determine if there is an additional knockdown.
C. The authors examine RNA in situ hybridization patterns of sox2 and sox3 following various manipulations, but do not use anti-Sox2 and anti-Sox3 antibody labeling, which would provide more quantifiable information about changes in patterning.
(2) The manuscript lacks important experimental details and appropriate quantification of results.
A. It is unclear for most of the experiments described in this manuscript how many individual embryos were examined for each experiment and how robust the results are for each condition. Only Figure 3 includes information about the numbers for each experiment, and in all cases, the experimental manipulations are not fully penetrant, and there is no statistical analysis.
B. It is not clear at what stage most of the RNA in situ hybridizations were performed.
C. The manuscript lacks quantification of many of the experiments, making it difficult to conclude their significance.
Reviewer #3 (Public review):
Summary:
This study aims to understand the molecular underpinnings of the complex process of periodic deposition of the neuromast organs of the embryonic posterior lateral line (PLL) sensory system in zebrafish. It was previously established that Fgf signaling in the trailing zone of the migrating PLL primordium is key to protoneuromast establishment, while Wnt signaling in the leading zone must be downregulated to allow new Fgf signaling-dependent protoneuromasts to form. Here, the authors evaluate the role of three SoxB transcription factors (Sox1a, Sox2, and Sox3) in this complex process, generating two novel CRISPR mutants as part of their study. They interrogate the interplay of the SoxB genes with the Fgf and Wnt signaling pathways during PLL primordium migration, using a combination of genetics, knockdown, and imaging approaches, including live time-lapse studies. They report a key role for the SoxB genes in regulating the pace of protoneuromast maturation as the primordium migrates, thus ensuring appropriate deposition and spacing of the neuromast organs.
Strengths:
Strengths of the study are the careful quantitative analysis. based on imaging approaches, of the impact of mutation or knockdown of SoxB genes, coupled with the use of heat shock inducible dominant negative strategies to address how SoxB genes interact with Wnt and Fgf signaling. Functional analyses convincingly uncover a SoxB regulatory network that serves to limit Wnt activity, as directly read out with a live Wnt reporter. The finding that Wnt inhibition (achieved using pharmacological reagents) rescues the SoxB deficiency phenotype provides compelling evidence of the centrality of the Wnt pathway in mediating SoxB function. Use of atoh1 markers to track the stages of development of the neuromasts provides an effective approach to following their maturation, and allows the authors to explore how SoxB/Wnt interplay ultimately translates into the establishment of functional neuromasts. Finally, loss of Sox2 function, together with loss of either Sox1a or Sox3, blocks maturation of the neuromasts, clearly establishing redundancy between these SoxB family genes.
The concepts introduced and explored in this study - of complex gene networks that work within a dynamic cellular environment to enable self-organization and ultimately stabilization of cell fate choices-provide a useful conceptual framework for future studies. This study is therefore of relevance to understanding the morphogenesis of self-organizing tissues more broadly.
Weaknesses:
A minor weakness is the use of SoxB morpholino (MO) knockdown reagents, which are interspersed with mutant analyses. Although the stable mutants are available, they would be challenging to couple with the reporter transgenes used for some of the experiments, providing a reasonable rationale for the use of MO reagents (although the authors don't overtly provide this rationale). Moreover, reduced penetrance of the Sox2 mutants over multiple generations is noted, but no detailed explanation for this finding is offered.
Given that the expression patterns of Sox1a and Sox3 are not merely different but are largely reciprocal, the mechanistic basis of their very similar double mutant phenotypes with Sox2 remains opaque. Related to this, the authors discuss that Sox1a/Sox2 double knockdown produces a more severe phenotype than Sox2/Sox3 double knockdown, yet this difference is not obviously reflected in the data, some of which is not shown.
Author response:
We would like to thank the three reviewers for the careful review and thoughtful comments on our manuscript. In addition to providing useful suggestions, they uncovered some embarrassing oversights on our part, related to experimental details including number of embryos, and quantification of variance in the observed changes for some of the experiments, which were inadvertently omitted in the submission. We provide below an initial response to the reviewer’s public reviews and expect to submit a revised manuscript comprehensively addressing all their concerns.
I would like to start by addressing some of their most critical comments related to validation of the tools used to reduce soxB1 gene family function in the embryo. In the absence of the critical supplementary data that we inadvertently failed to include, the reviewers were left with an understandable, but we feel erroneous impression, that there was insufficient validation of mutant and knockdown tools.
Reviewer #2 says “The sox2y589 mutant line is not properly verified in this manuscript, which could be done by examining ant-Sox2 antibody labeling, Western blot analysis or…”
This validation, which had been performed previously both with antibody staining and with western blot analysis, was inadvertently omitted from the supplementary data submitted with the paper. The western blot data is shown here.
Author response image 1.
Validation of sox2 mutant phenotype with Western blot.
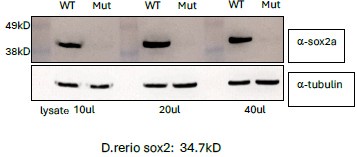
Lysates were prepared from 25 embryos selected as wild type or potentially mutant based on the “loss of L1” phenotype at 6 dpf. This polyclonal antibody recognizes within the last 16 amino acids of the C-terminal.
Author response image 2.
Validation of sox2 mutant phenotype with antibody staining.
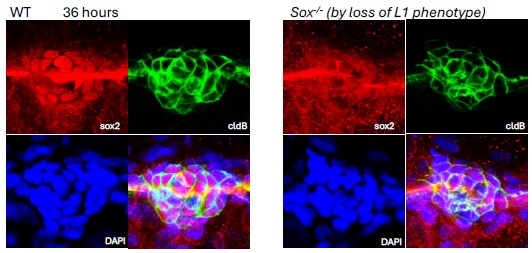
Though in this experiment there was considerable background in the red channel, and it shows the lateral line nerve, loss of nuclear Sox2 expression is evident in the deposited neuromast of an embryo identified as a mutant based on its delayed deposition of the L1 neuromast.
This data and a repeat of the antibody staining showing the primordium with loss of Sox2 will be included in a revised manuscript.
Furthermore, Reviewer #2 comments “the authors show that the anti-Sox2 and antiSox3 antibody labeling is reduced but not absent in sox2 MO1 and sox3 MO-injected embryos, but do not show antibody labeling of the sox2 MO and sox3 MO-double injected embryos to determine if there is an additional knockdown”
This will be included in a revised manuscript.
Reviewer #2:
The authors acknowledge that the sox2 MO1 used in this manuscript also alters sox3 function, but do not redo the experiments with a specific sox2 MO
This is not exactly true. Having discovered sox2 MO1 simultaneously reduces sox2 and sox3 function, three new morpholinos were obtained based on another paper (Kamachi et al 2008), which had quantitatively assessed efficacy of three sox2 specific morpholinos (sox2 MO2, sox2 MO3, and sox2 MO4). The effects of these morpholinos on the pattern of L1 deposition was compared to that of sox2 MO1. This comparison was shown in supplementary Figure 2 and is included below. It shows that the sox2 specific morpholinos resulted in a poorly penetrant delay in deposition of L1, comparable to that of a sox2 mutant, which was quantified in supplementary Figure 3B. The observations with these three sox2 specific morpholinos independently supported the observations made with the sox2 mutant that reduction of sox2 on its own results in a delay in deposition of the first neuromast with low penetrance and that to effectively examine the role of these SoxB1 genes in the primordium their function needs to be compromised in a combinatorial manner. A conclusion that was independently supported by observations made by crossing sox1a, sox2 and sox3 mutants (Figure 3 and Supplementary Figure 3). Therefore, even though the initial use of a sox2 morpholino, which simultaneously knocks down sox3, was unintentional, its use turned out to be useful. It allowed us to examine effects of knocking down sox2 and sox3 with a single morpholino. Furthermore, though this project was initiated more than 15 years ago to specifically understand sox2 function, our focus had shifted to understanding the role of soxB1 family members sox1a, sox2 and sox3 functioning together as an interacting system that regulates Wnt activity in the primordium. Considering this broader focus, reflected in the title of the paper, it was not a priority to repeat every experiment previously done with the sox2MO1 with the new sox2 specific morpholinos. Instead, having acknowledged the “limitations” of sox2MO1, we used it to better understand effects of combinatorial reduction of SoxB1 function.
Reviewer #1:
It is not exactly clear what underlies the apparent redundancy. It would be helpful if the soxb gene family member expression was reported after loss of each.
As suggested by reviewer #1, we had previously looked changes in expression of each of the soxB1 factors following loss of individual soxB1 factors but not included it in the supplementary data with the original submission. Independent of a reproducible and consistent expansion sox1a expression into the trailing zone, following loss of sox2 function, which is reported in the paper and quantified here where 10/10 mutant embryos showed the expansion (compare region within bracket in WT and sox2-/-), no consistent changes in the expression of other soxB1 family members was observed as part of a mechanism that might account for compensation when function of a particular soxB1 factor is soxB1 factor is lost. The data shown above together with more extensive quantification of changes will be included in a revised version of the manuscript. At this time the only consistent change was the expansion of sox1a to the trailing zone when lost. The data trailing zone when sox2 function is lost. This change reflects dependence of sox1a on Wnt activity and the fact that Wnt activity expands into the trailing zone when sox2 function is lost.
Author response image 3.
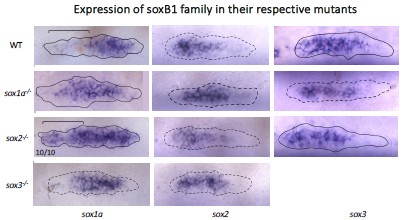
Reviewer #3:
Given that the expression patterns of Sox1a and Sox3 are not merely different but are largely reciprocal, the mechanistic basis of their very similar double mutant phenotypes with Sox2 remains opaque.
The simplest way to think about compensation for gene function in a network is to think of it being determined by expression of a homolog or another gene with a similar function being expressed in a similar or overlapping domain. However, it is more useful to think of Sox2 function in the primordium as part of a interacting network of SoxB1 factors whose differential regulatory mechanisms create a robust system that simultaneously regulates two key aspects of Wnt activity in the primordium; how high Wnt activity is allowed to get in the leading zone and how effectively it is shut off to facilitate protoneuromast maturation in the trailing zone. These features of Wnt activity influence both when and where nascent protoneuromasts will form in the wake of a progressively shrinking Wnt system and where they undergo effective maturation and stabilization prior to deposition. Changes in individual SoxB1 expression patterns provide some hints about how some SoxB1 factors may compensate when function of one or more of these factors is compromised. However, a deeper understanding of robustness and “compensation” will require a systems level understanding of this gene regulatory network with computational models, which we are currently working on in our group. It remains possible, for example, that how far into the trailing zone the Wnt activity has an influence is regulated at least in part by how high it is allowed to get in the leading zone by sox1a. Conversely, how high Wnt activity gets in the leading zone may be influenced by how effectively it is shut off in the trailing zone by sox2 and sox3, as this influences the size of the Wnt system, which in turn can influence the overall level of Wnt activity. In this manner Sox1a may cooperate with Sox2 and Sox3 to limit both how high Wnt activity is allowed to get in the primordium and to effectively shut it off in the trailing zone.
Reviewer #3:
Related to this, the authors discuss that Sox1a/Sox2 double knockdown produces a more severe phenotype than Sox2/Sox3 double knockdown, yet this difference is not obviously reflected in the data.
The severity of the sox1a/sox2 double mutant phenotype compared to that of the sox2/sox3 double mutant is shown in Figure 3 K and N, and quantified in Supplementary Figure 3A. Simultaneous loss of sox2 and sox3 results in a small but relatively penetrant delay in where the first stable neuromast is deposited (Figure 2 N). By contrast, loss of sox2 and sox1a together consistently results in a longer delay in deposition of the first stable (Figure 2 K). A new graph, shown below, which will be incorporated in the revised paper, shows that there is a significant difference in the pattern of L1 deposition in sox1a-/-, sox2-/- and sox2-/-, sox3-/- double mutants.
Author response image 4.
All 3 datasets found to be normally distributed by Shapiro-Wilk test. 1-way ANOVA showed significance (<0.0001), with Tukey’s multiple comparisons test showing significant difference between all 3 conditions. (***p=0.0008, ****p<0.0001)
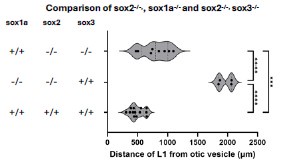
Reviewer #1:
It would be good to more clearly state why sox3 is not regulated by Wnt given its expression is inhibited by the delta TCF construct (Figure 2M).
The explanation for why we believe sox3 expression is determined by Fgf signaling, and not Wnt activity requires integrating what is observed both with induction of the delta TCF construct and the dominant negative Fgf receptor (DN FgfR). Loss of sox3 expression with induced expression of the delta TCF construct could result from loss of Wnt activity or the downstream loss of Fgf activity, which is ultimately dependent on Fgfs secreted by Wnt active cells in the leading domain. Distinguishing between these possibilities is based on inhibition of FGF signaling with the DN FgfR, described in the next paragraph. Heat Shock induced expression of DN FgfR expression results in loss of FGF signaling and the simultaneous expansion of Wnt activity into the trailing zone. As explained in the original text, loss of sox3 expression in this context, rather than its expansion, suggests its expression is determined by Fgf signaling not Wnt activity. We will emphasize that its loss, rather than its expansion, following induction of DN FgfR, indicates its expression is determined by Fgf signaling not Wnt activity.
Reviewer #2:
The manuscript lacks quantification of many of the experiments, making it difficult to conclude their significance.
One of the biggest inadvertent omissions of the paper was the inadequate quantification of some of the results. Quantification of results with considerable variation in the outcome, like the pattern of L1 deposition, was provided following manipulations where various combinations of sox1a, sox2, and sox3 function was lost (Figures 3, supplementary Figures 2 and 3) or where sox2MO1/sox3MO was used with or without IWR (Figure 5 and Figure 6). However, numbers for the experiments in Figures 2 were omitted in the Figure legend, where typically about 10 embryos for each manipulation were photographed, scored, and a representative image was used to make the figure. In these experiments there was a very consistent result with 100% of the embryos showing changes represented by each panel in Figure 2. The only exception was Figure 2Y where 9/10 embryos showed the described change. Similarly in Figure 4 there was a consistent result and 100% of embryos showed the change shown. Numbers and statistics for these results will be included in a revised manuscript.
Reviewer #2:
The statistical analysis in Figure 5 and Supplementary Figures 2 and 3 should be one-way ANOVA or Kruskal-Wallis with a Dunn's multiple comparisons test rather than pair-wise comparisons.
The analysis has been re-done following the reviewer’s suggestions. The analysis confirms the primary conclusions of the original submission, and this analysis will be incorporated in a revised manuscript. However, to improve the power of the analysis, experiments with low numbers of embryos will be repeated.
See redone graphs in Figure 5 and supplementary Figure 2 and 3.



