Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorZhao ShanYunnan University, Kunming, China
- Senior EditorDidier StainierMax Planck Institute for Heart and Lung Research, Bad Nauheim, Germany
Reviewer #1 (Public review):
Summary:
In this manuscript, Chengjian Zhao et al. focused on the interactions between vascular, biliary, and neural networks in the liver microenvironment, addressing the critical bottleneck that the lack of high-resolution 3D visualization has hindered understanding of these interactions in liver disease.
Strengths:
This study developed a high-resolution multiplex 3D imaging method that integrates multicolor metallic compound nanoparticle (MCNP) perfusion with optimized CUBIC tissue clearing. This method enables the simultaneous 3D visualization of spatial networks of the portal vein, hepatic artery, bile ducts, and central vein in the mouse liver. The authors reported a perivascular structure termed the Periportal Lamellar Complex (PLC), which is identified along the portal vein axis. This study clarifies that the PLC comprises CD34+Sca-1+ dual-positive endothelial cells with a distinct gene expression profile, and reveals its colocalization with terminal bile duct branches and sympathetic nerve fibers under physiological conditions.
Comments on revisions:
The authors very nicely addressed all concerns from this reviewer. There are no further concerns and comments.
Reviewer #3 (Public review):
Xu, Cao and colleagues aimed to overcome the obstacles of high-resolution imaging of intact liver tissue. They report successful modification of the existing CUBIC protocol into Liver-CUBIC, a high-resolution multiplex 3D imaging method that integrates multicolor metallic compound nanoparticle (MCNP) perfusion with optimized liver tissue clearing, significantly reducing clearing time and enabling simultaneous 3D visualization of the portal vein, hepatic artery, bile ducts, and central vein spatial networks in the mouse liver. Using this novel platform, the researchers describe a previously unrecognized perivascular structure they termed Periportal Lamellar Complex (PLC), regularly distributed along the adult liver portal veins.
Using available scRNAseq data, the authors assessed the CD34+Sca-1+ cells' expression profile, highlighting mRNA presence of genes linked to neurodevelopment, bile acid transport, and hematopoietic niche potential. Different aspects of this analysis were then addressed by protein staining of selected marker proteins in the mouse liver tissue. Next, the authors addressed how the PLC and biliary system react to CCL4-induced liver fibrosis, implying PLC dynamically extends, acting as a scaffold that guides the migration and expansion of terminal bile ducts and sympathetic nerve fibers into the hepatic parenchyma upon injury.
The work clearly demonstrates the usefulness of the Liver-CUBIC technique and the improvement of both resolution and complexity of the information, gained by simultaneous visualization of multiple vascular and biliary systems of the liver. The identification of PLC and the interpretation of its function represent an intriguing set of observations that will surely attract the attention of liver biologists as well as hepatologists. The importance of the CD34+/Sca1+ endothelial cell population and claims based on transcriptomic re-analysis require future assessment by functional experimental approaches to decipher the functional molecules involved in PLC formation, maintenance, and the involvement in injury response before establishing their role in biliary, arterial, and neural liver systems.
Strengths:
The authors clearly demonstrate an improved technique tailored to the visualization of the liver vasulo-biliary architecture in unprecedented resolution.
This work proposes a new morphological feature of adult liver facilitating interaction between the portal vein, hepatic arteries, biliary tree, and intrahepatic innervation, centered at previously underappreciated protrusions of the portal veins - PLCs.
Weaknesses:
The importance of CD34+Sca1+ endothelial cell sub-population for PLC formation and function was not tested and warrants further validation.
Comments on revisions:
I appreciate the author's effort to revise the text so it more rigorously adheres to the presented evidence. Following a thorough read of the revised text, a few remaining minor issues were identified in the Discussion.
(1) From where comes the hard evidence for PLC being the stem cell niche in the following sentence?
for the two following statements:
This suggests that the PLC may not only provide structural support but also serve as a perivascular stem cell niche specific to the portal region, potentially involved in hematopoiesis and tissue regeneration.
The PLC serves as a directional scaffold for ductal growth, a specialized stem cell niche, and a potential site of neurovascular coupling.
(2) In the following paragraph, I lack references to the previously published evidence of liver innervation guidance mechanisms, such as the mesenchyme-mediated guidance (CD31- population) Gannoun et al., 2023 https://doi.org/10.1242/dev.201642, an important context for your finding.
Further analysis showed significant upregulation of genes involved in neurodevelopment and axonal guidance in the CD34+Sca-1+ cluster, along with activation of neuronal signaling pathways. Immunostaining confirmed the presence of TH+ sympathetic nerve fibers wrapping around the PLC in a "beads-on-a-string" pattern (Fig. 6), consistent with a classic neurovascular unit(Adori et al., 2021). Previous studies have shown that sympathetic nerves enter the liver along collagen fibers of Glisson's capsule and interact with hepatic arteries, portal veins, and bile duct epithelium, supporting the PLC as a scaffold for intrahepatic neurovascular integration.
(3) Several sentences have issues with a lack of space between words.
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this manuscript, Chengjian Zhao et al. focused on the interactions between vascular, biliary, and neural networks in the liver microenvironment, addressing the critical bottleneck that the lack of high-resolution 3D visualization has hindered understanding of these interactions in liver disease.
Strengths:
This study developed a high-resolution multiplex 3D imaging method that integrates multicolor metallic compound nanoparticle (MCNP) perfusion with optimized CUBIC tissue clearing. This method enables the simultaneous 3D visualization of spatial networks of the portal vein, hepatic artery, bile ducts, and central vein in the mouse liver. The authors reported a perivascular structure termed the Periportal Lamellar Complex (PLC), which is identified along the portal vein axis. This study clarifies that the PLC comprises CD34+Sca-1+ dual-positive endothelial cells with a distinct gene expression profile, and reveals its colocalization with terminal bile duct branches and sympathetic nerve fibers under physiological conditions.
Comments on revisions:
The authors very nicely addressed all concerns from this reviewer. There are no further concerns or comments.
We sincerely thank the reviewer for the positive evaluation of the revised manuscript.
Reviewer #2 (Public review):
Summary:
The present manuscript of Xu et al. reports a novel clearing and imaging method focusing on the liver. The Authors simultaneously visualized the portal vein, hepatic artery, central vein, and bile duct systems by injected metal compound nanoparticles (MCNPs) with different colors into the portal vein, heart left ventricle, vena cava inferior and the extrahepatic bile duct, respectively. The method involves: trans-cardiac perfusion with 4% PFA, the injection of MCNPs with different colors, clearing with the modified CUBIC method, cutting 200 micrometer thick slices by vibratome, and then microscopic imaging. The Authors also perform various immunostaining (DAB or TSA signal amplification methods) on the tissue slices from MCNP-perfused tissue blocks. With the application of this methodical approach, the Authors report dense and very fine vascular branches along the portal vein. The authors name them as 'periportal lamellar complex (PLC)' and report that PLC fine branches are directly connected to the sinusoids. The authors also claim that these structures co-localize with terminal bile duct branches and sympathetic nerve fibers and contain endothelial cells with a distinct gene expression profile. Finally, the authors claim that PLC-s proliferate in liver fibrosis (CCl4 model) and act as scaffold for proliferating bile ducts in ductular reaction and for ectopic parenchymal sympathetic nerve sprouting.
Strengths:
The simultaneous visualization of different hepatic vascular compartments and their combination with immunostaining is a potentially interesting novel methodological approach.
Weaknesses:
This reviewer has some concerns about the validity of the microscopic/morphological findings as well as the transcriptomics results, and suggests that the conclusions of the paper may be critically viewed. Namely, at this point, it is still not fully clear that the 'periportal lamellar complex (PLC)' that the Authors describe really exists as a distinct anatomical or functional unit or these are fine portal branches that connect the larger portal veins into the adjacent sinusoid. Also, in my opinion, to identify the molecular characteristics of such small and spatially highly organized structures like those fine radial portal branches, the only way is to perform high-resolution spatial transcriptomics (instead of data mining in existing liver single cell database and performing Venn diagram intersection analysis in hepatic endothelial subpopulations). Yet, the existence of such structures with a distinct molecular profile cannot be excluded. Further research with advanced imaging and omics techniques (such as high resolution volume imaging, and spatial transcriptomics/proteomics) are needed to reproduce these initial findings.
We thank the reviewer for the thoughtful and constructive comments. In response to the reviewer’s concerns regarding the anatomical and molecular definition of the periportal lamellar complex (PLC), we have further clarified the scope and methodological boundaries of the present study in the revised manuscript.
Regarding the key question raised by the reviewer—namely, whether the PLC represents an independent anatomical or functional unit, or merely small portal venous branches connecting larger portal veins to adjacent sinusoids—we provide below a more detailed explanation of the criteria used to define the PLC in this study. The identification of the PLC is primarily based on periportal structures that can be reproducibly recognized by three-dimensional imaging across multiple mice, exhibiting a relatively consistent spatial distribution within the periportal region. The PLC could be stably observed across different MCNP dye color assignments and independent experimental batches. In addition, three-dimensional CD31 immunofluorescence consistently revealed vascular-associated signal distributions in the same periportal region, indirectly supporting its spatial association with the periportal vascular system.
At the morphological level, the PLC appears as a periportal vasculature-associated structure distributed around the main portal vein trunk and maintains a relatively consistent spatial proximity to portal veins, bile ducts, and neural components in three-dimensional space. This highly conserved spatial organization across multiple tissue systems supports the anatomical positioning of the PLC as a relatively distinct structural tissue unit within the periportal region.
The present study primarily focuses on a descriptive characterization of the three-dimensional anatomical organization and spatial relationships of the PLC based on volumetric imaging and vascular labeling strategies. As a complementary exploratory analysis, we reanalyzed endothelial cell populations potentially associated with the PLC using existing liver single-cell transcriptomic datasets. This analysis was intended to provide molecular-level information consistent with the structural observations and to offer preliminary clues to its potential biological functions, rather than to independently define the PLC at the spatial level or to functionally validate it.
We fully acknowledge the value of spatial transcriptomic and spatial proteomic technologies in revealing molecular heterogeneity within tissue architecture. However, under current technical conditions, these approaches are largely dependent on thin tissue sections and are limited by spatial resolution and signal mixing effects, which still pose challenges for resolving periportal structures with pronounced three-dimensional continuity, such as the PLC. In the future, further integration of high-resolution volumetric imaging with spatial omics technologies may enable a more refined understanding of the molecular features and potential functions of the PLC at higher spatial resolution.
Reviewer #3 (Public review):
Summary:
In the revised version of the manuscript authors addressed multiple comments, clarifying especially the methodological part of their work and PLC identification as a novel morphological feature of the adult liver portal veins. Tet is now also much clearer and has better flow.
The additional assessment of the smartSeq2 data from Pietilä et al., 2025 strengthens the transcriptomic profiling of the CD34+Sca1+ cells and the discussion of the possible implications for the liver homeostasis and injury response. Why it may suffer from similar bias as other scRNA seq datasets - multiple cell fate signatures arising from mRNA contamination from proximal cells during dissociation, it is less likely that this would happen to yield so similar results.
Nevertheless, a more thorough assessment by functional experimental approaches is needed to decipher the functional molecules and definite protein markers before establishing the PLC as the key hub governing the activity of biliary, arterial, and neuronal liver systems.
The work does bring a clear new insight into the liver structure and functional units and greatly improves the methodological toolbox to study it even further, and thus fully deserves the attention of the Elife readers.
Strengths:
The authors clearly demonstrate an improved technique tailored to the visualization of the liver vasulo-biliary architecture in unprecedented resolution.
This work proposes a new morphological feature of adult liver facilitating interaction between the portal vein, hepatic arteries, biliary tree, and intrahepatic innervation, centered at previously underappreciated protrusions of the portal veins - the Periportal Lamellar Complexes (PLCs).
Weaknesses:
The importance of CD34+Sca1+ endothelial cell subpopulation for PLC formation and function was not tested and warrants further validation.
We thank the reviewer for the careful and constructive comments regarding the functional validation of cell populations associated with the PLC. The central aim of this study is to establish and validate a novel volumetric imaging and vascular labeling strategy and to apply it to the periportal region of the liver, thereby revealing previously underappreciated structural organizational patterns at the three-dimensional level, rather than to perform a systematic functional validation of specific cellular subpopulations.
We agree that the precise roles of the CD34+Sca-1+ endothelial cell subpopulation in the formation and function of the periportal lamellar complex (PLC) have not been directly addressed through functional intervention experiments in the present study. Our conclusions are primarily based on three-dimensional imaging and spatial distribution analyses, which reveal a stable and consistent spatial association between this cell population and the PLC structure, but are not intended to independently support causal or functional inferences. The underlying functional mechanisms remain to be elucidated in future studies using genetic or functional perturbation approaches.
In light of these considerations, we have further refined the relevant statements in the revised manuscript to more clearly define the functional scope and limitations of the current study in the Discussion section, and to avoid functional interpretations that extend beyond the direct support of the data. At the same time, we consider functional validation of the PLC to be an important and promising direction for future investigation.
It should be emphasized that the present study is not primarily designed to provide direct functional validation, but rather to systematically characterize the three-dimensional structural features of the periportal lamellar complex (PLC) and its cellular associations using volumetric imaging and vascular labeling approaches. At this stage, we mainly provide spatial and histological evidence for the organizational relationship between the PLC structure and the CD34+Sca-1+ endothelial cell population, while their specific roles in PLC formation and functional regulation await further investigation.
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
I highly appreciate the Authors' endeavors to improve the manuscript. I am enlisting those points (from my original review) where I still have further comments.
(2) I would suggest this sentence:
"...the liver has evolved a highly complex and densely organized ductal vascular-neuronal network in the body, consisting primarily of the portal vein system, central vein system, hepatic artery system, biliary system, and intrahepatic autonomic nerve network [6, 7]."
We thank the reviewer for the valuable suggestion. We have revised the relevant sentence accordingly, and the revised wording is as follows:
“The liver has evolved a highly complex and densely organized vascular–biliary–neural network, primarily composed of the portal venous system, central venous system, hepatic arterial system, biliary system, and the intrahepatic autonomic neural network.”
(3) I suggest renaming 'clearing efficiency' to 'clearing time', and revise the last sentence like:
'...The results showed that the average transmittance increased by 20.12% in 1mm-thick cleared tissue slices.'
We thank the reviewer for this helpful suggestion. Accordingly, we have replaced the term “clearing efficiency” with “clearing time” and revised the final sentence to reflect this change. The revised wording is as follows:
“The results showed that the average transmittance increased by 20.12% in cleared tissue slices with a thickness of 1 mm.”
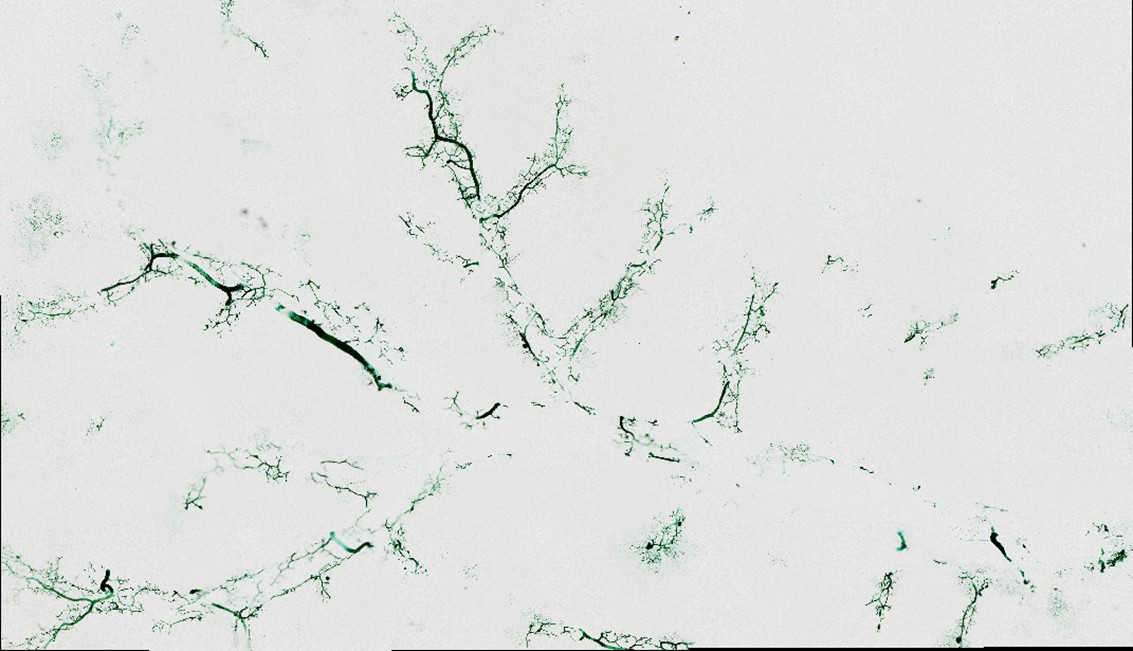
(4) While the dye perfusion was indeed on full lobe, FigS1F also seems to be rather a thick section instead of a full 3d reconstruction. This is OK, but please, be clear and specific about this in the respective part of the ms.
We thank the reviewer for the careful review and detailed comments. We would like to clarify that Fig. S1F shows whole-lobe imaging of the mouse left liver lobe obtained after dye perfusion at the whole-liver scale, rather than an image derived from a thick tissue section. Although this image does not represent a three-dimensional reconstruction, it does reflect imaging of the entire left liver lobe at the macroscopic level.
In addition, for the reviewer’s reference, we have provided in this response a representative image of a 200 μm-thick liver tissue section to directly illustrate the morphological differences between thick-section imaging and whole-lobe imaging. We note that the third and fourth panels in Fig. 1G of the main text already show local imaging results from 200 μm-thick sections; in contrast, the comparative image provided here presents a larger field of view and overall morphology. To avoid redundancy, this additional image is included solely for clarification in the present response and has not been incorporated into the revised manuscript or the supplementary materials.
(11) Regarding the 'transmission quantification':
'Regarding the comparative quantification of different clearing methods, as the reviewer noted, nearly all aqueous or organic solvent based clearing techniques can achieve relatively uniform transparency in 1 mm thick tissue sections, so differences at this thickness are limited.'
So, based on all these, I think, measuring/comparisons of clearing efficacy in the present form are kind of pointless --- one may consider omitting this part.
We thank the reviewer for the valuable comments. The purpose of the transmittance quantification in this study was not to provide a comprehensive comparison among different tissue-clearing methods, but rather to serve as a quantitative reference supporting the optimization of the Liver-CUBIC protocol. Accordingly, we have narrowed and clarified the relevant statements in the revised manuscript to define their scope and avoid overinterpretation.
The revised text now reads as follows:
“Importantly, Liver-CUBIC treatment did not induce significant tissue expansion (Figure 1B–D). In addition, quantitative transmittance measurements in 1-mm-thick cleared tissue slices showed an average increase of 20.12% (P < 0.0001; 95% CI: 19.14–21.09; Figure 1E).”
Author response image 1.
(16) It is OK, but please, indicate this clearly in the Methods/Results because in its present form it may be confusing for the reader: which color means what.
We thank the reviewer for this helpful request for clarification. We agree that the previous wording may have caused confusion regarding the meaning of different MCNP colors. Accordingly, we have revised the Methods section and the relevant figure legends to clearly state that the color assignment of MCNP dyes is not fixed across different experiments or figures. The use of different colors serves solely for visualization and presentation purposes, facilitating the distinction of anatomical structures in multichannel and three-dimensional imaging, and does not indicate any fixed or intrinsic correspondence between a specific color and a particular vascular or ductal system. We believe that this clarification will help prevent misinterpretation and improve the overall clarity of the manuscript.
(17) Still I think the hepatic artery is extremely shrunk, while the portal vein is extremely dilated. Please, note that in the referring figure (from Adori et al), hepatic artery and portal vein are ca 50 micrometers and 250 micrometers in diameter, respectively. In your figure, as I see, ca. 9-10 micrometers and 125 micrometers, respectively. This means 5x (Adori) vs. 13-14x differences (you). I would not say that this is necessarily problematic --- but may reflect some perfusion issues that may be good to consider.
We thank the reviewer for the careful comparison and acknowledge the quantitative differences pointed out. Compared with the study by Adori et al., the diameter ratio between the hepatic artery and the portal vein in our images does indeed differ to some extent. We believe that this discrepancy primarily arises from methodological differences in imaging and analysis strategies between the two studies.
In the work by Adori et al., periportal vasculature identification and three-dimensional segmentation were mainly based on 488 nm autofluorescence signals acquired from inverted tissues. This signal predominantly reflects the overall outline of periportal tissue regions rather than direct imaging of the vascular lumen itself. Consequently, the measured “vessel diameter” largely represents a spatial domain delineated by surrounding periportal structures, and does not necessarily correspond to the actual or functional luminal diameter of the vessel.
In contrast, the present study employed fluorescent MCNP dye perfusion under low perfusion pressure, combined with tissue clearing and three-dimensional optical imaging. Under these experimental conditions, the measured vessel diameters more closely reflect the perfusable luminal space of vessels in a fixed state, rather than their maximally dilated diameter, and are not defined by the morphology of surrounding tissues. This distinction is particularly relevant for the hepatic artery: as a high-resistance, smooth muscle–rich vessel, its diameter is highly sensitive to perfusion pressure and post-excision changes in vascular tone. In comparison, the portal vein exhibits greater compliance and is relatively less affected by these factors.
Based on these methodological differences, the observation of relatively smaller apparent hepatic arterial diameters—and consequently a higher arterial-to-portal vein diameter ratio—under dye perfusion–based optical imaging conditions is an expected outcome. Importantly, the primary focus of the present study is the identification and characterization of the periportal lamellar complex (PLC) as a three-dimensional lamellar tissue structure that can be stably and reproducibly recognized across different samples and imaging conditions, rather than absolute comparisons of vascular diameters.
(21) After the presented documentation, I still have some concerns that the 'periportal lamellar complex (PLC)' that the Authors describe is really a distinct anatomical or functional unit. The confocal panel in Fig. 4F is nice and high quality. However, as far as I see, it shows that CD34+/Sca-1+ immunostaining is not specific for the presumptive PLCs in the peri-portal region. Instead, Sca-1 immunoreactivity is highly abundant also in the midzone --- to which the supposed PLCs do not extend, according to the cartoon shown in panel D, same figure. Notably, this questions also the specificity of the single cell analysis.
We thank the reviewer for this detailed and important comment regarding the specificity of CD34+/Sca-1+ markers and the definition of the periportal lamellar complex (PLC).
It should be emphasized that the PLC is not defined on the basis of any single molecular marker, but rather by a reproducible periportal lamellar anatomical structure consistently revealed by three-dimensional imaging across multiple samples. The co-expression of CD34 and Sca-1 is interpreted within this clearly defined anatomical context and is used to characterize the molecular features of endothelial cells associated with the PLC structure.
As shown in Fig. 4F, the co-expression of CD34 and Sca-1 delineates a continuous, lamellar endothelial structure surrounding the portal vein. In contrast, outside the periportal region—including the midlobular areas—Sca-1 or CD34 expression can also be detected, but these signals appear scattered and discontinuous, lacking an organized lamellar topology.
In the single-cell transcriptomic analysis, we treated CD34+/Sca-1+ endothelial cells as an operational population to explore molecular features that may be enriched in the microenvironment of the periportal lamellar complex (PLC). Importantly, this analysis was intended to provide molecular clues associated with the PLC, rather than to precisely assign spatial locations or identities to individual cells.
Occasional isolated Sca-1+ signals detected outside the periportal region do not affect the anatomical definition of the PLC, nor do they alter the interpretation of the single-cell analysis. These analyses serve to provide supportive and exploratory molecular information for the structural identification of the PLC, rather than constituting decisive spatial evidence.
(23) '....In the manuscript, we have carefully stated that this analysis is exploratory in nature and have avoided overinterpretation. In future studies, high-resolution spatial omics approaches will be invaluable for more precisely delineating the molecular characteristics of these fine structures.'
I do not find these statements either in the Discussion or in the Results. I must reiterate my opinion that the applied methodical approach in the single cell transcriptomics part has severe limitations, and the readers must be aware of this.
We thank the reviewer for this further comment. We understand and acknowledge the reviewer’s concerns regarding the methodological limitations of single-cell transcriptomic analyses, and we agree that these limitations should be clearly communicated to readers in the main text.
We acknowledge that in the previous version of the manuscript, the exploratory nature of the single-cell transcriptomic analysis and its methodological boundaries were discussed only in the response to reviewers and were not explicitly stated in the manuscript itself. We thank the reviewer for pointing out this omission. In the revised manuscript, we have now added explicit clarifications in the main text to prevent potential overinterpretation of these results.
In the present study, our primary effort is focused on the descriptive characterization of the three-dimensional anatomical organization and spatial relationships of the PLC using volumetric imaging and vascular labeling strategies. As a complementary exploratory analysis, we reanalyzed existing liver single-cell transcriptomic datasets to examine endothelial cell populations exhibiting PLC-associated features, and performed differential gene expression and Gene Ontology enrichment analyses. Importantly, these results are intended to provide molecular-level support for the structural identification of the PLC and to offer preliminary insights into its potential biological functions. Accordingly, we have narrowed the presentation and interpretation of the single-cell analysis in both the Results and Discussion sections of the revised manuscript.
In addition, we have expanded the Discussion to address the limitations of current spatial transcriptomic approaches in validating a continuous three-dimensional structure such as the PLC. Most existing spatial transcriptomic methods rely on two-dimensional tissue sections of 8–10 μm thickness, whereas identification of the PLC depends on three-dimensional imaging of tissue volumes with thicknesses of ≥200 μm, making reliable reconstruction of its spatial continuity from single sections challenging. Furthermore, because each spatial transcriptomic capture spot often encompasses multiple adjacent cells, signal mixing effects further limit precise resolution of specific periportal microstructures.
Overall, we agree with the reviewer’s central point that the limitations of single-cell transcriptomic analyses should be clearly understood by readers. By explicitly clarifying the methodological boundaries and refining the related statements in the main text, we believe this concern has now been adequately addressed in the revised manuscript. We thank the reviewer for identifying this omission, which has helped to improve the rigor and clarity of the study.
Reviewer #3 (Recommendations for the authors):
(1) While interesting observations, suitable for discussion, the following sections are speculations, given that no functional characterization of PLC importance has been performed yet. This is the most felt when commenting on the role in hematopoiesis, which transiently takes place in the liver during embryogenesis (Khan et al 2016) but ceases to exist after ligation of the umbilical inlet. Adult Liver hematopoiesis remains controversial, and more solid evidence would need to be presented to support its existence in PLC regions.
265 - These findings suggest that the Periportal Lamellar Complex (PLC) is not only a morphologically and spatially distinct, low-permeability vascular unit surrounding the portal vein, but also likely serves as a critical nexus connecting the portal vein, hepatic artery, and liver sinusoids. Thus, the PLC constitutes a key node within the interactive vascular network of the mouse liver.
We thank the reviewer for the comments and suggestions regarding the potential functional interpretation of the periportal lamellar complex (PLC), particularly its possible association with hematopoietic function. We would like to clarify that the statement on page 265 was intended solely to describe the structural characteristics and spatial organization of the PLC within the periportal vascular network. Specifically, the original wording aimed to summarize the morphological features of the PLC and its spatial relationships among the portal vein, hepatic artery, and hepatic sinusoids.
Nevertheless, to minimize potential misunderstanding, we have revised this section to avoid unnecessary functional implications. The revised text now reads:
“These results suggest that the periportal lamellar complex (PLC) is a morphologically and spatially distinct vascular structure that surrounds the portal vein and may serve as a key organizational node coordinating the spatial relationships among the portal vein, hepatic artery, and hepatic sinusoids. Accordingly, the PLC represents an important structural element within the interactive vascular network of the mouse liver.”
This revision preserves the structural significance of the PLC while avoiding overinterpretation of its functional roles.
(2) The same is true also for this section, following Figure 3 - no functional experiment tested this. For example, diphtheria toxin is expressed in the CD34+Sca1+ population. Or at least a careful mapping of the developing liver, which would indicate if the PLC precedes or follows the BD development.
356 as a spatial positional cue guiding bile duct growth and branching but also as a regulatory node involved in coordinating bile drainage from the hepatic lobule into the biliary network.
To avoid potential misunderstanding, we have further refined and revised the statements in the manuscript regarding the functional interpretation of the periportal lamellar complex (PLC) and its relationship to bile duct development. We agree that cell ablation strategies are of great importance for functional validation studies. However, it should be noted that CD34 and Sca-1 are relatively broadly expressed markers during liver development, labeling multiple endothelial, mesenchymal, and progenitor cell populations, and their expression is not restricted to the PLC. Owing to this broad expression pattern, ablation of CD34+Sca-1+ cell populations would likely exert widespread effects on vascular and stromal structures, thereby complicating the distinction between direct PLC-specific effects and secondary developmental alterations. As such, this strategy may present technical limitations for specifically dissecting the role of the PLC in bile duct development. At the same time, given that the primary objective of this study is the systematic characterization of the three-dimensional anatomical features and spatial organization of the PLC, we have correspondingly revised the manuscript to restrict statements regarding the relationship between the PLC and bile ducts to spatial associations supported by the current data. Specifically, our results show that primary bile ducts run along the main portal vein trunk, secondary bile ducts exhibit directed branching toward the PLC region, and terminal bile duct branches tend to spatially cluster in the vicinity of the PLC, thereby forming a reproducible periportal spatial arrangement. Based on these observations, the PLC delineates a relatively conserved anatomical microenvironment within the portal region, whose spatial position is closely associated with the organization and terminal distribution of the intrahepatic bile duct network.
We believe that these revisions more accurately reflect the experimental evidence and the defined scope of the present study.
(3) The following statement ought to be rephrased or skipped, considering that CD34 and Sca1 (Ly6a) are markers of periportal endothelial cells (Pietilä et al., 2025, Gómez-Salinero et al., 2022) and as shown by the authors in their own Fig. 6D. In this context and the context of the CCL4 experiments, a "simple" proliferative progenitor portal vein endothelial cell phenotype, suggested also by the presence of DLL4 (Fig5A) and JAG1 (Pietilä et al., 2025) (Benedito et al., 2009) ought to be considered.
409 Notably, CD34 and Sca-1 (Ly6a) were co-expressed exclusively within PLC structures surrounding the portal vein, but absent from central vein ECs and midzonal LSECs (Figure 4F).
We thank the reviewer for pointing out the potential imprecision in this wording. We agree that both CD34 and Sca-1 (Ly6a) are well-established markers of periportal endothelial cells, as previously reported (Pietilä et al., 2025; Gómez-Salinero et al., 2022), and as also illustrated in Fig. 4F of our study.
Accordingly, the original statement suggesting that CD34 and Sca-1 are co-expressed exclusively within the PLC structure may indeed represent an overinterpretation. Following the reviewer’s suggestion, we have revised the relevant text on page 409 by removing the exclusive phrasing (“only in”) and by emphasizing instead that CD34+Sca-1+ endothelial cells are enriched in periportal regions associated with the PLC, rather than being specific to or confined within the PLC.
In addition, in the context of the CCl4-induced liver fibrosis model, we agree with the reviewer that the observed expression of DLL4 and JAG1 under fibrotic conditions is more appropriately interpreted as reflecting an activated or proliferative periportal endothelial progenitor–like phenotype, rather than defining a novel endothelial lineage. The corresponding statements in the revised manuscript have been adjusted accordingly.
(4) Again, these concluding sentences are based on correlative evidence of mRNA expression and literature but not experimental evidence.
436 These findings suggest that this unique endothelial cell subset in the periportal region may possess dual regulatory functions in both metabolic and hematopoietic modulation
441 results suggest that PLC endothelial cells may not only regulate periportal microcirculatory blood flow but also help establish a specialized microenvironment that potentially supports periportal hematopoietic regulation, contributing to stem cell recruitment, vascular homeostasis, and tissue repair.
We thank the reviewer for this thoughtful comment. We agree that these statements are primarily based on transcriptomic correlation analyses and support from previous literature, rather than direct functional experimental evidence.
Accordingly, in the revised manuscript, we have appropriately toned down and adjusted the relevant concluding statements to more accurately reflect their inferential nature. The revised wording emphasizes associations and potential involvement, rather than definitive functional roles. These changes preserve the overall scientific interpretation while aligning the level of inference more closely with the available evidence.
The revised text now reads:
“Finally, we found that the main trunk of the PLC is primarily composed of CD34+Sca-1+CD31+ endothelial cells (Fig. 4J). These CD34+Sca-1+ double-positive cells are mainly distributed in the basal region of the PLC structure and exhibit molecular features associated with hematopoiesis. Taken together, these results suggest that PLC endothelial cells may contribute to the establishment of a local microenvironment related to periportal hematopoietic regulation and may play potential roles in stem cell recruitment and maintenance of vascular homeostasis.”
(5) The following part is speculative and based on re-analysis from the dataset that was gathered after 6 more weeks of CCL4 treatment (12weeks Su et al., 2021), then in the linked experiments from the manuscript. And should be moved to discussion or removed.
504 Moreover, single-cell transcriptomic re-analysis revealed significant upregulation of bile duct-related genes in the CD34+Sca-1+ endothelium of PLC in fibrotic liver, with notably high expression of Lgals1 (Galectin-1) and Hgf (Figure 5G). Previous studies have shown that Galectin-1 is absent in normal liver parenchyma but highly expressed in intrahepatic cholangiocarcinoma (ICC), correlating with tumor dedifferentiation and invasion (Bacigalupo, Manzi, Rabinovich, & Troncoso, 2013; Shimonishi et al., 2001). Additionally, hepatocyte growth factor (HGF), particularly in combination with epidermal growth factor (EGF) in 3D cultures, promotes hepatic progenitor cells to form bile duct-polarized cystic structures (N. Tanimizu, Miyajima, & Mostov, 2007). Together, these findings suggest the PLC endothelium may act as a key regulator of bile duct branching and fibrotic microenvironment remodeling in liver fibrosis.
Collectively, our results demonstrate that the PLC, situated between the portal vein and periportal sinusoidal endothelium, constitutes a critical vascular microenvironmental unit. It may not only colocalize with bile duct branches under normal physiological conditions, but also through its basal CD34+Sca-1+ double-positive endothelial cells, potentially orchestrate bile duct epithelial proliferation, branching morphogenesis, and bile acid transport homeostasis via multiple signaling pathways. Particularly during liver fibrosis progression, the PLC exhibits dynamic structural extension, serving as a spatial scaffold facilitating terminal bile duct migration and expansion into the hepatic parenchyma (Figure 5H). These findings highlight the PLC endothelial cell population and the vascular-bile duct interface as key regulatory hubs in bile duct regeneration, tissue repair, and pathological remodeling, providing novel cellular and molecular insights for understanding bile duct-related diseases such as ductular reaction, cholangiocarcinoma, and cholestatic disorders, and offering potential targets for therapeutic intervention.
We thank the reviewer for this careful and thought-provoking comment. We understand and agree with the reviewer’s assessment that this section involves a degree of inference, as the analysis is based on a re-analysis of a previously published single-cell transcriptomic dataset from a CCl4-induced liver fibrosis model (Su et al., 2021), rather than on experimental data directly generated in the present study.
In response to the reviewer’s suggestion, we have carefully re-examined and revised the relevant paragraphs. Without altering the overall structure of the manuscript, we have appropriately moderated the wording to clarify that these results primarily describe the transcriptional features of PLC-associated CD34+Sca-1+ endothelial cells under fibrotic conditions, and their associations with bile duct–related gene expression, rather than providing direct functional evidence for their roles in bile duct branching or microenvironmental remodeling.
In addition, we have explicitly clarified in the main text the data source and methodological limitations of the single-cell transcriptomic analysis, and emphasized that these findings should be interpreted in conjunction with the spatial information revealed by three-dimensional imaging. Through these revisions, we aim to retain the value of this analysis in providing complementary molecular insight into PLC characteristics, while avoiding potential over-interpretation of its functional implications.
Formal suggestions:
(6) The following sentence would benefit from being more clearly written.
263 - The formation of PLC structures in the adventitial layer may participate in local blood flow regulation, maintenance of microenvironmental homeostasis.
We thank the reviewer for this helpful suggestion. The sentence has been revised to improve clarity by correcting the parallel structure and refining the wording.
The formation of PLC structures in the adventitial layer may participate in local blood flow regulation and the maintenance of microenvironmental homeostasis.
(7) The following sentence is misleading as it implies cell sorting, and "subsetted" rather than "sorted" should be used.
414 Based on this, we sorted CD34+Sca-1+ endothelial populations from the total liver EC pool (Figure 4G).
Thank you for your comment.
We have revised the term as suggested. This avoids the misleading implication of physical sorting, as our operation was analytical subsetting of the target subpopulation.
We appreciate your careful review.
(8) Correct typos, especially in the results section related to Fig. 6. and formatting issues in the discussion.
730 Morphologically, the PLC shares features with previously described telocytes (TCs)- 731 a recently identified class of interstitial cells in the liver observed via transmission electron
We thank the reviewer for pointing out this textual error. In the submitted version, the sentence describing the morphological similarity between the PLC and previously reported telocytes was inadvertently interrupted due to a punctuation issue. This has now been corrected to ensure sentence integrity and consistent formatting.



